
Abstract
Key message
Tree-ring growth of pollarded narrow-leaved ash trees in Central Spain reveals traditional management cessation in 1970 and property-specific management patterns.
Abstract
Tree pollarding was a dominant management strategy of European forests for centuries creating open agroforestry landscapes with important cultural and environmental values. This traditional practice has been widely abandoned in last decades with a subsequent impact in terms of biodiversity and cultural loss. Central Spain hosts the largest and best-preserved area of pollarded narrow-leaved ash (Fraxinus angustifolia Vahl.) woodlands in Europe. The main aim of this research is to obtain rigorous historical records of pollarding frequency to get adequate information for traditional ash management. We used dendrochronological techniques to evaluate temporal changes of pollarding frequency and rotation length. We analysed the stand level synchrony and the effect of land property on pollarding activity from 322 trees growing in eight pollard stands in Central Spain. Pollarding events were unequivocally identified at tree level by a characteristic change in growth pattern. We identified 2426 tree-level pruning events with the first event dated in 1777. Historical pruning recurrence ranged between 5 and 10 years with higher pollarding frequency on private lands. Pruning events within each site were synchronous, suggesting the existence of a rotational schema within each stand. Pruning frequency decreased drastically in the 1970s matching with the depopulation of rural areas and the general abandonment of traditional practices. Pollarding practices have recovered in recent decades although with lower intensity and lacking the synchronic historical patterns. Providing technical and economic support to make this traditional activity profitably would have strong environmental revenue due the multiple ecosystem services provided by pollarded ashes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Tree pollarding has been one of the dominant management strategy of many European forests for centuries (Petit and Watkins 2003; Read 2008). Pollarding involves lopping branches at a certain height above the ground, out of the reach of browsing livestock, to obtain a product from the tree (e.g., fodder, firewood, charcoal, osier wickers for basketry, timber or small-scale wood for tool handles; Read 2003). Moreover, pollarding does not require grazing restriction afterward for protecting the new sprouts from browsing as after coppicing trees down at ground level (Read et al. 2013). This multipurpose land management became a key component of subsistence farming economy in Europe (Sjölund and Jump 2013), enabling the maintenance of large open woodlands as agroforestry systems (Smith et al. 2012). Pollarding was applied to a wide array of tree species (Read 2000; Rozas 2004; Rozas et al. 2008; Allende Álvarez et al. 2019; Camarero et al. 2022), and these practices were tightly controlled by traditional regulations and bylaws since at least the sixteenth century (Allende Álvarez et al. 2013; Gomez et al. 2016). As a result, pollard woodlands and trees out of woodlands constitute landscape elements of great cultural and conservational value that represent long-term sustainable land management systems (Grove and Rackham 2003; Haeggström 2007; Mansion 2010).
Sharp changes in demography, economic and agricultural activities, stock raising and forestry practices, linked to the industrialisation and rural depopulation during the last two centuries across most part of Europe, has caused the progressive abandonment or reduction of traditional multipurpose land uses, pollarding practices included (Read 1996; Moe and Botnen 2000; Leppik et al. 2011; Castro et al. 2012; Sjölund and Jump 2013). The practice of pruning trees at height is both costly and risky (Dujesiefken et al. 2005), and leaves and branches from trees are no longer a critical resource for local subsistence farming economies (Nordén et al. 2018). Consequently, pollarding practice has been progressively reduced or abandoned, and most former pollard woodlands have evolved into closed forests, or were transformed into forest plantations or agricultural lands, with this process intensifying during the latter half of the twentieth century (Pankhurst 2013). The few remaining pollard landscapes represent a true European cultural heritage which is often maintained only for their cultural value.
As a consequence of frequent pruning, and the globally recognized growth-lifespan trade-offs (Brienen et al. 2020), pollarded trees grow slower and live longer that uncut trees, especially in fast-growing species (Lonsdale 1996). In fact, in many European regions, pollarded trees are currently the oldest existing trees (Read 2000; Read et al. 2013; Nordén et al. 2018). Moreover, repeated cutting creates large and gnarled trunks, where rainwater collects in hollows creating packs of decaying wood that are appropriate niches for saproxylic organisms (Sebek et al. 2013; Zapponi et al. 2017; Doerfler et al. 2020). Thus, pollard woodlands are biodiversity storages preserving species associated with old-growth forests and hollow trees, being pollarded trees the main reservoir of these species in many areas (Skarpaas et al. 2011; Siitonen and Ranius 2015). Long-term persistence of pollards is strongly dependent on the maintenance of cutting practices, since growth of lateral branches on hollow trees promotes structural instability leading to breakages and eventually tree death (Read 2000). This management dependency makes the origination and preservation of aged managed woodlands possible, leading pollards as a reservoir of organisms linked to old-growth forest. Pollard management maintenance poses a strong conservation issue in those areas, where open pollard woodlands remain, but cutting practices has disappeared or become less frequent, triggering a gradual loss of large veteran trees and heading to an environmental and cultural impoverishment.
Nowadays, the Iberian Peninsula hosts the largest extension of pollard tree landscapes in Europe, with more than 3 million hectares of pollard woodlands known as dehesas (Moreno et al. 2007). Dehesas woodlands mainly dominated by Quercus ilex L. and Q. suber L. are widely represented in the low-populated rural landscapes of western Spain and Portugal (Pulido et al. 2001; Bugalho et al. 2018; Plieninger et al. 2021). However, relatively large and well-preserved pollard woodlands of narrow-leaved ash (Fraxinus angustifolia Vahl.) are also extensive in Central Spain. Pollarded ash woodlands persist at the foothills of the Guadarrama Mountains in the Central System, north of Madrid city. The high palatability of their leaves led ash trees to be treated as pollards for fodder across Europe; however, this practice has been progressively reduced with the historic drop in the stocks of extensive livestock and subsistence farming (Read 1996; Di Pasquale et al. 2004; Leppik et al. 2011). Therefore, the uninterrupted maintenance of traditional pollarding in ash, since at least Middle Age in areas close to a large city as Madrid is remarkable. The persistence of this practice is compromised not only due to a progressive urbanisation of the area and industrialization of the agriculture and livestock sectors, but also to the loss of traditional knowledge linked to the massive exodus of rural population along the second half of last century. Currently, we lack basic information on the historical frequency of pruning practice and its impact on tree growth. Rigorous historical records of the variation patterns of this silvicultural treatment are essential, since this information may be used to further apply the necessary treatments to preserve this valuable ecological and cultural landscape.
Dendrochronological techniques provide high-resolution information on environmental changes and disturbances occurring along the lifespan of trees (Speer 2010), which are widely used to reconstruct historical disturbance regimes in forests (Lorimer and Frelich 1989; Nowacki and Abrams 1997; Di Filippo et al. 2017). However, in contrast with the rich literature of natural forest dynamics reconstruction, the analysis of pollard events has received considerably less attention (Haas and Schweingruber 1993; Slotte 2001; Rozas 2004, 2005; Camarero et al. 2022). In this work, we reconstructed and analysed the historical sequence of pollarding events, and changes in their frequency and synchronicity in eight pollarded ash woodlands along the southern piedmont of Guadarrama Mountain in Central Iberian Peninsula. Our aim was to (1) characterize the wood anatomical markers of pruning events to enable the reconstruction of management patterns, (2) identify historical changes in pollarding frequency at tree and woodland levels, and (3) evaluate whether property type (public versus private) has affected management practices to create a baseline for recovering these activities.
Materials and methods
Sites description
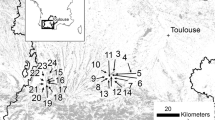
The eight woodlands constituted by narrow-leaved ash pollards that we studied are located on the southern slopes of the Guadarrama Mountains, Comunidad de Madrid, Spain, at altitudes ranging from 855 to 1067 m a.s.l. (Fig. 1). Climate is continental Mediterranean, with mean annual temperature ranging from 12 to 14 °C with a total annual precipitation of over 650 mm, with dry and hot summers (Fig. 1). Narrow-leaved ash woodlands occupy flat areas with moderate and seasonally hydromorphic soils (Caudullo and Houston Durrant 2016). The high phreatic level in these soils supports highly productive grasslands with a long growing period, eventually drying only at July and August. Currently, these areas are usually grazed by cattle, albeit sheep were also common in the past. We sampled three privately owned and five public woodlands showing hints of historical pollard management (Table 1).

Location of the eight pollarded woodlands (red squares) studied in Comunidad de Madrid, Spain (PU: public, PR: private), and distribution range of Fraxinus angustifolia in the central Iberian Peninsula (green area), according to Vallejo Bombín (2005). Inset map shows the location of the study area in Central Spain and the natural range of F. angustifolia in green (Caudullo et al. 2017). The climograph of PU1 site is shown
The large privately owned ash woodlands studied have a historical, architectural and cultural heritage of great interest. Among public owned woodlands, all but one belong to town councils with one, PU1, being a National Property (Patrimonio Nacional). Therefore, most of the public ash woodlands studied are part of the catalogue of public utility forests, mainly as municipal “dehesas boyales”, or “oxen pastures”. These properties are often enclosures on commons of villages with a livestock tradition for feeding working animals. In the wider surroundings of Madrid, villages have often grown and evolved into extended peri-urban housing areas, and this conurbation occasionally even surrounds the common pastures left as green islands within.
Sampling, ring-width measurement and chronologies computation
Sampling took place in summer–autumn 2021. Sampling was carried out following a wide transect in each of the ash woodlands studied. A minimum of 40 ash trees were selected at each of the 8 studied sites including only individuals with diameter wider than 20 cm and avoiding trees with external rot evidence. Trees available that met the specified criteria and were scattered across the chosen transect were selected for core extraction. A core was extracted at 1.3 m above ground level from each tree with a Pressler increment borer (Häglof, Sweden). We measured diameter and height from cored trees and georeferenced them using a submeter GPS device (Geo 7x, Trimble Ind., Sunnyvale, USA).
Cores were air-dried, glued to wooden mounts and surfaced with progressively finer grades of sandpaper for tree-ring analysis. Cores were digitized with CaptuRING® (García-Hidalgo et al. 2022) at 5897 dpi resolution and earlywood and latewood widths for each year were measured on the images with a precision of 0.004 mm with CooRecorder software (Cybys, Sweden). Cross-dating quality and measurement errors were checked and corrected using the software COFECHA (Holmes 1983). A mean chronology for each study site was calculated from those ring-width series highly inter-correlated. Three individual tree-ring width series out of 325 were omitted for further analyses due to low correlational values. At least 39 series corresponding to 39 trees per site were considered for further dendrochronological analysis (Table 1).
Pruning event identification
The extensive loss of the photosynthetic tissues associated with pollarding affects tree-ring formation (Haas and Schweingruber 1993; Schweingruber 2007), leading to abrupt and sustained growth depressions in the ring-width pattern (Rozas 2005; Bernard et al. 2006). Previous literature indicates that pruning in ash is associated with the formation of a distinctive second ring of smaller earlywood pores occurring instead of the formation of a typical most dense latewood (Haas and Schweingruber 1993). In the particular case of pollarded narrow-leaved ashes in Central Spain, the traditional management applied caused a different wood formation pattern. Since pollarding for fodder occurred in summer, at the end of the growing season, no resprouting took place until the next season, and no distinctive second ring of smaller earlywood pores was formed. A sudden growth reduction was observed the subsequent year after pruning, and earlywood width was not altered this year, but latewood is almost absent. Total ring-width decreased in the 2–3 following years after pruning, in comparison with the pre-pruning period, forming only narrower rings composed mostly of earlywood. After this period, latewood width and total ring width gradually increase over the next 3–4 years. Therefore, all these characteristics together allowed identifying all pollarding events carried out on each sampled ash throughout the tree life span (Fig. 2).

Wood-anatomy changes in narrow-leaved ash (Fraxinus angustifolia) wood cross section associated with pruning events. Differences in tree-rings growth when carrying out pollarding treatments (vertical shaded lines depict dated pruning events). The inset image shows one of the sampled ash plots (PU1) in the vicinity of the Royal Site of San Lorenzo de El Escorial
We dated discrete pruning events with an annual resolution on individual time series. A value of 1 was assigned to the first year in which tree rings presented hints of pruning, as indicated above. Otherwise, a value of 0 was assigned. For the subsequent analyses at site level, we considered a period comprised between the first year with at least four trees per site and the year 2021 (Table 1). With this data set, stand pruning recurrence at each sampling site was calculated as follows:
where k defines the time-span in years of the moving average (k = 10), t is the sequence of years within the period in which the pruning event is computed, n is the focal year in which the mean value of pruning events is computed, i is the tree of interest and finally, j is the number of trees in a sampling plot. The inverse step (x−1) allows us to convert discrete event information into a temporal scale indicating, therefore, the average elapsed time in years between two pruning events.
In addition, changes in pruning synchrony were assessed by means of a data set accounting for the total number of pruning events for every year and site grouped in 10-year wide periods lagged by 5 years moving time-windows. In each time-window we computed the variance (observed σ2) of the matrix consisting of events (1) and no events (0) per year. We also created two additional proxy data sets to assess the minimum and maximum variance (min σ2 and max σ2) possible on time-windows based on the existing pruning events. Decadal minimum variance was computed on a data set in which the total number of decadal events is homogeneously distributed along years. Decadal maximum variance was computed similarly in a data set with events concentrated in the minimum number of possible years. We calculated these values to reflect the real synchrony in relation to maximum and minimum possible pruning synchronies. Event synchrony was addressed for each window as follows:
The effect of land property and time in pruning recurrence was tested using a binomial generalised linear model. The model accounted for the first-order interaction between land property type (two-level factor variable; public and private) and time segmented in three 50-year windows (1871–1920, 1921–1970, 1971–2020; three-level factor variable). Time segmentation was done to assess historical difference in management, corresponding to different historical periods. Response variable was the data set of discrete events series at individual-scale. This model depicts the probability of finding a pruning event in a tree during a given land property type and during a given period. All analysis were performed in R environment (R Core Team 2021).
Results
We analysed 34643 tree-rings in the sampled trees, among which we identified 2426 pruning events at tree level along a 250-year period. Mean length of the analysed tree-ring series along a period with at least four trees per site, was 106 years, ranging from 88 years in PU2 to 145 in PU5 (Table 1). Several long-lived mature trees with more than 200 years were identified within the study areas. Since we were not able to reach the pith due to core-borer length limitations or presence of decomposed wood in the innermost parts in the largest trees, actual tree age was underestimated in several individuals. Average tree ring width ranged from 1.34 mm per year in PU5 to 2.07 mm in PR1 (Table 1). Series intercorrelation ranged from 0.563 to 0.643, and EPS values equal or superior to 0.85 in all sites indicated an adequate common signal among trees and high replication level (Table 1). The first pruning treatment was dated in 1777 in PR3.
Site level pollarding frequency, calculated in the period covered by at least four trees, could be analysed since the mid-nineteenth century for all the study sites (Table 1, Fig. 3), e.g., since 1830 in PU4. Pruning in privately owned plots occurred every 5–7 years until the 1970s decade. Three of the public sites (PU2, PU4, PU5) had a slightly longer pruning recurrence of up to 10 years, whereas PU1 and PU3 public sites showed even higher pruning recurrence until the 1970s decade. PU1, El Escorial site, showed a significant reduction of pruning events during the 1875–1915 period, probably related to decisions on its management by the Spanish Crown in 1875, just after the end of the first Spanish Republic (1873–1874).

Pruning recurrence of sampled plots defined as the inverse of the site-averaged 10-year simple moving average (SMA10) of trees within every private or public study site. Shaded areas depict periods with pruning recurrence higher than 20 years
The observed pollarding pattern showed only slight temporal differences in pruning frequency until the 1970s. For example, the average pruning frequency was extended to 15 years (e.g., 1920s, 1930s) or reduced to 7 years (e.g., 1940s) in PU5, to continue being carried out every 10 years until the general decline of the pollarding activity in the 1970s (Fig. 3). Since the 1970s decade a sharp decrease of pruning activity was evidenced leading to extended pruning turns. Pruning treatments were recovered in most sites during the 1980s decade, albeit at a lower and most irregular frequency.
Pruning synchrony was assessed for each site and decade (Fig. 4), except for those periods with infrequent events, i.e., with less than one event per decade. Pruning was synchronous across trees from the nineteenth century to the first half of twentieth century. Privately owned plots have high synchrony until the 1930–1950s, generally above 75% of the trees pruned in the same years, whereas pruning in public plots was more unevenly distributed for this period. Some public sites showed highly synchronous pruning events, such as PU4, but others exhibited periods of low synchrony, as occurred in PU2 in the 1900–1920 period. The most irregular and asynchronous pruning pattern was observed in both public and private sites since the 1970s, and matching the reduction in the number of events. Nevertheless, synchronized pruning events were also carried out during the last decade, as the high synchrony percentage in PU4 during the current century showed.

Pruning synchrony (%) observed for each private (PR) or public (PU) site. The number of decadal pruning events (histogram) is shown centred at the middle point of the 10 years moving windows. Shaded areas indicate periods without pruning events
To gain a better understanding of the interplay between pruning recurrence and synchrony patterns we graphed them simultaneously for each study site including the number of pollarding events occurring annually (Supplementary material, Figs. S2–S8). As an example, in the private study site PR3 we found a total of 316 pruning events among all the analysed series (Fig. 5). Periods with contrasting management were identified by changes in the frequency between pruning events throughout the study period (Fig. 5a). In the second half of the nineteenth century, pruning treatments were intensified and hence the recurrence between two pruning events was reduced from 15 to 5 years. This frequency remained steady until the beginning of the twentieth century. During the first half of the twentieth century, the period between successive pruning events remained almost constant between 5 and 7 years. Less frequent pruning events were observed from 1950 to 1970, carried out with a 7–9-year interval. After 1970 management regime changed with very few pruning events and less synchronous. Pruning activity resumed in the 1980 decade, but synchrony was not restored. Since then, pruning activity has remained sporadic and asynchronous (Fig. 5b).

Ensemble of pruning events analyses for PR3. a Sample depth (number of trees), pruning frequency as the average recurrence time in which pruning events are repeated (pruning recurrence, smoothed with a Gaussian kernel K = 5) and number of events that occurred at each year (pruning events). Shaded areas indicate decades without pruning events. b Evolution of synchrony patterns (pruning synchrony) and number of pruning events (pruning events) in 10-year periods with 5-year moving windows
During the last 150 years, pruning activity differed along the analysed periods and between public and private sites (Fig. 6). Model gave not support for the interaction between land property regime and time as mediators of pruning probability (Tables S1 and S2). Significant differences in the estimated marginal means were found regarding land use factors and between the two posterior and the current time period. Pruning significantly decreased since 1971 in comparison with the two previous periods. Pruning activity in private lands always maintained higher than in public sites, nevertheless, these differences have been getting close over time.

Model-estimated probability of pruning (odds ratios with standard errors) over 50-year time periods by land use (public or private sites). Different letters indicate significant differences in mean pruning frequency at p < 0.05 level
Discussion
Pollarding ash trees for fodder mainly occurs in summer when drought reduces the availability of fresh grass for cattle pasture (Haas et al. 1998; Vandermeulen et al. 2018) and fodder quality is optimal (FRAXIGEN 2005). This traditional practice enables livestock to withstand the most critical period of the year under Mediterranean climate, and at the same time reduces the costs of supplementary feeding acquisition (Halstead 1998; Papanastasis et al. 1998, 2008; Papachristou et al. 1999). Literature on ash species pollarding in Central Europe indicates that latewood formation is arrested after summer branch cutting with a characteristic small-vesseled band of earlywood produced to support branches resprouting in the same season (Haas and Schweingruber 1993). However, pollarded ash trees in Central Spain did not show this characteristic false ring. Such difference can be attributed to regional summer climate conditions when pruning activities take place. In contrast to the mild and relatively wet summers in Central Europe, summer in our study area is characterised by a hot and dry period that considerably limits ash resprouting ability and cambial resumption. Thus, resprouting is delayed until the start of the next growing season (F. Castro pers. obs.), when wet and mild conditions in spring enable the production of a wider than usual earlywood (Fig. 2), probably to support new sprouting and the treetop reconstruction. Such regrowth drains tree resources as appreciated in the low density of this ring with a low proportion of fibres than the latewood. In fact, the next two or three annual rings are almost completely composed by earlywood, probably because of a higher effort devoted to branch resprouting than to radial growth. After these years, latewood formation is gradually recovered until pre-cutting rings latewood growth levels are reached (Fig. 2).
Historical pruning recurrence in narrow-leaved ash woodlands of Central Spain ranged between 5 and 10 years (Fig. 3). The frequency of pollarding is highly variable worldwide, depending on species, the obtained product, site quality or local traditions. When the main purpose is to obtain wood for charcoal production, firewood or increase fruit production on slow-growing species, inter-pruning cycles are longer, ranging 10–50 years in beech (Dagley and Burman 1996; Read 2000), 15–20 years in Spanish holm oak (Gómez Gutiérrez and Pérez Fernández 1991) or 10–76 years in deciduous oaks (Rozas 2004; Haneca et al. 2009). In contrast, when the objective is fodder production, species of high palatability are usually selected and cutting cycles are shorter. In the United Kingdom, ash and holly were cut for fodder at 5–10-year frequency (Spray 1980), but frequency could be even higher (2–6 years) in common ash trees in Switzerland, Sweden or Northern Spain (Haas and Schweingruber 1993; Read 2003). The slightly lower frequency of narrow-leaved ash pruning could be related to the more limiting summer conditions, which imply a lack of the current year resprouting ability. In any case, the relative high frequency can be attributed to the rapid growth rate of ash, as well as to a fodder-oriented production that requires a shorter recovery period than a fuel or construction wood-oriented management.
Our reconstruction of pruning events reflected a sharp reduction of this management practice since the 1970s, with a sporadic recovery in last decades, but lacking the synchronic and systematic pattern that we observed in the previous periods. Land ownership was also relevant, with higher historical and current cutting frequency and synchrony found in privately owned woodlands than in public lands. Pollarding in Central Spain occurred few years after pre-pollarding secondary growth rates were almost completely recovered. Besides, pruning frequency in a population was strongly synchronized among trees, since most of the trees within a site were simultaneously cut in the same years. Since livestock needed fodder every summer, our results suggest that probably ash woodlands were subdivided into smaller management units to ensure a yearly fodder production through a rotational schema (Gómez-Sal et al. 1992; Díaz et al. 1997; Moreno and Pulido 2009). Results suggest that trees sampled belonged to one management unit in most of the sites (but see results from PU5 in Fig. 4). This means that the simultaneously pollarded trees belonged to the same cyclical pollarding unit within a rotational schema.
Land ownership affected pruning frequency with cutting cycle ranging from 5 to 7 years in private lands to 10–15 years in public properties, with this difference being maintained along the whole studied period, even under current management with a low pruning frequency. The issue of forest ownership has recently received growing attention in both research and policy ambits (Ficko et al. 2019; Weiss et al. 2019). The objectives and motivation of forest owners to manage the pollarded stands vary substantially, depending on the products and services provided, but there are also considerable geographical variations due to cultural reasons. Public forest owners generally actively manage their woodlands according to multi-purpose political, societal, conservational and business objectives, whereas there is less variation in the purpose of private woodlands. In public woodlands, while the local and shared production of natural resources was widely true before rural emigration, new social trends seek for environmental objectives such as the preservation of highly valuable cultural and aesthetic landscapes over the demand for income from private agroforestry uses. On the contrary, private woodland owners try to reach greater economic efficiency in forest management (i.e., higher harvesting frequency to maximize benefits), leading to shorter exploitation turns. Interestingly, we observed few signs of tree degradation in private lands in comparison with public ones, despite higher pruning recurrence.
Until the 1950s, we observed a fairly regular pruning pattern, evidencing a most systematic management carried out in these agroforestry systems. Traditional management of ash trees collapsed at the 1970s with a generalized and drastic decline of pollarding activity. This period matches with the rural exodus and urbanization of the society, and the strong upsurge of Spanish economy (De la Torre and García-Zúñiga 2014), with a rapid internal population reorganization that transformed radically rural landscapes driving a rapid abandonment of traditional activities (MacDonald et al. 2000; Rozas et al. 2008; Lasanta et al. 2021). As a consequence, many farmers and landowners abandoned traditional farming practices highly valuable for cultural heritage conservation (Plieninger et al. 2021). Albeit Madrid province received a huge number of immigrants, this phenomenon was concentrated in the capital and its conurbation. Population in rural Guadarrama piedmonts either declined or remained stable until the large increase in the 1990s (Hewitt and Escobar 2017), whereas rural economy shifted from the primary to the third sector intended to second residences construction and services.
One of the major concerns of pruning abandonment is the effect of cutting cessation on tree health and life expectancy of the pollard trees (Rozas 2005; Rotherham 2013; Sjölund and Jump 2013). Many pollarded stands have disappeared due to land-use changes (Götmark 2013) and most of the still existing populations have developed oversized branches on hollow trunks with a core of decaying wood that could collapse easily (Pankhurst 2013). Repeated cutting produces injuries in the trees and creates a bolling trunk of uneven shape (Read 2000). In fact, pruning-abandoned trees generally showed higher and unbalanced crowns, increasing the possibility of heavy branches breaking. The southern slopes of Guadarrama mountains have many surviving pollarded trees, and some of them are older than 200 years. We identified pollarded trees exceeding 250 years in the studied populations, which confirmed that periodical managing ensures high productivity and enables tree longevity, being currently the oldest living trees in the studied area. With respect to the time window in which pruning activity can be successfully recovered after its abandonment, our study brings valuable information on the ability of trees to return to pollarding cycles after 30-year periods without management, as, for example, 1885–1916 in PU1. Although, our approach is skewed, since we do not retrieve information on failure rates (i.e., dead trees after pollarding were reinstated), but it provides an estimation of available period for pruning activity recovery.
In general terms, we observed an evolution from the initial, synchronous and uniform treatments carried out until the mid-twentieth century, which was more intense in private stands, to irregular and asynchronous management patterns in recent decades. An increasing synchrony of pruning has been detected in some publicly owned woodlands responding to the current conservation efforts to restore and maintain functional pollarded open woodlands. Future management of pollarded woodlands is dependent upon availability of resources and staff with enough knowledge and experience to maintain this valuable resource which remains common in several Mediterranean, Balkan and Scandinavian countries. Having competent well-trained foresters is, therefore, a prerequisite for successful management and long-term survival (Pankhurst 2013), particularly to restore lapsed pollards which require a more sophisticated treatment (Read et al. 2013, 2021). Ash dieback (Hymenoscyphus fraxineus (T. Kowalski) Baral et al.) is also a concern for ash populations in Spain, including F. angustifolia, with potential impact in pollard populations, as the recent report of the presence of the pathogen in Spain remarks (Stroheker et al. 2021). Moreover, to recover and preserve pollarded stands, managers should be able to access site history for the trees especially if the last pollarding date is unknown. In this sense, we prove that dendrochronology is very appropriate to reconstruct the variation of management patterns over the history of these pollarded woodlands.
Traditional management in pollard ashes has the ability to provide valuable livestock food while maintaining a biodiverse ecosystem thanks to the creation of diverse wildlife niches. Unlike other pollarding systems, where cessation of traditional activities marked a no return point, pollarding has been recovered in most of the sites albeit at lower frequency and without the traditional synchronic approach (Fig. 5). This might be a consequence of the easy application of pollarding in our study area being technically easier, since cutting is performed at a relatively low height of at least 2 m above the ground and ash occupy flat areas, where machinery can operate. Narrow-leaved ash woodlands have optimal conditions to restore pollarding activity (Read et al. 2021), providing technical and economic support to make this traditional activity profitably would have strong environmental revenue. It would contribute to preserve this rich cultural landscape, its venerable trees and its associated biodiversity, while simultaneously, by reducing the need of supplementary food, it would also contribute to the reduction of the environmental impact of meat production, one of the largest greenhouse gas emission issues (IPCC 2022).
Conclusions
The unequivocal dating of pruning events in Central Spanish pollarded ash woodlands enabled to track changes in pruning frequency since the nineteenth century. Traditional management involved a 5–10-year rotation interval with a synchronous pruning within management units and higher frequency in private lands. This management schema collapsed in the 1970s decade during the rural exodus and urbanization of population of inner Spain. However, after several decades of almost no pollarding activity, pruning has recovered due to economic and/or conservational purposes in most of the stands albeit with much lower intensity and without the characteristic synchrony of traditional management patterns. Supporting this recovery would have multiple benefits, such as preserving historical and environmental values, but may also contribute to the reduction of the environmental impact of meat production.
References
Allende Álvarez F, Gomez-Mediavilla G, López-Estébanez N (2019) Pollard forest of Fraxinus angustifolia in the centre of Iberian Peninsula: protection and management. In: Allende Álvarez F, Gomez-Mediavilla G, López-Estébanez N (eds) Silvicultures - management and conservation. IntechOpen, UK
Allende Álvarez F, Gómez Mediavilla G, López Estébanez N, Sobrino García J (2013) Podas y trasmochos en las ordenanzas forestales del Sistema Central español y su impronta en el paisaje forestal actual. Cuad Soc Esp Cienc For 35–41
Bernard V, Renaudin S, Marguerie D (2006) Evidence of trimmed oaks (Quercus sp.) in north western France during the early middle ages (9th-11th centuries A.D.). In: Dufraisse A (ed) Charcoal analysis: new analytical tools and methods for archaeology. BAR Publishing, UK, pp 103–108
Brienen RJW, Caldwell L, Duchesne L et al (2020) Forest carbon sink neutralized by pervasive growth-lifespan trade-offs. Nat Commun 11:4241. https://doi.org/10.1038/s41467-020-17966-z
Bugalho MN, Pinto-Correia T, Pulido F (2018) Human use of natural capital generates cultural and other ecosystem services in montado and dehesa oak woodlands. In: Paracchini ML, Zingari PC, Blasi C (eds) Reconnecting natural and cultural capital: contributions from science and policy. Publications Office of the European Union, Luxembourg, pp 115–123
Camarero JJ, González de Andrés E, Colangelo M, de Jaime LC (2022) Growth history of pollarded black poplars in a continental Mediterranean region: a paradigm of vanishing landscapes. For Ecol Manag 517:120268. https://doi.org/10.1016/j.foreco.2022.120268
Castro A, Martínez De Murguía L, Fernández J et al (2012) Size and quality of wood used by Rosalia alpina (Linnaeus, 1758) (Coleoptera: Cerambycidae) in beech woodlands of Gipuzkoa (northern Spain). Munibe 60:77–100
Caudullo G, Houston Durrant T (2016) Fraxinus angustifolia in Europe: distribution, habitat, usage and threats. In: San-Miguel-Ayanz J, de Rigo D, Caudullo G et al (eds) European atlas of forest tree species. Publications Office of the European Union, Luxembourg
Caudullo G, Welk E, San-Miguel-Ayanz J (2017) Chorological maps for the main European woody species. Data Br 12:662–666. https://doi.org/10.1016/j.dib.2017.05.007
Dagley J, Burman P (1996) The management of the pollards of Epping forest: its history and revival. In: Read HJ (ed) Pollard and veteran tree management II. The Richmond Publishing Company, UK, pp 29–41
De la Torre J, García-Zúñiga M (2014) Was it a Spanish miracle? Development plans and regional industrialization, 1950–1975. In: Grabas C, Nützenadel A (eds) Industrial Policy in Europe after 1945. Palgrave Macmillan, London, pp 162–183
Di Filippo A, Biondi F, Piovesan G, Ziaco E (2017) Tree ring-based metrics for assessing old-growth forest naturalness. J Appl Ecol 54:737–749. https://doi.org/10.1111/1365-2664.12793
Di Pasquale G, Di Martino P, Mazzoleni S (2004) Forest history in the Mediterranean region. In: Mazzoleni S, di Pasquale G, Mulligan M et al (eds) Recent dynamics of the Mediterranean vegetation and landscape. John Wiley & Sons, Ltd, Chichester, UK, pp 13–20
Díaz M, Campos P, Pulido FJP (1997) The Spanish dehesas: a diversity in land-use and wildlife. In: Pain D, Pienkowski M (eds) Farming and Birds in Europe. Academic Press, Cambridge, pp 178–209
Doerfler I, Cadotte MW, Weisser WW et al (2020) Restoration-oriented forest management affects community assembly patterns of deadwood-dependent organisms. J Appl Ecol 57:2429–2440. https://doi.org/10.1111/1365-2664.13741
Dujesiefken D, Drenou C, Oven P, Stobbe H (2005) Arboricultural Practices. In: Konijnendijk C, Nilsson K, Randrup T, Schipperijn J (eds) Urban Forests and Trees. Springer, Berlin, Heidelberg, pp 419–441
Ficko A, Lidestav G, Ní Dhubháin Á et al (2019) European private forest owner typologies: a review of methods and use. For Policy Econ 99:21–31. https://doi.org/10.1016/j.forpol.2017.09.010
FRAXIGEN (2005) Ash species in Europe. Oxford Forestry Institute, University of Oxford, UK
García-Hidalgo M, García-Pedrero Á, Colón D et al (2022) CaptuRING: a do-it-yourself tool for wood sample digitization. Methods Ecol Evol. https://doi.org/10.1111/2041-210x.13847
Gomez G, López N, Allende F (2016) Las fresnedas trasmochadas del piedemonte del Sistema Central en Madrid (España): cambios y usos actuales. Estud Rural 6:32–47
Gómez Gutiérrez JM, Pérez Fernández MA (1991) Manejo del arbolado y del matorral. In: Gómez Gutiérrez JM (ed) El libro de las dehesas salmantinas. Junta de Castilla y León, Salamanca, pp 321–340
Gómez-Sal A, Rodríguez MA, De Miguel JM (1992) Matter transfer and land use by cattle in a dehesa ecosystem of central Spain. Vegetatio 99–100:345–354. https://doi.org/10.1007/BF00118241
Götmark F (2013) Habitat management alternatives for conservation forests in the temperate zone: review, synthesis, and implications. For Ecol Manag 306:292–307. https://doi.org/10.1016/j.foreco.2013.06.014
Grove AT, Rackham O (2003) The nature of Mediterranean Europe: an ecological history. Yale University Press, Yale
Haas JN, Schweingruber FH (1993) Wood-anatomical evidence of pollarding in ash stems from the Valais, Switzerland. Dendrochronologia 11:35–43
Haas JN, Karg S, Rasmussen P (1998) Beech leaves and twigs used as winter fodder: examples from historic and prehistoric times. Environ Archaeol 1:81–86. https://doi.org/10.1179/env.1996.1.1.81
Haeggström CA (2007) Les trognes dans l’art. In: Dumont E (ed) Les trognes en Europe. Rencontres autour des arbres têtards et des arbres d’emonde. Maison Botanique, Boursay, pp 134–140
Halstead P (1998) Ask the fellows who lop the hay: leaf-fodder in the mountains of northwest Greece. Rural Hist 9:211–234. https://doi.org/10.1017/S0956793300001588
Haneca K, Čufar K, Beeckman H (2009) Oaks, tree-rings and wooden cultural heritage: a review of the main characteristics and applications of oak dendrochronology in Europe. J Archaeol Sci 36:1–11. https://doi.org/10.1016/j.jas.2008.07.005
Hewitt R, Escobar F (2017) The territorial dynamics of fast-growing regions: unsustainable land use change and future policy challenges in Madrid, Spain. Appl Geogr 31:650–667. https://doi.org/10.1016/j.apgeog.2010.11.002
Holmes RL (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull 43:69–78
IPCC (2022) Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press
Lasanta T, Nadal-Romero E, Khorchani M, Romero-Díaz A (2021) A review of abandoned lands in Spain: from local landscapes to global management strategies. Geogr Res Lett 47:477–521. https://doi.org/10.18172/cig.4755
Leppik E, Jüriado I, Liira J (2011) Changes in stand structure due to the cessation of traditional land use in wooded meadows impoverish epiphytic lichen communities. Lichenol 43:257–274. https://doi.org/10.1017/S002428291100003X
Lonsdale D (1996) Pollarding success or failure; some principles to consider. In: Read HJ (ed) Pollard and veteran tree management II. The Richmond Publishing Company, UK, pp 100–104
Lorimer CG, Frelich LE (1989) A methodology for estimating canopy disturbance frequency and intensity in dense temperate forests. Can J for Res 19:651–663
MacDonald D, Crabtree JR, Wiesinger G et al (2000) Agricultural abandonment in mountain areas of Europe: environmental consequences and policy response. J Environ Manag 59:47–69. https://doi.org/10.1006/jema.1999.0335
Mansion D (2010) Les trognes, l’arbre paysan aux mille usages. Editions Ouest- France, Rennes
Moe B, Botnen A (2000) Epiphytic vegetation on pollarded trunks of Fraxinus excelsior in four different habitats at Grinde, Leikanger, western Norway. Plant Ecol 151:143–159. https://doi.org/10.1023/A:1026585911823
Moreno G, Pulido FJ (2009) The Functioning, Management and Persistence of Dehesas. In: Rigueiro-Rodríguez A, McAdam J, Mosquera-Losada MR (eds) Agroforestry in Europe. Springer, Netherlands, Dordrecht, pp 127–160
Moreno G, Obrador JJ, García A (2007) Impact of evergreen oaks on soil fertility and crop production in intercropped dehesas. Agric Ecosyst Environ 119:270–280. https://doi.org/10.1016/j.agee.2006.07.013
Nordén B, Jordal JB, Evju M (2018) Can large unmanaged trees replace ancient pollarded trees as habitats for lichenized fungi, non-lichenized fungi and bryophytes? Biodivers Conserv 27:1095–1114. https://doi.org/10.1007/s10531-017-1482-x
Nowacki GJ, Abrams MD (1997) Radial-growth averaging criteria for reconstructing disturbance histories from presettlement-origin oaks. Ecol Monogr 67:225–249. https://doi.org/10.1890/0012-9615(1997)067[0225:RGACFR]2.0.CO;2
Pankhurst M (2013) There’s more to a pollard than meets the eye! the views and experience of a woodland ranger. Arboric J 35:91–98
Papachristou TG, Platis PD, Papanastasis VP, Tsiouvaras CN (1999) Use of deciduous woody species as a diet supplement for goats grazing Mediterranean shrublands during the dry season. Anim Feed Sci Technol 80:267–279. https://doi.org/10.1016/S0377-8401(99)00056-5
Papanastasis VP, Platis PD, Dini-Papanastasi O (1998) Effects of age and frequency of cutting on productivity of Mediterranean deciduous fodder tree and shrub plantations. For Ecol Manag 110:283–292. https://doi.org/10.1016/S0378-1127(98)00293-X
Papanastasis VP, Yiakoulaki MD, Decandia M, Dini-Papanastasi O (2008) Integrating woody species into livestock feeding in the Mediterranean areas of Europe. Anim Feed Sci Technol 140:1–17. https://doi.org/10.1016/j.anifeedsci.2007.03.012
Petit S, Watkins C (2003) Pollarding trees: changing attitudes to a traditional land management practice in Britain 1600–1900. Rural Hist 14:157–176. https://doi.org/10.1017/S0956793303001018
Plieninger T, Flinzberger L, Hetman M et al (2021) Dehesas as high nature value farming systems: a social-ecological synthesis of drivers, pressures, state, impacts, and responses. Ecol Soc. https://doi.org/10.5751/ES-12647-260323
Pulido FJ, Díaz M, Hidalgo De Trucios SJ (2001) Size structure and regeneration of Spanish holm oak Quercus ilex forests and dehesas: effects of agroforestry use on their long-term sustainability. For Ecol Manag 146:1–13. https://doi.org/10.1016/S0378-1127(00)00443-6
R Core Team (2021) R: A language and environment for statistical computing. R Foundation for Statistical Computing
Read H (2000) Veteran trees: a guide to good management. English Nature, Peterborough
Read HJ (2008) Pollards and pollarding in Europe. Br Wildl 19:250–259
Read HJ, Dagley J, Elosegui JM et al (2013) Restoration of lapsed beech pollards: evaluation of techniques and guidance for future work. Arboric J 35:74–90. https://doi.org/10.1080/03071375.2013.747720
Read H, Bengtsson V, Castro A et al (2021) Restoration of lapsed beech pollards in the Basque region of Spain: comparison of different cutting techniques. Arboric J 43:140–155. https://doi.org/10.1080/03071375.2020.1854993
Read HJ (1996) Pollard and veteran tree management II. Incorporating the proceedings of the meeting hosted by the Corporation of London at Epping Forest in 1993. The Richmond Publishing Company, Berkshire
Read HJ (2003) A study of practical pollarding techniques in northern Europe. Pers Commun
Rotherham ID (2013) Managing veterans for future conservation. Arboric J 35:61–63. https://doi.org/10.1080/03071375.2013.826528
Rozas V (2004) A dendroecological reconstruction of age structure and past management in an old-growth pollarded parkland in northern Spain. For Ecol Manag 195:205–219. https://doi.org/10.1016/j.foreco.2004.02.058
Rozas V (2005) Dendrochronology of pedunculate oak (Quercus robur L.) in an old-growth pollarded woodland in northern Spain: establishment patterns and the management history. Ann for Sci 62:13–22. https://doi.org/10.1051/forest:2004091
Rozas V, Olano JM, DeSoto L, Bartolomé D (2008) Large-scale structural variation and long-term growth dynamics of Juniperus thurifera trees in a managed woodland in Soria, central Spain. Ann for Sci 65:809. https://doi.org/10.1051/forest:2008066
Schweingruber FH (2007) Wood structure and environment. Springer, Berlin, Heidelberg
Sebek P, Altman J, Platek M, Cizek L (2013) Is active management the key to the conservation of saproxylic biodiversity? Pollarding promotes the formation of tree hollows. PLoS ONE 8:1–6. https://doi.org/10.1371/journal.pone.0060456
Siitonen J, Ranius T (2015) The importance of veteran trees for saproxylic insects. In: Kirby KJ, Watkins C (eds) Europe’s changing woods and forests: from wildwood to managed landscapes. CABI, Croydon, UK, pp 140–153
Sjölund MJ, Jump AS (2013) The benefits and hazards of exploiting vegetative regeneration for forest conservation management in a warming world. Forestry 86:503–513. https://doi.org/10.1093/forestry/cpt030
Skarpaas O, Diserud OH, Sverdrup-Thygeson A, Ødegaard F (2011) Predicting hotspots for red-listed species: multivariate regression models for oak-associated beetles. Insect Conserv Divers 4:53–59. https://doi.org/10.1111/j.1752-4598.2010.00109.x
Slotte H (2001) Harvesting of leaf-hay shaped the Swedish landscape. Landsc Ecol 16:691–702. https://doi.org/10.1023/A:1014486331464
Smith J, Pearce BD, Wolfe MS (2012) A European perspective for developing modern multifunctional agroforestry systems for sustainable intensification. Renew Agric Food Syst 27:323–332. https://doi.org/10.1017/S1742170511000597
Speer JH (2010) Fundamentals of tree-ring research. University of Arizona Press, Tucson, Arizona
Spray M (1980) Holly as a fodder in England. Agric Hist Rev 29:97–100
Stroheker S, Queloz V, Nemesio-Gorriz M (2021) First report of Hymenoscyphus fraxineus causing ash dieback in Spain. New Dis Reports 44:e12054. https://doi.org/10.1002/ndr2.12054
Vallejo Bombín R (2005) El mapa forestal de España escala 1:50.000 (MFE50) como base del tercer inventario forestal nacional. Cuad Soc Esp Cienc for 19:205–210
Vandermeulen S, Ramírez-Restrepo CA, Marche C et al (2018) Behaviour and browse species selectivity of heifers grazing in a temperate silvopastoral system. Agrofor Syst 92:705–716. https://doi.org/10.1007/s10457-016-0041-x
Weiss G, Lawrence A, Hujala T et al (2019) Forest ownership changes in Europe: state of knowledge and conceptual foundations. For Policy Econ 99:9–20. https://doi.org/10.1016/j.forpol.2018.03.003
Zapponi L, Mazza G, Farina A et al (2017) The role of monumental trees for the preservation of saproxylic biodiversity: re-thinking their management in cultural landscapes. Nat Conserv 19:231–243. https://doi.org/10.3897/natureconservation.19.12464
Acknowledgements
We thank the comments provided by the reviewers to improve the manuscript. Special thanks are due to other members of the Cambium Research Group (María A. García López and Jaime Madrigal-González) and María Sánchez who contributed to sample collection, and to Juan Carlos Rubio and Alfonso Martínez for assistance in sample preparation. We appreciate the collaboration of public and private environmental staff (P. Sanjuanbenito García, Y. Baselga Aymerich, M.M. García Herguido, A. Bonilla Rodríguez, M.A. Hontoria Suárez). We are also grateful to forest managers (Parque Nacional de la Sierra de Guadarrama and Comunidad de Madrid) and to public (Patrimonio Nacional, Ayuntamiento de Soto del Real, Ayuntamiento de El Berrueco, Ayuntamiento de Moralzarzal and Ayuntamiento de Villavieja del Lozoya) and private (Finca Prado Nuevo, Finca La Granjilla and Finca Prado Herrero) owners for granting access and sampling permission.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This work was funded by the Junta de Castilla y León projects CLU-2019-01, VA171P20 and IR2020-1-UVA08, and the Spanish MCIN/AEI/10.13039/501100011033 project PROWARM (PID2020-118444GA-100). DC-P was supported by the project "CLU-2019-01 - iuFOR Institute Unit of Excellence" of the University of Valladolid, funded by the Junta de Castilla y León and co-financed by the European Union (ERDF "Europe drives our growth"). GS-B was supported by a Postdoctoral grant (IJC2019-040571-I) funded by MCIN/AEI/10.13039/501100011033. MG-H was supported by Grant PRE2018-084106 funded by MCIN/AEI /10.13039/501100011033 and "ESF Investing in your future".
Author information
Authors and Affiliations
Contributions
JMO and FC contributed to the study conceptualization. Methodology, material preparation and fieldwork were performed by all authors. DC-P carried out the data collection. Formal analysis was performed by HH-A, JMO and DC-P. The first draft of the manuscript was written by DC-P and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Communicated by Eryuan Liang.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Candel-Pérez, D., Hernández-Alonso, H., Castro, F. et al. 250-Year reconstruction of pollarding events reveals sharp management changes in Iberian ash woodlands. Trees 36, 1909–1921 (2022). https://doi.org/10.1007/s00468-022-02343-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-022-02343-8