
Abstract
The current study aims to differentiate, morphologically, between the masticatory muscles in black-tip sharks and shovelnose ray fish, which control the opening and closure of jaws. This work was applied to four specimens of each species, obtained from the Red Sea in Suez. Anatomical dissection was conducted on the cephalic region is order to expose the musculature. The obtained results illustrate each muscle's position, origin, and insertion. The common muscles involved in the feeding of both species were the muscles of mandibular arch: adductor mandibulae, levator palatoquadrati, while the muscles of hyoid arch were levator hyomandibulae, also the hypobranchial arch contained coracomandibularis, coracohyoideus, and coracoarcualis. All muscles of the jaws are responsible for the abduction of mouth except Adductor mandibulae M, Intermandibularis M. involved in closing the mouth, in addition, Preorbitalis M. assists in protruding of upper jaw.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The black-tip shark (Carcharhinus limbatus) is one of the cartilaginous fish belonging to the family Carcharhinidae, present in the Mediterranean and Red Seas. This shark is found in tropical and subtropical waters, along the southeast coast of the United States, where it migrates northward to Georgia and the Carolinas in summer and southward to Florida in winter and also in the Gulf of Mexico and most likely the Caribbean Sea and in South Africa Akel and Karachle (2017). The shovelnose ray (Glaucostegus Typus) is known as the common guitarfish. It belongs to the family: Rhinobatidae). This fish inhabits subtropical and temperate waters, living in shallow waters over mudflats, sandflats, estuaries, and rock reefs as mentioned by Shaker and Rezk (2017).
The upper and lower jaws (mandibular arch), hyoid arch, and branchial region of the majority of fishes are mechanically connected for feeding and breathing. Jaw suspension occurs with muscular action that connects the palatoquadrate (upper jaw) and the cranium (Motta and Wilga 2001; Motta et al. 2002). The muscle action leads to the jaws' protrusion, and lowering of the tongue and pharynx as it manifested in megamouth sharks by the sudden bucco-pharyngeal expansion, (Motta and Wilga 2001).
Some cartilaginous fishes are suited for ram feeding, suction feeding, and filter-feeding and exert a powerful compressive force; these types of feeding mechanisms requires functional adaptation in the jaw musculature (Dean and Motta 2004). There are diet variations between black-tip sharks and shovelnose rays. Black-tip sharks feed on large prey such as triggerfish, catfish, sardines, tilapia, octopi, and lobster (Castro 1996; Alewijnse and Wells 2020) while shovelnose rays eat coastal benthic invertebrates, principally crustaceans such as crabs and shrimps, followed by cephalopods (Valenzuela-Quiñonez et al. 2018). The variety of feeding performance in both species affects the bite force that occurs between eating large prey which is different from that of small prey (Alewijnse and Wells 2020; Valenzuela-Quiñonez et al. 2018).
Species of elasmobranch fishes adapted for consuming hard prey differ morphologically in bite force compared to the consumption of smaller, less durable prey. The force measurements affect how they crush various sizes of prey (Huber et al. 2005). Huber et al. (2005) added that bite force is principally informative in linking morphological and behavioral variables related to prey capture because biting capacity is dictated by cranial muscle morphology which is affected by dietary diversity and changes in feeding.
The present study provides a comparative morphological description of the masticatory muscles, describing their origins and insertion, so as to determine the action generated by each muscle that is involved in feeding and the biting mechanisms. The study shows the variation between the musculature during the opening and closure of the mouth in both cartilaginous fishes.
Material and methods
Our study was conducted according to the international ethical standards set by the Institutional Animal Care and Use Committee (IACUC) (Vet. CU. 01122022537).
The present study applies to four specimens from each cartilaginous fish: the black-tip shark and shovelnose ray collected from the Red Sea in the Suez region, obtained recently dead from commercial shark fishers, about 2-3 kg body weight, and 70-90 cm total length. These eight specimens were preserved in a mixed solution of (formalin 10%, concentrated liquid phenol 88%, glycerin, and water) and were brought to the anatomy lab, where two specimens from both species were dissected by removal of skin and connective tissue on the cephalic region to expose the musculature. In this manner the mandibular, hyoid arch, and hypobranchial muscles are clarified. The other two specimens of each species were prepared for a skeleton, by maceration technique according to (Shaker 2013). That technique is done with manual cleaning using a scalpel and forceps.; removal of the skin, eye, brain, and musculature part of fish to illustrate the muscular attachment. The prepared skull was soaked in warm water for one hour, then immersed in a mixture of (1 part commercial hydrochloric acid 40% and 40 parts water) and left for 3-6 hours The specimen was neutralized in water and ammonia for 20 minutes. Finally, the skull was bleached with hydrogen peroxide 6% for 3-5 hours and preserved in a permanent solution (Formalin 5%) in a glass jar. The results were photographed by using a digital camera 20 mp, 16x. Wilga (2005), Dino and Gerardo (2007) were utilized to denominate the anatomical terms in the study.
Results
The black-tip sharks have a long, narrow pointed snout with a large curved sub-terminal mouth filled with long sharp serrated teeth, but guitarfish have flattened heads, their mouths are small and straight, and on ventral side of their heads, with pebble-like teeth. The slight variation in head structure can affect the orientation of muscles around the head. In black-tip sharks, the muscles are arranged medially, laterally, and ventrally while in guitarfish are mostly ventrally.
I. I. Muscles of black-tip shark
The muscle concerned with feeding in sharks were classified into muscles of the mandibular arch, including the adductor mandibulae, levator labii superioris, levator palatoquadrati, depressor mandibularis, and intermandibularis; the muscles of hyoid arch include the levator hyomandibulae and the hypobranchial muscles which contain the coracomandibularis, coracohyoideus, and coracoarcualis.
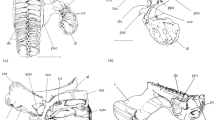
M. Adductor mandibulae (Fig. 1/AM) is a bipennated muscle located just caudal to the mouth commissure on the posterior part of the palatoquadrate (upper jaw) and inserted on the lateral surface of Meckel’s cartilage (lower jaw). It is divided into three parts: anterior and, posterior division of dorsal adductor mandible (the m. adductor mandibulae dorsalis) (Fig. 1/AMDA, AMDP, respectively), The anterior portion had its fibers oriented dorsoventrally, while the posterior portion had ventrodorsal direction. The third part is represented as the ventral adductor mandibulae (the m. adductor mandibulae ventralis, Fig. 1/AMV). It has elongated shape, firmly attached to the ventral surface of Meckel’s cartilage and inserted in the mandibular symphysis.

Photograph of the muscles of mastication in a black-tip shark (Carcharhinus limbatus ((lateral view). LLS, levator labii superioris; AMDA, Anterior part of adductor mandibulae dorsalis; AMDP, posterior part of adductor mandibulae dorsalis; AMV, adductor mandibulae ventralis; Lpq, levator palatoquadrati; Lhm, levator hyomandibulae; E, eye; N, nostril; Pq, palatoquadrate; Mc, Meckel’s cartilage; Pop, postorbital process; Ga, gill arch; po, postorbitalis muscle
M. Levator labii superioris (Fig. 1/LLS) consists of one part that occupies a large area between the nasal capsule and the orbit. It is also termed as preorbitalis M. It originates from the posterior wall of the nasal capsule, lateral-to-lateral ethmoid cartilage, with its muscular fiber directed oblique caudo-ventral then united with the anterior margin of posterior part of adductor mandibulae dorsalis muscle.
M. Levator palatoquadrati (Fig. 1/Lpq) originates from the otic capsule and pterotic cartilage of the neurocranium and is inserted on the lateral border of the palatoquadrate cartilage. It is located caudo-dorsal to the eyeball, medial to postorbital process, and rostral to the cranial margin of adductor mandibulae.
M. Levator hyomandibulae (Figs. 1, 3/Lhm) originates on the otic capsule of the chondrocranium just caudal the levator palatoquadrati and is inserted on the caudodorsal surface of the hyomandibular cartilages. It extends medially with a dorsoventral orientation to the dorsal margin of the adductor mandibulae dorsalis.
M. Intermandibularis (Figs. 2, 3/IM) is a thin transverse sheet of muscle composed of medial and lateral parts. It is attached to the posterior border of Meckel’s cartilage and mandibular symphysis and inserted on the ventral aponeurosis of the m. constrictor hyoideus ventralis. (Fig. 2/Vhc).

Photograph of the muscles of mastication in a black-tip shark (Carcharhinus limbatus) (ventral view). Cr: cranial direction, Cd: caudal direction. LLS, levator labii superioris; IM, intermandibularis; CM, coracomandibularis; CH, coracohyoideus; CA, coracoarcualis; Lec, lateral ethmoid cartilage; N, nostril; Cb, coracoid bar; Ga, gill arch; PF, pectoral fin; Vhc,. constrictor hyoideus ventralis M

Photograph of the chondrocranium of a black-tip shark (Carcharhinus limbatus) with its muscular attachment. (A: ventral view, B: lateral view). Cr: cranial direction, Cd: caudal direction. AMD, adductor mandibulae dorsalis muscle; AMv, adductor mandibulae ventralis muscle; LLS, levator labii superioris; Lpq, levator palatoquadrati; IM, intermandibularis; Lhm, levator hyomandibulae; CM, coracomandibularis; CH, coracohyoideus; CA, coracoarcuals
M. Coracomandibularis (Figs. 2, 3/CM) is a cylindrical single muscle, lying at the mid-ventral part of the floor of the mouth. It is found at the mandibular symphysis and originates on the coracoarcualis.
M. Coracohyoideus (Figs. 2, 3/CH) are paired elongated muscles deep laterally to the coracomandibularis m. It is inserted on the ventral surface of the basihyal cartilage and originates on the anterior margin of m. coracoarcualis.
M. Coracoarcualis (Fig. 2/CA) are paired muscles that have a broadly triangular shape. It originates on a coracoid bar at the pectoral girdle and then it joins with CM and CH.
II. Muscles of shovelnose ray fish
The muscles of the mandibular arch include the adductor mandibulae complex, preorbitalis, and levator palatoquadrati. The muscles of the hyoid arch are the levator hyomandibulae and depressor mandibularis. The hypobranchial muscles include the coracomandibularis, coracohyoideus, coracoarcualis and coracohyomandibularis.
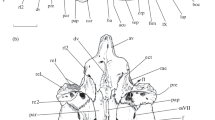
The adductor mandibulae complex is composed of three divisions; medial (Fig. 4/AMM), anterior lateral, and posterior lateral (Figs. 4, 5/AMLA, AMLP), all of them originate on the palatoquadrate (upper jaw) and are inserted on the Meckel’s cartilage (lower jaw). It is located beneath the dorsal rectus muscle. The medial part surrounds the mouth commissure and partially overlaps with the preorbitalis muscle.

Photograph of the muscles of mastication in shovelnose ray fish (Glaucostegus typus) (ventral view). Cr: cranial direction, Cd: caudal direction. AMLA: adductor mandibulae lateral anterior, AMLP: adductor mandibulae lateral posterior, AMM: adductor mandibulae medialis, Pro: preorbitalis M., DM: depressor mandibularis, CM: coracomandibularis, CA: coracoarcuals, CHM: coraco hyomandibularis, PQ: palatoquadrate, Mc: Meckel’s cartilage, VSBc: ventral superficial branchial muscle

Photograph of the deep layer muscles of mastication in shovelnose ray (Glaucostegus typus). Cr: cranial direction, Cd: caudal direction. AMLA: adductor mandibulae lateral anterior, AMP: adductor mandibulae lateral posterior, Pro: preorbitalis M., Lpq: levator palatoquadrati, CM: coracomandibularis, CH: coracohyoideus, CA: coracoarcuals, CHM: Coracohyomandibularis, Pq: palatoquadrate, Mc: Meckel’s cartilage, VSBc: ventral superficial branchial muscle
M. Preorbitalis (Figs. 4, 5, 7/Pro) originates on the nasal capsule ventral to the preorbital process and lies over the levator palatoquadrati muscle. It is inserted on the lower jaw via an elongated tendon joined to the adductor mandibulae. It assists in the jaw protrusion.
M. Depressor mandibularis (Figs. 4, 5, 7/DM) appears as a thin muscular sheet, beneath the dorsal rectus, where it is inserted on the lower jaw.
M. Levator palatoquadrati (Figs. 5, 6/LPq) it originates on the otic region of the chondrocranium and is inserted in the palatoquadrate.

Photograph of levator palatoquadrati in shovelnose ray (Glaucostegus typus) (deep layer). Cr: cranial direction, Cd: caudal direction, D: dorsal direction, V: ventral direction. Lpq: levator palatoquadrati, Lhm: levator hyomandibulae, PQ: palatoquadrate, Mc: Meckel’s cartilage, hyomandibular cartilage
M. Levator hyomandibularis (Fig. 6/ Lhm) is just behind levator palatoquadrati and inserted in the lateral surface of the hyomandibular cartilage.
M. Coracomandibularis (Figs. 4, 5, 7/CM) is a single broad muscle, lying at the mid-ventral part of the floor of the mouth. It is inserted on the Meckel’s cartilage (lower jaw) and originates on coracoid bar of the pectoral girdle.

Photograph of the chondrocranium of shovelnose ray (Glaucostegus typus) with its muscular attachment. Cr: cranial direction, Cd: caudal direction. AM: adductor mandibulae, Pro: preorbitalis M., DM: depressor mandibularis, CM: coracomandibularis, CH: coracohyoideus, CHM: coracohyomandibularis
M. Coracohyoideus (Figs. 4, 5, 7/CH) are paired muscles on each side of the coracomandibularis m., inserting on the ventral surface of the basihyal cartilage.
M. Coracoarcualis (Figs. 4, 5/CA) is paired with broad triangular muscle. It originates on coracoid bar and located in coracoid cartilage.
M. Coracohyomandibularis (Figs. 4, 5, 7/CHM) originates along the hyobranchial cartilage and ends with a tendon at hyomandibular cartilage.
LLS, levator labii superioris; AMDA, anterior part of adductor mandibulae dorsalis; AMDP, posterior part of adductor mandibulae dorsalis; AMV, adductor mandibulae ventralis; Lpq, levator palatoquadrati; Lhm, levator hyomandibulae; E, eye; N, nostril; Pq, palatoquadrate; Mc, Meckel’s cartilage; Pop, postorbital process; Ga, gill arch; po, postorbitalis muscle.
Discussion
In accordance with Huber et al. (2005) in horn sharks, bite force during opening and closure of the mouth and at the feeding mechanism is controlled by different factors including the movement of jaws through cranial muscles involved in adduction and abduction. He also determined that the horn shark is capable of generating bite forces of higher magnitude than comparably sized spiny dogfish (Huber and Motta 2004) and that bite force was in a way suitable for processing hard prey. Our study noted that whole muscles of mastication in black-tip sharks were relatively thick musculature while those of shovelnose fish were relatively thin. That variation in muscle may affect the bite force of both species which is affected by their variation in diet.
The muscles of the mandibular arch in black-tip sharks are classified into adductor mandibulae, levator labii superioris (preorbitalis), levator palatoquadrati and intermandibularis. These results are similar to Dean and Motta (2004), Huber et al. (2005), Wilga (2005), Nakaya et al. (2008) and Huber et al. (2011), while those of shovelnose rays are the same, except only that intermandibularis is non-existed.
According to Dean and Motta (2004), Wilga (2005), Nakaya et al. (2008), Huber et al. (2011), Saber and Hassanin (2014), the muscles of hyoid arch in black-tip sharks and shovelnose rays include the levator hyomandibulae, while the hypobranchial muscles in both species, including coracomandibularis, coracohyoideus, coracoarcualis, except for coracohyomandibularis found only in shovelnose rays.
The adductor mandibulae muscle of black-tip shark is composed of three parts muscles; the adductor mandibulae dorsalis subdivided into an anterior and posterior part, the adductor mandibulae ventralis muscles, that result similar to Gerry et al. (2008), Huber and Motta (2004) but the latter author added another part called superficial quadratomandibularis, while Soares and de Carvalh (2013 b), Nakaya et al. (2008) showed two divisions; dorsal and ventral parts in carcharhiniformes and lamniformes. In shovelnose rays, it is formed of three muscles—medial, anterior lateral, and posterior lateral. These muscles are responsible for jaw-closing by elevating the floor of the mouth.
Our study named the adductor mandibulae muscles as well as Soares and de Carvalho (2013a, b), Gidmark et al. (2012) and Nakaya et al. (2008), while Motta and Wilga (1995), Dean and Motta (2004), Huber and Motta (2004) Wilga (2005) referred that muscles as quadratomandibularis muscle which formed of four division. Motta and Wilga (1995), Wilga (2005) termed the four divisions: quadratomandibularis dorsalis superficial, quadratomandibularis dorsalis medial, quadratomandibularis dorsalis deep, and quadratomandibularis ventralis,
The adductor mandibulae complex M. in both black-tips shark and shovelnose rays was inserted on the lateral surface of lower jaw, even though Huber et al (2005) describe that muscle as quadratomandibularis–preorbitalis complex as having an extensive insertion on the lateral face of both upper and lower jaws, that related to the protrusion of the upper jaw.
The present study as well as the studies of Soares and de Carvalho (2013a, b) in Carcharhiniformes and lamniform families revealed that the m. levator labii superioris originated on the posterior wall of the nasal capsule. However, Soares and de Carvalho (2013a) in squalomorphs indicated some variation as this muscle originated on the lateral ethmoid cartilage but Goto (2001) recorded that in species of Brachaeluridae, Hemiscylliidae, Ginglymostomatidae, Stegostomatidae, and Rhincodontidae, the origin of the levator labii superioris extends to the dorsal surface of the neurocranium.
According to Soares and de Carvalho (2013b), in Lamniformes and Carcharhiniformes, m. levator labii superioris is considered as the cranio-dorsal portion of the adductor mandibulae dorsalis, although Wilga (2005), Huber et al. (2005) termed the same muscle preorbitalis in L. nasus and I. oxyrinchus that extends anteriorly to insert onto the skin just cranial to the quadratomandibularis. According to the origin and insertion of the levator labii superioris muscle, it can reveal a great opening for the mouth.
In line with Wilga (2005) the preorbitals muscle in shovelnose rays was documented as one part as in Mitsukurina owstoni, Carcharias taurus, even though the later author found the preorbitalis in Alopias vulpinus, Lamna nasus, and I. oxyrinchus had two divisions: the original ventral head and a new dorsal head that are distinct from each other. In regard to the origin and insertion of preorbitalis muscle, it acts as an upper jaw protrusion and lower jaw adductor as discussed by Huber et al. (2011)
In accordance with our findings in shovelnose ray fish, Huber et al. (2011) clarified that the depressor mandibularis muscle was firmly attached to Meckel’s cartilage and abducted the lower jaw. The intermandibularis compressed the mouth cavity during prey manipulation through elevation of the floor of the mouth as the result of Soares and de Carballo (2013a), Dino and Gerardo (2007).
In the current investigation, the levator palatoquadrati originates on the otic capsule of the chondrocranium and is inserted on the palatoquadrate that are similar to Dino and Gerardo (2007), Wilga (2005), Dean and Motta (2004). Motta et al. (2002), Nakaya et al. (2008), Wilga (2005), Dean and Motta (2004) agreed with our observation that the levator hyomandibulae originates on the otic capsule of the chondrocranium and is inserted on the caudodorsal surface of the hyomandibular cartilages.
With regard to the hypobranchial muscles, the current study supports the findings of Huber et al. (2011), Nakaya et al. (2008), Wilga (2005), Dean and Motta (2004) who state that the coracomandibularis, the coracohyoideus and the coracoarcuals are responsible for the abduction of the lower jaw. The coracomandibularis assists the opening of the mouth through the depression of the lower jaw. These muscles lay in series on the floor of the mouth, attaching to the posterior border of Meckel’s cartilage in shovelnose rays, while in black-tip sharks it found at mandibular symphysis and hyoid arch, respectively, as mentioned by Huber et al. (2005), Huber and Motta (2004), Wilga and Motta (2000).
Conclusion
Our anatomical description of the masticatory muscles shows variation between the two species and classified according to their origin and insertion morphologically, as shown in Table 1, where the intermandibularis muscle is involved in masticatory muscles of black-tip sharks (adductor muscle) that not found in shovelnose rays; moreover, the coracohyomandibularis muscle is instituted in shovelnose rays. Although the number of muscles in both species is the same, macroscopically the muscles of black-tip sharks are thicker than those of shovelnose rays. The variation in the jaw’s muscular configuration may clarify the feeding types and the force involved in capturing prey in both species.
Data availability statement
The authors confirm that the data used to support the findings of this study are available under this article.
References
Alewijnse SR, Wells RJD (2020) Diet of the Blacktip Shark (Carcharhinus limbatus scientific names should be in italics) in the Northwestern Gulf of Mexico. Gulf and Caribbean Research 31:SC25–SC30. https://doi.org/10.18785/gcr.3101.12
Castro JI (1996) Biology of the blacktip shark Carcharhinus Limbatus, off the Southeastern United States. Bull Mar Sci Miami 59:508–522
Dean M, Motta PJ (2004) Anatomy and functional morphology of the feeding apparatus of the lesser electric ray, Narcine basiliensis (Elasmobranchii: Batoidea). Journal of Morphology 262: 462–483. https://doi.org/10.1002/jmor.10245
Dino P and Gerardo De J (2007) The Dissection of Vertebrates, A Laboratory Manual, chapter3, the shark. Academic Press, Elsevier, pp. 27–45. https://doi.org/10.1093/icb/icv038
Gerry SP, Ramsay JB, Dean MN, Wilga CD (2008) Evolution of asynchronous motor activity in paired muscles: effects of ecology, morphology, and phylogeny. Integr Comp Biol 48:272–82. https://doi.org/10.1093/icb/icn055
Gidmark NJ, Staab KL, Brainerd EL, Hernandez LP (2012) Flexibility in starting posture drives flexibility in kinematic behavior of the kinethmoid-mediated premaxillary protrusion mechanism in a cyprinid fish, Cyprinus carpio. J Exp Biol 215:2262–72. https://doi.org/10.1242/jeb.070516
Goto T (2001) Comparative anatomy, phylogeny and cladistic classification of the order Orectolobiformes (Chondrichthyes, Elasmobranchii). Mem Grad Sch Fish Sci Hokkaido Univ 48:1–100. http://hdl.handle.net/2115/22014
Hernandez LP, Staab KL (2015) Bottom feeding and beyond: how the premaxillary protrusion of cypriniforms allowed for a novel kind of suction feeding. Integr Compar Biol 55:74–84. https://doi.org/10.1093/icb/icv038
Huber DR, Eason TG, Hueter RE, Motta PJ (2005) Analysis of the bite force and mechanical design of the feeding mechanism of the durophagous horn shark Heterodontus francisci. Journal of Experimental Biology 208: 3553–3571. https://doi.org/10.1242/jeb.01816.
Huber DR, Soares MC, de Carvalho MR (2011) Cartilaginous Fishes Cranial Muscles. In: Farrell A.P., (ed.), Encyclopedia of Fish Physiology: From Genome to Environment 1:449–462. San Diego: Academic Press. https://doi.org/10.1016/B978-0-12-374553-8.00238-0
Huber DR, Motta PJ (2004) Comparative analysis of methods for determining bite force in the spiny dogfish Squalus acanthias. J Exp Zool 301:26–37. https://doi.org/10.1002/jez.a.20003
Hulsey CD, Hollingsworth PR, Holzman R (2010) Co-evolution of the premaxilla and jaw protrusion in cichlid fishes (Heroine: Cichlidae). Biol J Linn Soc 100:619–629. https://doi.org/10.1111/j.1095-8312.2010.01468
Kyne P, Bennett MB (2002). Diet of the eastern shovelnose ray, Aptychotrema rostrata (Shaw & Nodder, 1794), from Moreton Bay, Queensland, Australia Marine and Freshwater Research 53. https://doi.org/10.1071/MF01040
Motta PJ, Hueter RE, Tricas TC, Summers AP (2002) Kinematic analysis of suction feeding in the nurse shark, Ginglymostoma cirratum (Orectolobiformes, Ginglymostomatidae). Copeia 24–38. https://doi.org/10.1643/0045
Motta PJ, Wilga CD (1995) J Morphol 226:309–329. https://doi.org/10.1002/jmor.1052260307
Nakaya K, Matsumoto R, Suda K (2008) Feeding strategy of the megamouth shark Megachasma pelagios (Lamniformes: Megachasmidae). J Fish Biol 73:17–34. https://doi.org/10.1111/j.1095-8649.2008.01880.x
Saber AS, Hassanin A (2014) Some Morphological Studies on the Jaw Joint of the Australian Saltwater Crocodile (Crocodylus porosus). No dashVet Anat. 7:55–74. https://doi.org/10.21608/JVA.2014.44742
Shaker N, Rezk H (2017) Anatomical Study on the Gills with its Respiratory Circulation in Shovelnose Ray Fish (Glaucostegus Typus). AJVS 52:125–132. https://doi.org/10.5455/ajvs.250234
Shaker AN (2013) Construction of Anatomical Modules in Some Types of Fish with Special Reference to Reproductive System., Ph. D, Thesis, Cairo University
Soares MC, de Carvalho MR (2013a) a) Mandibular and Hyoid Muscles of Galeomorph Sharks (Chondrichthyes: Elasmobranchii), with Remarks on Their Phylogenetic Intrarelation ships. J of Morphology 274:1111–1123. https://doi.org/10.1002/jmor.20166
Soares MC, de Carvalho MR (2013) b) Comparative myology of the mandibular and hyoid arches of sharks of the order hexanchiformes and their bearing on its monophyly and phylogenetic relationships (Chondrichthyes: Elasmobranchii). J. Morphol 274:203–214. https://doi.org/10.1002/jmor.20088
Staab KL, Ferry L, Hernandez LP (2012) Comparative kinematics of cypriniform premaxillary protrusion. Zoology 115:65–77. https://doi.org/10.1016/j.zool.2011.09.007
Staab KL, Holzman R, Hernandez LP, Wainwright LP (2012b) Independently evolved upper jaw protrusion mechanisms show convergent hydrodynamic function in teleost fishes. J Exp Biol 215:1456–1463. https://doi.org/10.1242/jeb.066308
Valenzuela-Quiñonez F, Galván-Magaña F, Ebert D, Aragón-Noriega E (2018) Feeding habits and trophic level of the shovelnose guitarfish (Pseudobatos productus) in the upper Gulf of California. J. Mar. Biolog. Assoc. UK 98:1783–1792. https://doi.org/10.1017/S0025315417000832
Wilga CD (2005) Morphology and evolution of the jaw suspension in lamniform sharks. J Morphol 265:102–119. https://doi.org/10.1002/jmor.10342
Wilga CD, Motta PJ (2000) Durophagy in sharks: feeding mechanics of the hammerhead Sphyrna tiburo. J Exp Biol. 203:2781–2796. https://doi.org/10.1242/jeb.203.18.2781
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). This study was funded by SDFT according to the agreement between Springer Nature, EKB and Cairo University.
Author information
Authors and Affiliations
Contributions
NAS and AMI designed the protocol, collected the samples, and performed the descriptive morphological features of the masticatory muscles in the cartilaginous fishes. Both authors drafted and reviewed the manuscript, and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
All authors are no conflict of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Shaker, N.A., Ibrahium, A.M. Anatomical variation between the muscles of mastication in black-tip shark (Carcharhinus limbatus) and shovelnose ray Fish (Glaucostegus typus). Zoomorphology 143, 117–126 (2024). https://doi.org/10.1007/s00435-023-00627-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00435-023-00627-2