
Abstract
Key message
Stem radius changes measured at high temporal resolution in a drought-limited southern boreal forest were primarily related to soil temperature and partly soil moisture variation, but only weakly to VPD.
Abstract
Forest productivity at the southern fringe of the boreal forest biome in Inner Asia is strongly drought-limited, as is evident from dendrochronological analyses. Using electronic point dendrometers, we studied the climate response of stem radius changes at high temporal resolution (10-min intervals) in a mixed larch–birch forest in northern Mongolia in a drought year and two subsequent moist years. Larch trees showed stronger stem radius fluctuations than birches, and this difference was more pronounced in dry than in moist years. Stem radius changes were most tightly related to soil temperature variation, while soil moisture was the dominant controlling factor only in birch in the dry year. Correlations with the atmospheric vapor pressure deficit (VPD), and even more so with air temperature, were much weaker. While the linkage between radius change and VPD might primarily reflect diurnal transpiration-driven stem shrinkage and expansion, soil temperature is thought to directly affect cambial cell division and elongation during wood formation. We conclude that the phenology of stemwood increment is strongly controlled by soil temperature even in drought-limited southern boreal forests under continental climate due to the combination of cold and dry climate.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Drought limitation of tree growth and forest productivity has emerged as a key topic of forest health in large parts of the boreal forest (Buermann et al. 2014; Tei et al. 2017; Babst et al. 2019), which were limited in the past primarily by low summer temperatures and nitrogen deficiency alone (Dulamsuren and Hauck 2021). Therefore, climatic influences on tree growth have attracted increased attention in recent years in numerous dendrochronological studies (e.g., Dulamsuren et al. 2010a; b; Huang et al. 2010; Hellmann et al. 2016; Khansaritoreh et al. 2017). Climate response analyses of tree ring width reveal either correlation of increasing increment with elevated growing season temperatures in temperature-limited systems (Dulamsuren et al. 2014; Tumajer and Lehejček 2019) or with increasing precipitation and low summer temperatures if drought becomes limiting (Dulamsuren et al. 2013; Mamet et al. 2015). In some studies, a switch from temperature limitation to drought limitation was detected, if data from different time periods were analyzed (Lloyd and Bunn 2007).
While the significance of either low temperature limitation or drought limitation for annual stem increment is well established in tree growth analysis, much less is known about the climatic control of stem radius changes measured at high temporal resolution. From an annual growth perspective, cumulative stem radius changes as recorded by dendrometers reflect the annual stem increment in a similar manner as derived from tree-ring analysis. However, factors controlling intra-annual variation in stem radius must not necessarily be the same as variables limiting annual wood production. This is evident from the fact that stem radius changes recorded at high temporal resolution do not only reflect stemwood production, but also fluctuations in the water content of elastic stem tissues (Irvine and Grace 1997; Zweifel et al. 2000, 2001). Furthermore, in contrast to tree-ring width data, dendrometer readings also mirror the timing of cell division and cell elongation (Oberhuber et al. 2014).
Short-term variation in the water content of elastic and inelastic stem tissues beyond the water-conducting xylem elements is the result of changing radial water potential gradients. Falling xylem water potentials due to diurnal transpiration maxima at noon and afternoon during maximum evaporative demand cause reversible daily radial stem shrinkages and thus a diurnal stem radius periodicity (Dietrich et al. 2018). Drought events that reduce predawn and midday xylem water potentials over extended periods can reinforce radial potential gradients in the stem and cause stem shrinkage that lasts over several days or even weeks and is thought to reflect periods of tree water deficits (Zweifel et al. 2005; Oberhuber et al. 2015; Schäfer et al. 2019). Water flowing from bark, phloem, cambium, and xylem parenchyma tissues to xylem conduits due to a steeper radial water potential gradient during drought may help to maintain transpiration during periods of water deficit and postpone stomatal closure (Brough et al. 1986; Zweifel et al. 2001). Zweifel and Häsler (2001) quantified the contribution of radial water translocation in the stem to canopy transpiration with 2–5 l or 5% of the water loss in 13–28 m tall Picea abies trees. This contribution was less in a ca. 85 m tall Sequoiadendron giganteum tree, where 1.5–1.8% of the trees’ transpiration came from stem water storage (Williams et al. 2021).
Stem shrinkage may also be caused by low temperatures when local freezing of xylem sap reduces water vapor pressure and generates a radial water potential gradient between frozen and liquid water reservoirs (Zweifel and Häsler 2000; Améglio et al. 2001). In this context, it is important to mention that the xylem of exposed branches in the outer canopy can experience freezing even before frost occurs near the ground and that ice formation spreads downward through the xylem conduits from the crown to the tree base (Jordan and Smith 1994; Zweifel and Häsler 2000). Freezing can spread remarkably quickly along the water flow pathway in the xylem with a propagation velocity of several decimeters per minute (Single 1964; Levitt 1980). The tree trunk then starts to freeze from the xylem, while the surrounding cambial and phloem tissues with higher solute concentrations remain unfrozen. Frost-induced stem shrinkage can, therefore, occur when air temperatures at the standard measuring height of 2 m and soil temperatures are still above zero. In turn, stem expansion due to thawing in the xylem, which reverses frost-induced stem shrinkage, may still occur at air temperatures around 10 °C (Zweifel and Häsler 2000).
Even though study results are inconsistent (Irvine and Grace 1997), most authors agree that the xylem parenchyma plays a subordinate role in generating turgor-driven stem radius changes (Brough et al. 1986; Zweifel et al. 2000). Rather, most of the contraction is happening in the phloem and cambium. Additional water presumably originating from elastic and inelastic bark tissues may enter the xylem conduits through pits after having passed the phloem, cambium and wood rays through apoplastic and symplastic radial transport (Zweifel et al. 2000; Borchert and Pockman 2005; Treydte et al. 2021). In many trees, the bark functions as an important water reservoir with direct contact to the atmosphere, which does only store rainwater but also absorbs water vapor (Ilek et al. 2021).
The complex interaction of factors causing stem radius fluctuations suggests that the climatic variables controlling either annual stem increment or the short-term positive and negative radius fluctuations are likely to differ, because annual ring width reflects only the irreversible (positive) component of stem radius change, while additional reversible (positive and negative) components of radius fluctuations do exist. Under conditions of ample water supply, root temperature plays an important role not only for root growth but also for the increment of aboveground organs, as it affects water uptake and thus leaf water status and photosynthetic activity (Lyr and Garbe 1995; Lyr 1996; Solfjeld and Johnsen 2006; Alvarez-Uria and Körner 2007). Surprisingly, some studies from boreal and subalpine forests did not find a significant effect of soil temperature on radial growth (Deslauriers et al. 2003; Lupi et al. 2012; Oberhuber et al. 2014).
In the present study, we analyze stem radius changes recorded with electronic point dendrometers in the boreal forest at its southern fringe in Inner Asia, where climatic droughts usually limit annual stem increment, exacerbated through strong climate warming in recent decades (Dulamsuren et al. 2010a, 2011). Stem radius changes were comparatively analyzed in Larix sibirica (Siberian larch), a dominant late-successional boreal conifer in southern Siberia and Inner Asia, and Betula platyphylla (Siberian silver birch), a broadleaved pioneer tree with boreal-temperate distribution. We had the opportunity to monitor stem radius changes and stand microclimate in a year with a severe summer drought in the normally precipitation-richest month July, and in two subsequent moister years with marked precipitation peak in July, which is typical for the regional climate. In view of the pronounced drought limitation of annual stem increment in Inner Asia’s boreal forests at the edge to the steppe grassland, we tested the hypothesis (1) that air humidity and soil moisture are more closely related to stem radius changes than air and soil temperatures. Given that Mongolia’s boreal forest region remains a cold place with widespread subzero mean annual temperatures despite prevailing drought limitation of forest productivity (Dulamsuren et al. 2010a, b), we tested the hypothesis (2) that soil temperature is an additional factor that influences stem radial dynamics in moist years. Furthermore, we tested the hypothesis (3) that moisture effects on stem radius change are greater in birch than in larch, since the transpiration of early-successional broadleaf trees (such as birches and poplars) is known to follow more closely temporal variation in VPD than that of late-successional conifers with more stringent stomatal regulation (Ponton et al. 2006; Dulamsuren et al. 2009a, b; Zha et al. 2010).
Materials and methods
Study area
The study was conducted in Mongolia’s mountain taiga region at 48°40′ N, 106°55′ E in the western Khentey Mountains near the town of Tunkhel, ca. 85 km north of the Mongolian capital Ulan Bator and 230 km south of Lake Baikal. The Khentey Mountains range from Ulan Bator northeastward into Siberia (where they are named Yablonoviy Mountains) to Transbaikalia. Geologically, the mountain range consists of Proterozoic and Paleozoic rock, predominantly granite. Forests below 1500 m a.s.l. are mostly composed of Larix sibirica Ledeb., but are frequently replaced by Betula platyphylla Sukaczev after natural and anthropogenic disturbance, which is mainly related to fire, windthrow and logging (Dulamsuren 2004). A local timber industry was developed in Tunkhel, which is located 12 km from our study site, in the 1960s. Field work was carried out in the interior of a mixed L. sibirica-B. platyphylla forest on a north-facing mountain slope (48°40′5″ N, 106°55′12″ E) at an elevation of ca. 1400 m a.s.l. (Dulamsuren 2021).
Climate of the study region
To characterize the macroclimate and assess long-term climate trends of the study region in the Khentei Mountains, we retrieved monthly means of air temperature and precipitation sums from the CRU TS 4.04 dataset edited by the Climate Research Unit of the University of East Anglia, Norwich and the Met Office in Exeter, UK. All climatological data were downloaded from https://climexp.knmi.nl, selecting a resolution of 0.5° × 0.5° for the grid field of 48.5–49.0°N and 106.5–107.0°E. One-month values of the Standardized Precipitation-Evapo-transpiration Index (SPEI1; Vicente-Serrano et al. 2010) were calculated with the R package ‘SPEI’ version 1.7 using values of potential evapotranspiration, which were calculated after Thornthwaite (1948) based on the downloaded mean temperature values and a latitude of 48.67°N.
The climate of the study area is dominated by the Siberian High, which usually develops from September to April with its center over Lake Baikal, before it collapses. It then allows the inflow of warm and moist air masses, generating a marked precipitation peak in summer. Under high atmospheric pressure, the climate is extremely dry and cold in winter with mean January temperatures around −25 °C and minima as low as −45 to −50 °C, that are reached in most winters (Dulamsuren and Hauck 2008). For 1901 to 2019, mean annual (July/January) temperature of the study region was −1.7 ± 0.1 °C (17.6 ± 0.1 °C/−24.0 ± 0.3 °C). Mean annual precipitation in this period was 329 ± 5 mm with 90% received from May to September (297 ± 5 mm) and 68% (226 ± 4 mm) from June to August. Mean annual temperature has increased at a rate of 0.31 K decade−1 from −2.1 °C in 1960 to −0.3 °C in 2019, while mean annual precipitation has decreased by 10 mm decade−1 from 358 to 298 mm in the same interval.
Climatic conditions in the study years
The summers of 2017–2019 differed in their temperature and moisture regimes (Fig. 1). A marked summer drought spell occurred in late June and July 2017, whereas 2018 and 2019 were moist years with clear precipitation peaks in July. While July precipitation amounted to 46 mm in 2017, it was 119 mm in 2018 and 97 mm in 2019. The long-term average of July precipitation (1901–2019) was 85 mm. The one-month SPEI value (SPEI1) for July was −2.1 in 2017 as compared to 1.3 and 0.1 in 2018/19. The dry July 2017 was followed by a wet August (119 mm), whereas August precipitation sums in 2018 (90 mm) and 2019 (70 mm) were closer to the long-term average (1901–2019) for this month of 79 mm. While the annual precipitation in 2018 (354 mm) exceeded the long-term average of 329 mm, it was below average in 2017 (298 mm) and 2019 (290 mm). The similar annual precipitation amounts in 2017 and 2019 show that the main climatic drought element in 2017 was a dry July, while much of the precipitation deficit was compensated by higher rainfall in August. As evidenced from the long-term SPEI series for the study region, 2017 was among the driest years since 1901 (Fig. S1). Yet, despite the moist July, 2018 was similarly dry as 2017 according to mean SPEI and even reached the lowest minimum SPEI on record due to an extremely dry May (SPEI = −3.0).

Monthly values of a mean air temperature, b precipitation sums and c one-month values of the Standardized Precipitation-Evapotranspiration Index (SPEI1) from 2017 to 2019 modeled from the CRU TS 4.04 dataset edited by the Climate Research Unit of the University of East Anglia, Norwich and the Met Office in Exeter, UK (https://climexp.knmi.nl) at a resolution of 0.5° × 0.5° for the grid field of 48.5–49.0°N and 106.5–107.0°E. Months are indicated by numbers. MAT Mean annual temperature and AP annual precipitation sums are given in blue and precipitation sums for June/July in black type. July data are highlighted in red
Dendrometer recording
Electronic point dendrometers were installed on each five nearby birch and larch trees in the stand on the northern side of the tree boles to avoid direct insolation. Sample trees were selected among the dominant and subdominant trees to represent the typical range in diameter at breast height (1.3 m, dbh) in this stand and were spread over an area of ca. 50 m × 50 m. Since larch trees were, on average, taller than birch trees, birch trees of a mean dbh of 26 ± 2 cm and a mean height of 19 ± 1 m and larch trees with a dbh of 40 ± 2 cm and a height of 26 ± 2 m were selected (Table 1). Dendrometer readings were originally planned for the growing seasons 2017 and 2018, but were extended to 2019, after the weather conditions in 2017 and 2018 turned out to be highly contrasting. Dendrometer recording took place from May to October in 2017, April to October in 2018, and April to late September in 2019, thereby covering the entire growing season in the study region. In 2017, data recording was interrupted from June 12th to July 10th due to technical failure, i.e., a software error in the routine controlling data recording that did not affect the data quality in the remaining periods. All April and May data points of the three years were omitted from further analysis to exclude potential artifacts in the dendrometer recordings caused by dendrometer installation in spring 2017 and frost effects on sensor readings in the early growing season such as stem expansion with thawing, which can hardly be separated from the onset of initial growth (Zweifel and Häsler 2000; Tardif et al. 2001).
Sensors used as point dendrometers were RS Pro conductive polymer potentiometers (RS Components, Frankfurt/Main, Germany), which were originally developed for automotive applications. The potentiometers had a shaft of 3 mm diameter and 10 mm length. The devices were fixed to the stem with two parallel screws 34 mm apart and driven 80 mm into the wood, thus leaving full flexibility for measuring short-time radial shrinking as well as growth movements of the stem. The dead bark was removed before sensor mounting. The sensor thus recorded radius variations in the phloem, cambium and xylem. Phloem depth was 0.82 ± 0.08 mm in birch, but much thinner in larch. To avoid penetration of the spring-loaded sensor into the wood, a plate of stainless steel with 10 mm diameter was attached to the tip of the measuring shaft. The raw sensor signal of each sensor (resistance measured in Ω) was calibrated with a high-precision electronic sliding caliper. Sensors were connected to a CR1000 data logger via an AM16/32 multiplexer (Campbell Scientific, Logan, Utah, USA). The sensor signal was measured every 10 s and stored as 10-min averages. Annual radial increment was calculated from annual maximum values of the dendrometer readings. The calculation of tree water deficits (TWD) followed the zero growth concept of Zweifel et al. (2016). This concept assumes that no growth takes places during TWD, and thus TWD at a given point in time t is calculated as the maximum stem radius (SRmax) before stem shrinkage minus the stem radius at time t (SRt). The end of a TWD phase is reached when SRt reaches (or exceeds) SRmax again. Transient reductions of the stem radius lasting for less than 24 h, which result from diurnal variation in transpiration, were excluded from the analysis.
Field measurements of microclimate and soil moisture
Air temperature (Ta) and relative air humidity were recorded hourly from 2017 to 2019 using HOBO U23 ProV2 temperature/relative humidity data loggers (Onset Computer Cooperation, Bourne, Massachusetts, USA). These loggers were placed at 1.5 m height on the north side of five birch and five larch trunks, which were selected at random positions in the stand. Data were used to calculate VPD following Buck (1981). The results showed that Ta, relative air humidity and the dependent VPD were not influenced by the tree species (linear regression slopes for values from birch vs. larch were 1; r = 1.00, P < 0.001). Therefore, mean values across all 10 sample trees are presented.
Soil temperature and soil moisture were measured at one central location in the sample plot employing the same CR1000 Data Logger as for the dendrometers. Soil temperature (Ta) was recorded at 1, 5, 10, 20, 30, 40, 50, and 60 cm depth. Volumetric soil water content (SWC) and soil matric potential (SMP) were measured every 2 h at 15, 30, and 50 cm depth. Volumetric water content was recorded with horizontally installed CS616 time-domain reflectometry probes (Campbell Scientific, Logan, Utah, USA). The raw signal, i.e., travel time t (in µs) of the voltage pulse along the 30 cm-long sensors, was converted to SWC (in m3 m−3) with a quadratic equation and coefficients according to the manufacturer’s manual: SWC = –0.0663 – 0.0663 t + 0.0007 t2.
For recording SMP, ceramic heat dissipation sensors (Model 229, Campbell Scientific) were chosen for their wide measuring range, especially under dry conditions (−10 to −2500 kPa). This sensor type has been in extensive use in soil water monitoring networks in the USA since the 1990s (Schneider et al. 2003) and has, thus, been subject to constant theoretical improvement (Flint et al. 2002). Raw values, i.e., the temperature rise after a 30 s-heat pulse inside the ceramic, were converted into SMP by a logistic equation:
with empirical parameters fitted by Zhang et al. (2019), including the maximum matric potential measurable with this equation (|SMP|max, 2083 kPa), the steepness of the curve a (3.35 °C−1), and the inflection point \(\Delta {T}_{\mathrm{inf}}\) (3.17 °C). \({\Delta T}_{\mathrm{ref}}\) is the normalized reference temperature difference, calculated according to Schneider et al. (2003) using the temperature difference measured in the field and sensor-specific temperature difference under wet and dry conditions (measured for each sensor in the laboratory). Where logarithmic values were advantageous in diagrams, the pF value was used to characterize SMP, which was derived from SMP (in hPa) according to pF = log (−SMP).
Statistical analysis
Arithmetic means ± standard errors (SE) are presented throughout the paper, except for climate data, where the specification of standard deviations (SD) seemed more appropriate. Statistical testing was done with SAS 9.4 software. Data were tested for normal distribution with the Shapiro–Wilk test. Four-way analysis of variance (ANOVA) was calculated to quantify the effect of climatic and edaphic variables (soil temperature, SWC) on stem radius changes. Soil temperatures at 60 cm and SWC at 15 cm were selected to represent soil thermal and moisture conditions in the ANOVA, as the correlation of soil temperature increased, but that with SWC decreased, with increasing soil depth, and soil temperature and SWC from different depth levels were inter-correlated. SWC was preferred over SMP in the model, as the former was more closely correlated with stem radius change. Linear correlations were examined with Pearson’s correlation coefficient. ANOVA and correlation analyses were calculated for the summer months (June–August) to exclude potential frost influences in the late growing season, which are not related to growth, but can generate stem shrinkages.
Results
Microclimate in the studied forest
According to our microclimate measurements in the study plot, the summer of 2017 was drier than those of 2018 and 2019 (Table 2; Fig. 2). As the 2017 drought episode occurred in late June and July, these contrasts become especially evident from a comparison of Ta July means (19.8 vs. 15.6 and 17.8 °C in 2017, 2018 and 2019) and VPD (1.27 vs. 0.43 and 0.66 kPa). The onset of the 2017 summer drought was marked by a gradual decline of SMP in the sample plot from early June (June 3rd), when soil moisture reached field capacity (−10 kPa; Fig. S1), to a minimum in late July (July 25th; Fig. 3a). In these 7.5 weeks, SMP fell to minima of −415 kPa at 15 cm, −133 kPa at 30 cm, and −183 kPa at 50 cm soil depth (Fig. 3a). One month later (on August 20th), the soil had re-saturated to field capacity at all surveilled depths (Fig. S2). This high soil moisture level was kept until the end of the growing season. In summer 2018, SMP never fell below −55 kPa at any depth and reached a growing season mean (early May to late September) of −9.5 kPa, which is close to field capacity (Fig. 3b). The low SMP in summer 2017 was associated with low SWC. Yet, the July minimum of SMP had a larger influence on the growing season mean than the SWC minimum, causing greater SWC differences between the years than in SMP (Table 2). Growing season Ts means did not reflect inter-annual Ta differences and were not affected by soil moisture differences between the years at any depth from 1 to 60 cm (Table 2).

a Air temperature (Ta) and b VPD between May 2017 and July 2019 in the studied forest (mean values from 10 temperature/relative humidity sensors). Months are indicated by numbers

Seasonal course of soil matric potential (SMP) at 15, 30, and 50 cm depth in the growing seasons of a the dry year 2017 and b the moist year 2018
Weak frost occurred sporadically in the late growing season. Ta fell below zero on a single day in late August 2017 (minimum −0.17 °C) and on 3 days in mid/late September 2018 (−1.6 to −1.8 °C). Subzero air temperatures were not measured in the observation period of 2019. Ground frost did not occur in late summer/early fall. The deep soil at 60 cm was still weakly frozen (−0.02 °C) at the start of the observation period in the third year on June 1, 2019.
Ta ranges during the summer months (June–August) covered a span from weak frost to 32 °C (Table S1). Minimum Ts in the profile to 60 cm depth varied between 1.7 and 4.4 °C in 2017 (June to August), between 1.4 and 5.6 °C in 2018, and between 0.0 and 3.5 °C in 2019 (Table S1). The corresponding Ts maxima ranged from 8.8–19.0 °C in 2017, 9.6–18.2 °C in 2018, and 8.7–17.4 °C in 2019.
Stem radius change
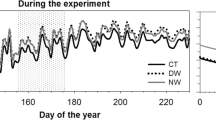
In both species, the radial stem increment in the drought year 2017 was lower than in the moist years 2018 and 2019 (Fig. 4). Stem increment generally continued until July. From smoothened curves where most of the diurnal variation was removed by a cubic spline with a 90% frequency–response cut-off, it becomes evident that stem radius changes generally showed a stronger high-frequency variation in the drought year than in the moist years, and also in larch as compared to birch. At the end of the growing season, the curves of stem radius change showed a downward curvature, except for larch in 2019.

Cumulative radial stem increment of a L. sibirica (N = 5) and b B. platyphylla (N = 5) in the growing seasons (June 1st–September 20th) of the dry year 2017 and the moist years 2018/19. Curves were smoothened to remove high-frequency variability using cubic spline functions with a 90% frequency–response cut-off. Missing data from June 12 to July 10, 2017 due to technical failure
Correlation of stem radius change with climatic and edaphic parameters
Stem radius changes in the summer months exhibited a much stronger relationship to soil temperature than to atmospheric climatic variability (Table 3). Across tree species and years, soil temperature emerged as environmental variable with closest association with stem radius change. In the drought year 2017, however, SWC showed in birch a stronger effect than temperature (Table 3). In this year and the following year, the interaction of Ts and SWC also exerted a large effect on stem radius variability in the ANOVA. Overall, the ANOVA models composed of VPD, SWC, Ta, and Ts could mostly explain as much as 90–100% of total stem radius variability. For larch, R2 was lower in the drought year 2017 (64%) and in the following year (81%). Correlations of stem radius change with Ts became stronger with increasing soil depth (Fig. 5a). They were particularly strong in the moist years 2018/19, where Pearson coefficients varied between 0.87 and 0.99 at soil depths from 10 to 60 cm. This close relationship was found for both larch and birch. In the dry year 2017, larch showed lower, but nevertheless relatively strong correlations with soil temperature at 10–60 cm depth with r ranging from 0.52 to 0.83. By contrast, birch revealed only a weak relationship with soil temperature in the drought year 2017; r varied between 0.10 and 0.47 at 10–60 cm depth in that year. Near the surface at 1 and 5 cm depth, Ts was not related to stem radius change in the drought year (r between −0.23 and 0.30). In the moist years 2018/19, correlations were weak for Ta at 1 cm depth (r between 0.52 and 0.58) and relatively strong at 5 cm depth (r between 0.71 and 0.81).

Linear correlations of high-resolution (30 min) stem radius change in L. sibirica and B. platyphylla with a soil temperature (Ts), b SWC soil water content, and c soil matric potential (SMP) at varying soil depths from 1 to 60 cm during summer (June to August) in the dry year 2017 and the moist years 2018/19. All correlations were significant at P ≤ 0.001 (Pearson correlations)
Correlations of SWC with stem radius change decreased with soil depth, except for larch in the drought year 2017, where the correlation was very weak with r ranging from 0.16 to 0.21 (Fig. 5b). Close correlation with SMP was only found for birch in the drought year (r between −0.81 and −0.87). High VPD was negatively correlated with stem radius changes in both species in all years (Fig. 6), though these correlations were not as strong as with Ts. Rather weak negative correlations occurred with Ta in the drought year, whereas Ta had no effect on stem radius change in the moist years (Fig. 6).

Linear correlations of stem radius change in L. sibirica and B. platyphylla with air temperature (Tair) and atmospheric VPD vapor pressure deficit during summer (June to August) in the dry year 2017 and the moist years 2018/19. Levels of significance: **P ≤ 0.01; n.s. not significant; otherwise P ≤ 0.001 (Pearson correlations)
Tree water deficits
Both tree species showed stem shrinkages beyond the diurnal variation throughout the growing season (Fig. 7). In most cases, stem shrinkage proceeded in parallel in the two species, suggesting that they are attributable to common climatic drivers. In addition to transient stem shrinkages during peak growing season, longer lasting trends of decreasing stem radius occurred at varying points of time in the late growing season. Since they became dominant in September (with downward trend of most stem radius curves; Fig. 4), the September data have been omitted from the above climate response analyses. These stem shrinkages at the end of the growing season were at least partly associated with mild frost events. In 2017, late growing season stem shrinkages started in late August and continued (despite transient ups and downs) until the end of the growing season in late September (Fig. 7a). In 2018, such stem shrinkage started in September, later than in 2017 (Fig. 7b). In 2019, steep trends of stem shrinkage did not occur in larch and were in birch less clearly developed than in the other years (Fig. 7c).

Tree water deficit in birch (N = 5) and larch (N = 5) from July 11th to September 20th in a the drought year 2017 and b, c the moist years 2018 and 2019
Discussion
Soil temperature was found to exert the strongest effect on stem radius changes measured at high temporal resolution, when relationships to atmospheric (Ta) and soil (Ts) temperatures, VPD, and soil moisture were analyzed. This is unexpected, since the boreal forests in southern Siberia and Inner Asia are generally known for the strong drought limitation of growth due to their position close to the Central Asian steppe grassland biome (Dulamsuren et al. 2013; Liu et al. 2013; Khansaritoreh et al. 2017). This climatic drought limitation explicitly includes the studied forest in the western Khentei Mountains, where recent climate change has aggravated drought limitation since the late twentieth century (Dulamsuren et al. 2010a, b, 2011). Yet, water availability was the most important explanatory factor of radius changes only in the drought year 2017 and solely influenced birch but not larch. Thus, our first hypothesis assuming a consistent dominant influence of water availability has to be rejected. The dominant impact of Ts on stem radius change was all the more unexpected, as it is inconsistent with the results of similar studies from boreal forests with Abies balsamea and Picea mariana in eastern Canada (Deslauriers et al. 2003; Lupi et al. 2012) and subalpine conifer stands of the Austrian Alps (among others Larix decidua, Oberhuber et al. 2014). The authors of these studies agreed in assuming a subordinate role of Ts on stem radius changes, even though water availability was more favorable in those study regions than in our study area in Mongolia. However, our results from Mongolia parallel findings from Picea crassifolia forests at the north-eastern edge of the Tibetan Plateau, which are, like Mongolia’s boreal forest region, characterized by the combination of drought and low temperatures (Tian et al. 2017; Wan et al. 2022). Nevertheless, it cannot be ruled out that drought might become more critical under future drier and warmer conditions.
The importance of Ts for stem radius variation was clearly greater in the moist years 2018/19 than in the drought year 2017, thus supporting our second hypothesis. The existence of a lower threshold for water supply, at which Ts becomes relevant, is also evident from the significant interaction of Ts with soil moisture in the ANOVA (Table 3) and from the uncoupling of stem radius change from Ts at the sun-exposed and thus drought-prone soil surface. In the drought year 2017, soil moisture displaced Ts as the most dominant independent variable for birch, but not larch, which is consistent with our third hypothesis that birch depends more on the variability of water supply due to its less sensitive stomatal regulation than late-successional larch with its more stringent stomatal control. However, this was only valid during drought, and the dominant relationship of Ts with stem radius variation was observed in both tree species.
Relevant experimental studies, which examined the impact of low root temperatures on growth, photosynthesis and water relations of trees were mostly carried out with seedlings in hydroponic culture and thus under conditions of ample water supply (Vapaavuori et al. 1992; Lyr and Garbe 1995; Lyr 1996; Solfjeld and Johnsen 2006). Under such conditions, cooling of the root systems resulted in reduced root and shoot growth and caused only transient reductions in stomatal conductance and transpiration, but permanently reduced the photosynthetic carbon gain as long as low temperatures were effective (Lyr and Garbe 1995). The reduction in net photosynthesis likely is attributable to the cold inhibition of cell division and cell elongation, which is mediated by the downregulation of cytokinin and gibberellin synthesis at low temperatures, and subsequently leads to the accumulation of non-structural carbohydrates (NSC) (Lyr and Garbe 1995; Eremina et al. 2016). NSC accumulation can lead to the downregulation of photosynthesis despite high stomatal conductance (Neubauer et al. 2011; Sugiura et al. 2017) and thus may uncouple transpirative water loss from carbon assimilation. In addition, water uptake can be impaired by increased viscosity of water and reduced membrane permeability at low temperatures in the soil-root interface (Aroca et al. 2012).
Experiments in hydroponic culture showed reductions in shoot growth at root zone temperatures between 2 and ca. 10 °C in temperate and boreal tree species (Vapaavuori et al. 1992; Lyr and Garbe 1995; Schenker et al. 2014). These experiments included Betula pendula, which is closely related to B. platyphylla, and Larix decidua (Lyr 1996; Solfjeld and Johnsen 2006). Lyr 1996 found a temperature optimum for whole-plant biomass increment at a root zone temperature of 15 °C for both L. decidua and B. pendula. Soil temperatures in Mongolia remained during summer (June–August) regularly below this optimum temperature with a mean temperature range from 5.6 to 9.0 °C recorded at 20–60 cm soil depth and of 9.1 to 10.0 °C at 10 cm depth, while temperature minima varied between 0.0 and 4.6 °C during summer at 10–60 cm depth. In soil profiles to 80 cm depth in Mongolia, L. sibirica showed no significant decrease in root density with soil depth (Chenlemuge et al. 2013), suggesting that Ts measured at 60 cm is highly relevant for larch growth. The explanatory powers of the ANOVAs with most R2 values for the entire model reaching values of 0.9 to 1.0, combined with the high F values found for Ts (Table 3), strongly suggest that the correlations between stem radius change and Ts are causal. The strong linear correlation with Pearson coefficients between 0.87 and 0.99 (at 10–60 cm depth) in the moist years (Fig. 5a) also suggests a direct impact of low temperature on stem radius dynamics. Our results are consistent with finding from Picea crassifolia forests on the north-eastern Tibetan Plateau, where Tian et al. (2017) found a significant impact of Ts on dendrometer values. These spruce forests are drought-limited ecosystems (Liang et al. 2016) in a similar manner as the larch–birch forests studied in Mongolia. This coincidence suggests that Ts is a key determinant of the phenology of wood formation in these dry and cold forest ecosystems, even though drought is limiting annual wood production. Ts has to be regarded independently of Ta, as inter-annual differences in Ts did not closely follow the variation of Ta. Among others, this is attributable to the decoupling of annual mean values of Ts and Ta by the long-lasting snowpack in Mongolia’s boreal forests (Grundstein et al. 2005).
The much weaker, but consistently observed, negative correlation between stem radius change and VPD is most likely caused by diurnal stem shrinkage due to low xylem pressures during maximum transpiration at noon and in the afternoon (Intrigliolo and Castel 2006; Dulamsuren et al. 2009a; Turcotte et al. 2011). The detected correlation between stem radius change and VPD thus most likely does not reflect a climate effect on radial growth, but rather is the consequence of the diurnal periodicity of stem radius changes caused by radial water potential gradients and the resulting water flow from peripheral stem tissues toward the xylem (Zweifel et al. 2000; Treydte et al. 2021).
Long-lasting stem shrinkage, as observed late in the growing season at the end of August and in September, was probably induced by frost (Zweifel and Häsler 2000; Dong et al. 2019), as the onset of these events coincided with the occurrence of weak frost. The flow of water from the bark, phloem and cambium to frozen xylem conduits may influence the frost resistance of these tissues, as they lose water, and thus should become more resistant to freezing (Pramsohler and Neuner 2013). The exact mechanisms underlying the observed persistent trends for late-growing-season stem shrinkage remain to be studied (Zweifel and Häsler 2000). Even though the beginning of stem shrinkage started in parallel with the onset of weak frost events, especially the weak frost on August 28, 2017 was not of long duration, but lasted for just a day. Yet, stem radius did not return to its original value in that growing season. Even though late-growing-season stem shrinkage may have little impact on annual radial growth and apparently does not influence the climate–growth relationships detected in the frost-free growing season, this phenomenon deserves further study to unravel its importance for boreal forests exposed to both winter cold and summer drought.
Conclusions
Our results obtained with electronic point dendrometers indicate that the phenology of stemwood formation in southern boreal forests of Mongolia is primarily controlled by soil temperatures (Ts), even though annual stem increment is primarily limited by drought. This result differs from findings made in more oceanic boreal climates (Deslauriers et al. 2003; Lupo et al. 2012), but is consistent with data from oroboreal spruce mountain forests of the north-eastern Tibetan Plateau. Since both Mongolia’s boreal forest zone and the north-eastern Tibetan Plateau share cold continental climatic conditions with low Ts even during summer, low summer precipitation and high potential evapotranspiration (Wagner et al. 2015), it is plausible to assume that co-limitation of wood formation by low soil temperatures might be widespread in such dry and cold climates. Dulamsuren and Hauck (2021) recently showed that also co-limitation by nitrogen is effective in drought-limited southern boreal forests of Mongolia. Taken together, these results suggest that while drought is the key factor, controlling stemwood formation and thus forest productivity at the southern fringe of the boreal forest in Inner Asia, Ts and nitrogen availability are both important co-limiting factors in these forests. It should be clarified how widely distributed such co-limitation by Ts and nitrogen occurs in primarily drought-limited boreal forests, because drought limitation of forest productivity is generally increasing in the boreal forest biome as the result of climate change (Buermann et al. 2014).
Data availability
Data can be requested from the authors.
References
Alvarez-Uria P, Körner C (2007) Low temperature limits of root growth in deciduous and evergreen temperate tree species. Funct Ecol 21:211–218
Améglio T, Cochard H, Ewers FW (2001) Stem diameter variations and cold hardiness in walnut trees. J Exp Bot 52:2135–2142
Aroca R, Porcel R, Ruiz-Lozano JM (2012) Regulation of root water uptake under abiotic stress conditions. J Exp Bot 63:43–57
Babst F, Bouriaud O, Poulter B, Trouet V, Girardin MP, Frank DC (2019) Twentieth century redistribution in climatic drivers of global tree growth. Sci Adv. https://doi.org/10.1126/sciadv.aat4313
Borchert R, Pockman WT (2005) Water storage capacitance and xylem tension in isolated branches of temperate and tropical trees. Tree Physiol 25:457–466
Brough DW, Jones HG, Grace J (1986) Diurnal changes in water content of stems of apple trees, as influenced by irrigation. Plant Cell Environ 9:1–7
Buck AL (1981) New equations for computing vapor pressure and enhancement factor. J Appl Meteorol 20:1527–1532
Buermann W, Parida BR, Jung M, MacDonald GM, Tucker CJ, Reichstein M (2014) Recent shift in Eurasian boreal forest greening response may be associated with warmer and drier summers. Geophys Res Lett 41:1995–2002
Chenlemuge T, Hertel H, Dulamsuren Ch, Khishigjargal M, Leuschner C, Hauck M (2013) Extremely low fine root biomass in Larix sibirica forests at the southern drought limit of the boreal forest. Flora 208:488–496
Deslauriers A, Morin H, Urbinati C, Carrer M (2003) Daily weather response of balsam fir (Abies balsamea (L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada). Trees 17:477–484
Dong M, Wang B, Jiang Y, Ding X (2019) Environmental controls of diurnal and seasonal variations in the stem radius of Platycladus orientalis in northern China. Forests 10:784
Dulamsuren Ch (2004) Floristische Diversität, Vegetation und Standortbedingungen in der Gebirgstaiga des Westkhentey, Nordmongolei. Ber Forschungszentr Waldökosyst A 191:1–290
Dulamsuren Ch (2021) Organic carbon stock losses by disturbance: comparing broadleaved and late-successional conifer forests in Mongolia’s boreal forest. For Ecol Manag 499:119636
Dulamsuren Ch, Hauck M (2008) Spatial and seasonal variation of climate on steppe slopes of the northern Mongolian mountain taiga. Grassland Sci 54:217–230
Dulamsuren Ch, Hauck M (2021) Drought stress mitigation by nitrogen in boreal forests inferred from stable isotopes. Glob Change Biol 27:5211–5224
Dulamsuren Ch, Hauck M, Bader M, Osokhjargal D, Oyungerel Sh, Nyambayar S, Runge M, Leuschner C (2009a) Water relations and photosynthetic performance in Larix sibirica growing in the forest-steppe ecotone of northern Mongolia. Tree Physiol 29:99–110
Dulamsuren Ch, Hauck M, Bader M, Osokhjargal D, Oyungerel Sh, Nyambayar S, Leuschner C (2009b) The different strategies of Pinus sylvestris and Larix sibirica to deal with summer drought in a northern Mongolian forest-steppe ecotone suggest a future superiority of pine in a warming climate. Can J for Res 39:2520–2528
Dulamsuren Ch, Hauck M, Leuschner C (2010a) Recent drought stress leads to growth reductions in Larix sibirica in the western Khentey, Mongolia. Glob Change Biol 16:3024–3035
Dulamsuren Ch, Hauck M, Khishigjargal M, Leuschner HH, Leuschner C (2010b) Diverging climate trends in Mongolian taiga forests influence growth and regeneration of Larix sibirica. Oecologia 163:1091–1102
Dulamsuren Ch, Hauck M, Leuschner HH, Leuschner C (2011) Climate response of tree-ring width in Larix sibirica growing in the drought-stressed forest-steppe ecotone of northern Mongolia. Ann For Sci 68:275–282
Dulamsuren Ch, Wommelsdorf T, Zhao F, Xue Y, Zhumadilov BZ, Leuschner C, Hauck M (2013) Increased summer temperatures reduce the growth and regeneration of Larix sibirica in southern boreal forests of eastern Kazakhstan. Ecosystems 16:1536–1549
Eremina M, Rozhon W, Poppenberger B (2016) Hormonal control of cold stress responses in plants. Cell Mol Life Sci 73:797–810
Flint AL, Campbell GS, Ellett KM, Calissendorff C (2002) Calibration and temperature correction of heat dissipation matric potential sensors. Soil Sci Soc Am J 66:1439–1445
Grundstein A, Todhunter P, Mote T (2005) Snowpack control over the thermal offset of air and soil temperatures in eastern North Dakota. Geophys Res Lett 32:L08503
Hellmann L, Agafonov L, Ljungqvist FC, Churakova O, Düthorn E, Esper J, Hülsmann L, Kirdyanov AV, Moiseev P, Myglan VS, Nikolaev AN, Reinig F, Schweingruber FH, Solomina O, Tegel W, Büntgen U (2016) Diverse growth trends and climate responses across Eurasia’s boreal forest. Environ Res Lett 11:074021
Huang J, Tardif JC, Bergeron Y, Denneler B, Berninger F, Girardin MP (2010) Radial growth response of four dominant boreal tree species to climate along a latitudinal gradient in the eastern Canadian boreal forest. Glob Change Biol 16:711–731
Ilek A, Siegert CM, Wade A (2021) Hygroscopic contributions of bark water storage and controls exerted by internal bark structure over water vapor absorption. Trees 35:831–843
Intrigliolo DS, Castel JR (2006) Usefulness of diurnal trunk shrinkage as a water stress indicator in plum trees. Tree Physiol 26:303–311
Irvine J, Grace J (1997) Continuous measurements of water tensions in the xylem of trees based on the elastic properties of wood. Planta 202:455–461
Jordan DN, Smith WK (1994) Energy balance analysis of nighttime leaf temperatures and frost formation in a subalpine environment. Agric for Meteor 71:359–372
Khansaritoreh E, Dulamsuren Ch, Klinge M, Ariunbaatar T, Bat-Enerel B, Batsaikhan G, Ganbaatar K, Saindovdon D, Yeruult Y, Tsogtbaatar J, Tuya D, Leuschner Ch, Hauck M (2017) Higher climate warming sensitivity of Siberian larch in small than large forest islands in the fragmented Mongolian forest steppe. Glob Change Biol 23:3675–3689
Levitt J (1980) Responses of plants to environmental stresses. Chilling, freezing, and high temperature stresses. Elsevier Academic Press, New York
Liang EY, Leuschner C, Dulamsuren Ch, Wagner B, Hauck M (2016) Global warming-related tree growth decline and mortality on the north-eastern Tibetan Plateau. Clim Change 134:163–176
Liu H, Williams AP, Allen CD, Guo D, Wu X, Anenkhonov OA, Liang EY, Sandanov DV, Yin Y, Qi Z, Badmaeva NK (2013) Rapid warming accelerates tree growth decline in semi-arid forests of Inner Asia. Glob Change Biol 19:2500–2510
Lloyd AH, Bunn AG (2007) Responses of the circumpolar boreal forest to 20th century climate variability. Environ Res Lett 2:045013
Lupi C, Morin H, Deslauriers A, Rossi S (2012) Xylogenesis in black spruce: does soil temperature matter? Tree Physiol 32:74–82
Lyr H (1996) Effect of the root temperature on growth parameters of various European tree species. Ann For Sci 53:317–323
Lyr H, Garbe V (1995) Influence of root temperature on growth of Pinus sylvestris, Fagus sylvatica, Tilia cordata and Quercus robur. Trees 9:220–223
Mamet SD, Chun KP, Metsaranta JM, Barr AG, Johnstone JF (2015) Tree rings provide early warning signals of jack pine mortality across a moisture gradient in the southern boreal forest. Environ Res Lett 10:084021
Neubauer SG, Renau-Morata B, Guardiola JL, Molina R-V (2011) Photosynthesis down-regulation precedes carbohydrate accumulation under sink limitation in Citrus. Tree Physiol 31:169–177
Oberhuber W, Gruber A, Kofler W, Swidrak I (2014) Radial stem growth in response to microclimate and soil moisture in a drought-prone mixed coniferous forest at an inner Alpine site. Eur J For Res 133:467–479
Oberhuber W, Kofler W, Schuster R, Wieser G (2015) Environmental effects on stem water deficit in co-occurring conifers exposed to soil dry ness. Int J Biometeorol 59:417–426
Ponton S, Flanagan LB, Alstad KP, Johnson BG, Morgenstern K, Kljun N, Black TA, Barr AG (2006) Comparison of ecosystem water-use efficiency among Douglas-fir forest, aspen forest and grassland using eddy covariance and carbon isotope techniques. Glob Change Biol 12:294–310
Pramsohler M, Neuner G (2013) Dehydration and osmotic adjustment in apple stem tissue during winter as it relates to the frost resistance of buds. Tree Physiol 33:807–816
Schäfer C, Rötzer T, Thurm EA, Biber P, Kallenbach C, Pretzsch H (2019) Growth and tree water deficit of mixed Norway spruce and European beech at different heights in a tree and under heavy drought. Forests 10:577
Schenker G, Lenz A, Körner C, Hoch G (2014) Physiological minimum temperatures for root growth in seven common European broad-leaved tree species. Tree Physiol 34:302–313
Schneider JM, Fisher DK, Elliott RL, Brown GO, Bahrmann CP (2003) Spatiotemporal variations in soil water: First results from the ARM SGP CART network. J Hydrometeorol 4:106–120
Single WV (1964) Studies on frost injury to wheat. II. Ice formation within the plant. Aust J Agric Res 15:869–875
Solfjeld I, Johnsen Ø (2006) The influence of root-zone temperature on growth of Betula pendula Roth. Trees 20:320–328
Sugiura D, Watanabe CKA, Betsuyaku E, Terashima I (2017) Sink-source balance and down-regulation of photosynthesis in Raphanus sativus: effects of grafting, N and CO2. Plant Cell Physiol 58:2043–2056
Tardif J, Flannigan M, Bergeron Y (2001) An analysis of the daily radial activity of 7 boreal tree species, northwestern Québec. Environ Monit Assess 67:141–160
Tei S, Sugimoto A, Yonenobu H, Matsuura Y, Osawa A, Sato H, Fujinuma J, Maximov TC (2017) Tree-ring analysis and modelling approaches yield contrary response of circumboreal forest productivity to climate change. Glob Change Biol 23:5179–5188
Tian Q, He Z, Xiao S, Peng X, Ding A, Lin P (2017) Response of stem radial growth of Qinghai spruce (Picea crassifolia) to environmental factors in the Qilian Mountains of China. Dendrochronologia 44:76–83
Treydte K, Lehmann MM, Wyczesany T, Pfautsch S (2021) Radial and axial water movement in adult trees recorded by stable isotope tracing. Tree Physiol 41:2248–2261
Tumajer J, Lehejček J (2019) Boreal tree-rings are influenced by temperature up to two years prior to their formation: a trade-off between growth and reproduction. Environ Res Lett 14:124024
Turcotte A, Rossi S, Deslauriers A, Krause C, Morin H (2011) Dynamics of depletion and replenishment of water storage in stem and roots of black spruce measured by dendrometers. Front Plant Sci 2:21
Vapaavuori EM, Rikala R, Ryyppö A (1992) Effects of root temperature on growth and photosynthesis in conifer seedlings during shoot elongation. Tree Physiol 10:217–230
Vicente-Serrano SM, Beguería S, López-Moreno JI (2010) A multiscalar drought index sensitive to global warming: the Standardized Precipitation Evapotranspiration Index. J Clim 23:1696–1718
Wagner B, Liang EY, Li X, Dulamsuren Ch, Leuschner C, Hauck M (2015) Carbon pools of semi-arid Picea crassifolia forests in the Qilian Mountains (north-eastern Tibetan Plateau). For Ecol Manag 343:136–143
Wan Y, Yu P, Li X, Wang X, Wang B, Yu Y, Zhang L, Liu X, Wang S (2022) Divergent seasonal patterns of Qinghai spruce growth with elevation in northwestern China. Forests 13:388
Williams CB, Næsborg RR, Ambrose AR, Baxter WL, Koch GW, Dawson TE (2021) The dynamics of stem water storage in the tops of Earth’s largests trees – Sequioadendron giganteum. Tree Physiol 41:2262–2278
Zha T, Barr AG, van der Kamp G, Black TA, McCaughey JH, Flanagan LB (2010) Interannual variation of evapotranspiration from forest and grassland ecosystems in western Canada in relation to drought. Agric For Meteorol 150:1476–1484
Zhang Y, Ochsner TE, Fiebrich CA, Illston BG (2019) Recalibration of sensors in one of the world’s longest running automated soil moisture monitoring networks. Soil Sci Soc Am J 83:1003–1011
Zweifel R, Häsler R (2000) Frost-induced reversible shrinkage of bark of mature subalpine conifers. Agric For Meteorol 102:213–222
Zweifel R, Häsler R (2001) Dynamics of water storage in mature subalpine Picea abies: temporal and spatial patterns of change in stem radius. Tree Physiol 21:561–569
Zweifel R, Item H, Häsler R (2000) Stem radius changes and their relation to stored water in stems of young Norway spruce. Trees 15:50–57
Zweifel R, Item H, Häsler R (2001) Link between diurnal stem radius changes and tree water relations. Tree Physiol 21:869–877
Zweifel R, Zimmermann L, Newberry DM (2005) Modeling tree water deficit from microclimate: an approach to quantifying drought stress. Tree Physiol 25:147–156
Zweifel R, Haeni M, Buchmann N, Eugster W (2016) Are trees able to grow in periods of stem shrinkage? New Phytol 211:839–849
Acknowledgements
We are thankful to the German Corporation for International Cooperation (GIZ) and to K. Schmidt-Corsitto and M. Trockenbrodt in particular for enabling us to use their field station in Tunkhel. We thank T. Ariunbaatar, Kh. Ganbaatar, G. Byamba-Yondon, Yo. Yeruult and B. Bat-Enerel for their help with changing and loading the batteries of the data loggers in the field.
Funding
The study was supported by a grant of the German Science Foundation (DFG, Du 1145/4-1) to Ch. Dulamsuren. Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
CD designed the study. HC designed the dendrometers and the setup for measuring soil microclimate. CD, HC and MH conducted field work. CD and HC analyzed data. CD wrote the paper. All authors commented and worked on the paper.
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare no conflict of interest.
Additional information
Communicated by Eryuan Liang.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dulamsuren, C., Coners, H., Leuschner, C. et al. Climatic control of high-resolution stem radius changes in a drought-limited southern boreal forest. Trees 37, 797–810 (2023). https://doi.org/10.1007/s00468-022-02384-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-022-02384-z