
Abstract
Pyroleae species are perennial understory shrubs, many of which are partial mycoheterotrophs. Most fungi colonizing Pyroleae roots are ectomycorrhizal (ECM) and share common mycobionts with their Pyroleae hosts. However, such mycobiont sharing has neither been examined in depth before nor has the interspecific variation in sharing among Pyroleae species. Here, we examined root-associated fungal communities in three co-existing Pyroleae species, including Pyrola alpina, Pyrola incarnata, and Orthilia secunda, with reference to co-existing ECM fungi on the surrounding trees in the same soil blocks in subalpine coniferous forests. We identified 42, 75, and 18 fungal molecular operational taxonomic units in P. alpina, P. incarnata, and O. secunda roots, respectively. Mycobiont sharing with surrounding trees, which was defined as the occurrence of the same mycobiont between Pyroleae and surrounding trees in each soil block, was most frequent among P. incarnata (31 of 44 plants). In P. alpina, sharing was confirmed in 12 of 37 plants, and the fungal community was similar to that of P. incarnata. Mycobiont sharing was least common in O. secunda, found in only 5 of 32 plants. Root-associated fungi of O. secunda were dominated by Wilcoxina species, which were absent from the surrounding ECM roots in the same soil blocks. These results indicate that mycobiont sharing with surrounding trees does not equally occur among Pyroleae plants, some of which may develop independent mycorrhizal associations with ECM fungi, as suggested in O. secunda at our research sites.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Most land plants perform photosynthesis and can live autotrophically. In contrast, however, some plants have evolved to obtain carbon from root-colonizing fungi (Leake 2004; Bidartondo 2005). Such carbon dependence on fungi is called mycoheterotrophy (MH) and ranges from full MH among achlorophyllous plants to partial MH (or mixotrophy), in which plants use both photosynthetic and fungus-derived carbon (Selosse and Roy 2009; Merckx 2013). In some plant species, the trophic mode changes with developmental stage. For example, most orchid species exhibit full MH during germination and the early stages of development (initial mycoheterotrophy) but become autotrophic as they mature (Warcup and Talbot 1967; Warcup 1973; Hynson et al. 2013a). However, at least 514 angiosperm species and 1 liverwort species are full MHs throughout their entire life cycle (Merckx 2013). Such a full MH lifestyle has multiple evolutionary origins and is evident in at least 46 independent plant lineages (Merckx 2013).
Mature partial MH plants may be more diverse than full MH plants (Selosse and Roy 2009; Merckx 2013). However, the exact diversity of partial MH plants is unknown and understudied (Selosse et al. 2016), partly because we cannot easily distinguish them from autotrophic plants by appearance, as both have green leaves. Analysis of 13C natural abundance ratios (δ13C) is potentially an effective approach for discriminating partial MH plants because fungi consistently have higher δ13C than autotrophic plants do. Enriched δ13C in some plants compared with surrounding autotrophic ones could potentially indicate the acquisition of fungal carbon and their partial MH status. Using this approach, some species of Pyroleae (syn. Pyroloideae, Angiosperm Phylogeny Website ver. 13, Selosse et al. 2016) have been suggested to be partial MH (Tedersoo et al. 2007; Zimmer et al. 2007; Hynson and Bruns 2009; Matsuda et al. 2012). Green orchids have also been referred to as partial MH species using this approach (Gebauer and Meyer 2003; Julou et al. 2005; Motomura et al. 2008; Roy et al. 2009). However, δ13C levels in plants are also affected by other factors, including water stress, light conditions, and stomatal traits (Farquhar et al. 1989), and enriched δ13C values alone are not sufficient to confirm partial MH status in some cases. Moreover, δ13C enrichment in green Pyroleae is inconsistent among species, conspecific populations, or conspecific individuals within a population (Tedersoo et al. 2007; Zimmer et al. 2007; Hynson and Bruns 2009; Hynson et al. 2015; Johansson et al. 2015). Thus, additional methods of evaluation are necessary to identify species as partial MH.
Additional evidence of partial MH plants is the identity of root-colonizing fungi. In forest ecosystems, many full MH orchids and Monotropoideae (an achlorophyllous subfamily in Ericaceae) species are colonized by ectomycorrhizal (ECM) fungi, while their autotrophic ancestors are associated with other fungal groups (Leake 2004; Bidartondo 2005; Selosse and Roy 2009; Strullu-Derrien et al. 2016). ECM fungi are ubiquitous in forest soil and receive abundant photosynthetic products from host trees (Hobbie 2006; Smith and Read 2008). By establishing common mycorrhizal networks (physical connection through shared ECM fungal mycelia) with surrounding trees, full MH plants can gain access to the abundant photosynthates of trees (Leake 2004; Bidartondo 2005; Selosse and Roy 2009; Motomura et al. 2010; Ogura-Tsujita et al. 2012). Such carbon transfer from trees to full MH plants has been demonstrated in 14C tracer experiments (Bjorkman 1960; McKendrick et al. 2000; Bidartondo et al. 2003). Green-leaved plants that are phylogenetically close to full MH and are associated with ECM fungi are often assumed to obtain carbon through common mycorrhizal networks in a similar way (Selosse et al. 2016).
A diverse set of root-colonizing ECM fungi, including Cortinarius, Tricholoma, Russula, Thelephora/Tomentella, Sebacina, Wilcoxina, and Inocybe, have been shown to colonize Pyroleae (Tedersoo et al. 2007; Zimmer et al. 2007; Massicotte et al. 2008; Vincenot et al. 2008; Hynson and Bruns 2009; Toftegaard et al. 2010; Hashimoto et al. 2012; Matsuda et al. 2012), which are phylogenetically rather close to full MH Monotropoideae (Lallemand et al. 2016). Because these ECM fungi are usually hosted by surrounding trees, it has been hypothesized that Pyroleae and surrounding trees share common mycorrhizal networks. However, this hypothesis has not been tested rigorously, as most of the previous studies did not examine the ECM tips of surrounding trees. A pioneer work by Hashimoto et al. (2012) demonstrated that 4 of 34 fungal RFLP types confirmed in Pyrola asarifolia (syn. P. incarnata) at one stand and 3 of 14 RFLP types at another stand were shared with coexisting Betula trees in the same stands (Table S3 in Hashimoto et al. 2012). Such stand scale fungal sharing, however, does not necessarily indicate physical mycorrhizal networks connecting them (Nara 2006), because the different individuals (genets) of the same fungal species detected in remote soil samples do not form physical networks for carbon transfer between the plants. Thus, it is also possible that Pyroleae plants are independently associated with ECM fungi regardless of the presence of surrounding trees, as in the autotrophic ericaceous shrubs Arbutoideae (Molina and Trappe 1982; Horton et al. 1999; Richard et al. 2005). Recently, Uesugi et al. (2016) advanced the approach of Hashimoto et al. (2012) and showed the sharing of common ECM fungi between Pyrola japonica and ECM trees in the same soil blocks (five of six soil blocks examined). Although the lack of the sharing could not completely deny the networks (e.g., some may extend the networks outside the sampled soil block), the fungal sharing in such a small space gives a strong evidence of the mycorrhizal networks connecting them. This approach deserves application to quantitative and detailed analyses of the mycorrhizal networks between Pyroleae and surrounding ECM trees, since ECM networks are regarded as the key to partial MH in Pyroleae and its ecological adaptation to the dark forest floor.
The colonization of diverse ECM fungi on Pyroleae roots (Tedersoo et al. 2007; Zimmer et al. 2007; Vincenot et al. 2008; Hynson and Bruns 2009; Toftegaard et al. 2010; Hashimoto et al. 2012) contrasts the high fungal specificity in closely related Monotropoideae, each of which is associated with a single fungal species or a very narrow fungal group (Bidartondo 2005 and references therein). Association with different fungi hypothetically enables coexisting Monotropoideae species to avoid competition for fungal carbon resources and niche overlap. While multiple Pyroleae species often coexist in the same forest, it is still unclear how they avoid niche overlap without differentiating mycobionts. Recently, a few Pyroleae species have been shown to associate with fungal communities biased toward a certain fungal group (Matsuda et al. 2012; Hynson et al. 2015; Uesugi et al. 2016). Such fungal preference could potentially enable sympatric niche (mycobiont community) differentiation among coexisting Pyroleae species. Although some pioneer studies have described root-colonizing fungal communities in some coexisting Pyroleae species (Tedersoo et al. 2007; Zimmer et al. 2007), no statistical comparison has been made in these descriptive studies because of the limited number of samples. Therefore, to advance our understanding of the differentiation of root-colonizing fungal communities among coexisting Pyroleae species, it is necessary to compare the communities statistically using sufficient sample sizes.
In this study, we examined root-colonizing fungal communities in the three co-existing Pyroleae species, Pyrola alpina, Pyrola incarnata, and Orthilia secunda, as well as ECM fungi on surrounding trees in the same soil blocks. More than 100 Pyroleae plants were evaluated, and the following hypotheses were tested: (1) ECM fungi sharing between Pyroleae plants and surrounding trees is frequent, regardless of the Pyroleae species; (2) co-existing Pyroleae species are associated with different sets of ECM fungi to avoid competition for mycobiont resources; and (3) co-existing Pyroleae species establish mycelial networks with different tree species to reduce niche overlap.
Materials and methods
Study site
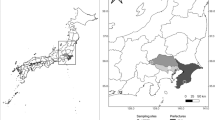
Three forest sites on the northern side of Mt. Fuji (Table S1), Yamanashi Prefecture, Japan, were investigated. The mean annual temperature and precipitation are 3.6 °C and 2766 mm, respectively (The Japan Meteorological Agency 2016). The sites are boreal forests composed of Abies veitchii and Larix kaempferi mixed with Salix reinii, Alnus sieboldiana, and Betula ermanii shrubs. Three Pyroleae species, P. alpina, P. incarnata, and O. secunda, also co-exist on the forest floors (Electronic Supplementary Fig. S1).
Sampling
Between 31 and 34 soil blocks (15 × 15 × 10 cm, length × width × depth) containing at least one Pyroleae plant per block were collected at each site in September 2013 and 2014. Pyroleae species were at least 10 m apart. In total, 95 soil blocks, including 113 Pyroleae plants, were collected (37, 44, and 32 individuals of P. alpina, P. incarnata, and O. secunda, respectively). Soil samples were collected in individual plastic bags and stored in a cool box prior to analysis.
Root samples for molecular analyses
All roots were manually separated into Pyroleae roots (by tracing back from the plant base along rhizomes), washed with tap water, and examined using a dissecting microscope (SZX7, Olympus, Tokyo). The root tips of two Pyrola species had distinguishable morphological traits with obvious mantle formation or sparse extraradical hyphae (Massicotte et al. 2008; Vincenot et al. 2008) and, thus, were classified into morphotypes (Fig. S1) (Massicotte et al. 2008). Up to three root tip replicates (depending on availability) per morphotype were subjected to DNA analysis. O. secunda, however, had no obvious morphotype or mantle (Fig. S1) (Massicotte et al. 2008). Examination of O. secunda roots using 10 random root fragments from each of nine plants in 2013 revealed that fungal colonization occurred mostly in swollen lateral root regions, often accompanied by brown pigments (Fig. S1). Notably, such findings were rare in other root regions. To improve fungal detection frequency, we excised the former root regions of O. secunda, sectioned them into 2–4 mm fragments and randomly selected 4–10 fragments from each plant for fungal identification. To evaluate common mycelial networks between Pyroleae plants and surrounding trees, ECM roots from each soil block were collected and morphotyped, and fungi were identified, as described elsewhere (Murata et al. 2013). In total, 853 Pyroleae and 1273 ECM root samples were used for molecular identification.
Fungal identification
DNA extraction and molecular identification followed that described elsewhere (Miyamoto et al. 2015). Briefly, each Pyroleae or ECM root tip was placed in a 2.0-ml tube, pulverized using a bead-beater, and DNA was extracted using a modified cetyl trimethyl ammonium bromide method. The internal transcribed spacer (ITS) regions of fungal rDNA were amplified by polymerase chain reaction (PCR) using the forward primer ITS1F and the reverse primers ITS4, LR21, LR22, and LBW with a Multiplex PCR kit (QIAGEN, Hilden, Germany), following the manufacturer’s instructions. PCR products were assessed on 1.2% agarose gels (0.5× TBE buffer) using Gel Red (Biotium, Fremont, CA, USA) under UV light (Benchtop 2UV Transilluminator, UVP, Cambridge, UK). Amplification products were purified using a PCR clean-up kit containing exonuclease I and shrimp alkaline phosphatase (GE Healthcare, Hertfordshire, UK). Sanger sequencing was performed using the Big Dye Terminator version 3.1 Cycle Sequencing kit (Applied Biosystems, Foster City, CA, USA) and ITS1 or ITS4. Sequencing was performed on the ABI 3130 XL Genetic Analyzer (Applied Biosystems). Sequences were manually corrected using the chromatograms and assembled into molecular operational taxonomic units (OTUs) using ATGC ver. 7 (GENETYX, Tokyo, Japan) with a ≥97% similarity threshold. One representative sequence for each OTU was used for BLAST searches to identify the OTU (hereafter referred to as “species” for simplicity). To identify the host tree species associated with each mycobiont, the trnL region of chloroplast DNA was amplified using the trnC–trnD and trnE–trnF primer pairs and then sequenced (Liu et al. 2011).
Statistical analyses
The frequency of each fungal species was defined as the number of plants (or soil blocks) colonized by that fungus in each Pyroleae species. The relative abundance of a fungal species was defined as the percentage of Pyroleae root tips containing each fungal species. To compare fungal diversity, Jackknife2 richness estimator for each Pyroleae species was calculated with EstimateS ver. 9.1.0. (Colwell et al. 2012). To evaluate the fungal communities among plant species, non-metric multidimensional scaling (NMDS) was performed using the vegan package in R ver. 3.2.3 (R Core Team 2015). To calculate Bray–Curtis distances, 999 permutations were performed based on a fungal community matrix containing the fungal frequency data for each plant species at each site. Singleton fungal species and fungi from unidentified plant hosts were excluded prior to analysis. Statistical differences in root-associated fungal communities among sites, among Pyroleae species, and between Pyroleae and ECM trees were determined using ADONIS (permutation-based multivariate analysis of variance). A total of 999 permutations were performed for all such analyses.
Network architecture
The architecture of mycobiont sharing was visualized based on the Fruchterman-Reingold algorithm (Fruchterman and Reingold 1991) implemented in the igraph package for R (Ognyanova 2015), using the mycobiont sharing data between each Pyroleae species and surrounding ECM trees within the same soil blocks. The same analysis was performed for the data set including all plant-fungal occurrence in three study sites for visualizing land scape level fungal sharing (Toju et al. 2013), although it does not necessarily indicate direct (physical) mycorrhizal networks for MH carbon transfer.
Results
General description of the root-associated fungal community in Pyroleae and ectomycorrhizal trees
A total of 180 putative mycorrhizal fungal species were identified from the roots of Pyroleae and surrounding trees. Basidiomycetes were dominant (164 spp.), while 16 species (9%) belonged to ascomycetes (Table S2). The Jackknife2 estimator of species richness was 307.
Mycobionts in O. secunda
Forty-eight (33.8%) of 142 root tips from 32 plants were sequenced. Mycobionts were not identified from 12 plants. In total, 18 fungal species were identified using a >97% ITS similarity threshold, and ~44 species were identified in O. secunda roots using Jackknife2 (Table 1, Fig. 1 and Table S2). The most frequent lineage in O. secunda roots was Wilcoxina, which was found in 22 of 48 root fragments and in 12 of 20 plant individuals (Tables 2 and S2). Phialocephala was the second most frequent lineage, found in four root fragments from four plants. Russula and Sebacina were detected once in each of three plant individuals. The fungal richness (mean ± SE) in O. secunda roots per soil block was 1.6 ± 0.2 or 1.0 ± 0.2 if uncolonized plants were included (Table 1).

Species accumulation curves of a observed species richness and b estimated richness (Jackknife2) for each Pyroleae species and all ectomycorrhizal trees coexisting on Mount Fuji, Japan
Mycobionts in P. alpina
Ninety-five (40.8%) of 233 root tips from 37 plants were sequenced. These sequences were grouped into 42 species (~105 using Jackknife2; Table 1 and Fig. 1), including the diverse ECM fungal lineages. Cortinarius, Amphinema, and Thelephora/Tomentella were found in 11, 8, and 5 P. alpina individuals, respectively (Table 2, Table S2). Other ECM fungal genera, including Sebacina, Russula, Inocybe, Hebeloma, Hygrophorus, and Piloderma, were detected in two to four plant individuals. Most mycobionts belonged to Basidiomycetes, while only two belonged to Ascomycetes. The fungal richness in P. alpina roots per soil block was 1.8 ± 0.2 (Table 1).
Mycobionts in P. incarnata
Three hundred and seven (63.6%) of 483 tips from 44 plants were sequenced and grouped into 75 fungal species (~156 using Jackknife2, Table 1). Similar to P. alpina, mycobionts in P. incarnata roots were dominated by the ECM fungal genera Cortinarius and Thelephora/Tomentella (Table 2, Table S2), which were detected in 29 plants. Other frequently observed genera were Laccaria, Amphinema, Sebacina, Russula, Inocybe, and Hebeloma, each of which was found in five to nine plants. As in P. alpina, Basidiomycetes ECM fungi were dominant (Fig. 2). The fungal richness in P. incarnata roots per soil block was 3.4 ± 0.2, which was significantly higher than that in the other two Pyroleae species (p < 0.05, Tukey’s test; Table 1).

Relative abundances of fungal lineages associated with roots of Pyroleae and ectomycorrhizal trees coexisting in subalpine forests on Mount Fuji, Japan. Data from Table S2
Ectomycorrhizal fungi on surrounding trees in Pyroleae-containing soil blocks
Of 1267 ECM tips from surrounding trees, 768 (61%) were sequenced. In total, 142 fungal species (243 using Jackknife2) were identified from a total of 95 soil blocks. The dominant ECM fungi were Cortinarius (59 soil blocks), Russula/Lactarius (47 blocks), Cenococcum (42 blocks), and Thelephora/Tomentella (42 blocks), followed by Inocybe (23 blocks), Suillus (14 blocks), Piloderma (12 blocks), Laccaria (11 blocks), Hygrophorus (10 blocks), Hebeloma (8 blocks), and Sebacina (5 blocks) (Table S2). Clavulina, Amphinema, and Amanita were each found in three or fewer soil blocks. Using chloroplast DNA sequences, host tree species were identified in 85% of ECM tips. Larix, Abies, and Betula were dominant ECM hosts, found in 70, 47, and 33 soil blocks, respectively (Table 1). The ECM fungal richness values (mean ± SE) per soil block containing O. secunda, P. alpina, and P. incarnata were 4.0 ± 0.4, 4.4 ± 0.2, and 3.6 ± 0.1, respectively (p > 0.05, Table 1).
Mycorrhizal fungal communities among Pyroleae and canopy trees on a landscape scale
Of 180 fungal species identified, 66 were found in both Pyroleae and ECM trees, after pooling all soil blocks from the three sites. These common species on a landscape scale included 12 of 18, 26 of 42, and 53 of 75 fungal species in O. secunda, P. alpina, and P. incarnata roots, respectively (Table S2). Cenococcum and Lactarius were rarely confirmed in Pyroleae roots, in contrast to their abundance on ECM tips of the surrounding trees (Fig. 2 and Table 2).
We used NMDS and ANONIS analyses to evaluate the differences in root-associated fungal communities among Pyroleae and ECM tree species. After excluding the minor hosts (i.e. <5 soil blocks per site, singleton fungal species, fungi on unidentified hosts), the data set used for the community analyses contained 107 fungal species. Fungal communities were separated by plant species rather than by sites in the NMDS plot (Fig. 3). Fungal communities of O. secunda were separated from those of other plant species. Fungi in P. alpina roots were partially separated from those in ECM trees, with the exception of Larix in Site 3. Additionally, fungal communities of P. incarnata were in close proximity to those of ECM trees at the same site.

Non-metric multidimensional scaling plot showing similarity in root-associated fungal communities of Pyroleae and surrounding ectomycorrhizal trees on Mount Fuji, Japan. Hosts represented by <six soil samples were excluded before the analyses. Bray-Stress = 0.184. Circles represent the communities on ectomycorrhizal trees, where Abies (A), Betula (B), and Larix (L) are in white, black, and gray, respectively. Other open symbols represent the communities on Pyroleae spp., where Orthilia secunda (Os), Pyrola alpina (Pa), and Pyrola incarnata (Pi) are in squares, diamonds, and triangles, respectively. Site numbers are shown after host abbreviations
The ADONIS test confirmed that the data variance in the fungal communities was significantly explained by plant species (F 5, 12 = 2.44, r 2 = 0.50, p = 0.001), while the effect of sites was not significant (F 2, 15 = 1.02, r 2 = 0.12, p = 0.417). We further compared the fungal communities among Pyroleae species, after excluding ECM trees and found a significant difference among Pyroleae species (F 2, 6 = 2.68, r 2 = 0.47, p = 0.007), but not among sites (F 2, 6 = 0.67, r 2 = 0.18, p = 0.833).
While the occurrence of common fungal species between Pyroleae and ECM trees on the landscape scale does not necessarily indicate direct physical connection for MH carbon transfer, P. incarnata is connected to ECM trees by many fungal species in a landscape-scale network plot (Fig. S2). In contrast, O. secunda and ECM trees had far fewer common fungal species across the study sites (Fig. S2).
Mycobiont sharing between Pyroleae and canopy tree species on a soil block scale
Mycobiont sharing between Pyroleae and canopy trees was defined as the presence of the same fungal species in both plant groups within a soil block, assuming common mycorrhizal networks between them. In O. secunda, mycobiont sharing was rare, as it was confirmed in only 5 of 31 (16.1%) mycobionts in O. secunda roots and in 5 of 32 O. secunda individuals (Table 3, Fig. 4a). Three canopy tree genera (Larix, Abies, and Alnus) shared mycobionts with O. secunda, although they represented only 5 of 51 ECM trees in the same blocks (Table 3, Fig. 4a). Of the five shared mycobionts, three belonged to Cortinarius. Although Wilcoxina was the most dominant mycobiont lineage in O. secunda roots (Table 2, Fig. S2), it was not confirmed in any of the surrounding ECM tips within the same soil blocks (p < 0.001).


Confirmed mycobiont sharing between Pyroleae and ectomycorrhizal (ECM) trees within the same soil blocks collected in subalpine forests on Mount Fuji, Japan. a Orthilia secunda. b Pyrola alpina. c Pyrola incarnata. Data from Table 3. White circles represent mycobiont species confirmed in roots of both Pyroleae and ECM trees within the same soil blocks. Line width is proportional to the number of occurrence, i.e., the number of soil blocks or plant individuals that contained the fungus. Because of the sharing within the small space (soil block), line connections between plants in these panels strongly suggest physical networks enabling MH carbon transfer. Note that very few thin connections to ECM trees from O. secunda but many (and some thick) connections from Pyrola incarnata
In P. alpina, sharing was confirmed in 12 of 37 plants by 19 of 57 (33.3%) mycobiont individuals (Table 3). Three canopy tree genera, including Larix, Abies, and Betula, shared mycobionts with P. alpina (Fig. 4b), accounting for 16 of 61 ECM trees confirmed in the same blocks (Table 3). Of 19 mycobionts (13 species) shared with surrounding ECM trees, 8 (three species), 2, and 2 belonged to Cortinarius, Hebeloma, and Inocybe, respectively (Table 3, Fig. 4b). Although Amphinema was the second most dominant mycobiont lineage in P. alpina (Table 2), it was not confirmed in surrounding ECM tips within the same blocks (p < 0.001, Fig. S2). Conversely, Cenococcum was frequently found in the surrounding ECM tips but rarely found in P. alpina roots (Table 2, Fig. S2).
Mycobiont sharing with canopy trees was most frequently observed in P. incarnata (Fig. 4c). Such sharing was confirmed in 30 of 44 plant individuals by 44 of 150 (29.3%) mycobionts (Table 3). Of 44 mycobiont individuals (27 species) shared with surrounding canopy trees, 23 (8 species) and 8 (8 species) belonged to Cortinarius and Thelephoraceae, respectively (Table 3). Trees sharing common mycobionts with P. incarnata included Larix, Abies, Betula, and Salix (Fig. 4c), collectively representing more than half of the trees detected within the same soil blocks. Similar to P. alpina, Cenococcum was rarely found in P. incarnata roots; however, it was frequently found in the surrounding ECM tips (p < 0.001, Table 2). The occurrence of Lactarius was also negatively biased against P. incarnata (p < 0.01, Table 2). Conversely, Thelephoraceae was more prevalent in P. incarnata roots, compared with its prevalence in the surrounding ECM tips (p < 0.01, Table 2).
The occurrence of mycobiont sharing was significantly different among Pyroleae species (Fig. 4a–c). The lowest proportion of sharing was found in O. secunda, while the highest was found in P. incarnata (Table 3, p < 0.001, Fisher’s exact test). This was further confirmed by network plots (Fig. 4), where there are few connections between O. secunda and ECM trees but many connections between P. incarnata and ECM trees within the same soil blocks.
While Larix, Abies, and Betula roots coexisted with all Pyroleae species, the proportion of shared trees was significantly different among Pyroleae (Table 3). For example, of the 18 Larix trees coexisting with O. secunda, only 2 were shared with common mycobionts. The proportion of Larix trees shared with P. incarnata was much higher (p = 0.009, Table 3). Such differences in the shared tree proportion among Pyroleae species were also observed in Abies (p = 0.001) (Table 3).
Discussion
Root-associated fungi in three coexisting Pyroleae species were dominated by putative ECM fungal lineages, most of which have been confirmed in ECM trees in this study or in previous studies on Mt. Fuji (Nara et al. 2003; Miyamoto et al. 2014; Miyamoto et al. 2015). The dominance of ECM fungal lineages has been repeatedly shown in previous studies as mentioned in the “Introduction.” Most of these studies assumed common mycorrhizal networks linking Pyroleae and surrounding ECM trees; however, such networks have not been rigorously tested. Only one study documented the actual sharing of mycobionts between P. japonica and surrounding trees in five of six soil blocks (Uesugi et al. 2016), but the small sample size needed further confirmation. In the current study, P. incarnata shared common mycobionts with surrounding trees in nearly 70% of soil blocks, indicating that common mycorrhizal networks are common in this species. However, mycobiont sharing was confirmed in only 15% of soil blocks containing O. secunda and in 32% soil blocks containing P. alpina. The limited sharing in O. secunda was also confirmed in the analyses based on mycobiont individuals, i.e., only 16% mycobionts confirmed in O. secunda (yet all of them were minor, potentially indicating surface adhesion) was shared with surrounding trees, while about 30% mycobionts in two Pyrola species (many of them were dominant colonizers detected in multiple roots) were shared with the trees. Such an observation contradicts our first hypothesis. Since common mycorrhizal networks are nearly ubiquitous among ECM trees of the same species (Lian et al. 2006) and between different ECM trees (Nara and Hogetsu 2004; Nara 2006), the lower frequencies of mycobiont sharing between coexisting pairs were unexpected. Thus, some Pyroleae species, especially O. secunda in this study, may avoid sharing common mycobionts with surrounding trees and possibly maintain independent associations with ECM fungi. While the detection of ECM fungi in Pyroleae roots has been regarded as evidence of mycorrhizal networks with surrounding trees, we should be cautious about such assumptions.
Given the methodological limitations of this study (e.g. small soil blocks and small root sections for DNA analyses), it is difficult to completely exclude the potential for mycorrhizal networks between Pyroleae and ECM trees by the lack of their detection. Indeed, the fungal sharing data documented here would surely be underestimates due to undetected minor colonization, potential networks beyond the soil blocks, and ephemeral connections in other seasons. However, the lower frequencies of mycobiont sharing in O. secunda or P. alpina, compared with that of P. incarnata, are irrelevant to such methodological limitations, since the same sampling and analytical methods were used throughout this study. Similarly, the number of colonized roots was lower in O. secunda and P. alpina compared with P. incarnata. We are not sure how such a lower frequency of mycobiont sharing and lower rate of fungal colonization in O. secunda could effectively function in exploring for fungal carbon or in being epiparasitic to surrounding trees for carbon. Less morphological differentiation in fungus-colonized roots of O. secunda (diffuse roots with no fungal mantle; Fig. S1) may also be indicative of the limited carbon dependency on fungi, since full MH relatives (Monotropoideae) exhibit considerable morphological root differentiation (e.g. a “root ball” composed of a dense cluster of fungus-colonized short roots with well-developed mantles). While the root system of P. incarnata was still diffuse compared with that of Monotropoideae, fungus-colonized roots had well-developed fungal mantles (Fig. S1). Collectively, mycoheterotrophic levels may be high in P. incarnata but low or absent in O. secunda. Interestingly, our data regarding stable isotope signatures and photosynthetic traits examined within the same sites also support this hypothesis (Jia et al. in preparation).
Root-associated ECM fungi detected in two Pyrola species were diverse; however, the frequency of Cenococcum in both Pyrola species was significantly less than that of surrounding ECM tips. Similarly, the frequency of Lactarius was also lower in both Pyrola species but not statistically different (p = 0.119) in P. alpina. Antagonism toward Ascomycetes and Lactarius was suggested by Hashimoto et al. (2012) and was statistically confirmed, for the first time, by this study. Conversely, preferences toward specific fungal lineages were also statistically confirmed in the two Pyrola species (e.g. Thelephoraceae in P. incarnata and Amphinema in P. alpina). Compared with the two Pyrola species, O. secunda was associated with fewer fungal lineages and was dominated exclusively by Wilcoxina. The colonization of Wilcoxina in O. secunda roots has been documented in previous studies, but in one or two plants mainly due to the limited number of samples used (Tedersoo et al. 2007; Zimmer et al. 2007). Wilcoxina forms ectendomycorrhiza on conifer and broadleaf trees in nurseries, burned sites, and disturbed sites (Yang and Korf 1985; Egger 1996), but its ecology in old growth forests is unknown.
The occurrence of Wilcoxina on O. secunda roots and not on the surrounding ECM roots suggests an independent symbiotic association of O. secunda with Wilcoxina. Because Wilcoxina mycelium is described as “short distance exploration type” (Agerer 2001; Smith and Read 2008), its mycelia would less likely to extend beyond the soil block to form networks without colonizing nearby ECM roots. Moreover, in Tedersoo et al. (2007), 10 of 19 fungal species detected in O. secunda roots were non-ECM fungal lineages, in which root endophytes like Phialocephala and Helotiales were far more abundant than ECM fungi in relative abundance. This corresponds with our results and may imply less ECM networks between O. secunda and surrounding trees even in Europe, yet rigorous tests like our approach would be needed before conclusion. In fact, the mycoheterotrophy levels for O. secunda estimated from 13C signatures are inconsistent among sites, suggesting a lack of tripartite networks in some locations (Tedersoo et al. 2007; Zimmer et al. 2007; Johansson et al. 2015).
With contrasting fungal preference/antagonism, the three co-existing Pyroleae species were associated with significantly different communities of root-colonizing fungi. Although the two Pyrola species were associated with similar ECM fungal lineages, they differed in mycobiont sharing patterns. P. incarnata was incorporated into surrounding ECM networks intensively, whereas P. alpina was not so intensively. These data support our second hypothesis and suggest that the co-existing Pyroleae species avoid niche overlaps by associating with different fungi (Bever et al. 2010). Given the higher frequency of mycobiont sharing among P. incarnata with surrounding trees, carbon supply from surrounding trees through mycorrhizal networks would be critical in this species (Tedersoo et al. 2007; Zimmer et al. 2007; Matsuda et al. 2012). By contrast, the lower frequency of mycobiont sharing and association with unique fungal communities in O. secunda raise doubts on carbon supply from surrounding trees in some locations. O. secunda may seek other resources, such as nitrogen, from root-associated fungi (Hynson et al. 2013b).
The few connections to surrounding trees in O. secunda partly collapse our third hypothesis. Yet, the two Pyrola species were associated with slightly different proportions of the surrounding trees, i.e., P. incarnata was connected to Abies, Betula, and Larix rather evenly (Table 3, Fig. 4c), while P. alpina shared common mycotionts with Larix mostly (Table 3, Fig. 4b). Thus, the two Pyrola species may partly avoid niche overlaps by differentiating host trees for sharing, in addition to the different abundance of the sharing.
Pyroleae is a unique plant lineage that includes autotrophic (Zimmer et al. 2007), partial MH (Tedersoo et al. 2007; Zimmer et al. 2007), and full MH (Hynson and Bruns 2009) species and provides an opportunity to investigate its evolution from an autotrophic to full MH species (Lallemand et al. 2016). In Orchidaceae, the evolution of full MH species is hypothesized to occur with changes in root-associated fungi from the saprotrophic Rhizoctonia to ECM fungi and from a broad to narrow range of mycorrhizal fungi (Leake 2004; Bidartondo 2005; Selosse and Roy 2009; Motomura et al. 2010; Ogura-Tsujita et al. 2012). However, full MH Pyrola aphylla was associated with a broad range of ECM fungi, suggesting that the situation in Orchidaceae is not applicable to the evolution of mycoheterotrophy in Pyroleae (Hynson and Bruns 2009). While most previous studies on Pyroleae mycobiont communities are descriptive based on fewer samples, the quantitative and community analyses based on enough samples including surrounding ECM roots would be necessary for detecting inconspicuous but significant preference or antagonism to certain fungal lineage. The accumulation of such data in other geographical locations and other Pyroleae species would advance our understanding about the evolution of their mycobiont associations, and its geographical variance or local adaptation.
References
Agerer R (2001) Exploration types of ectomycorrhizae—a proposal to classify ectomycorrhizal mycelial systems according to their patterns of differentiation and putative ecological importance. Mycorrhiza 11:107–114. doi:10.1007/s005720100108
Bever JD et al (2010) Rooting theories of plant community ecology in microbial interactions. Trends Ecol Evol 25:468–478. doi:10.1016/j.tree.2010.05.004
Bidartondo MI (2005) The evolutionary ecology of myco-heterotrophy. New Phytol 167:335–352. doi:10.1111/j.1469-8137.2005.01429.x
Bidartondo MI, Bruns TD, Weiss M, Sergio C, Read DJ (2003) Specialized cheating of the ectomycorrhizal symbiosis by an epiparasitic liverwort. Proc R Soc Lond B 270:835–842. doi:10.1098/rspb.2002.2299
Bjorkman E (1960) Monotropa Hypopitys L.—an Epiparasite on tree roots. Physiol Plant 13:308–327. doi:10.1111/j.1399-3054.1960.tb08034.x
Colwell RK, Chao A, Gotelli NJ, Lin SY, Mao CX, Chazdon RL, Longino JT (2012) Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J Plant Ecol 5:3–21. doi:10.1093/jpe/rtr044
Egger KN (1996) Molecular systematics of E-strain mycorrhizal fungi: Wilcoxina and its relationship to Tricharina (Pezizales). Can J Bot 74:773–779
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annu Rev Plant Physiol 40:503–537. doi:10.1146/annurev.arplant.40.1.503
Fruchterman TM, Reingold EM (1991) Graph drawing by force-directed placement. Software: Pract Exper 21(11):1129–1164
Gebauer G, Meyer M (2003) 15N and 13C natural abundance of autotrophic and myco-heterotrophic orchids provides insight into nitrogen and carbon gain from fungal association. New Phytol 160:209–223. doi:10.1046/j.1469-8137.2003.00872.x
Hashimoto Y, Fukukawa S, Kunishi A, Suga H, Richard F, Sauve M, Selosse MA (2012) Mycoheterotrophic germination of Pyrola asarifolia dust seeds reveals convergences with germination in orchids. New Phytol 195:620–630. doi:10.1111/j.1469-8137.2012.04174.x
Hobbie EA (2006) Carbon allocation to ectomycorrhizal fungi correlates with belowground allocation in culture studies. Ecology 87:563–569. doi:10.1890/05-0755
Horton TR, Bruns TD, Parker VT (1999) Ectomycorrhizal fungi associated with Arctostaphylos contribute to Pseudotsuga menziesii establishment. Can J Bot 77:93–102
Hynson NA, Bruns TD (2009) Evidence of a myco-heterotroph in the plant family Ericaceae that lacks mycorrhizal specificity. Proc Biol Sci 276:4053–4059. doi:10.1098/rspb.2009.1190
Hynson NA, Madsen TP, Selosse MA, Adam IKU, Ogura-Tsujita Y, Roy M, Gebauer G (2013a) The physiological ecology of mycoheterotrophy. In: Merckx V (ed) Mycoheterotrophy: the biology of plants living on fungi. Springer, pp 297–342. doi:10.1007/978-1-4614-5209-6_8
Hynson NA, Weiss M, Preiss K, Gebauer G, Treseder KK (2013b) Fungal host specificity is not a bottleneck for the germination of Pyroleae species (Ericaceae) in a Bavarian forest. Mol Ecol 22:1473–1481. doi:10.1111/mec.12180
Hynson NA, Bidartondo MI, Read DJ (2015) Are there geographic mosaics of mycorrhizal specificity and partial mycoheterotrophy? A case study in Moneses uniflora (Ericaceae). New Phytol 208:1003–1007. doi:10.1111/nph.13587/full
Johansson VA, Mikusinska A, Ekblad A, Eriksson O (2015) Partial mycoheterotrophy in Pyroleae: nitrogen and carbon stable isotope signatures during development from seedling to adult. Oecologia 177:203–211. doi:10.1007/s00442-014-3137-x
Julou T, Burghardt B, Gebauer G, Berveiller D, Damesin C, Selosse MA (2005) Mixotrophy in orchids: insights from a comparative study of green individuals and nonphotosynthetic individuals of Cephalanthera damasonium. New Phytol 166:639–653. doi:10.1111/j.1469-8137.2005.01364.x
Lallemand F, Gaudeul M, Lambourdière J, Matsuda Y, Hashimoto Y, Selosse MA (2016) The elusive predisposition to mycoheterotrophy in Ericaceae. New Phytol 212:314–319. doi:10.1111/nph.14092
Leake JR (2004) Myco-heterotroph/epiparasitic plant interactions with ectomycorrhizal and arbuscular mycorrhizal fungi. Curr Opin Plant Biol 7:422–428. doi:10.1016/j.pbi.2004.04.004
Lian CL, Narimatsu M, Nara K, Hogetsu T (2006) Tricholoma matsutake in a natural Pinus densiflora forest: correspondence between above- and below-ground genets, association with multiple host trees and alteration of existing ectomycorrhizal communities. New Phytol 171:825–836. doi:10.1111/j.1469-8137.2006.01801.x
Liu ZW, Wang ZH, Zhou J, Peng H (2011) Phylogeny of Pyroleae (Ericaceae): implications for character evolution. J Plant Res 124:325–337. doi:10.1007/s10265-010-0376-8
Massicotte HB, Melville LH, Tackaberry LE, Peterson RL (2008) A comparative study of mycorrhizas in several genera of Pyroleae (Ericaceae) from western Canada. Botany-Botanique 86:610–622. doi:10.1139/B08-027
Matsuda Y, Shimizu S, Mori M, Ito S, Selosse MA (2012) Seasonal and environmental changes of mycorrhizal associations and heterotrophy levels in mixotrophic Pyrola japonica (Ericaceae) growing under different light environments. Am J Bot 99:1177–1188. doi:10.3732/ajb.1100546
McKendrick SL, Leake JR, Read DJ (2000) Symbiotic germination and development of myco-heterotrophic plants in nature: transfer of carbon from ectomycorrhizal Salix repens and Betula pendula to the orchid Corallorhiza trifida through shared hyphal connections. New Phytol 145:539–548. doi:10.1046/j.1469-8137.2000.00592.x
Merckx VSFT (2013) Mycoheterotrophy: the biology of plants living on fungi. Springer, New York. doi:10.1007/978-1-4614-5209-6
Miyamoto Y, Nakano T, Hattori M, Nara K (2014) The mid-domain effect in ectomycorrhizal fungi: range overlap along an elevation gradient on Mount Fuji, Japan. ISME J 8:1739–1746. doi:10.1038/ismej.2014.34
Miyamoto Y, Sakai A, Hattori M, Nara K (2015) Strong effect of climate on ectomycorrhizal fungal composition: evidence from range overlap between two mountains. ISME J 9:1870–1879. doi:10.1038/ismej.2015.8
Molina R, Trappe JM (1982) Lack of mycorrhizal specificity by the ericaceous hosts Arbutus menziesii and Arctostaphylos uva-ursi. New Phytol 90:495–509. doi:10.1111/j.1469-8137.1982.tb04482.x
Motomura H, Yukawa T, Ueno O, Kagawa A (2008) The occurrence of crassulacean acid metabolism in Cymbidium (Orchidaceae) and its ecological and evolutionary implications. J Plant Res 121:163–177. doi:10.1007/s10265-007-0144-6
Motomura H, Selosse MA, Martos F, Kagawa A, Yukawa T (2010) Mycoheterotrophy evolved from mixotrophic ancestors: evidence in Cymbidium (Orchidaceae). Ann Bot 106:573–581. doi:10.1093/aob/mcq156
Murata M, Kinoshita A, Nara K (2013) Revisiting the host effect on ectomycorrhizal fungal communities: implications from host-fungal associations in relict Pseudotsuga japonica forests. Mycorrhiza 23:641–653. doi:10.1007/s00572-013-0504-0
Nara K (2006) Ectomycorrhizal networks and seedling establishment during early primary succession. New Phytol 169:169–178. doi:10.1111/j.1469-8137.2005.01545.x
Nara K, Hogetsu T (2004) Ectomycorrhizal fungi on established shrubs facilitate subsequent seedling establishment of successional plant species. Ecology 85:1700–1707. doi:10.1890/03-0373
Nara K, Nakaya H, Hogetsu T (2003) Ectomycorrhizal sporocarp succession and production during early primary succession on Mount Fuji. New Phytol 158:193–206. doi:10.1046/j.1469-8137.2003.00724.x
Ognyanova K (2015) Network visualization with R. POLNET 2015 Workshop, Portland. Available at http://www.kateto.net
Ogura-Tsujita Y, Yokoyama J, Miyoshi K, Yukawa T (2012) Shifts in mycorrhizal fungi during the evolution of autotrophy to mycoheterotrophy in Cymbidium (Orchidaceae). Am J Bot 99:1158–1176. doi:10.3732/ajb.1100464
R Core Team (2015) A language and environment for statistical computing. R foundation for statistical computing, Vienna. Available from http://www.R-project.org/
Richard F, Millot S, Gardes M, Selosse MA (2005) Diversity and specificity of ectomycorrhizal fungi retrieved from an old-growth Mediterranean forest dominated by Quercus ilex. New Phytol 166:1011–1023. doi:10.1111/j.1469-8137.2005.01382.x
Roy M, Watthana S, Stier A, Richard F, Vessabutr S, Selosse MA (2009) Two mycoheterotrophic orchids from Thailand tropical dipterocarpacean forests associate with a broad diversity of ectomycorrhizal fungi. BMC Biol 7:51. doi:10.1186/1741-7007-7-51
Selosse MA, Roy M (2009) Green plants that feed on fungi: facts and questions about mixotrophy. Trends Plant Sci 14:64–70. doi:10.1016/j.tplants.2008.11.004
Selosse MA, Bocayuva MF, Kasuya MCM, Courty PE (2016) Mixotrophy in mycorrhizal plants: extracting C from mycorrhizal networks. In: Martin F (ed) Molecular mycorrhizal symbiosis. Wiley, pp 451–471
Smith SE, Read DJ (2008) Mycorrhizal symbiosis, 3rd edn. Academic, London
Strullu-Derrien C, Kenrick P, Selosse MA (2016) Origins of the mycorrhizal symbioses. In: Martin F (ed) Molecular mycorrhizal symbiosis. Wiley, pp 1–20. doi:10.1002/9781118951446.ch1
The Japan Meteorological Agency (2016) Mesh average 2016. Japan meteorological business support center, Tokyo
Tedersoo L, Pellet P, Koljalg U, Selosse MA (2007) Parallel evolutionary paths to mycoheterotrophy in understorey Ericaceae and Orchidaceae: ecological evidence for mixotrophy in Pyroleae. Oecologia 151:206–217. doi:10.1007/s00442-006-0581-2
Toftegaard T, Iason GR, Alexander IJ, Rosendahl S, Taylor AFS (2010) The threatened plant intermediate wintergreen (Pyrola media) associates with a wide range of biotrophic fungi in native Scottish pine woods. Biodivers Conserv 19:3963–3971. doi:10.1007/s10531-010-9940-8
Toju H et al (2013) How are plant and fungal communities linked to each other in belowground ecosystems? A massively parallel pyrosequencing analysis of the association specificity of root-associated fungi and their host plants. Ecol Evol 3:3112–3124. doi:10.1002/ece3.706
Uesugi T, Nakano M, Selosse MA, Obase K, Matsuda Y (2016) Pyrola japonica, a partially mycoheterotrophic Ericaceae, has mycorrhizal preference for russulacean fungi in central Japan. Mycorrhiza 26:819–829. doi:10.1007/s00572-016-0715-2
Vincenot L, Tedersoo L, Richard F, Horcine H, Koljalg U, Selosse MA (2008) Fungal associates of Pyrola rotundifolia, a mixotrophic Ericaceae, from two Estonian boreal forests. Mycorrhiza 19:15–25. doi:10.1007/s00572-008-0199-9
Warcup JH (1973) Symbiotic germination of some Australian terrestrial orchids. New Phytol 72:387–392. doi:10.1111/j.1469-8137.1973.tb02046.x
Warcup JH, Talbot PHB (1967) Perfect states of Rhizoctonias associated with orchids. New Phytol 66:631. doi:10.1111/j.1469-8137.1967.tb05434.x
Yang CS, Korf RP (1985) A monograph of the genus Tricharina and of a new, segregate genus, Wilcoxina (Pezizales). Mycotaxon 24:467–531
Zimmer K, Hynson NA, Gebauer G, Allen EB, Allen MF, Read DJ (2007) Wide geographical and ecological distribution of nitrogen and carbon gains from fungi in pyroloids and monotropoids (Ericaceae) and in orchids. New Phytol 175:166–175. doi:10.1111/j.1469-8137.2007.02065.x
Acknowledgements
We thank Naoki Shiomi for field assistance, Dr. Masao Murata and Dr. Zhugui Wen for lab assistance, and Dr. Kenshiro Oshima for assistance of sequencing. This work was supported by JSPS KAKENHI Grant Number 15K14750.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Supplementary Fig S1
(DOCX 412 kb)
Supplementary Fig S2
(DOCX 412 kb)
Supplementary Table S1
(DOCX 13 kb)
Supplementary Table S2
(DOCX 63 kb)
Supplementary Table S3
(DOCX 22 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Jia, S., Nakano, T., Hattori, M. et al. Root-associated fungal communities in three Pyroleae species and their mycobiont sharing with surrounding trees in subalpine coniferous forests on Mount Fuji, Japan. Mycorrhiza 27, 733–745 (2017). https://doi.org/10.1007/s00572-017-0788-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00572-017-0788-6