
Abstract
The question of whether any specific plant species are typically growing along railway tracks (the so-called “railway-wandering plants”) has been discussed for many years. This study proves the existence of a form of Geranium robertianum species growing along railway tracks in North–Eastern Poland. Floristic studies have been carried out in 246 areas along railway tracks. This particular species was found in 70 studied areas (28 %). Comparative studies were carried out on 12 plant populations growing in the fieldwork and in glasshouse cultivation. Plants growing along the railway tracks in Waliły were different from all other studied populations. They were small (smaller by 31 %, max. by 57 % than other plants), with little leaf blades representing different shapes and colour. In studies of light absorption by photosynthetic apparatus (chlorophyll fluorescence) under conditions of exposure to high light intensity, the plants from Waliły were proved to have a better adaptation capacity to stress conditions. Increased levels of anthocyanins—which provided better protection of the photosynthetic apparatus against insolation—were shown. The protective properties against water deficiencies and excessive insolation were genetically preserved and were found in the second generation of plants cultivated in a glasshouse. For the first time, a new plant form of G. robertianum—a “railway-wandering plant” adapted to the conditions prevailing along railway tracks—was confirmed to exist. The form has developed probably after 1886, when the Białystok–Zubki railway was built, featuring the Waliły railway station.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
For many years, botanists have taken interest in plants growing almost exclusively along railway tracks. Polish authors named such species “railway plants” (Kornaś et al. 1959; Zając and Zając 1969; Sendek 1973 and reference literature). It was finally concluded that such plants did not actually exist. Railway tracks are considered to be the habitat of native and invasive species, and can be compared to “an environmental corridor” enabling the colonization of new territories (Tikka et al. 2001). Geranium robertianum L. is mentioned among plant species commonly growing along railway tracks in various European countries, e.g. Germany or Austria (Brandes 1993, 2005; Lindacher 1995; Zarzycki et al. 2002; Tofts 2004).
In Poland, in the period between 2007 and 2008, comprehensive floristic studies were also carried out along railway tracks (Galera et al. 2011). The studies revealed common occurrence of G. robertianum species, covering the entire sections of railway tracks.
This prevalence of G. robertianum along tracks can be quite surprising, especially taking into consideration the habitat requirements of this particular species (Tofts 2004). This species prefers partly shaded, nitrogen and phosphorus rich alkaline soils (pH level above 5.5) (Grime et al. 1988). Meanwhile, completely different habitat conditions are typical along the railway tracks. Easily permeable soil consisting of crushed stones and river gravel is the reason for poor availability of water and nutritional deficits. Plants growing along railway tracks are typically exposed to strong sunlight (insolation). In addition, once every few months the plant cover is intentionally devastated by spraying of herbicides (Roundap 360). Moreover, toxic compounds such as heavy metals, PAHs, and PCBs are typically present in the railway soil (Malawska and Wiłkomirski 2001; Wiłkomirski et al. 2011).
Thus, the question arises why is G. robertianum so commonly growing along railway tracks despite such specific, seemingly adverse habitat conditions?
The aim of the studies was to determine prevalence and habitat preferences of G. robertianum plants along railway tracks in North–Eastern Poland. The next question was how to explain the prevalence of G. robertianum along railway tracks under such unfavourable habitat conditions. Both fieldwork observations and measurements of plants cultivated in a glasshouse were carried out, with particular attention devoted to the railway track population of G. robertianum in Waliły (NE Poland). Waliły is a railway station situated along the railway Białystok–Zubki Białostockie, one of the oldest railways in Poland, opened in 1886 (www.kolej.onet.pl).
Materials and methods
Prevalence of G. robertianum along railway tracks in North–Eastern Poland
Geranium robertianum occurrence along railway tracks representing considerably different habitat conditions was determined based on fieldwork observations carried out in 246 areas, 20 m2 each. The study was conducted along the railway tracks of North–Eastern Poland and included:
-
66 areas along operating tracks located at 11 sites.
-
180 areas along closed tracks located at 30 sites.
Locations of the sites and the arrangement of the areas within each of sites are presented in Wiłkomirski et al. (2011).
The occurrence of G. robertianum along the tracks was analysed in terms of:
-
railway track utilization (comparison of operating railway tracks and railway tracks closed at different time periods—from 1982 to 2000),
-
local conditions (indication whether railway tracks are surrounded by leafy forests, coniferous forests, meadows, or ruderal areas),
-
shading (% share of shaded area),
-
trampling (trampling-free, weak, relatively weak and high trampling intensity),
-
mechanical composition of the track ballast (2 basic types of soil: crushed stones with the prevalence of coarse fractions and fine gravel),
-
plant layer density (% of area covered by plants).
The habitat conditions of G. robertianum along railway tracks in North–Eastern Poland were evaluated by means of the principal component analysis (PCA). Calculations were carried out on the basis of covariance matrix, using Statistica ver. 9 (Statsoft Inc.).
Collection of study material in fieldwork
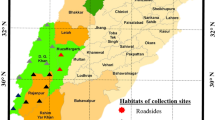
There were six localities of G. robertianum identified along railway tracks of North–Eastern Poland where G. robertianum heavily prevailed (Table 1). Another six localities were selected from among natural habitats of G. robertianum in different parts of Poland: forests near the watercourses, humid and shady bushes, insolated ravine screes (Fig. 1; Table 1). Observations and measurements were taken during three vegetative seasons (2007–2010) at 12 localities (Fig. 1; Table 1), where herbarium specimens of the plants as well as 10 largest leaves from 20 plants from each of the sites were collected. The leaf surfaces were measured and the colour and morphological structure of fruits were compared. Seeds from all 12 localities were collected for experimental purpose to be cultivated in a glasshouse. At each site, seeds were collected from more than 50 plants.

Map of Poland with locality numbers of Geranium robertianum (no. 1–12)—locality description presented in Table 1
Soil studies
Soil specimens were collected from six localities along railway tracks according to the previously reported methodology (Wiłkomirski et al. 2011; Galera et al. 2011). Soil samples were collected at each locality, three samples were collected between the tracks and three were collected at the outer sides of railway tracks (at roughly 70 cm distance from the tracks). In total, six soil samples were collected at each locality at the depth of 0–20 cm. Samples were subsequently mixed together and analysed in six duplicates. The soil samples were sieved with sieves of 1 and 0.25 mm mesh size. Comparative soil samples were collected in the area surrounding the railway tracks—at a distance of approximately 20–100 m (reference site). Soil was analysed with the use of standard methods. Acidity was tested with a pH meter. For this purpose, 10 g of soil was mixed with 25 cm3 of water or 1M KCl. The measurements were taken after 24 h (standard procedure). Nitrogen content in the soil was analysed using the Kiejdahl’s method. A weighed amount of 5 g of soil was mineralized and analysed in a Kiejdahl’s apparatus. Concentration of phosphorus bioavailability was tested using the Egnar’s method in Rhiem’s modification (spectrophotometry). Exchangeable cations concentration (Ca, K, Mg, Na) was tested after the extraction of 5 g of soil with 0.1 M ammonium acetate solution (24 h) with an ASA method (Galera et al. 2011).
Glasshouse cultivation
Plants grown from seeds collected at 12 localities (Table 1) were cultivated in a glasshouse, in horticultural soil during the period of 6 months (January–June). In periods of low light availability, irradiation of plants was applied (light intensity: 120 μmol m2 s−1, photoperiod: 16:8, temperature: 22 ± 3 °C). The experiment was repeated three times, each time the plant cultivation was carried out from seeds collected on field. Preliminary studies have shown that the seeds collected in the field were infected, which hampered their germination. For this reason, they require sterilization.
Prior to planting, the seeds (100–120 from each population) were sterilized in 70 % ethanol for 1 min, rinsed with water, and in Clorox (50 %) for 4 min and 10× rinsed with water. The seeds were put in water for 24 h (imbibition period) and were seeded into Petri dishes covered with a filter paper (30 seeds in each dish). After sprouting (approx. 2 weeks), seedlings were transferred to flower pots with horticultural substrates. After 5 months of cultivation, the plants reached the peak vegetative phase. After next 3 months, the plants entered the rest phase. None of the cultivated plans burst into blossom in the first year of cultivation. Only some plants reached the generative phase in the next year.
Approximately 30 (20–40) plants of each population were cultivated in the afore-mentioned method. During three consecutive years, about 500 plants were cultivated in total.
Plant morphology
After 5 months of cultivation, the following measurements of each plant were carried out: number of leaves, surface area of the largest leaves (10 leaves) and the length of petioles. The leaf surface was measured with Winfolia software (WinFolia 2007b For Leaf Analysis, Regent Instruments INC). Manually transverse sections of leaves and petioles were analysed with the use of light and stereoscopic microscopes. Thirty plants per population were examined in this manner.
Seed sprouting
Seed viability was analysed with a standard TTC method (tetrazolium test). Mericarps swollen in water were incubated in 1 % solution of 2,3,5-triphenyltetrazoline (TTC) for 48 h at 30 °C. Seed sections (in mericarps) were analysed under a stereoscopic microscope. The red colour of embryo tissues confirmed high seed viability, whereas discoloured embryos were found dead.
Seed germination was tested with standard methods (Wierzbicka and Obidzińska 1998). For this purpose, sterilized seeds were placed on Petri dishes, on wet filter paper; 150 (3 × 50) seeds per population. The number of germinated seeds was determined for a period of 2 weeks.
The amount of anthocyanins
Measuring the amount of anthocyanins in leaves was carried out in vivo in two selected populations of plants—Waliły (railway track population) and Zajezierce (natural population).
These populations characterized with major morphological differences, including differences in the colour of the leaves. Plants were grown for 3 months in the greenhouse—10 plants in each population. Determining the amount of anthocyanins was performed using the modified method of Fuleki and Francis (1968). 0.5 g of middle-aged leaves of each plant was homogenized in 4 ml of methanol acidified with HCl (1 %) and left for 4 h at 4 °C. Samples were centrifuged for 30 min and the absorbance was measured at a wavelength of λ = 530 nm and λ = 657 nm.
Chlorophyll a fluorescence in vivo
Plant physiological state was analysed on the basis of fluorescence parameters determination (Romanowska 1999; Kalaji and Łoboda 2009). Chlorophyll a fluorescence was measured in vivo in the selected two plant populations from Waliły (railway population) and Zajezierce (natural population). Plants of both populations (five plants in each) were cultivated for 6 months in a glasshouse at a low level of light intensity (120 μmol m2 s−1, temperature 22 °C). Fluorescence was measured in five leaves per plant (middle-aged leaves were selected) with a fluorometer—Fluorescence Monitoring System (Hansatech). The leaves were adapted to lack of light for 15 min. Chlorophyll a fluorescence was tested under the following conditions:
-
1.
Modulated measurement light: roughly 0.05 μmol m2 s−1 (excitation-free).
-
2.
Saturation light: 4,500 μmol m2 s−1 impulse duration: 1 s.
-
3.
Actinic light intensity: 60–1,020 μmol m2 s−1 (each PPFD 8 min) : 60, 120, 180, 240, 420, 600, 820, 1,020 μmol m2 s−1.
Chlorophyll fluorescence capacity coefficients: Fv/Fm, qp (photochemical quenching of fluorescence) and NPQ (non-photochemical quenching of fluorescence) were measured according to Gently et al. (1989).
After all measurements were completed, the same plants were irradiated under high light intensity (1,200 μmol m2 s−1, temperature 28 °C) for 7 days. Chlorophyll a fluorescence was measured on the leaves of all plants according to the same procedure. In total, 50 leaves were analysed.
Statistical analysis
The significance of measurement results was tested by a univariate analysis of variance (ANOVA), with a post hoc Tukey’s test (for p = 0.05). XL Stat software by Addinsoft was used for calculations.
Results
Occurrence of G. robertianum along railway tracks in North–Eastern Poland
During floristic observations carried out in 246 research areas, G. robertianum was found in 70 areas, comprising 28 % of all studied areas. This species was present at sites representing varied characteristics:
-
railway track utilization (intensively and extensively operating railway tracks and railway tracks closed in different time periods—from 1993 to 2000),
-
local conditions (whether railway tracks are surrounded by leafy forests, coniferous forests, meadows, or ruderal areas),
-
shading (0–80 % share of shaded area),
-
trampling (G. robertianum occurred along trampling-free tracks and tracks exposed to high trampling intensity),
-
track ballast (crushed stones with the prevalence of coarse fractions and fine gravel),
-
plant layer density (10–90 % share of area covered by plants).
Based on the results of PCA analysis, no correlations were shown between the prevalence of G. robertianum and all of the analysed characteristics. The populations of G. robertianum prevailing at the studied areas were highly diversified in terms of the environmental conditions along railway tracks.
Biometric characteristics of G. robertianum—fieldwork
Geranium robertianum plants field studied differed by size, habit and leaf colour, depending on specific habitat (Figs. 2, 3). As a rule, the plants at insolated positions were small with red leaves (Fig. 4). Along railway tracks, specimens of G. robertianum prevailed in clusters, and the plant habit was different (Fig. 2) from that of plants at natural localities (Fig. 3).

Geranium robertianum, a railway track at the Waliły railway station

Geranium robertianum, a natural locality, screes, Pieniny National Park

Leaf surface of Geranium robertianum plants collected at: natural localities SB Stara Białowieża, ZAJ Zajezierce; railway tracks WAL Waliły railway station, HAJ Hajnówka—closed track, IŁA Iława railway siding, CIS Cisówka dyke, HZ Hajnówka switch, BF Białystok Fabryczny. Asterisk—statistically significant difference
Leaf size
To characterize the field plants, the largest leaf surface was measured (Fig. 4). The leaves of plants growing along railway tracks localities were typically smaller than those of plants growing at natural localities (10.1 and 16.0 cm2 on average, respectively).
Significant differences were also found among the plants growing along railway tracks (Fig. 4). The smallest leaves were developed in plants from Hajnówka and Waliły localities (3.9 and 5.3 cm2, respectively) and the largest—in plants from Białystok Fabryczny (18.2 cm2). The leaf area could differ by as much as fivefold.
Mericarps
Mericarps were found to be of the same size at all studied localities and had the corrugated surface (Fig. 5a–f). Differences were shown only in the colour of mericarps. Mericarps in Waliły were red–brown (Fig. 5a–c), whereas the mericarps of all other plants were dark brown (Fig. 5d–f).

a–c Red mericarps of Geranium robertianum—collected from plants on railway tracks at the Waliły railway station. 4× magnification. d–f Brown mericarps of G. robertianum—collected from plants in natural localities in Zajezierce. 4× magnification
Chemical composition of the railway track ballast (soil)
As G. robertianum shows high demand with regard to pH and soil fertility, chemical soil composition at the railway tracks localities was tested. For the comparison, chemical composition of soil from the vicinity of railway tracks—reference site—was also analysed (Table 2).
It was revealed that the pH level of the track soil was balanced and alkaline in all studied railway track sections within the range of 7.37–7.80 (7.65 on average). Similar pH results were obtained in measurements with KCl. In this case, pH of the soil was within the range of 6.98–7.57 (7.32 on average). pH levels of the surrounding soil were lower: 7.43 and 6.89 (with KCl) on average (Table 2).
The concentration of all studied chemical elements in railway track soil was similar or higher compared with the surrounding soil (reference site). The level of nitrogen in track soil was relatively high, 0.312 % on average, as compared to 0.284 % in the surrounding soil (reference site). Along railway tracks, the total level of phosphorus was 30.5 mg/100 g on average, and 16.4 mg/100 g in the surrounding soil. The average concentration of sodium was 2.5 mg/100 g and 1.3 mg/100 g along railway tracks in the surrounding soil (reference site). Similarly, the concentration of potassium was 11.3 mg/100 g and 6.4 mg/100 g, respectively, the concentration of magnesium—13.2 mg/100 g and 9.3 mg/100 g, respectively, the concentration of calcium—411 mg/100 g and 249 mg/100 g, respectively (Table 2).
Obtained results indicate that in all studied localities, pH level of the soil was slightly alkaline and nutritional conditions were most often better than in the surrounding soil.
Biometric characteristics of the plants cultivated in a glasshouse
Further studies were carried out to examine if the differences between plants from different localities can be attributed only to different habitat conditions, or if they were genetically preserved. For this purpose, the first generation of plants from 12 localities (further called populations) was cultivated in parallel, under similar glasshouse conditions. Plants from Waliły stood out among all the studied populations. They were the smallest and characterized with red leaves.
30–50 % of seeds from different populations germinated during 14 days. The plants were analysed with a tetrazolium method; some of the germs were dead and, therefore, unable to sprout.
Leaf blade shape and colour
The leaves of G. robertianum plants have palmatisect leaf blade (Fig. 6). However, the shape of plants from the Waliły railway tracks differed significantly from the other studied populations: the leaves were small, clearly cut-out, of narrow stipules, whereas the leaves of plants of the remaining populations (growing along railway tracks and in natural populations) were larger, with wide stipules (Fig. 6).

Comparison of habit and colour of two leaves of Geranium robertianum plants after 5 months of glasshouse cultivation with moderate light intensity. On the left—a plant leaf collected in Waliły (railway tracks), on the right—a plant leaf from Zajezierce (natural population)
Another characteristic typical for the plants from Waliły railway track population as compared to all other plants was the colour of the leaf blade. On the dorsal side (upper), all the leaves of plants from the Waliły population were dark green with purple colour, whereas the leaves of all remaining plants were green or light green. However, underneath, all leaves of plants growing along track populations, including the leaves of Waliły plants, were purple. The leaves of natural plant populations were most often green underneath (Fig. 6).
The red colour of the underside of Waliły population leaves resulted from red dyeing of epidermal or parenchyma cells of leaf underside (anthocyanins), which was confirmed by transverse sections of the leaves observed under light microscope.
The comparative culture of different plant populations of G. robertianum was repeated three times, for three consecutive years. Each time, the plants from the Waliły railway tracks were distinguishably small, represented a different shape and leaf colour and a quite uniform morphology of all individuals in the population.
The amount of anthocyanins
Using the method of Fuleki and Francis (1968), the amount of anthocyanins was examined in leaves of both populations of G. robertianum. It was found that the average concentration of anthocyanins in leaves from population from Waliły was 379 (±18.7) μg/g fresh weight, while in the population from Zajezierce only 82 (±9) μg/g fresh weight.
These results demonstrate that the difference in colour between the leaves from two populations was caused by more than fourfold higher levels of anthocyanins in the leaves of plants from Waliły population compared to the population of Zajeciere.
Assimilation surface of the plants
Among all populations of plants cultivated in glasshouse and examined after reaching the complete vegetative development level, a small size was observed in the Waliły railway track population. A single leaf surface of this particular population was 25.14 cm2 on average, while the average leaf surface of all other populations was 38.82 cm2. The average difference of 35.3 % (max. 45.2 %) in the leaf blade surface between the Waliły population and of all other populations was statistically significant. The biggest leaves (an average surface of 45.9 cm2) were found in plants from the Hajnówka railway population. However, no statistically significant differences were found between the remaining populations of the plants.
The number of leaves per plant in particular populations was 33 on average (from 28 to 38) and no significant differences were found between the studied populations. Based on the average surface of the leaf blade surface and the number of leaves in each population, the assimilation surface of the plants has been calculated (Fig. 7). Of all studied populations, the Waliły railway track population again distinguished itself with the smallest assimilation surface per plant—867 cm2—on average (a statistically significant difference compared to other populations). The assimilation surface of the remaining plant populations (both from the railroad and natural) was from 1,035 cm2 on average (the Pieniny National Park) up to 1,526 cm2 (Zajezierce population).

The surface of a single Geranium robertianum plant collected at a natural locality and at railway tracks—after 5 months of glasshouse cultivation: PPN Pieniny National Park, LŁ Forestry Łuknajno, POW Powsin, ZAJ Zajezierze, WAL Waliły railway station, HAJ Hajnówka closed track, IŁA Iława railway siding, CIS Cisówka dyke, HZ Hajnówka switch. Asterisk statistically significant difference
The assimilation surface of all leaves per plant from the Waliły population was smaller by 31 % on average than in all other populations (max. difference was 57 %—with the Zajezierce population).
The average length of petiole was 18.5 cm and was similar in all studied plants (from 15.0 cm to 21.4 cm, and in the Waliły population—16.7 cm on average).
The plants cultivated in a glasshouse and obtained from seeds collected at railroad localities in Waliły differed significantly from all other plants. The Waliły population plants were distinguished by a smaller size and the smallest assimilation surface of plants. The characteristic feature of this population was a very small intra-population variability of the plants. All cultivated plants were almost identical in terms of morphological traits.
The comparison of physiological conditions of populations from railways and natural habitats based on chlorophyll a fluorescence parameters
Physiological condition of plants originating from two most different populations—from Waliły (a railway population) and Zajezierce (natural population)—was analysed. The examined plants were cultivated for 6 months under low light intensity (120 μM m2 s−1, temperature 22 °C), and then exposed to high light intensity for 7 days (1,200 μM m2 s−1, temperature 28 °C). Chlorophyll a fluorescence parameters were subsequently measured and analysed (Table 3, Figs. 8, 9, 10, 11).

Photochemical quenching of fluorescence (qp) in the leaves of Geranium robertianum plants after 6 months of cultivation under moderate light intensity (120 μmol m2 s−1)

Non-photochemical quenching of fluorescence (NPQ) in the leaves of Geranium robertianum plants after 6 months of cultivation under moderate light intensity (120 μmol m2 s−1)

Photochemical quenching of fluorescence (qp) in the leaves of Geranium robertianum plants after 6 months of cultivation under moderate light intensity (120 μmol m2 s−1) subsequently exposed to very high light intensity for 7 days (1,200 μmol m2 s−1)

Non-photochemical quenching of fluorescence (qp) in the leaves of Geranium robertianum plants after 6 months of cultivation under moderate light intensity (120 μmol m2 s−1) subsequently exposed to very high light intensity for 7 days (1,200 μmol m2 s−1)
Low light intensity
There were no differences found between Waliły populations (growing along railway tracks) and Zajezierce population (natural population) grown under low light intensity (Table 3). The average level of minimal fluorescence (F0) was similar in both populations: 328 (±45) (Waliły) and 377 (±23) (Zajezierce). The Fv/Fm ratio reflects maximum quantum efficiency of photosystem II (PSII) of leaves adapted to the dark. The coefficient was similar in both populations with the value of 0.826 in Waliły population and 0.821 in Zajezierce population. The above coefficient values reflect high photochemical yield of photosystem II (Table 3).
Figure 8 shows photochemical quenching of fluorescence (qp) as a coefficient of the amount of energy consumed in photochemical reactions. The level of photochemical quenching was similar in both populations. NPQ coefficient (Fig. 9) indicating a non-photochemical quenching of energy was similar in both studied plant populations.
Energy consumption in photochemical reactions was identified on the basis of in vivo measurements of chlorophyll a fluorescence parameters Fv/Fm, qp, and NPQ. The photosynthetic apparatus of all studied plants (growing along railway track and in natural populations) cultivated in low light intensity (120 μmol m2 s−1) were shown to have ability to efficiently use the light. This is a measure of the good physiological condition of plants from both studied populations and of absence of differences between both populations.
High light intensity
To investigate the reactions of both plant populations (Waliły railway track population, and Zajezierce natural population) to light stress, the plants were exposed to high light intensity for the period of 7 days (1,200 μmol m2 s−1).
Fluorescence of chlorophyll a parameters was different in both plant populations (Table 3; Figs. 10, 11). Under conditions of light stress, the plants representing the Waliły population maintained good fluorescence parameters compared to those under conditions of low light intensity (Table 3). Basic fluorescence (F0) was 409 (±11) on average. The Fv/Fm coefficient (PSII yield) was 0.731, which indicates a slight reduction of photochemical efficiency of photosystem II (PSII).
However, these parameters differed diametrically in plants from Zajezierce (natural population). Minimal fluorescence (F0) was 1,045(±74). (Fm) was 1,433(±303), and the Fv/Fm coefficient was only 0.257(±0.10) (Table 3). These are very strong indicators of damage of PSII. This indicates a very weak photochemical efficiency of photosystem II in natural plant population (Zajezierce). Photochemical quenching of fluorescence (qp) shown in Fig. 10 indicates decreased efficacy of the photochemical reactions in plants from Zajezierce (natural population) compared to plants from Waliły (population growing along railway tracks). Also, NPQ coefficient was significantly lower in natural plant population (Zajezierce), which reflects a very weak yield of non-photochemical quenching in these plants (Fig. 11).
The above results indicate significant differences between response to light stress in the two populations of G. robertianum.
The experiments were carried out on the second generation of plants cultivated for 6 months under the same conditions in glasshouse. The Waliły plants revealed (inherited) adaptations to stressful environmental conditions and were shown to be better adapted to excessive light, characteristic of such localities, as compared to the natural plant population from Zajezierce.
Discussion
As a result of this study, G. robertianum was shown to quite frequently grow along railway tracks in Poland (28 %). Interestingly, G. robertianum was found to grow in different habitat conditions, which may indicate surprisingly high ecological amplitude of the species and/or a significant differentiation of the studied populations in terms of habitat requirements.
The question arises why G. robertianum is able to grow along operating railway tracks in clearly unfavourable conditions (see the “Introduction”). The answer to this question was investigated in this study.
Adaptation or acclimations
It is commonly known that plants are capable to adjust to stress conditions by acclimation (non-hereditary changes resulting from phenotype plasticity) or adaptation (hereditary changes).
The habit of G. robertianum plants growing along operating or recently closed railway tracks in North–Eastern Poland differed from the habit of plants growing under natural conditions. Plants growing along railway tracks were most often of small size, with leaves even five times smaller than those of plants from natural habitats. Since conditions along railway tracks are generally unfavourable for plant growth, the plants are of smaller size as a result of the acclimation process so that they are able to maintain water balance. This assumption was confirmed in some cases—the leaves of plants from Hajnówka growing along a closed track were the smallest among all studied plants (3.9 cm2 on average). The leaves of the next generation of plants cultivated in favourable glasshouse conditions were as large as 45.9 cm2. They were the largest of all studied leaves. In this case, acclimation process took place.
On the other hand, G. robertianum plants from Waliły represented a much more complex case. The leaves of Waliły plants were small as well (5.3 cm2). However, the leaves of the second generation of plants cultivation in favourable glasshouse conditions were still the smallest among all plants cultivated in glasshouse. The average surface of the leaves was 25.14 cm2 and was by 35.3 % (max. 45.2 %) smaller than in case of other plants.
Therefore, the smaller size of G. robertianum plants from Waliły railway track can be explained by two reasons: unfavourable conditions along the tracks (acclimation process) and a genetically preserved characteristic (adaptation processes). This was confirmed in G. robertianum plants cultivated under favourable glasshouse conditions, which remained the smallest in the three consecutive years of cultivation.
The correlations between differences of plant size and leaf shape and habitat conditions of many plant species were investigated in numerous studies (Briggs and Walters 1997), including Geranium sanguineum (Lewis 1969). However, no such observations were carried out in anthropogenic habitats along railway tracks.
Soil pH level
In natural conditions, G. robertianum grows in neutral or alkaline soil. It appeared obvious that railway tracks leading across a variety of localities will also represent different pH levels. However, it was shown that soil pH levels were similar and equalled 7.65 on average. The alkaline soil was favourable for G. robertianum growth. The alkalinity of soil under railway tracks can be possibly explained by the alkalinity of the railway track ballast (crushed stones)—the results obtained for numerous railway track sections were very similar (Galera et al. 2011).
Soil fertility
Soil fertility is perhaps another factor which could limit the occurrence of G. robertianum along railway tracks. This species requires a fertile soil and usually occurs in nitrogen and phosphorus-rich soil.
The concentration of nitrogen, phosphorus and other elements (Ca, K, Mg, Na) in the soil under railway tracks was examined in this study. Note that the soil was mixed with the crushed stones of railway embankment. The soil was particularly scarce under operating railway tracks. However, the soil was rich in nitrogen in all studied localities. Nitrogen concentration (approx. 0.3 %) was as high as in other fertile soils in Poland (brown and black soils) (Lityński 1982). As compared to the surrounding soils, the concentration of nutritional elements in the railway soil was comparable or higher, which reflects favourable nutritional conditions for plants. High concentration of nitrogen and other elements in the railway track soil can be explained by passenger traffic, with faecal matter removed from railway coaches directly to the railway tracks.
Thus, soil fertility contributes to the growth of G. robertianum along railway tracks instead of limiting it.
Water deficit
In natural localities, G. robertianum prevails in shady and semi-shady forests where water is abundant. Therefore, water deficit, which probably occurs along railway tracks, would appear to prevent the species from growing along railway tracks.
In the present study, plant leaves growing along railway tracks were smaller by approximately 40 % than those from the natural habitats. This was probably caused by hard habitat conditions related to water deficit along the tracks. Small leaves reduce assimilation area hence it is easier to keep the water balance (acclimation processes).
However, water deficit along the tracks was an equally powerful selective factor. In one of the studied populations, G. robertianum plants were still distinguished by small leaf assimilation surface. Under beneficial water and soil conditions (glasshouse cultivation), the leaves of plants growing along the Waliły railway tracks were smaller, with limited assimilation surface as compared to all other plants (difference by 31 %, max. 57 %).
Thus, through selection a new form of G. robertianum developed in Waliły, which is better adapted to maintain economical water balance. This characteristic was shown to be hereditary.
Excessive exposure to light
In natural localities, G. robertianum typically grow under conditions of partial shading, and the excessive insolation in exposed sections of railway tracks should limit the propagation of this species along railway tracks. Excessive light intensity may inhibit photosynthesis. Photosynthesis efficiency is correlated with chlorophyll a fluorescence (Romanowska 1999; Kalaji and Łoboda 2009). Fluorescence parameters reflect the physiological condition of plants. Chlorophyll a fluorescence reveals stress conditions and stress tolerance of the plants (Romanowska et al. 2006; Drożak and Romanowska 2006).
During the course of this study, chlorophyll a fluorescence parameters were analysed for G. robertianum cultivated under conditions of low light intensity—120 μmol m2 s−1, i.e. under intensity corresponding to conditions prevailing in partly shaded forest. A very good light utilization by photosynthetic apparatus was observed in all studied plants.
However, after exposure to stress, i.e. high light intensity—1,200 μmol m2 s−1, differences were observed in the response of different populations. A significant damage of photosynthetic apparatus was shown in G. robertianum plants from natural populations (Zajezierce). On the other hand, the photosynthetic apparatus of G. robertianum plants from the railway track population (Waliły) still maintained its high performance. Thus, track population of plants from Waliły has got genetically preserved characteristic (adaptation), which finally protects photosynthetic apparatus from light intensity (insolation).
To adapt to high light intensity, plants produce high levels of anthocyanins in leaves (Romanowska 1999). Anthocyanins are secondary metabolites (flavonoids). They are collected in vacuoles of the plant cells and colour the leaves red. They can absorb light and are believed to protect the plants from UV radiation (Grime et al. 2004).
The leaves of G. robertianum plants typically become red under conditions of excessive light. In on-site studies, G. robertianum plants of intensive red colour were observed—both along railway tracks (Fig. 2) and under natural conditions, e.g. at screes in the Pieniny National Park (Fig. 3). This characteristic feature was also described by other authors (e.g. Tofts 2004). However, in the present studies, the leaves of Waliły plant population were unusually red. Thus, the Waliły railway track population having four times more anthocyanins in the leaves was additionally protected from excessive light as compared to the natural plant population (Zajezierce).
Hence, excessive insolation along railway tracks can influence G. robertianum growth along railway tracks. Our research shows that a new plant form (the lowest taxonomic rank used for plants) has developed along the railway tracks, having genetically preserved increased anthocyanins levels in the leaves to protect the photosynthetic apparatus against excessive light. This particular feature is hereditary and has been developed in an adaptation processes.
This paper discusses the question of how is it possible for G. robertianum to successfully grow in unfavourable conditions along the railway tracks.
In this study, it was shown that a number of conditions typically occurring along the tracks are in fact favourable for the growth of G. robertianum, in particular: soil pH level and its fertility. However, two factors probably inhibit the growth of G. robertianum: water deficit and excessive light intensity (insolation).
These two factors have apparently led to micro-evolution processes and a new form of G. robertianum plant has developed in Waliły—“a railway-wandering form”, well-adapted to growth conditions observed along the railway tracks and distinguished by:
-
Small size and the smallest assimilation surface (by 31 %).
-
Different leaf habit—small stipules.
-
Leaf colour—more purple (4 times higher level of anthocyanins)
-
Seed split colour—red.
The question of whether any specific plant species are typically growing along railway tracks (“railway-wandering plants”) and not occurring anywhere else has been discussed in the studies for many years (Zając and Zając 1969; Brandes 2005; Büscher et al. 2008). It appears, however, that such species are extremely difficult to identify. The majority of species growing along railway tracks subsequently populate ruderal habitats of different types.
The present study reveals for the first time the development of a typical “railway-wandering plant” formed in a micro-evolution process. A new plant form of G. robertianum was identified along the railway track in Waliły, which was perfectly adapted to railway conditions. The railway line in Waliły has existed for 130 years, which was sufficient for the new plant to adapt to the anthropogenic habitat. The “form” is the lowest taxonomic rank used for plants. The formation of new plant forms in anthropogenic habitats has been also discussed with reference to areas heavily polluted with heavy metals, such as zinc-lead dumps (e.g. Przedpełska and Wierzbicka 2007; Olko et al. 2008; Wierzbicka and Panufnik 1998; Załęcka and Wierzbicka 2002; Wierzbicka and Pielichowska 2003), which has been recently confirmed in genetic studies (Abratowska et al. 2012).
References
Abratowska A, Wąsowicz P, Bednarek PT, Telka J, Wierzbicka M (2012) Morphological and genetic distinctiveness of the metallicolous and non-metallicolous populations of Armeria maritima s.l. (Plumbaginaceae) in Poland. Plant Biology 14:585–595
Brandes D (1993) Eisenbahnanlage Untersuchungsstand der Geobotanik. Tuexenia 13:415–444 (in German)
Brandes D (2005) Kormophytendiversität innerstädtischer Einsenbahnaulagen. Tuexenie 25:269–284 (in German)
Briggs D, Walters SM (1997) Plant variation and evolution. Cambridge University Press, Cambridge
Büscher D, Keil P, Loos GH (2008) Neue Ausbreitungstendenzen von primär als Eisenbahnwanderer aufgetretenen Pflanzenarten im Ruhrgebiet: Die Beispiele Evagrostis minor, Geranium purpureum und Saxifraga tridactylis. Brauschweiger Geobotanische Arbeiten 9:97–106 (in German)
Drożak A, Romanowska E (2006) Acclimation of mesophyll and bundle sheath chloroplasts of maize to different irradiances during growth. Biochim Biophys Acta 1757:1539–1546
Fuleki T, Francis FJ (1968) Quantitative methods for anthocyanins. 1. Extraction and determination of total anthocyanins in cranberries. J Food Sci 33:72
Galera H, Sudnik-Wójcikowska B, Wierzbicka M, Wiłkomirski B (2011) Encroachment of forest species into operating and abandoned railway areas In North-eastern Poland. Plant Biosystems 145:23–36
Gently B, Briantais JM, Baker NR (1989) The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophs Acta 990:87–92
Grime JP, Hodgson JG, Hunt R (1988) Comparative Plant Ecology, Allen & Unwin, London
Grime JP, Hodgson JG, Hunt R, Groud KS (2004) Nature’s swiss army knife; the diverse protective roles of antocyanins in leaves. J Biomed Biotechnol 5:314–320
Kalaji MH. Łoboda T (2009) Fluorescencja chlorofilu w badaniach stanu fizjologicznego roślin, SGGW Publishing house, Warsaw (in Polish)
Kornaś J, Leśniowska I, Skrywanek A (1959) Obserwacja nad florą linii kolejowych i dworców towarowych w Krakowie. Fragm Flor Geobot 5(2):199–216 (in Polish)
Lewis CM (1969) Genecological differentiation of leaf morphology in Geranium sanguineum L. New Phytol 68:481–503
Lindacher R (1995) Phanart. Datenbank der Gefässpflanzen Mitteleuropas. Erklärung der Kennzahlen, Aufbau und Inhalt. Veröffentlichung des Geobotanischen Institutes Tech. Hochschule, Stiftung Rübel, Zurich 125:1–436 (in German)
Lityński T (1982) Żyzność gleby i odżywianie roślin, Państwowe Wydawnictwo Naukowe, Warszawa (in Polish)
Malawska M, Wiłkomirski B (2001) An analysis of soil and plant (Taraxacum officinale) contamination with heavy metals and polycyclic aromatic hydrocarbons (PAHs) in the area of the railway junction Iława Główna, Poland. Air Soil Pollut 127:339–349
Olko A, Abratowska A, Żyłkowska J, Wierzbicka M, Tukiendorf A (2008) Armeria maritima from a calamine heap—Initial studies on physiologic—metabolic adaptations to metal en riched soil. Ecotoxicol Environ Saf 69:209–218
Przedpełska E, Wierzbicka M (2007) Arabidopsis arenosa (Brassicaceae) from a lead-zinc waste heap in southern Poland—a plant with high tolerance to heavy metals. Plant Soil 299:43–53
Romanowska E (1999) Adaptacje błon tylakoidowych do zmiennych warunków świetlnych środowiska. Postępy Biochemii 45:1–14 (in Polish)
Romanowska E, Wróblewska B, Drożak A, Siedlecka M (2006) High Light intensity protects photosynthetic apparatus of pea plants against exposure to lead. Plant Physiol Biochem 44:387–394
Sendek A (1973) Flora synantropijna terenów kolejowych węzła kluczborsko-oleskiego. Rocz. Muz. Górnośl. Bytom, Przyr 6:3–174 (in Polish)
Tikka PM, Högmander H, Koski PS (2001) Road and railway verges serve as dispersal corridors for grassland plants. Landscape Ecol 16(7):659–666
Tofts RJ (2004) Geranium robertianum L. Biological flora on the British isles. J Ecol 92:537–555
Wierzbicka M, Obidzińska J (1998) The effect of lead on seed imbibition and germination in different plant species. Plant Sci 137:155–171
Wierzbicka M, Panufnik D (1998) The adaptation of Silene vulgaris to growth on a calamine waste heap (S. Poland). Environ Pollut 99:1–12
Wierzbicka M, Pielichowska M (2003) Adaptation of Biscutella laevigata L, a metal hyperaccumulator, to growth on a zinc-lead waste heap in southern Poland. I: Differences between waste-heap and mountain populations. Chemosphere 54:1663–1674
Wiłkomirski B, Sudnik-Wójcikowska B, Galera H, Wierzbicka M, Malawska M (2011) Railway transportation as a serious of organic and inorganic pollution. Water Air Soil Pollut 218:333–345
Zając EU, Zając A (1969) Flora synantropijna linii kolejowej Czechowice—Zebrzydowice. Fragm Flor Geobot 15:271–282 (in Polish)
Załęcka R, Wierzbicka M (2002) The adaptation of Dianthus carthusianorum L. (Caryophyllaceae) to growth on a zinc-lead heap in southern Poland. Plant Soil 246:249–257
Zarzycki K, Trzcińska-Tacik H, Różański W (2002) Ecological indicato values of vascular plants of Poland. W. Szafer Institute of Botany, Krakow
Acknowledgments
We wish to acknowledge our indebtedness to the Ministry of Science and Higher Education for Grants No. NN305 052440 and UMO-2011/03/B/N28/03044 which made this work possible. The authors wish to thank Dr. Paweł Wąsowicz for performing the PCA analysis. This study was conducted with the participation of the students of the Biology Faculty and the Inter-departmental Studies of Environment Protection of the University of Warsaw for their involvement in the present study: Agnieszka Gmiter, Marcin Skrzecz, Hanna Drzazga—examination of G. robertianum plants; Dorota Ziętek, Jakub Wąsowicz-Kiryło, Patrycja Kondej—chemical analyses. The authors of this study wish to thank all the students who participated in collective research.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Wierzbicka, M., Galera, H., Sudnik-Wójcikowska, B. et al. Geranium robertianum L., plant form adapted to the specific conditions along railway: “railway-wandering plant”. Plant Syst Evol 300, 973–985 (2014). https://doi.org/10.1007/s00606-013-0936-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00606-013-0936-1