
Abstract
Key message
Lowered pollen and seed quality of B. humilis compared to its widespread tree relatives may threaten its populations at the southwestern edge of the species range.
Abstract
Inability to produce vital propagules is the main threat to the persistence of rare plants at the edges of species distribution. In the present investigation, factors responsible for the low germinability of shrub birch (Betula humilis) seeds in the peripheral populations were analysed. As shrub birch populations are found in phosphorus (P)-limited peatlands with high groundwater levels, the impact of groundwater PO43− ion contents and groundwater table for B. humilis flowering was also studied. Qualities of the B. humilis seeds and pollen were compared to those of tree congeners silver birch (B. pendula) and downy birch (B. pubescens), which usually represent a high reproductive performance. We revealed that the poor germination ability of B. humilis seeds was caused by the high proportions of empty seeds (44.8%) and seeds with partially developed embryos (47.3%). The insufficient numbers of viable seeds may be a consequence of low-quality shrub birch pollen, as only 1.6% of pollen grains sprouted. The germinability of shrub birch pollen was significantly lower than that of Betula tree pollen; thus, there is a risk of pollination of B. humilis female flowers by pollen grains of tree birches. Positive relationships between the P concentrations in seeds and seed germinability were revealed in tree birches, but not in B. humilis. The availability of PO43− ions in the groundwater did not affect B. humilis flowering, but male inflorescences were more numerous in stands with higher groundwater levels. Consequently, the maintenance of peatlands is crucial for the efficient sexual reproduction of shrub birches.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Human-driven climate change, habitat fragmentation and degradation resulting from the human demography and economy, overexploitation of organisms, pollution, and invasion of alien species are the ultimate reasons for the current species extirpation (Borges et al. 2019; Gaitán-Espitia and Hobday 2021). Plants, as sedentary organisms, seem to be especially vulnerable to environmental changes. The proximate cause of plant species decline results from their inability to produce vital propagules necessary to replace generations to sustain populations (Hamrick and Godt 1996; Shivanna and Tandon 2014). Unfortunately, the formation of seed sets, seed germination or seedling establishment is often impossible in adverse habitats (Barrett 2015). Sexual reproduction can be especially disturbed in the marginal populations of rare plant species, as they usually occupy suboptimal environmental conditions and are smaller and more isolated than populations from the centre of species distribution (Jump and Woodward 2003; Rasmussen and Kollmann 2004). In temperate regions, most endangered and slow-growing plants, often poor competitors, are found at P (phosphorus)-restricted sites (Fujita et al. 2014). In the P-poor habitats the number of species is limited; thus, less competitive plants have a chance to develop (Wassen et al. 2005; Fujita et al. 2014). However, at P-limited sites, plants invest less in growth and sexual reproduction, which results in delayed propagation, shorter flowering periods, reduced fruiting, and lower numbers of seeds (Nord et al. 2011; Fujita et al. 2014). In turn, spatial isolation of peripheral localities can prevent gene flow; consequently, a decrease in population viability due to the genetic drift and inbreeding depression can be observed (Ellstrand and Elam 1993). Furthermore, it is likely that lack of conspecific, genetically different mates in small, isolated populations will increase the risk of hybridisation with a common congener (Rhymer and Simberloff 1996). All these phenomena can drive small, isolated populations to extinction (Rasmussen and Kollmann 2004).
One of the endangered plant species in western and central Europe is the shrub birch (Betula humilis Schrk.). The southwestern boundary of the continuous range of B. humilis is located in Poland, where its population decreased approximately fivefold during the twentieth century due to meliorations and draining in the past and present-day climate change (Załuski et al. 2014). Shrub birch can inhabit diverse habitats, from entirely natural habitats of the species to their drained variants (Jabłońska 2012). However, the plant as an inferior competitor can survive on poor, subneutral and alkaline fens and spring mires and alkaline wet meadows, where conditions for development of other species are difficult (Jabłońska 2012; Załuski et al. 2014). When the groundwater table is low (> 10 cm below the peat surface), B. humilis suffers from the overgrowing by competitive forest and scrub plants (Jabłońska 2012; Załuski et al. 2014). Thus, the subsequent threats for marginal populations of B. humilis are its two closely related congeners, the silver birch (Betula pendula Roth) and downy birch (Betula pubescens Ehrh.). Although tree birches always coexist with endangered shrub birch, they are selectively superior to this species in human- or climate-impacted habitats. They represent a wide ecological tolerance and produce pollen grains in large quantities, and their seeds exhibit high germination capacity (Atkinson 1992; Franiel 2012).
To our knowledge, there is no information about pollen production in the B. humilis populations. In turn, the production of seeds at the southwestern margin of the species distribution seems to be disturbed. It was evidenced that the number of germinated seeds was significantly lower in the marginal Polish localities (median values from 0 to 16.5 per individual) compared to subcentral populations situated in Belarus (median values from 1 to 32 per individual) (Chrzanowska et al. 2016). The outcome of second analysis of B. humilis seed germination capacity conducted in six marginal Polish populations was even more distressing. Bona et al. (2019) revealed that only 2.7% of seeds were able to sprout, and the median numbers of germinated seeds per shrub birch individual ranged from 0 to 3.5. Habitat conditions appear to be the ultimate factor of poor germination capacity of shrub birch propagules as seeds from localities with the high groundwater table and no canopy cover or canopy cover of 50% were significantly heavier and reached higher germination capacity than seeds sampled in dry and fully shaded stands (Chrzanowska et al. 2016). However, the proximate reason responsible for low sprouting success of the B. humilis seeds remains unknown. In our opinion, there are several possible explanations for the reduced germination success of endangered B. humilis. The first explanation is a substantial contribution of empty seeds (without embryo), as was discovered in relict central European localities of dwarf birch (Betula nana L.) (Schwartz and Poschlod 2015; Jadwiszczak et al. 2017). Empty seed production can result from a decrease in the quality or quantity of pollen (pollen limitation), which can significantly reduce maternal plants’ reproductive success (Knight et al. 2005; Balogh and Barrett 2018). Another possible explanation for the germination failure of shrub birches is inadequate phosphorus contents in the seeds. It is likely that B. humilis individuals populating P-limited sites produce seeds with low P contents. As P is of prime importance for early development of seedlings (White and Veneklaas 2012 and references therein), P deficiency in the seeds could limit their germination ability.
In general, after applying Holm–Bonferroni's correction, Chrzanowska et al. (2016) revealed that the chemical parameters of the studied habitat did not influence the reproductive efficiency of shrub birch. However, the regression models for the total number of inflorescences (F = 4.489, P = 0.045) and the number of male inflorescences (F = 4.267, P = 0.026) per flowering individual were close to the significance. These outcomes resulted from a significant impact of the PO43− concentration variable on the number of male inflorescences and, then, on the total number of catkins per flowering specimen. This might suggest positive association between the abundance of phosphorus and the number of male inflorescences in the B. humilis populations. The quantity of pollen is positively correlated with the number of flowers in the anemophilous tree species (Tormo Molina et al. 1996); therefore, it still seems possible that P concentrations in groundwater can positively impact the reproductive capacity of shrub birch.
The main objective of this study was to reveal the causes of the low germination ability of shrub birch seeds in populations situated at the southwestern margin of the species range. To assess the sexual reproduction performance of endangered B. humilis, data obtained for this species were contrasted with pollen and seed quality of B. pendula and B. pubescens inhabiting the same peatlands. The following hypotheses were tested: (1) the low germinability of shrub birch seeds results from a lack of embryos present in seed coats; (2) shrub birch pollen quality is lower than that of tree birches; (3) B. humilis individuals with higher P content in seeds manifest higher seed germination capacity, and finally (4) flowering intensity of B. humilis does depend on groundwater P.
Materials and methods
Sample collection and laboratory analyses
Seed quality of birches
All seed analyses were carried out using randomly selected propagules, after removing seeds having visible damage. Seed quality of 180 B. humilis individuals from the six largest stands located in northeastern Poland: SOL, JEZ, ROS, MB, GP and SUS situated in the Biebrza National Park (Fig. 1) was studied. Seed samples were obtained from the germination experiment conducted previously (Bona et al. 2019). As authors revealed very low germinability of shrub birch seeds, we have decided to determine the degree of embryo development in unsprouted seeds. Thus, non-germinated shrub birch seeds were examined under a binocular microscope and classified in terms of the degree of embryo development as fully developed (FDE), partially developed (i.e., < 60%; PDE) or empty seeds with no embryo (ES), according to Holm (1994).

Location of sampling sites of shrub birch and tree birches in Poland
To assess B. humilis seed quality, seeds of this species were compared to those of 63 B. pendula and 21 B. pubescens control specimens inhabiting the same peatlands. Seeds from tree birches were used in the germination experiment and embryo development analysis. A total wet mass of 100 seeds per individual was measured with an accuracy of 0.0001 g. One hundred seeds per individual were used to test germination capability, according to the method of Holm (1994).
The seeds from 131 shrub birch individuals and 66 tree birch specimens not used in the germination experiment were used in the analyses of the total phosphate (Pt), organic phosphate (Po) and inorganic phosphate (Pi) contents. Taking into account that birch seeds are very small and light, we were not able to estimate the phosphate contents in individual propagules. Instead, 200 seeds per individual were considered in the analysis, and the concentrations obtained were divided by the seed number to estimate the average phosphate levels per seed. Before the measurements were taken, 200 seeds of each individual were weighed and homogenized in a frozen 10% trichloroacetic acid (TCA) solution in a TissueLyser LT bead mill (Qiagen). The mixture obtained was then used to estimate the Pt, Po and Pi contents in each sample. The Pi content was determined according to the method of Ames (1966) using ascorbic acid, sulfuric acid and ammonium molybdate. To evaluate Pt, the seed extract was incubated in a mixture of 60% perchloric acid and 2 N nitric acid in a ratio of 1:3 at 60 °C for 18 h. The Pt content was then analysed according to the same method as that used for Pi. The P concentrations were analysed using a UV/Vis Spectrophotometer DU730 (Beckman Coulter) at a wavelength of λ = 700 nm. The calculations of Po were performed according to the formula Po = Pt − Pi.
Pollen quality of birches
Detailed laboratory analyses of pollen quality were carried out for six B. humilis stands: SOL, JEZ, ROS, MB, as well as SUS and GRZ located in the Biebrza National Park (Fig. 1). Pollen quality was also tested for samples of tree birches growing in the same peatlands.
Male catkins with mature flowers containing pollen were sampled from 71 B. humilis, 54 B. pendula and 21 B. pubescens individuals. Because the periods of pollen release of the shrub birch and tree birches overlap only partially, male catkins were collected from tree birch specimens twice in the peatlands: once during the maximum pollen release of B. pendula and B. pubescens (MPBp) and again approximately two weeks later, when B. humilis flowers were releasing pollen grains (MPBh). The catkins were collected in paper bags and stored at 4 °C until the analyses were conducted. Alam and Grant (1971) revealed that pollen of birch species stored at 2–5 °C lost its viability about three times slower than pollen stored at room temperature.
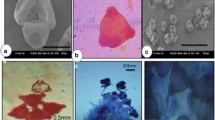
Up to 7 days after male catkins were collected, the pollen quality was tested using two methods: germination experiments and staining with acetocarmine solution. In vitro germination was conducted in a medium containing 2.0% sucrose and 0.8% agar in distilled water, according to the method outlined in Alam and Grant (1971). The pollen was spread evenly on a medium on microscopic slides. The slides were placed in closed Petri dishes with moist filter paper and left in the light at room temperature for 4–5 h to germinate. The slides were analysed under an Olympus CX40 light microscope. A total of 400 pollen grains were counted for each individual to determine the proportion of germinated grains. Pollen grains with tubes longer than half of their own diameter were recognized as having germinated (Fig. 2a; Alam and Grant 1971). The effectiveness of this method was shown previously for other birches: Betula populifolia, Betula cordifolia and Betula papyrifera (Alam and Grant 1971). Another pollen sample from each catkin was stained on microscopic slides with 2% acetocarmine solution, as was previously described in a study of the willow Salix myrtilloides (Pogorzelec et al. 2016). The slides were analysed within 1 h under a light microscope. From each individual, 300 grains were analysed to determine the proportion of stained grains. Fully stained pollen grains were considered viable, while in nonviable pollen cells, the cytoplasm remained transparent (Fig. 2b).

The B. humilis pollen grains tested with two methods: in vitro germination (a) and staining with 2% acetocarmine (b); unstained and stained pollen grains are indicated by the white and black arrows, respectively
Flowering of B. humilis and PO4 3− concentrations in the groundwater
Analyses of the flowering intensities and PO43− concentrations in the groundwater were conducted in the seven B. humilis stands analysed for pollen and/or seed quality as well as in 10 additional populations situated in northern (populations codes: LS, JM, TM, ZYT) and eastern Poland (populations codes: BIK, UU, KB, MOS, KAR, SPL) (Fig. 1). Following the description in Jabłońska (2012), in each B. humilis stand, the habitats were assessed as dry when the water table was more than 10 cm below the peat surface or wet when the water table was less than 10 cm below the peat surface. Betula humilis shoots of a height of 1–2 m were chosen to count inflorescences. Depending on the shrub birch abundance, the total numbers of specimens examined to determine flowering intensity in each population ranged from 8 to 35. In total, male and female catkins were counted on 514 individuals. Depending on the groundwater availability, the groundwater samples were taken manually at a depth of 0–70 cm below peat surface in 3–6 plots evenly distributed across populations. The concentrations of PO43− ions in the groundwater samples were determined spectrophotometrically using ammonium molybdate and ascorbic acid (Hach Company 1992).
Data analyses
Unequal numbers of both tree birch specimens collected in particular locations result from the fact that B. pendula prefers drier localities, while B. pubescens can grow in stands with higher groundwater levels. Since the samples of both tree species did not differ significantly in terms of seed and pollen quality, they were considered in the study as one group. Randomized one-way ANOVA tests with 10,000 randomizations, post hoc tests and the Holm–Bonferroni’s correction were conducted to compare the proportions of stained and germinated pollen grains among the shrub birch populations. The quality of pollen was also analysed among three groups: the shrub birch, tree birches sampled during MPBp, and tree birches sampled during MPBh.
Randomized linear regression (10,000 randomizations) analyses were performed to test the relationships between the different parameters of seed quality (germination ability and the degree of embryo development of non-germinated seeds) and Pi and Po concentrations in the seed sets. The regression analysis was also used to detect a relationship between the seed germinability and seed weights. To visualize results of regressions, the chord diagram plots were created in R software using the circlize package (Gu et al. 2014). Two-sample randomization tests (test statistic—median difference; 10,000 randomizations) were carried out to compare the Pi, Po and Pc concentrations, seed masses, seed germinabilities and embryo developments between shrub birches and tree birches. As the collected shrub birch seeds were significantly heavier than the tree birch seeds, the phosphate concentrations were calculated per seed weight for the interspecific comparison. A randomized one-way ANOVA test was conducted to compare the above parameters among the shrub birch populations.
Randomized linear regression was carried out to determine whether the median values of male, female and total inflorescences were related to the concentrations of PO43− ions in the groundwater. A randomized one-way ANOVA test was performed to compare the concentrations of groundwater PO43− ions, the number of male and female catkins and the total number of catkins among the B. humilis populations. These parameters were also analysed between stands with high and low groundwater levels using two-sample randomization tests. All statistical analyses were performed in Rundom Pro 3.14 (Jadwiszczak 2009).
Results
Seed quality
Results of seed quality analyses, i.e., percent of germinated seeds (GS), seeds with fully developed embryo (FDE), seeds with partially developed embryo (PDE), empty seeds with no embryo (ES), and mean phosphate concentrations per seed in shrub birch and tree birches populations are given in Online Resource 1. The linear regression analyses showed a significant, albeit weak negative relationship between the Pi concentration in the B. humilis seeds and PDE seeds (R2 = 0.061, b = − 32.69, P = 0.002; Fig. 3a). In the tree birch seeds, a positive relationship was found between Pi and GS (R2 = 0.197, b = 58.74, P = 0.0005) as well as between Po and GS (R2 = 0.221, b = 25.08, P = 0.0003; Fig. 3b). In turn, the relationship between Po and ES was significantly negative (R2 = 0.185, b = − 29.53, P = 0.0005). Shrub birch seed mass was positively correlated with GS (R2 = 0.096, b = 404.02, P = 0.0002). ANOVA showed that two shrub birch population pairs only (ROS and SUS, JEZ and ROS) were differentiated in terms of the Po contents in their seeds (F = 4.181, P = 0.0014), while the differences in Pi (F = 3.324, P = 0.0077) and Pt (F = 3.671, P = 0.0037) were nonsignificant between populations after Holm–Bonferroni’s correction (Table 1 and Online Resource 2). The differences in seed quality (seed mass, FDE, PDE and ES) were significant between B. humilis populations (Table 1 and Online Resource 2). Two-sample randomization tests revealed that the shrub birch populations were characterized by higher Po (median difference = 0.150, P = 0.0002) and Pt concentrations (median difference = 0.177, P = 0.0001) in their seeds, higher seed masses (median difference = 0.004, P = 0.0001), lower germination abilities (median difference = − 5, P = 0.0001), higher numbers of seeds with partially developed embryos (median difference = 32, P = 0.0001) and lower numbers of seeds with no embryos (median difference = − 19, P = 0.0002) compared to the control samples of tree birches (Fig. 4).

Chord diagrams showing the results of regression analyses between organic phosphate (Po) contents in seeds and the seeds quality parameters (green colour) and inorganic phosphate (Pi) contents in seeds and the seeds quality parameters (violet colour) in (a) B. humilis and (b) tree birches. Variables included in the analyses: GS number of germinated seeds per individual, FDE number of seeds with fully developed embryo, PDE number of seeds with partially developed embryo, ES number of empty seeds. Width of the arc connecting the variables reflects the value of the coefficient of determination (R2), according to the numerical scale. Asterisks indicate significant relationships after Holm–Bonferroni’s correction

Box plots showing differences in seed quality parameters (P contents in seeds, seed mass and the degree of embryo development) between the shrub birch (Bh) and tree birches (Bp). Pi inorganic phosphate, Po organic phosphate, Pt total phosphate, GS germinated seeds per individual, FDE seeds with fully developed embryo, PDE seeds with partially developed embryo, ES empty seeds. The upper and lower whiskers show the maximum and minimum values, respectively. The ends of the boxes are on the first and third quartiles. The horizontal lines in the boxes show the median values. Different letters indicate significant differences between groups revealed by two-sample randomization test after Holm–Bonferroni’s correction
Pollen quality
Among the B. humilis populations, the differences in pollen grain stainability and germinability were nonsignificant between sites after Holm–Bonferroni’s correction was applied (F = 0.924, P = 0.459 and F = 3.096, P = 0.013, respectively; Table 1). Despite the very high stainability of B. humilis pollen (95.5%) as well as that of the tree birch pollen collected during the maximum pollen release of B. pendula and B. pubescens (97.9%) and the maximum pollen release of B. humilis (96.3%), the pollen germination ability was low in all the groups studied (1.6%, 32.5% and 5.1%, respectively; Online Resource 3).
ANOVA revealed that pollen stainability did not differ among the B. humilis and tree birch samples collected during the two flowering periods (F = 0.38, P = 0.695). However, the differences in pollen germinability among the B. humilis and tree birch samples collected during the two flowering periods were significant (F = 171.34, P = 0.0001; Fig. 5). Post hoc tests showed that the tree birch pollen germination ability was significantly higher when collected during MPBp compared to the pollen that was collected later during MPBh (Padj = 0.0003). The number of germinated pollen grains was also significantly higher in the tree birch samples collected during MPBp and MPBh than in the collected B. humilis pollen (Padj = 0.0002 and Padj = 0.0001, respectively).

Box plot showing differences in the number of germinated pollen grains of B. humilis (Bh) and tree birches (Bp) collected during maximum pollen release of B. humilis (MPBh) and maximum pollen release of tree birches (MPBp). The upper and lower whiskers show the maximum and minimum values, respectively. The ends of the boxes are on the first and third quartiles. The horizontal lines in the boxes show the median values. Different letters indicate significant differences between groups (ANOVA; P < 0.05)
Flowering and P concentration in the groundwater
Median values of the male, female and total numbers of inflorescences per individual as well as the median PO43− concentrations in groundwater in the B. humilis populations are given in Online Resource 4. Very limited flowering was found in the most northern ZYT population, where many shrubs did not flower at all or only had few inflorescences (median number of total catkins per individual = 2). Flowering was the most intense in the KAR locality (72.5); however, only eight specimens were found at this site. ANOVA revealed that the number of male, female and total catkins per individual differed significantly between populations (F = 7.824, P = 0.0001; F = 4.133, P = 0.0001; F = 8.857, P = 0.0001, respectively; Table 1 and Online Resource 5).
Two populations, ROS and ZYT, were characterized by very high concentrations of PO43− ions in the groundwater (3.574 mg l−1 and 2.616 mg l−1, respectively; Online Resource 4). Nevertheless, the differences in the PO43− concentrations among stands were nonsignificant after applying Holm–Bonferroni’s correction (F = 16.493, P = 0.0001; Table 1). The regression analysis showed no relationships between the PO43− concentrations in groundwater and the medians of male, female or total inflorescences per individual in the studied B. humilis populations (R2 = 0.027, P = 0.552; R2 = 0.014, P = 0.660; R2 = 0.021, P = 0.592, respectively). There were no differences in the contents of PO43− ions between stands with high and low groundwater levels (median difference = − 0.0405, P = 0.253). Significantly higher numbers of male (median difference = 11, P = 0.0007) and total inflorescences (median difference = 17, P = 0.0001) were found in shrub birch stands characterized by high groundwater levels compared to drier locations (Fig. 6).

Box plot showing differences in the number of B. humilis inflorescences between sites with high (HWL) and low groundwater levels (LWL). The upper and lower whiskers show the maximum and minimum values, respectively. The ends of the boxes are on the first and third quartiles. The horizontal lines in the boxes show the median values. Different letters indicate significant differences between groups revealed by two-sample randomization test after Holm–Bonferroni’s correction
Discussion
Pollen and seed quality
In the present study, we revealed mean values of 44.8% empty seeds (ES) and 47.3% seeds with partially developed embryos (PDE) per B. humilis population. Thus, the substantial proportion of non-vital propagules seems to be directly responsible for the previously reported low reproductive success of the shrub birch populations (Chrzanowska et al. 2016; Bona et al. 2019), and this observation is in agreement with our hypothesis. Chrzanowska et al. (2016) found a significantly lower number of sprouted seeds in the Polish marginal localities of shrub birch compared to the subcentral stands situated in Belarus. A few years later, the germination ability of shrub birch seeds collected from six populations in northeastern Poland was even worse (Bona et al. 2019). We found that the germination ability of the B. humilis seeds was significantly lower than that of the tree birches. The production of seeds with no embryos is not a rare phenomenon in the birch taxa. For example, the median numbers of empty seeds in B. nana populations have been measured as ranging from 86 to 99 per 100 seeds (Jadwiszczak et al. 2017). Seeds without embryos constituted 57.6% of the B. pendula seed set and 58.5% of B. pubescens seed set in northern Sweden (Holm 1994). Empty seeds are often produced by birches in the absence of pollination and under circumstances of embryo developmental arrest (Atkinson 1992; de Groot et al. 1997).
Lack of effective pollination can be a consequence of low pollen quality and/or quantity (pollen limitation). We used two methods to study B. humilis pollen quality: staining with acetocarmine solution and germination assays (Alam and Grant 1971). The results obtained were quite different between the two methods: 95.5% of the shrub birch pollen grains were considered viable based on the intensive colour of the protoplasts, but only 1.6% of cells germinated in vitro. Based on various studies determining pollen quality, Dafni and Firmage (2000) reported that pollen staining could sometimes overestimate pollen viability. As the stained protoplasts do not indicate the ability of shrub birch pollen to germinate, the germination capacity of pollen grains tested in vitro turned out to be a better indicator of pollen quality than pollen staining; thus, we made conclusions about the quality of the pollen from the results of the in vitro germination experiment exclusively. Among the B. humilis populations, the ability of pollen to germinate was seen in 0.8% of the pollen grains in ROS up to only 3.1% in the JEZ. Therefore, we assume that significant pollen limitation is a highly probable cause of the production of substantial numbers of empty seeds. Reduced sexual reproduction efficiency caused by quantitative and qualitative pollen limitations has been observed in numerous plant populations (Byers 1995; Nagamitsu et al. 2006; Amat et al. 2011).
An insufficient amount of viable pollen of a particular species increases the risk of that species being pollinated by the abundant pollen of closely related species. The likelihood of interspecific crosses significantly increases in small populations where the chance of pollination within the same species is greatly reduced (Rhymer and Simberloff 1996; Knight et al. 2005). Our study showed that the germination ability of B. humilis pollen grains was significantly lower than that of the tree birches analysed during both terms: maximum pollen release of B. humilis and maximum pollen release of tree birches. This observation is congruent with our hypothesis; however, it seems to be especially distressing because it suggests that even during the mid-flowering time of the endangered B. humilis, its pollen quality is lower than the pollen quality of two common tree birches, B. pendula and B. pubescens, growing in the same area. This phenomenon can increase the probability of interspecific pollination and potentially can result in the formation of hybrid progeny. The analysis of amplified fragment length polymorphism (AFLP) markers in the B. humilis populations located at the southwestern margin of the species distribution revealed three specimens of hybrid origin, providing evidence of the existence of an incomplete reproductive barrier between B. humilis and its tree relatives (Bona et al. 2018). As three hybrid specimens were found in dry habitats, the authors implied that deteriorating environmental conditions could increase the number of interspecific crosses.
If the parental taxa have incompatible genomes, then one of the outcomes of interspecific hybridisation is arrested embryo development (Zenkteler 1990; Lester and Kang 1998; Oneal et al. 2016). In our study, the number of seeds with partially developed embryos in the shrub birch samples (mean PDE = 47.3%) was approximately twice as high as that in the tree birches (mean PDE = 24.4%). Thus, it cannot be excluded that the co-occurrence of different birch species in peatlands can generate seeds with nonviable embryos as a result of interspecific gene exchange.
Self-pollination of usually outcrossing species may also prevent the proper development of embryos (Bellusci et al. 2009; Liao et al. 2014). Most birch species are self-incompatible (Atkinson 1992; de Groot et al. 1997); thus, self-pollination could significantly decrease their reproductive success. Moreover, the self-pollination rate is higher in clonal plants representing clustered growth types, in which the mixing of shoots of different clones is highly limited (Barrett 2015). In the studied shrub birch populations, clonal growth was common, and most neighbouring specimens shared the same genotype (Bona et al. 2019). However, Bona et al. (2019) denied that clonality was the main reason for seed production failure, as seed germinability was similarly low in populations representing different clonal propagation patterns. Nevertheless, the self-pollination rate and the efficiency of the sexual reproduction of B. humilis by selfing have not been studied thus far.
Environmental conditions shaping sexual reproduction parameters
High phosphate availability in the environment often positively affects the sexual reproduction of plants (e.g., Lau and Stephenson 1994; Wang et al. 2011). However, the endangered species very often occur at P-poor sites and limit investment in the sexual reproduction in the temperate regions (Fujita et al. 2014). For example, they can produce a small number of seeds but the P concentration per seed is high (Fujita et al. 2014). Taking into account the way the phosphate concentrations were calculated per seed in the B. humilis specimens as well as control samples of B. pendula and B. pubescens, all relationships between parameters of shrub birch seed quality (GS, FDE, PDE, ES) and Pi or Po were nonsignificant or very weak. Hence, our hypothesis concerning the positive relationships between seed P and seed germination capacity in the B. humilis individuals was not confirmed. However, the relationships between Pi and GS as well as Po and GS were significantly positive, whereas significant and negative relationship was found between Po and ES in the tree birches. It seems that P contents did not affect shrub birch seed quality but could increase tree birch seed quality and simultaneously enhance their competitive strength against B. humilis. Shrub birch seeds had significantly higher Po concentrations per seed mass than tree birches. High P reserves allow seedlings to establish faster and accelerate seedling growth (Zhu and Smith 2001; White and Veneklaas 2012), which are of great importance in poorly competitive species. However, the studied tree birch seeds still showed better germination abilities than did the B. humilis seeds. Thus, as P concentrations in silver and downy birches seeds explained up to 22.1% of variability, P contents were not a main factor affecting the seed germinability and successful seedling establishment in peatlands.
Our study revealed that the numbers of male, female, and total inflorescences of the shrub birch were not related to P availability in the groundwater. The studied shrub birch populations differed significantly in terms of their reproductive output parameters, i.e., flowering and seed quality, but it seems that groundwater P availability was not a limiting factor in the B. humilis stands. For example, the MOS population, which had the lowest concentration of PO43− ions in the groundwater among all sites (0.031 mg l−1), was characterized by an above-average median number of male inflorescences and the highest median number of female inflorescences (30 and 15.5, respectively). At the ROS site, the groundwater concentration of PO43− ions was the highest among all sites (3.574 mg l−1). Although the flowering intensity of B. humilis was quite abundant here, the seed germination was very low (GS = 1.7%; Bona et al. 2019). Highest numbers of germinated seeds were noted in MB and SOL populations, where PO43− concentrations were 0.159 and 0.145 mg l−1, respectively. The above outcomes allow us to definitively reject the hypothesis that the shrub birch flowering intensity may be related to PO43− concentrations in groundwater and to decisively confirm the remark of Chrzanowska et al. (2016). In general, our observations are congruent with those presented by Serafin et al. (2018), who revealed that the B. humilis specimens functioned well when the total P concentrations in the water ranged from 0.08 to 0.32 mg l−1 and when PO43− = 0.1 mg l−1. However, as P is the limiting macronutrient in many ecosystems and its availability is relatively low and is rarely adequate for optimal growth (Abel et al. 2002; Elser et al. 2007), we cannot exclude that soil P enrichment could positively affect the growth and reproduction of shrub birch. On the other hand, fertilization of peatlands can cause invasions of non-peatland species, such as Betula trees (Sottocornola et al. 2007; Serafin et al. 2018). This would be particularly detrimental to the preservation of shrub birch populations; therefore, these practices are not recommended.
As B. humilis is a poor competitor in the western and central European peatlands, high water levels may be beneficial for this species (Jabłońska 2006, 2012). We found that groundwater levels had a significant impact on the formation of the male inflorescences. This result was surprising, as Chrzanowska et al. (2016) showed no relationship between the groundwater table and the flowering intensity of shrub birch. Nevertheless, our research, which was conducted on a larger sample set, indicated that draining mires can negatively affect flowering and, consequently, forming new generations. Chrzanowska et al. (2016) also suggested that sprouting shrub birch seeds could be more successful in flooded habitats, as seeds from sites with higher water levels had higher masses, and heavier seeds showed better germination ability. Our study confirmed the positive relationship between the seed mass and germination ability of B. humilis. Given the above information, we conclude that the protection of peatlands to maintain the current habitats of relict shrub birch populations is of great importance for the preservation of the species.
Conclusions
In this study, we investigated the reproductive performance of B. humilis in peatland stands to indicate reasons for low reproductive efficiency at the end of the species range. The pollen and seed qualities of the shrub birches were compared with those of the sympatrically growing tree birches B. pendula and B. pubescens. The shrub birch seed quality analysis showed that the direct cause of the reduced seed germinability reported in previous studies (Chrzanowska et al. 2016; Bona et al. 2019) was a small share of seeds containing fully developed embryos. Our study revealed very low-quality shrub birch pollen, and we consider this to be the main reason for the lack of embryos in seeds. Moreover, as the germination ability of the studied shrub birch pollen grains was significantly lower than that of the pollen of the close congeners, interspecific pollination caused by predominant pollen of B. pendula and B. pubescens cannot be excluded.
The study did not confirm the hypothesis of Chrzanowska et al. (2016) concerning the positive relationship between the seed germination capacity and P accumulation in seeds in the B. humilis individuals. In contrast, despite smaller overall P concentrations in the studied B. pendula and B. pubescens seeds compared to the shrub birch seeds, Po and Pi were positively related to the number of germinated seeds in tree birches. The availability of PO43− ions in the groundwater did not affect the observed number of shrub birch inflorescences. However, the numbers of male inflorescences were significantly higher in localities with high groundwater levels than in drier stands. This observation confirms the importance of water abundance in shrub birch stands. High water levels limit the overgrowth of B. humilis stands by other plants, including closely related tree birches, which reduces the risk of interspecific mating and facilitates the preservation of populations of this poorly competitive species.
Author contribution statement
AB and KAJ designed the study, analysed the data and wrote the manuscript. SK revised the manuscript. AB, SK and KAJ carried out the fieldwork. AB, SK, KAJ and GP performed the laboratory work. All authors read and approved the final manuscript.
Availability of data and materials
The datasets analysed during the current study are available from the corresponding author on request.
References
Abel S, Ticconi CA, Delatorre CA (2002) Phosphate sensing in higher plants. Physiol Plant 115(1):1–8. https://doi.org/10.1034/j.1399-3054.2002.1150101.x
Alam MT, Grant WF (1971) Pollen longevity in birch (Betula). Can J Bot 49:797–798. https://doi.org/10.1139/b71-119
Amat ME, Vargas P, Gómez JM (2011) Pollen quality limitation in the Iberian critically endangered genus Pseudomisopates (Antirrhinaceae). Plant Ecol 212(6):1069–1078. https://doi.org/10.1007/s11258-010-9887-6
Ames BN (1966) Assay of inorganic phosphate, total phosphate and phosphatases. Methods Enzymol 8:115–118. https://doi.org/10.1016/0076-6879(66)08014-5
Atkinson MD (1992) Betula pendula Roth (B. verrucosa Ehrh.) and B. pubescens Ehrh. J Ecol 80:837–870. https://doi.org/10.2307/2260870
Balogh CM, Barrett SCH (2018) The influence of floral morph ratios and low plant density on mating and fertility in a tristylous colonizing species. Botany 96:533–545. https://doi.org/10.1139/cjb-2018-0032
Barrett SCH (2015) Influences of clonality on plant sexual reproduction. PNAS 112:8859–8866. https://doi.org/10.1073/pnas.1501712112
Bellusci F, Pellegrino G, Musacchio A (2009) Different levels of inbreeding depression between outcrossing and selfing Serapias species. Biol Plant 53(1):175–178. https://doi.org/10.1007/s10535-009-0029-8
Bona A, Petrova G, Jadwiszczak KA (2018) Unfavourable habitat conditions can facilitate hybridisation between the endangered Betula humilis and its widespread relatives B. pendula and B. pubescens. Plant Ecol Divers 11:295–306. https://doi.org/10.1080/17550874.2018.1518497
Bona A, Kulesza U, Jadwiszczak KA (2019) Clonal diversity, gene flow and seed production in endangered populations of Betula humilis Schrk. Tree Genet Genomes 15:50. https://doi.org/10.1007/s11295-019-1357-2
Borges PAV, Gabriel R, Fattorini S (2019) Biodiversity erosion: causes and consequences. In: Leal Filho W, Azul A, Brandli L, Özuyar P, Wall T (eds) Life on Land. Encyclopedia of the UN Sustainable Development Goals. Springer, Cham. https://doi.org/10.1007/978-3-319-71065-5_78-1
Byers DL (1995) Pollen quantity and quality as explanations for low seed set in small populations exemplified by Eupatorium (Asteraceae). Am J Bot 82(8):1000–1006. https://doi.org/10.1002/j.1537-2197.1995.tb11564.x
Chrzanowska A, Jadwiszczak KA, Kłosowski S, Banaszek A, Sozinov OV (2016) Sexual reproduction efficiency and genetic diversity of endangered Betula humilis Schrk. populations from edge and sub-central parts of its range. Folia Geobot 51:161–173. https://doi.org/10.1007/s12224-016-9244-1
Dafni A, Firmage D (2000) Pollen viability and longevity: practical, ecological and evolutionary implications. Plant Syst Evol 222:113–132. https://doi.org/10.1007/BF00984098
de Groot W, Thomas P, Wein RW (1997) Betula nana L. and Betula glandulosa Michx. J Ecol 85:241–264. https://doi.org/10.2307/2960655
Ellstrand NC, Elam DR (1993) Population genetic consequences of small population size: implications for plant conservation. Annu Rev Ecol Syst 24:217–242. https://doi.org/10.1146/annurev.es.24.110193.001245
Elser JJ, Bracken ME, Cleland EE, Gruner DS, Harpole WS, Hillebrand H, Ngai JT, Seabloom EW, Shurin JB, Smith JE (2007) Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol Lett 10(12):1135–1142. https://doi.org/10.1111/j.1461-0248.2007.01113.x
Franiel I (2012) The biology and ecology of Betula pendula Roth on post-industrial waste dumping grounds: the variability range of life history traits. Dissertation, University of Silesia, Poland
Fujita Y, Venterink HO, van Bodegom PM, Douma JC, Heil GW, Hölzel N, Jabłońska E, Kotowski W, Okruszko T, Pawlikowski P, de Ruiter P, Wassen MJ (2014) Low investment in sexual reproduction threatens plants adapted to phosphorus limitation. Nature 505:82–86. https://doi.org/10.1038/nature12733
Gaitán-Espitia JD, Hobday AJ (2021) Evolutionary principles and genetic considerations for guiding conservation interventions under climate change. Glob Chang Biol 27:475–488. https://doi.org/10.1111/gcb.15359
Gu Z, Gu L, Eil SR, Schlesner M, Brors B (2014) Circlize implements and enhances circular visualization in R. Bioinformatics 30:2811–2812. https://doi.org/10.1093/bioinformatics/btu393
Hach Company (1992) Hach water analysis handbook. Hach Co., Loveland, p 831
Hamrick JL, Godt MJW (1996) Effects of life-history traits on genetic diversity in plants. Philos Trans R Soc Lond B Biol Sci 351:1291–1298. https://doi.org/10.1098/rstb.1996.0112
Holm S-O (1994) Reproductive patterns of Betula pendula and B. pubescens coll. along a regional altitudinal gradient in northern Sweden. Ecography 17:60–72. https://doi.org/10.1111/j.1600-0587.1994.tb00077.x
Jabłońska E (2006) Comparison of habitat conditions at Betula humilis sites in north-eastern and south-eastern Poland. Polish J Environ Stud 15(5d):181–187
Jabłońska E (2012) Vegetation with Betula humilis in Central Europe. Phytocoenologia 42:259–277. https://doi.org/10.1127/0340-269X/2012/0042-0527
Jadwiszczak P (2009) Rundom Pro 3.14. Software for classical and computer-intensive statistics. http://pjadw.tripod.com. Accessed 7 November 2019
Jadwiszczak KA, Kłosowski S, Zalewska I, Banaszek A, Chrzanowska A (2017) Genetic diversity and sexual reproduction in relict populations of Betula nana. Silva Fennica 51:5643. https://doi.org/10.14214/sf.5643
Jump AS, Woodward FI (2003) Seed production and population density decline approaching the range-edge of Cirsium species. New Phytol 160:349–358. https://doi.org/10.1046/j.1469-8137.2003.00873.x
Knight TM, Steets JA, Vamosi JC, Mazer SJ, Burd M, Campbell DR, Dudash MR, Johnstion MO, Mitchell RJ, Ashman TL (2005) Pollen limitation of plant reproduction: pattern and process. Annu Rev Ecol Evol Syst 36:467–497. https://doi.org/10.1146/annurev.ecolsys.36.102403.115320
Lau TC, Stephenson AG (1994) Effects of soil phosphorus on pollen production, pollen size, pollen phosphorus content, and the ability to sire seeds in Cucurbita pepo (Cucurbitaceae). Sexual Plant Reprod 7:215–220. https://doi.org/10.1007/BF00232740
Lester RN, Kang JH (1998) Embryo and endosperm function and failure in Solanum species and hybrids. Ann Bot 82(4):445–453. https://doi.org/10.1006/anbo.1998.0695
Liao T, Yuan D-Y, Zou F, Gao C, Yang Y, Zhang L, Tan X-F (2014) Self-sterility in Camellia oleifera may be due to the prezygotic late-acting self-incompatibility. PLoS ONE 9(6):e99639. https://doi.org/10.1371/journal.pone.0099639
Nagamitsu T, Kawahara T, Kanazashi A (2006) Pollen-limited production of viable seeds in an endemic dwarf birch, Betula apoiensis, and incomplete reproductive barriers to a sympatric congener, B. ermanii. Biol Conserv 129:91–99. https://doi.org/10.1016/j.biocon.2005.10.025
Nord EA, Shea K, Lynch JP (2011) Optimizing reproductive phenology in a two-resource world: a dynamic allocation model of plant growth predicts later reproduction in phosphorus-limited plants. Ann Bot 108:391–404. https://doi.org/10.1093/aob/mcr143
Oneal E, Willis JH, Franks RG (2016) Disruption of endosperm development is a major cause of hybrid seed inviability between Mimulus guttatus and Mimulus nudatus. New Phytol 210(3):1107–1120. https://doi.org/10.1111/nph.13842
Pogorzelec M, Serafin A, Banach-Albińska B, Szczurowska A, Parzymies M, Bronowicka-Mielniczuk U (2016) Pollen viability of Salix myrtilloides L.—an endangered species in Poland. Acta Agrobot 69:1679–1688. https://doi.org/10.5586/aa.1679
Rasmussen KK, Kollmann J (2004) Poor sexual reproduction on the distribution limit of the rare tree Sorbus torminalis. Acta Oecol 25:211–218. https://doi.org/10.1016/j.actao.2004.02.001
Rhymer JM, Simberloff D (1996) Extinction by hybridization and introgression. Ann Rev Ecol Evol Syst 27(1):83–109. https://doi.org/10.1146/annurev.ecolsys.27.1.83
Schwartz BU, Poschlod P (2015) Die Letzten ihrer Art in Bayern—Das Eiszeitrelikt Zwergbirke (Betula nana L.). Eine Bestandsanalyse mit biologisch-ökologischen Untersuchungen. [The last of its kind in Bavaria—the ice age relict Betula nana. Results of biological, ecological, and population viability studies]. Anliegen Nat 37(1):19–30
Serafin A, Urban D, Bronowicka-Mielniczuk U, Szczurowska A (2018) To what degree can the specifics of occurrence of glacial relic Betula humilis Schrank be an indicator of habitat conditions of moderate climate peatlands? Water 10(8):1062. https://doi.org/10.3390/w10081062
Shivanna KR, Tandon R (2014) Reproductive ecology of flowering plants: a manual. Springer, New Delhi, p 178
Sottocornola M, Boudreau S, Rochefort L (2007) Peat bog restoration: effect of phosphorus on plant re-establishment. Ecol Eng 31(1):29–40. https://doi.org/10.1016/j.ecoleng.2007.05.001
Tormo Molina R, Muñoz Rodríguez A, Palaciso IS, López FG (1996) Pollen production in anemophilous trees. Grana 35(1):38–46. https://doi.org/10.1080/00173139609430499
Wang S, Jiang J, Li T, Li H, Wang C, Wang Y, Liu G (2011) Influence of nitrogen, phosphorus, and potassium fertilization on flowering and expression of flowering-associated genes in white birch (Betula platyphylla Suk.). Plant Mol Biol Rep 29:794–801. https://doi.org/10.1007/s11105-010-0281-2
Wassen MJ, Olde Venterink H, Lapshina ED, Tanneberger F (2005) Endangered plants persist under phosphorus limitation. Nature 437(7058):547–550. https://doi.org/10.1038/nature03950
White PJ, Veneklaas EJ (2012) Nature and nurture: the importance of seed phosphorus content. Plant Soil 357:1–8. https://doi.org/10.1007/s11104-012-1128-4
Załuski T, Jabłońska E, Pawlikowski P, Pisarek W, Kucharczyk M (2014) Betula humilis Schrank. In: Kaźmierczakowa R, Zarzycki K, Mirek Z (eds) Polska czerwona księga roślin (Polish plant red book). W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków, pp 92–95
Zenkteler M (1990) In vitro fertilization and wide hybridization in higher plants. Crit Rev Plant Sci 9:267–279. https://doi.org/10.1080/07352689009382290
Zhu YG, Smith SE (2001) Seed phosphorus (P) content affects growth, and P uptake of wheat plants and their association with arbuscular mycorrhizal (AM) fungi. Plant Soil 231(1):105–112. https://doi.org/10.1023/A:1010320903592
Funding
This work was supported by the Polish National Science Centre under the Grant no. 2016/23/N/NZ8/03054.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by DesRochers.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bona, A., Kłosowski, S., Jadwiszczak, K.A. et al. Flowering and quality of seeds and pollen in endangered populations of Betula humilis. Trees 36, 313–324 (2022). https://doi.org/10.1007/s00468-021-02207-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-021-02207-7