
Abstract
Managed and wild pollinators often cohabit in both managed and natural ecosystems. The western honeybee, Apis mellifera, is the most widespread managed pollinator species. Due to its density and behaviour, it can potentially influence the foraging activity of wild pollinators, but the strength and direction of this effect are often context-dependent. Here, we observed plant–pollinator interactions in 51 grasslands, and we measured functional traits of both plants and pollinators. Using a multi-model inference approach, we explored the effects of honeybee abundance, temperature, plant functional diversity, and trait similarity between wild pollinators and the honeybee on the resource overlap between wild pollinators and the honeybee. Resource overlap decreased with increasing honeybee abundance only in plant communities with high functional diversity, suggesting a potential diet shift of wild pollinators in areas with a high variability of flower morphologies. Moreover, resource overlap increased with increasing trait similarity between wild pollinators and the honeybee. In particular, central-place foragers of family Apidae with proboscis length similar to the honeybee exhibited the highest resource overlap. Our results underline the importance of promoting functional diversity of plant communities to support wild pollinators in areas with a high density of honeybee hives. Moreover, greater attention should be paid to areas where pollinators possess functional traits similar to the honeybee, as they are expected to be more prone to potential competition with this species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
As a managed and super-generalist pollinator, the western honeybee, Apis mellifera Linnaeus, plays a fundamental role in the pollination of both crops (Garibaldi et al. 2013) and wild plants (Hung et al. 2018). However, managed honeybees might adversely impact wild pollinator communities, as they are often extremely abundant, have a prolonged flight season, and tend to forage on the most abundant and rewarding floral resources (Goulson 2003). Nevertheless, observed effects are often idiosyncratic and seem to depend on local conditions, on the composition of wild pollinator communities, and on the different methodological approaches adopted (Goulson 2003; Cane and Tepedino 2017; Mallinger et al. 2017).
Against this background, functional traits of both plants and pollinators can help to identify the likelihood, strength and direction of the interactions between managed and wild pollinators (Violle et al. 2007; Eklöf et al. 2013; Schleuning et al. 2015; Bergamo et al. 2020). Floral morphological traits are fundamental in shaping plant–pollinator interactions (Junker et al. 2013). Plant species with greater flower size and longer flowering periods are usually more generalist, being attractive to many pollinator species, while flowers with deep corolla are usually accessible only to a few specialized pollinator species (Lázaro et al. 2020). Although the effect of functional diversity of plant communities on pollinators is still debated (Fornoff et al. 2017; Uyttenbroeck et al. 2017; Goulnik et al. 2020), one expectation is that increased functional diversity should reduce the plant resource overlap between wild pollinators and a dominant species such as the honeybee by providing a larger number of alternative nectar and pollen resources (Fig. 1).

Expected effects of functional diversity of plant community and trait similarity between wild pollinator community and the honeybee on plant–pollinator interactions. We hypothesise that: a in sites with a low functional diversity of plant community and a low trait similarity between wild pollinator community and the honeybee, the resource overlap between wild pollinators and the honeybee would be generally low, as pollinator species with functional traits different from those of the honeybee would exploit different resources; b in sites with a high functional diversity of plant community and a low trait similarity between wild pollinator community and the honeybee, the resource overlap would be even lower, as pollinator species would spread on different floral resources; c in sites with a low functional diversity of plant community and a high trait similarity between wild pollinator community and the honeybee, pollinator species would share an important portion of plants with the honeybee, therefore, resulting in a high resource overlap; d in sites with a high functional diversity of plant community and a high trait similarity between wild pollinator community and the honeybee, the resource overlap would decrease, as pollinator species would have much more resources to forage on. Increasing honeybee abundance and higher temperatures would intensify the observed effects
Similarly, pollinator traits can affect both how pollinators interact with plant species and how they interact with each other (Albrecht et al. 2012; Garibaldi et al. 2015; Woodcock et al. 2019). In particular, the competition of wild pollinators with honeybees in areas with a high abundance of managed pollinators could be stronger for central-place foragers, which are forced to collect pollen and nectar near to their nest (Walther-Hellwig et al. 2006), and for oligolectic pollinators, which have a limited ability to shift to alternative resources (Cane and Tepedino 2017). On the contrary, large-sized pollinators with longer proboscis usually have a larger diet breadth, as they are able to exploit a wider range of resources compared to smaller ones (Greenleaf et al. 2007; Lara-Romero et al. 2019). Hence, we expect that a high trait similarity between wild pollinators and the honeybee should increase their resource overlap (Fig. 1).
Environmental variables can also have a strong effect on species phenology and behaviour. Air temperature and weather, in particular, modulate the activity of pollinators (Trøjelsgaard and Olesen 2013; Giannini et al. 2015). For example, bumblebees are often active at low temperatures and under unfavourable weather conditions (Goulson 2010), while butterflies are strongly negatively affected by low air temperatures (Abrahamczyk et al. 2011). Honeybees are more sensitive to low temperatures than many wild pollinators (Jaffé et al. 2010), so potential competition between wild pollinators and honeybees should be more severe at high temperatures (Fig. 1).
A promising approach to elucidate potential mechanisms shaping the interactions between plants and pollinators is the use of network tools integrated with functional trait analysis. Here, we investigated how functional traits of both plants and pollinators, together with the abundance of honeybees and temperature, affected the foraging behaviour of wild pollinators. In particular, this study aimed to explore how functional richness and dispersion of plant communities influenced the resource overlap between wild pollinators and the honeybee, also testing the effect of trait similarity between wild pollinators and the honeybee. We observed plant–pollinator networks in 51 grasslands in Northern Italy and computed the resource overlap between each wild pollinator species and the honeybee. We calculated functional richness and functional dispersion of plant communities using flower corolla length, flower colour, and flower shape, while trait similarity between wild pollinators and the honeybee was calculated using proboscis length, body size, type of foraging range, and taxonomic family.
Materials and methods
Sampling design
Fieldwork was carried out in 51 grasslands in Northern Italy (Alps and Prealps), approximately 50 × 30 m in size. Grasslands were selected across a steep elevational gradient ranging from 150 to 2100 m a.s.l., and had a wide range of honeybee abundance (Table S1, Fig. S1). The selection of the sites was adjusted during the sampling season to have statistical independence between temperature and honeybee abundance (Pearson’s correlation = 0.11, p value = 0.41). Each sampling site was at least 0.53 km from the nearest one (mean = 4.60 km). We were not able to determine the exact number of beehives near the sampling sites, but the mean density in the study area was c. 5 beehives per km2 (data provided by the National Data Bank of the Zootechnical Registry established by the Ministry of Health at the National Service Centre of the “G. Caporale” Institute of Teramo).
Sampling of ecological interactions
Between May and September 2019, we observed plant–pollinator interactions in the selected sites. Sampling occurred between 9:00 and 17:00 only with air temperature > 15 °C, low wind, no rain, and cloud coverage < 70%. Each site was visited only once. At each site, we identified all flowering plant species and assessed their relative abundance. All the individuals of each plant species were then observed for 15 min in total, during which all hymenopterans and dipterans touching the reproductive parts of flowers were counted and collected. Both plant and pollinator species were identified in the field when possible, otherwise, plants were collected and prepared in a herbarium, while pollinators were placed in vials filled with 70% ethanol. Later identification was entrusted to experts (Filippo Prosser and LM for plants, and AC, MM, DP, and PC for pollinators). During the sampling, we also measured the air temperature using a Tinytag Plus 2 TGP-4017 data logger.
Resource overlap between wild pollinators and the honeybee
Starting from the observed interactions, we built 51 bipartite plant–pollinator networks, one for each sampling site. For each network, we calculated the resource overlap between each wild pollinator species (i.e., excluding the honeybee) and the honeybee using Morisita’s index (Morisita 1959) in the R package spaa (Zhang 2016). The index ranges from 0 to 1, with increasing values indicating an increase in the plant resource overlap between the two pollinator species. In each network, we then calculated the community weighted mean (hereafter, CWM) resource overlap between wild pollinators and the honeybee as the mean resource overlap value of all wild pollinator species weighted by their abundance. We used CWM resource overlap instead of resource overlap values of single species as no model using species as replicates met statistical assumptions, even after changing the distribution or transforming the variables. All analyses were performed using R version 3.6.1 (R Core Team 2019).
Functional traits of plant species
For each flowering plant species, we measured flower corolla length with a calliper, and recorded flower type after Kugler (1970) and flower colour (Table S2). These are among the most important morphological traits for the definition of pollinator feeding niches: flower colour affects the attractiveness and selectivity of flowers, while flower type and corolla length determine the accessibility of flowers to pollinators (Junker et al. 2013). We then calculated two indices of functional diversity of plant communities for each network, i.e., the standardized functional richness and the functional dispersion, which provide complementary information (Villéger et al. 2008; Laliberté and Legendre 2010). First, for each network, we built a Euclidean distance matrix by projecting flowering plant species into a three-dimensional trait space with each axis corresponding to a functional trait. The distance matrix was analysed through Principal Coordinate Analysis (PCoA), and the PCoA axes were then used as new combined traits to compute the functional diversity indices. Categorical variables were transformed into dummy variables (i.e., binary). Functional richness measures the functional space filled by the plant community, i.e., the volume of the convex hull. For each network, we standardized the index value by the “global” functional richness, including all plant species in all networks (Laliberté et al. 2014). Its value ranges from 0 to 1, with increasing values of the index indicating an increase in community functional richness. Functional dispersion additionally takes into account the relative abundance of plant species. The index represents the dispersion of plant species in the trait space, i.e., the distance of species to the centroid of all species in the community, weighted by their relative abundance. Its value ranges from 0 to infinity, with increasing values indicating an increase in functional dispersion, i.e., a strong difference in traits between dominant plant species and low abundant ones. Both indices were calculated using the R package FD (Laliberté et al. 2014).
Functional traits of pollinator species
For each pollinator species, we selected one to four individuals, depending on the availability, and extracted the proboscis which was measured along with total body length (body size). We derived from the literature two additional traits: type of foraging range (two classes: central-place forager, for species which build a nest, and non-central-place forager), and taxonomic family (Table S3; Additional References in ESM). As for corolla shape and length, proboscis length and body size affect the way a pollinator species can exploit a floral resource. The type of foraging range does not directly influence resource selection, but it determines how far pollinators can travel to collect pollen and nectar. Finally, the taxonomic family is often linked to floral preferences or particular mouthpart morphology. Using these traits, we estimated the trait similarity between each wild pollinator species and the honeybee using Gower’s similarity coefficient (Gower 1971) as described by Podani (1999), calculated using the R package FD (Laliberté et al. 2014). For each site, we then determined the CWM trait similarity between the community of wild pollinators and the honeybee by calculating the mean trait similarity value of all wild pollinator species (i.e., excluding the honeybee) weighted by their abundance.
Potential collinearity between predictors
Before performing the statistical analyses described below, we analysed potential collinearity in our data by computing the variance inflation factors (VIFs) using the R package car (John and Weisberg 2019). Plant species richness and standardized functional richness of plant community were strongly correlated (Pearson’s correlation = 0.876, p value < 0.001), as well as temperature and elevation (Pearson’s correlation = 0.751, p value < 0.001). We, therefore, chose to build our models using plant standardized functional richness and temperature as explanatory variables. Functional traits of pollinators were also correlated with each other (Table S4, Fig. S2), so their effect on resource overlap was analysed separately. The explanatory variables of the six global models described in the next paragraph fitted without the interactions had VIFs < 1.5, indicating low collinearity.
Statistical analyses
For the statistical analyses, we followed an information-theoretic approach (Burnham and Anderson 2002), which allows comparing the fit of a set of models rather than selecting one single best model based on p values. The first global model (Model 1) included resource overlap between wild pollinator community and the honeybee as response variable, and the main effects of honeybee abundance, temperature, standardized functional richness of plant community, and trait similarity between wild pollinator community and the honeybee as explanatory variables. The model also included all the interactions that could strongly affect the resource overlap, i.e., the two-way interactions between honeybee abundance and plant standardized functional richness, between honeybee abundance and trait similarity between wild pollinator community and the honeybee, between plant standardized functional richness and trait similarity between wild pollinator community and the honeybee, and the three-way interaction between honeybee abundance, plant standardized functional richness and trait similarity between wild pollinator community and the honeybee. The structure of the second model (Model 2) was similar, but standardized functional richness of plant community was replaced by functional dispersion of plant community.
Second, we explored the effect of single pollinator traits on resource overlap. We, therefore, built four linear mixed-effect models, one for proboscis length (Model 3), one for body size (Model 4), one for type of foraging range (Model 5), and one for taxonomic family (Model 6). Proboscis length and body size of wild pollinators were categorized according to trait values of the honeybee, which possesses a proboscis of c. 5 mm and body size of c. 12 mm. Proboscis length categories for wild pollinators were: proboscis shorter than the honeybee < 3.9 mm, proboscis similar to the honeybee = 4–6.9 mm, and proboscis longer than the honeybee > 7 mm. Body size categories for wild pollinators were: smaller than the honeybee < 7.9 mm, similar to the honeybee = 8–14.9 mm, and larger than the honeybee > 15 mm. We categorized continuous trait variables due to the poor outcome of model residual diagnostics using traits as continuous variables. For taxonomic family, we aggregated families with less than ten collected individuals, i.e., Cimbicidae, Megalodontesidae, Melittidae, and Scoliidae. For each network, we calculated the CWM resource overlap between wild pollinators and the honeybee for each trait category, e.g., for body size, we had one value of CWM resource overlap for wild pollinators smaller than the honeybee, one for wild pollinators similar in size, and one for wild pollinators larger than the honeybee. Each global model included honeybee abundance, temperature, trait category, and the interaction between honeybee abundance and trait category as explanatory variables, and network identity as random factor. In all models described above, the continuous explanatory variables were scaled to mean 0 and standard deviation 1 to make slopes comparable (Gelman 2008). For a summary of the six global models, see Table S5.
Within each set, models were ordered based on their second-order Akaike information criterion corrected for small sample size (AICc), with lower values indicating models that better fit the data. For each model, we calculated the ΔAICc, i.e., the difference between the model AICc and the lowest AICc of the entire set of models (with the best model having ΔAICc = 0), and the Akaike model weight, which indicates the probability that the model is the best one. As a measure of goodness-of-fit, we estimated the R2. Lastly, we calculated the model-averaged partial coefficient for each explanatory variable using all models within each set and estimated the 95% confidence intervals around model-averaged partial coefficients. We presented in the tables all models with ΔAICc < 6 (Harrison et al. 2018). All multi-model analyses were conducted using the R package MuMIn (Barton 2020).
Lastly, we tested for potential spatial autocorrelation of residuals of all models using Moran’s I in the R package ape (Paradis and Schliep 2019). The analyses highlighted no spatial autocorrelation in any of the model (Model 1 p value = 0.692; Model 2 p value = 0.478; Model 3 p value = 0.336; Model 4 p value = 0.842; Model 5 p value = 0.539; Model 6 p value = 0.075).
Methodological considerations
In this study, we opted to sample many sites with a single visit, as we wanted to include a wide range of plant and pollinator functional traits and temperatures. In network ecology, it is common practice to aggregate data collected in multiple sampling events within a single plant–pollinator network (e.g., Montero-Castaño and Vilà 2017; Norfolk et al. 2018; Valido et al. 2019). However, this operation can potentially create artificial species assemblages, i.e., cumulative communities composed of species observed in different days, weeks or seasons, often with non-overlapping phenology (CaraDonna et al. 2020; Schwarz et al. 2020). Using single visit networks, we aimed at exploring the realized interactions between co-occurring individuals of honeybees and wild pollinators, rather than achieving high sampling completeness of pollinator species or interactions. Our interactions can, therefore, be interpreted as short-term, behavioural responses.
Results
General results
Across the 51 networks combined, we observed 262 plant species (Table S2) and 325 pollinator species or morphospecies (Table S3), for a total of 10,841 pollinator visits to flowers. During the 255 h of observation, we recorded 1497 unique plant–pollinator interactions. We identified to the species level 99% of collected wild pollinators (Table S3). We observed an average of 81 wild pollinator individuals (min = 16, max = 332), and of 24 pollinator species (min = 9, max = 49) per site (Table S1). The honeybee was found in all sites and was the most abundant pollinator with 6718 collected individuals (min = 2, max = 768, mean = 132), and the most generalist one, visiting 111 flowering plant species. Other common, abundant and generalist species were Eristalis tenax (Linnaeus), a hoverfly species found at 39 sites with 597 individuals that visited 76 flowering plant species, Bombus pascuorum (Scopoli), a bumblebee species found at 35 sites with 411 individuals that visited 45 flowering plant species, and Sphaerophoria scripta (Linnaeus), a hoverfly species found at 37 sites with 366 individuals that visited 77 flowering plant species. Pollinator proboscis length ranged from 0.4 mm for Entomognathus brevis (Vander Linden) to 16 mm for Bombus gerstaeckeri Morawitz, while body length ranged from 4 mm for Hylaeus taeniolatus Förster and H. imparilis Förster to 22.5 mm for Xylocopa violacea Linnaeus (Table S3).
We observed an average of 20 flowering plant species (min = 8, max = 35) per site (Table S1). The most frequently visited species were Rubus sp. L. (931 total visits, 97% by the honeybee), Centaurea nigrescens Willd. (823 total visits, 84% by the honeybee), and Epilobium angustifolium L. (560 total visits, 93% by the honeybee), while the species most frequently visited only by wild pollinators were Galeopsis pubescens Besser (278 visits), Leucanthemum vulgare Lam. (191 visits), and Erigeron annuus (L.) Pers. (153 visits). Few plant species (N = 9) were exclusively visited by honeybees, while many species were exclusively visited by wild pollinators (N = 102), among which there were many umbellifers such as Daucus carota L., Anthriscus sylvestris (L.) Hoffm., and Heracleum sphondylium L. The most generalist plant species were Ranunculus acris L. (attracting 40 pollinator species), Trifolium pratense L. (attracting 39 pollinator species), and E. annuus (attracting 37 pollinator species). Flower corolla length ranged from 0.05 mm of open disc flowers to 33 mm of Calystegia sepium (L.) R. Br. (Table S2).
Overall functional traits of plants and pollinators
For Model 1, fifteen models showed a ΔAICc < 6 (Table S6). Model averaging indicated that both plant and pollinator functional traits affected the resource overlap between wild pollinator community and the honeybee (Fig. 2). The impact of plant functional traits on resource overlap varied with honeybee abundance: resource overlap decreased as honeybee abundance increased in sites with high plant functional richness, while there was no change in resource overlap with increasing honeybee abundance in sites with low plant functional richness (Fig. 3a). Moreover, resource overlap increased as trait similarity between wild pollinator community and the honeybee increased (Fig. 3b). Temperature and other interactions did not affect the resource overlap (Table S6, Fig. 2).

Model estimates from the model-averaging procedure based on the set of models with all functional traits of both plants and pollinators (Model 1). Explanatory variables of the global model are honeybee abundance (Apis, ln-transformed), temperature (Temp), standardized functional richness of plant community (FRic), trait similarity between wild pollinator community and the honeybee (TSim), and the interactions Apis × FRic, Apis × TSim, FRic × TSim, and Apis × FRic × TSim. All explanatory variables were scaled to mean 0 and standard deviation 1. Dots indicate the model estimated means, while error bars indicate the 95% confidence intervals for the expected values of the variables

Partial residual plots showing the effect of a the interaction between honeybee abundance (ln-transformed) and standardized functional richness of plant community, with the three standardized functional richness levels representing the 10th, 50th, and 90th percentiles, and b trait similarity between wild pollinator community and the honeybee on resource overlap between wild pollinator community and the honeybee (ln-transformed) (Model 1). The shaded areas indicate the 95% confidence intervals for the expected values
For Model 2, twenty-eight models showed a ΔAICc < 6 (Table S7). The resource overlap was affected only by the trait similarity between wild pollinator community and the honeybee (Fig. S3).
Single functional traits of pollinators
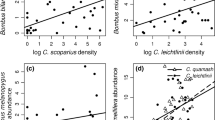
For Model 3, the multi-model inference analysis selected five models with a ΔAICc < 6 (Table S8a). Proboscis length was the only variable affecting the resource overlap between wild pollinator community and the honeybee (Fig. 4a), i.e., pollinators with proboscis length similar to the honeybee showed the highest overlap (Fig. 5a).

Model estimates from the model-averaging procedure based on the four sets of models considering single traits of pollinators, i.e., a proboscis length (Model 3), b body size (Model 4), c type of foraging range (Model 5), and d taxonomic family (Model 6). Explanatory variables of the four global models are honeybee abundance (Apis, ln-transformed), temperature (Temp), the levels of the four trait categories (ProbS proboscis similar to the honeybee, ProbL proboscis longer than the honeybee, BodyS body size similar to the honeybee, BodyL body size larger than the honeybee, ForNC non-central forager, Apid Apidae, Coll Colletidae, Cono Conopidae, Crab Crabronidae, Hali Halictidae, Mega Megachilidae, other other families, i.e., Cimbicidae, Megalodontesidae, Melittidae, and Scoliidae, Syrp Syrphidae, Tach Tachinidae, Tent Tenthredinidae, Vesp Vespidae) and the interactions between honeybee abundance and each levels of the traits. All continuous explanatory variables were scaled to mean 0 and standard deviation 1. Dots indicate the model estimated means, while error bars indicate the 95% confidence intervals for the expected values of the variables

Partial residual plots showing the effect of a proboscis length (Model 3), b body size (Model 4), c type of foraging range (Model 5), and d taxonomic family (Model 6) on resource overlap between wild pollinator community and the honeybee (ln-transformed). The shaded areas indicate the 95% confidence intervals for the expected values
For Model 4, five models had a ΔAICc < 6 (Table S8b). Body size was the only variable affecting resource overlap between wild pollinator community and the honeybee (Fig. 4b), i.e., resource overlap increased with increasing body size (Fig. 5b). Models for body size showed the highest values of R2 compared to other functional traits (Table S8).
For Model 5, six models had a ΔAICc < 6 (Table S8c). Again, only the trait category strongly affected resource overlap between wild pollinator community and the honeybee (Fig. 4c), i.e., central-place foragers showed a higher overlap with honeybees compared to non-central-place foragers (Fig. 5c).
For Model 6, four models showed a ΔAICc < 6 (Table S8d). The taxonomic family strongly affected resource overlap between wild pollinator community and the honeybee (Fig. 4d). Bees of family Apidae showed a higher resource overlap than the other families (Fig. 5d), but the resource overlap was also relatively high for other families such as Conopidae, Halictidae, Megachilidae, and Syrphidae (Fig. 5d). We did not find an interactive effect of honeybee abundance and trait category in any of the models (Fig. 4), meaning that the difference in resource overlap between trait categories was independent of honeybee abundance.
Discussion
Incorporating functional traits into ecological network analyses helped to elucidate the degree of resource overlap between wild pollinators and the honeybee. In particular, a low functional diversity of plant community combined with a high trait similarity between wild pollinators and the honeybee appeared to increase the risk of potential negative impacts of a high honeybee abundance on wild pollinator communities.
In areas with a high abundance of managed pollinators, resource overlap between wild pollinators and the honeybee could be mitigated by a high functional richness of plant community, in which pollinators could shift to alternative food resources, as opposed to areas with a low functional richness. To our knowledge, this is the first time that plant functional diversity was used to explore the changes in the resource overlap between wild pollinators and the honeybee. Previous works highlighted a similar effect of plant diversity and honeybee abundance on pollinator communities, with a reduction of potential competition in sites rich in plant species despite an increase in honeybee abundance (Rodríguez et al. 2021). Similarly, heterogeneous landscapes have been shown to support wild pollinators by reducing competition with honeybees (Herbertsson et al. 2016), while a lower availability of differentiated floral resources might increase competition among pollinator species (Thomson 2016; Wignall et al. 2020a, b). However, in contrast with previous research, we found that the resource overlap between wild pollinators and the honeybee never increased with increasing honeybee abundance (Lindström et al. 2016; but see Hudewenz and Klein 2015), even in sites with low plant functional diversity. This might be related to the honeybee foraging behaviour, as it often focuses on the most abundant and rewarding resources, especially in areas with low diversity of plants (Magrach et al. 2017). On the other hand, the lower resource overlap observed in sites with high functional diversity of plant community and high honeybee abundance could be related to the foraging behaviour of wild pollinators that could be forced to forage on plants that are not visited by honeybees. However, while we found an effect of functional richness of plant community, we observed no effect of functional dispersion. This could be partly explained by the fact that many sites were characterized by the same dominant plant species (e.g., E. annuus and Melilotus albus Medikus) and many different species with lower abundances, so functional dispersion values were similar across sites.
As expected, the resource overlap increased with increasing trait similarity between wild pollinators and the honeybee. Species with similar functional traits usually exploit similar floral resources (Fontaine et al. 2006; Albrecht et al. 2012), so potential competition is expected to be higher for wild pollinators which share traits with the honeybee. First, proboscis length is one of the main constraints of resource selection, affecting whether a pollinator species can obtain nectar from specific flowers. Pollinators are usually more efficient when foraging on plants with flower corolla length matching their mouthpart length (Inouye 1980; Madjidian et al. 2008; Klumpers et al. 2019). For example, hoverflies with a short proboscis tend to prefer flowers that are flat or have a shallow corolla (Fontaine et al. 2006), while long-tongued bumblebees tend to forage on flowers with deep corolla (Balfour et al. 2013). While pollinator species with proboscis shorter or longer than the honeybee mostly foraged on plant species that were not visited by honeybees, pollinators with a similar proboscis visited the same plant species, therefore, increasing their potential competition. Second, body size determines how far pollinators are able to forage, with large pollinators usually having a longer foraging range compared to small species (Gathmann and Tscharntke 2002; Greenleaf et al. 2007). Here, we found that body size was a key functional trait, driving the resource overlap between wild pollinators and the honeybee. The latter increased with increasing body size, even if we expected a higher overlap for species similar in size to the honeybee. Potential competition with honeybees was, therefore, higher for large species, such as bumblebees. Third, we also observed an increase in resource overlap for central-place foragers. These species are obliged to forage relatively near the nest, based on their foraging range, and are, therefore, unable to expand their foraging area, even when the local density of honeybees is high (Walther-Hellwig et al. 2006). Fourth, many Hymenoptera families such as Apidae, Halictidae, and Megachilidae showed a high level of resource overlap with honeybees. Surprisingly, both thick-headed flies (Diptera: Conopidae) and hoverflies (Diptera: Syrphidae), which we expected to mostly visit open disc flowers, also showed a relatively high resource overlap. While the potential negative effects of honeybees on wild pollinators have often focused on wild bees (e.g., Mallinger et al. 2017), other groups of insects might also be affected.
As the honeybee is not particularly active at low temperatures (Jaffé et al. 2010), we expected that its effect on wild pollinators would be stronger in sites with relatively high temperatures. However, similarly to what was observed in other works (e.g., Corcos et al. 2020; Seoane et al. 2021), we did not find any effect of temperature on resource overlap between wild pollinators and the honeybee, even if the observed temperature range was large (min = 18 °C, max = 38 °C).
Conclusions
Honeybees have been introduced worldwide, and, therefore, often cohabit with wild pollinators. As their hives can host more than 50,000 individuals, their abundance in natural and managed habitats can be extremely high. Here, we showed that the potential interactions between wild pollinators and honeybees depended on functional traits of both plants and pollinators. In particular, our results highlight the potential role of plant functional diversity in supporting wild pollinators in areas with high honeybee density by decreasing the resource overlap between wild pollinators and the honeybee. Moreover, as pollinator species with traits similar to those of the honeybee tended to visit the same plant species, they could be more vulnerable to potential competition. From a conservation point of view, particular attention should be paid to the potential effects of beekeeping in sites where pollinator species of conservation concern possess functional traits similar to those of the honeybee. More research is needed to quantify potential short- and long-term effects of high honeybee abundance on fitness, health, and population dynamics of wild pollinators.
Availability of data and material
Once the paper will be accepted, the data supporting the results will be archived in a Zenodo Digital Repository.
Code availability
Once the paper will be accepted, the R code will be archived in a Zenodo Digital Repository.
Change history
24 July 2022
“Missing Open Access funding information has been added in the Funding Note.”.
References
Abrahamczyk S, Kluge J, Gareca Y et al (2011) The influence of climatic seasonality on the diversity of different tropical pollinator groups. PLoS ONE 6:e27115. https://doi.org/10.1371/journal.pone.0027115
Albrecht M, Schmid B, Hautier Y, Müller CB (2012) Diverse pollinator communities enhance plant reproductive success. Proc R Soc B Biol Sci 279:4845–4852. https://doi.org/10.1098/rspb.2012.1621
Balfour NJ, Garbuzov M, Ratnieks FLW (2013) Longer tongues and swifter handling: why do more bumble bees (Bombus spp.) than honey bees (Apis mellifera) forage on lavender (Lavandula spp.)? Ecol Entomol 38:323–329. https://doi.org/10.1111/een.12019
Barton K (2020) MuMIn: multi-model inference. R package version 1.43.17. https://CRAN.R-project.org/package=MuMIn
Bergamo PJ, Streher NS, Wolowski M, Sazima M (2020) Pollinator-mediated facilitation is associated with floral abundance, trait similarity and enhanced community-level fitness. J Ecol 108:1334–1346. https://doi.org/10.1111/1365-2745.13348
Burnham KP, Anderson DR (2002) Model selection and inference: a practical information-theoretic approach. Springer
Cane JH, Tepedino VJ (2017) Gauging the effect of honey bee pollen collection on native bee communities. Conserv Lett 10:205–210. https://doi.org/10.1111/conl.12263
CaraDonna PJ, Burkle LA, Schwarz B et al (2020) Seeing through the static: the temporal dimension of plant–animal mutualistic interactions. Ecol Lett 24:149–161. https://doi.org/10.1111/ele.13623
Corcos D, Cappellari A, Mei M et al (2020) Contrasting effects of exotic plant invasions and managed honeybees on plant–flower visitor interactions. Divers Distrib 26:1397–1408. https://doi.org/10.1111/ddi.13132
Eklöf A, Jacob U, Kopp J et al (2013) The dimensionality of ecological networks. Ecol Lett 16:577–583. https://doi.org/10.1111/ele.12081
Fontaine C, Dajoz I, Meriguet J, Loreau M (2006) Functional diversity of plant-pollinator interaction webs enhances the persistence of plant communities. PLoS Biol 4:e1. https://doi.org/10.1371/journal.pbio.0040001
Fornoff F, Klein AM, Hartig F et al (2017) Functional flower traits and their diversity drive pollinator visitation. Oikos 126:1020–1030. https://doi.org/10.1111/oik.03869
Garibaldi LA, Steffan-Dewenter I, Winfree R et al (2013) Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 339:1608–1611. https://doi.org/10.1126/science.1230200
Garibaldi LA, Bartomeus I, Bommarco R et al (2015) Trait matching of flower visitors and crops predicts fruit set better than trait diversity. J Appl Ecol 52:1436–1444. https://doi.org/10.1111/1365-2664.12530
Gathmann A, Tscharntke T (2002) Foraging ranges of solitary bees. J Anim Ecol 71:757–764. https://doi.org/10.1046/j.1365-2656.2002.00641.x
Gelman A (2008) Scaling regression inputs by dividing by two standard deviations. Stat Med 27:2865–2873. https://doi.org/10.1002/sim
Giannini TC, Garibaldi LA, Acosta AL et al (2015) Native and non-native supergeneralist bee species have different effects on plant-bee networks. PLoS ONE 10:1–13. https://doi.org/10.1371/journal.pone.0137198
Goulnik J, Plantureux S, Théry M et al (2020) Floral trait functional diversity is related to soil characteristics and positively influences pollination function in semi-natural grasslands. Agric Ecosyst Environ 301:107033. https://doi.org/10.1016/j.agee.2020.107033
Goulson D (2003) Effects of introduced bees on native ecosystems. Annu Rev Ecol Evol Syst 34:1–26. https://doi.org/10.1146/annurev.ecolsys.34.011802.132355
Goulson D (2010) Bumblebees: behaviour, ecology, and conservation. Oxford University Press
Gower JC (1971) A general coefficient of similarity and some of its properties. Biometrics 27:857–871. https://doi.org/10.2307/2528823
Greenleaf SS, Williams NM, Winfree R, Kremen C (2007) Bee foraging ranges and their relationship to body size. Oecologia 153:589–596. https://doi.org/10.1007/s00442-007-0752-9
Harrison XA, Donaldson L, Correa-Cano ME et al (2018) A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018:1–32. https://doi.org/10.7717/peerj.4794
Herbertsson L, Lindström SAM, Rundlöf M et al (2016) Competition between managed honeybees and wild bumblebees depends on landscape context. Basic Appl Ecol 17:609–616. https://doi.org/10.1016/j.baae.2016.05.001
Hudewenz A, Klein AM (2015) Red mason bees cannot compete with honey bees for floral resources in a cage experiment. Ecol Evol 5:5049–5056. https://doi.org/10.1002/ece3.1762
Hung K-LJ, Kingston JM, Albrecht M et al (2018) The worldwide importance of honey bees as pollinators in natural habitats. Proc R Soc B Biol Sci 285:20172140. https://doi.org/10.1098/rspb.2017.2140
Inouye DW (1980) The effect of proboscis and corolla tube lengths on patterns and rates of flower visitation by bumblebees. Oecologia 45:197–201. https://doi.org/10.1007/BF00346460
Jaffé R, Dietemann V, Allsopp MH et al (2010) Estimating the density of honeybee colonies across their natural range to fill the gap in pollinator decline censuses. Conserv Biol 24:583–593. https://doi.org/10.1111/j.1523-1739.2009.01331.x
John A, Weisberg S (2019) An R companion to applied regression. Sage, Thousand Oaks
Junker RR, Blüthgen N, Brehm T et al (2013) Specialization on traits as basis for the niche-breadth of flower visitors and as structuring mechanism of ecological networks. Funct Ecol 27:329–341. https://doi.org/10.1111/1365-2435.12005
Klumpers SGT, Stang M, Klinkhamer PGL (2019) Foraging efficiency and size matching in a plant–pollinator community: the importance of sugar content and tongue length. Ecol Lett 22:469–479. https://doi.org/10.1111/ele.13204
Kugler H (1970) Blütenökologie. Gustav Fischer Verlag, Jena
Laliberté E, Legendre P, Shipley B (2014) FD: measuring functional diversity from multiple traits, and other tools for functional ecology. R package version 1.0-12. https://CRAN.R-project.org/package=FD
Laliberté E, Legendre P (2010) A distance-based framework for measuring functional diversity from multiple traits. Ecology 91:299–305. https://doi.org/10.1890/08-2244.1
Lara-Romero C, Seguí J, Pérez-Delgado A et al (2019) Beta diversity and specialization in plant–pollinator networks along an elevational gradient. J Biogeogr 46:1598–1610. https://doi.org/10.1111/jbi.13615
Lázaro A, Gómez-Martínez C, Alomar D et al (2020) Linking species-level network metrics to flower traits and plant fitness. J Ecol 108:1287–1298. https://doi.org/10.1111/1365-2745.13334
Lindström SAM, Herbertsson L, Rundlöf M et al (2016) Experimental evidence that honeybees depress wild insect densities in a flowering crop. Proc R Soc B Biol Sci 283:1–8. https://doi.org/10.1098/rspb.2016.1641
Madjidian JA, Morales CL, Smith HG (2008) Displacement of a native by an alien bumblebee: lower pollinator efficiency overcome by overwhelmingly higher visitation frequency. Oecologia 156:835–845. https://doi.org/10.1007/s00442-008-1039-5
Magrach A, González-Varo JP, Boiffier M et al (2017) Honeybee spillover reshuffles pollinator diets and affects plant reproductive success. Nat Ecol Evol 1:1299–1307. https://doi.org/10.1038/s41559-017-0249-9
Mallinger RE, Gaines-Day HR, Gratton C (2017) Do managed bees have negative effects on wild bees? A systematic review of the literature. PLoS ONE 12:1–32. https://doi.org/10.1371/journal.pone.0189268
Montero-Castaño A, Vilà M (2017) Influence of the honeybee and trait similarity on the effect of a non-native plant on pollination and network rewiring. Funct Ecol 31:142–152. https://doi.org/10.1111/1365-2435.12712
Morisita M (1959) Measuring of the dispersion of individuals and analysis of the distributional patterns. Mem Fac Sci Kyushu Univ Ser E Biol 2:215–235
Norfolk O, Gilbert F, Eichhorn MP (2018) Alien honeybees increase pollination risks for range-restricted plants. Divers Distrib 24:705–713. https://doi.org/10.1111/ddi.12715
Paradis E, Schliep K (2019) Ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35:526–528. https://doi.org/10.1093/bioinformatics/bty633
Podani J (1999) Extending Gower’s general coefficient of similarity to ordinal characters. Taxon 48:331–340. https://doi.org/10.2307/1224438
R Core Team (2019) R: a language and environment for statistical computing. R Foundation for statistical computing, Vienna, Austria
Rodríguez S, Pérez-Giraldo LC, Vergara PM et al (2021) Native bees in Mediterranean semi-arid agroecosystems: unravelling the effects of biophysical habitat, floral resource, and honeybees. Agric Ecosyst Environ 307:107188. https://doi.org/10.1016/j.agee.2020.107188
Schleuning M, Fründ J, García D (2015) Predicting ecosystem functions from biodiversity and mutualistic networks: an extension of trait-based concepts to plant-animal interactions. Ecography 38:380–392. https://doi.org/10.1111/ecog.00983
Schwarz B, Vázquez DP, CaraDonna PJ et al (2020) Temporal scale-dependence of plant–pollinator networks. Oikos 129:1289–1302. https://doi.org/10.1111/oik.07303
Seoane J, Silvestre M, Hevia V et al (2021) Abiotic controls, but not species richness, shape niche overlap and breadth of ant assemblages along an elevational gradient in central Spain. Acta Oecol 110:103695. https://doi.org/10.1016/j.actao.2020.103695
Thomson DM (2016) Local bumble bee decline linked to recovery of honey bees, drought effects on floral resources. Ecol Lett 19:1247–1255. https://doi.org/10.1111/ele.12659
Trøjelsgaard K, Olesen JM (2013) Macroecology of pollination networks. Glob Ecol Biogeogr 22:149–162. https://doi.org/10.1111/j.1466-8238.2012.00777.x
Uyttenbroeck R, Piqueray J, Hatt S et al (2017) Increasing plant functional diversity is not the key for supporting pollinators in wildflower strips. Agric Ecosyst Environ 249:144–155. https://doi.org/10.1016/j.agee.2017.08.014
Valido A, Rodríguez-Rodríguez MC, Jordano P (2019) Honeybees disrupt the structure and functionality of plant-pollinator networks. Sci Rep 91(9):4711. https://doi.org/10.1038/s41598-019-41271-5
Villéger S, Mason NWH, Mouillot D (2008) New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 89:2290–2301. https://doi.org/10.1890/07-1206.1
Violle C, Navas M-L, Vile D et al (2007) Let the concept of trait be functional! Oikos 116:882–892. https://doi.org/10.1111/j.2007.0030-1299.15559.x
Walther-Hellwig K, Fokul G, Frankl R et al (2006) Increased density of honeybee colonies affects foraging bumblebees. Apidologie 38:124–124. https://doi.org/10.1051/apido:200701
Wignall VR, Brolly M, Uthoff C et al (2020a) Exploitative competition and displacement mediated by eusocial bees: experimental evidence in a wild pollinator community. Behav Ecol Sociobiol 74:152. https://doi.org/10.1007/s00265-020-02924-y
Wignall VR, Harry IC, Davies NL et al (2020b) Seasonal variation in exploitative competition between honeybees and bumblebees. Oecologia 192:351–361. https://doi.org/10.1007/s00442-019-04576-w
Woodcock BA, Garratt MPD, Powney GD et al (2019) Meta-analysis reveals that pollinator functional diversity and abundance enhance crop pollination and yield. Nat Commun 10:1–10. https://doi.org/10.1038/s41467-019-09393-6
Zhang J (2016) Package ‘spaa’: Species association analysis. R package version 0.2.2. https://CRAN.R-project.org/package=spaa
Acknowledgements
We would like to thank Giacomo Falzini and Alessandro Marsilio for their support in the fieldwork. We also thank Filippo Prosser (Museo Civico di Rovereto, Italy) for the identification of plants.
Funding
Open access funding provided by Università degli Studi di Padova within the CRUI-CARE Agreement. The research was partly supported by the University of Padua STARS Consolidator Grant (STARS-CoG-2017, BICE project) and partly by the European Union’s Horizon 2020 329 project “Safeguard” (Grant agreement ID: 101003476) funded under Societal Challenges - Climate 330 action, Environment, Resource Efficiency and Raw Materials.
Author information
Authors and Affiliations
Contributions
AC and LM formulated the idea and designed the sampling. AC and GB conducted fieldwork. AC, MM, DP and PC identified pollinator species, and LM identified plant species. AC analysed the data. AC and LM led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Communicated by Heloise Gibb.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cappellari, A., Bonaldi, G., Mei, M. et al. Functional traits of plants and pollinators explain resource overlap between honeybees and wild pollinators. Oecologia 198, 1019–1029 (2022). https://doi.org/10.1007/s00442-022-05151-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-022-05151-6