
Abstract
Several animals use Earth’s magnetic field in concert with other sensor modes to accomplish navigational tasks ranging from local homing to continental scale migration. However, despite extensive research, animal magnetic reception remains poorly understood. Similarly, the Earth’s magnetic field offers a signal that engineered systems can leverage to navigate in environments where man-made positioning systems such as GPS are either unavailable or unreliable. This work uses a behavioral strategy inspired by the migratory behavior of sea turtles to locate a magnetic goal and respond to wind when it is present. Sensing is performed using a number of distributed sensors. Based on existing theoretical biology considerations, data processing is performed using combinations of circles and ellipses to exploit the distributed sensing paradigm. Agent-based simulation results indicate that this approach is capable of using two separate magnetic properties to locate a goal from a variety of initial conditions in both noiseless and noisy sensory environments. The system’s ability to locate the goal appears robust to noise at the cost of overall path length.















Similar content being viewed by others


References
The World Factbook. https://www.cia.gov/library/publications/the-world-factbook/geos/sh.html. Accessed 23 Mar 2017
Anderson JD (2001) Fundamentals of aerodynamics, 3rd edn. McGraw-Hill, New York
Bingman VP, Cheng K (2005) Mechanisms of animal global navigation: comparative perspectives and enduring challenges. Ethol Ecol Evol 17:295–318
Baker TC, Kuenen LPS (1982) Pheromone source location by flying moths: a supplementary non-anemotactic mechanism. Science 216:424–427
Brothers JR, Lohmann KJ (2015) Evidence for geomagnetic imprinting and magnetic navigation in the natal homing of sea turtles. Curr Biol 25:392–396
Browning NA, Grossberg S, Mingolla E (2009) Cortical dynamics of navigation and steering in natural scenes: motion-based object segmentation, heading, and obstacle avoidance. Neural Netw 22:1383–1398
Bostrőm JE, Akesson S, Alerstam T (2012) Where on earth can animals use a geomagnetic bi-coordinate map for navigation? Ecography 35:1039–1047
Boxerbaum AS (2012) Continuous wave peristaltic motion in a robot. Ph.D. Dissertation, Case Western Reserve University
Canciani A, Raquet JF (2016) Absolute positioning using the earth’s magnetic anomaly field. Navigation 63:111–126
Carew TJ (2000) Behavioral neurobiology: the cellular organization of natural behavior. Sinauer Associates Inc., Sunderland
Charlton RE, Cardé RT (1990) Factors mediating copulatory behavior and close-range mate recognition in the male gypsy moth, Lymantria dispar (L.). Can J Zool 68:1995–2004
Conn AR, Cheinberg K, Vicente LN (2009) Introduction to derivative-free optimization. Society for Industrial and Applied Mathmatics, Philidelphia
Coombes S, Graben Pb, Potthast R, Wight J (2014) Neural fields: theory and applications. Springer, Berlin
Crassidis JL, Junkins JL (2012) Optimal estimation of dynamic systems. CRC Press, Boca Raton
Dargahi J, Najarian S (2004) Human tactile perception as a standard for artificial tactile sensing—a review. Int J Med Robot Comput Assist Surg 1:23–35
Dey S (2014) Fluvial hydrodynamics. Springer, Berlin
Endres CS, Putman NF, Lohmann KJ (2009) Perception of airborne odors by loggerhead sea turtles. J Exp Biol 212:3823–3827
Endres CS, Lohmann KJ (2013) Detection of coastal mud odors by loggerhead sea turtles: a possible mechanism for sensing nearby land. Mar Biol 160:2951–2956
Endres CS, Putman NF, Ernst DA, Kurth JA, Lohmann CMF, Lohmann KJ (2016) Multi-modal homing in sea turtles: modeling dual use of geomagnetic and chemical cues in island-finding. Front Behav Neurosci 10:19. doi:10.3389/fnbeh.2016.00019
Goyret J, Markwell PM, Raguso RA (2007) The effect of decoupling olfactory and visual stimuli on the foraging behavior of Manduca sexta. J Exp Biol 210:1398–1405
Grasso FW, Consi TR, Mountain DC, Atema J (2000) Biomimetic robot lobster performs chemo-orientation in turbulence using an pair of spatially separated sensors: progress and challenges. Robot Auton Syst 30:115–131
Grasso FW, Atema J (2002) Integration of flow and chemical sensing for guidance of autonomous marine robots in turbulent flows. Environ Fluid Mech 2:95–114
Jensen KK (2010) Light-dependent orientation responses in animals can be explained by a model of compass cue integration. J Theor Biol 262:129–141
Johnsen S, Lohmann KJ (2005) The physics and neurobiology of magnetoreception. Nat Rev Neurosci 6:703–712
Johnsen S, Lohmann KJ (2008) Magnetoreception in animals. Phys Today 61:29–35
Keshavan J, Humbert JS (2010) MAV stability augmentation using weighted outputs from distributed hair sensor arrays. In: American Control Conference, pp 4445–4450
Knecht DJ, Shuman BM (1985) The geomagnetic field. In: Jursa A (ed) Handbook of geophysics and the space environment. Air Force Geophysics Laboratory, Springfield, VA
Lilienthal AJ, Loutfi A, Duckett T (2006) Airborne chemical sensing with mobile robots. Sensors 2006:(6)1616–1678
Lohmann KJ, Hester JT, Lohmann CMF (1999) Long-distance navigation in sea turtles. Ethol Ecol Evol 11:1–23
Lohmann KJ, Lohmann CMF, Putman NF (2007) Magnetic maps in animals: nature’s GPS. J Exp Biol 210:3697–3705
Lohmann KJ, Luschi P, Hays GC (2008) Goal navigation and island-finding in sea turtles. J Exp Mar Biol Ecol 356:83–95
Lohmann KJ, Putman NF, Lohmann CMF (2012) The magnetic map of hatchling loggerhead sea turtles. Curr Opin Neurobiol 22:336–342
Mouritsen H, Hore P (2012) The magnetic retina: light-dependent and trigeminal magnetoreception in migratory birds. Curr Opin Neurobiol 22:343–352
Painter KJ, Hillen T (2015) Navigating the flow: individual and continuum models for homing in flowing environments. J R Soc Interface 12(112):20150647. doi:10.1098/rsif.2015.0647
Postlethwaite CM, Walker MM (2011) A gemoetric model for initial orientation errors in pigeon navigation. J Theor Biol 269:273–279
Putman NF, Lohmann KJ (2008) Compatibility of magnetic imprinting and secular variation. Curr Biol 18:R596–R597
Putman NF, Endres CS, Lohmann CMF, Lohmann KJ (2011) Longitude perception and bicoordinate magnetic maps in sea turtles. Curr Biol 21:463–466
Putman NF, Verley P, Shay TJ, Lohmann KJ (2012) Simulating transoceanic migrations of young loggerhead sea turtles: merging magnetic navigation behavior with an ocean circulation model. J Exp Biol 215:1863–1870
Putman NF, Verley P, Endres CS, Lohmann KJ (2015) Magnetic navigation behavior and the oceanic ecology of young loggerhead sea turtles. J Exp Biol 218:1044–1050
Putman NF (2015) Inherited magnetic maps in salmon and the role of geomagnetic change. Integr Comp Biol 55(3):396. doi:10.1093/icb/icv020
Renaudin V, Afzal MH, Lachapelle G (2010) New method for magnetometers based orientation estimation. In: Position location and navigation symposium, pp 348–356
Reppert SM, Gegear RJ, Merlin C (2010) Navigational mechanisms of migrating monarch butterflies. Trends Neurosci 33:399–406
Ritz T, Adem S, Schulten K (2000) A model for photoreceptor-based magnetoreception in birds. Biophys J 78(2):707–718
Rutkowski AJ, (2008) A Biologically-inspired sensor fusion approach to tracking a wind-borne odor in three dimensions. Ph.D. Dissertation, Case Western Reserve University
Rutter BL, Taylor BK, Bender JA, Blmel MA, Lewinger WA, Ritzmann RE, Quinn RD (2011) Descending commands to an insect leg controller network cause smooth behavioral transitions. In: International conference on intelligent robots and systems, pp. 215–220
Rutter BL, Taylor BK, Bender JA, Blűmel MA, Lewinger WA, Ritzmann RE, Quinn RD (2011) Sensory coupled action switching modules (SCASM) for modeling and synthesis of biologically inspired coordination. In: International conference on climbing and walking robots
Servedio MR et al (2014) Not just a theory –the utility of mathematical models in evolutionary biology. PLoS Biol 12(12):12
Shaw J, Boyd A, House M, Woodward R, Falko M, Cowin G, Saunders M, Baer B (2015) Magnetic particle-mediated magnetoreception. J R Soc Interface 12(110):20150499. doi:10.1098/rsif.2015.0499
Shockley JA, Raquet JF (2014) Navigation of ground vehicles using magnetic field variations. Navigation 61:237–252
Stoddard PK, Mardsen EJ, Williams TC (1983) Computer simulation of autumnal bird migration over the western North Atlantic. Anim Behav 31:173–180
Szczecinski NS (2012) Massively distributed neuromorphic control for legged robots modeled after insect stepping. MS Thesis, Case Western Reserve University
Szczecinski NS, Brown AE, Bender JA, Quinn RD, Ritzmann RE (2013) A neuromechanical simulation of insect walking and transition to turning of the cockroach Blaberus discoidalis. Biol Cybern 108:1–21
Talley JL (2010) Males chasing females: a comparison of flying Manduca sexta and walking Periplaneta americana male tracking behavior to female sex pheromones in different flow environments. Ph.D. Dissertation, Case Western Reserve University
Taylor BK (2016) Validating a model for detecting magnetic field intensity using dynamic neural fields. J Theor Biol 408:53–65
Taylor BK (2012) Tracking fluid-borne odors in diverse and dynamic environments using multiple sensory mechanisms. Ph.D. Dissertation, Case Western Reserve University
Taylor BK, Rutkowski AJ (2015) Bio-inspired magnetic field sensing and processing. In: Proceedings of the Institute of Navigation’s Pacific PNT conference
Trappenberg TP (2010) Fundamentals of computational neuroscience, 2nd edn. Oxford University Press, New York
Wajnberg E, Acosta-Avalos D, Alves OC, de Oliveria JF, Srygley RB, Esquivel DMS (2010) Magnetoreception in eusocial insects: an update. J R Soc Interface 7:S207–S225
Walker M (1998) On a wing and a vector: a model for magnetic navigation by homing pigeons. J Theor Biol 192:341–349
Webb B (2000) What does robotics offer animal behaviour? Anim Behav 60:545–558
Willis MA, Arbas EA (1991) Odor-modulated upwind flight of the sphinx moth, Manduca sexta (L.). J Comp Physiol A 169:427–440
Willis MA, Avondet JL, Finnell AS (2008) Effects of altering flow and odor information on plume tracking behavior in walking cockroaches, Periplaneta americana (L.). J Exp Biol 211:2317–2326
Wilson H, Cowan JD (1973) A mathematical theory of the functional dynamics of cortical and thalamic nervous tissue. Kybernetik 13:55–80
Wilson H (1999) Spikes, decisions, and actions: the dynamical foundations of neuroscience. Oxford University Press, New York
Wiltschko R, Wiltschko W (1995) Magnetic orientation in animals. Springer, Berlin
Wiltschko R, Stapput K, Thalau P, Wiltschko W (2010) Directional orientation of birds by the magnetic field under different light conditions. J R Soc Interface 7:S163–S177
Winklhofer M (2010) Magnetoreception. J R Soc Interface 7:S131–S134
Zhang K (1996) Representation of spatial orientation by the intrinsic dynamics of the head-direction cell ensemble: a theory. J Neurosci 16:2112–2126
Zill S, Schmitz J, Bűschges A (2004) Load sensing and control of posture and locomotion. Arthropod Struct Dev 33:273–286
Acknowledgements
Dr. Taylor thanks Drs. Adam Rutkowski and Jennifer Talley (AFRL/RWWI), Ms. Pamela Card and Mr. Martin Wehling (AFRL/RWWI), Mr. Sergio Cafarelli (AFRL/RWWG), and Mr. Brian Ortiz-Muñoz (University of Colorado at Boulder) for their discussions, insights, editorial suggestions. He also thanks Dr. Arunava Banerjee (University of Florida) and Dr. Ken Lohmann (University of North Carolina - Chapel Hill) for their insights on this work.
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix
A Determining sigmoid parameters based on expected sensing values
Recall the general form of the sigmoids used in Eqs. 1 and 2
If the environment’s minimum and maximum stimulus values are known (i.e., values of s), along with outputs that correspond to these stimuli (i.e., values of A), then it is possible to determine the parameters K and b algebraically, provided that the rate parameter a is specified. Let \(r = s - s_\mathrm{goal}\). Then Eq. 7 becomes
Specifying minimum and maximum values for the stimulus gives \(r_\mathrm{max} = s_\mathrm{max} - s_\mathrm{goal}\) and \(r_\mathrm{min} = s_\mathrm{min} - s_\mathrm{goal}\). Substituting these values into Eq. 8 gives the following
Here, \(A_\mathrm{min}\) and \(A_\mathrm{max}\) are the outputs corresponding to the minimum and maximum stimuli, respectively. These are two linear equations in the unknowns K and b. Solving Eqs. 10 and 10 yields the following closed form expressions.
In Eqs. 11 and 12, if the stimulus value is allowed to tend toward \(\pm \infty \), limits can be appropriately taken to obtain the final values of K and b.
B Proxy magnetic environment
The contours for the proxy magnetic environment are created using the velocity potential and streamlines created by a lifting cylinder (Anderson 2001). The flow properties here are not those that are used by the agent when it responds to a wind. Properties described here are solely for the purpose of creating a proxy magnetic environment. Expressed in Cartesian coordinates, the streamlines and potential lines are given, respectively, as the following.
Here, x and y are spatial coordinates, and \(V_{\infty }\), \({\varGamma }\), and R are the aerodynamic properties of a lifting cylinder. \(V_{\infty }\) is the flow velocity, \({\varGamma }\) is the circulation, and R is the cylinder’s radius. Increasing the value of \({\varGamma }\) relative to \(V_{\infty }\) increases the overall curvature of the streamlines and potential lines, creating a more nonlinear contour map. If \({\varGamma }\) were zero, the streamlines in Fig. 3a and potential lines in Fig. 3b would be, respectively, horizontal and vertical, except for the immediate vicinity of the cylinder at the origin. Increasing R increases the radius of the cylinder, which increases the spatial range of contours that lie within but do not cross the cylinder boundary. From the perspective of this work, \(V_{\infty }\), \({\varGamma }\), and R are constants that adjust the shape of the environment’s contours (values given in Table 4).
C General mathematical approach
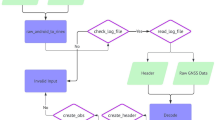
This section provides the overall mathematical approach outlined in Fig. 7 as it is implemented. The intent is to frame the discussion presented in “Appendix 1.” Referring to Fig. 7, the process starts by sensing a physical quantity \(p_{j^*}\) (e.g., the magnetic field, the wind, olfactory cues). This sensor measurement is converted to a nervous system representation based on the ellipses described in Eq. 1. This is done by computing the parameters A and B using Eqs. 2 and 3 (rewritten for clarity).
Here, q denotes the qth sensor in the distributed sensor array. In general, several sensor modes could be combined with each other at this stage. This is represented with the following notation
where H is a function that is used in analogy to different populations of neurons inhibiting or exciting each other. In the current implementation, H is used to combine raw data from several sensor modes into one raw effective (jth) sensor mode. For example, in (Jensen 2010), this corresponded to using the absolute value of a difference, or
When combining a circular distribution and an elliptical distribution, the absolute value operation produces the oblong black distribution shown in Fig. 4b. In Jensen’s work, this created a raw effective sensor mode that is a combination of light-based sensing and magnetic sensing.
With the raw effective sensor modes computed, effective sensor modes are created that are weighted versions of the raw effective sensor modes. One could think of this as creating a behaviorally relevant representation of the world. For example, even though one sensing mode might be constant in time, data from a different sensing mode that are changing over time might alter how the agent responds to the static signal. This is done by creating weights that are functions of the following form
where the weight \(w_i\) can be any function that is designed to elicit the necessary behavior, and i and j are independent integers that denote different sensor modes. Once the weights are computed, effective sensor modes of the following form are computed.
Here, the function f could be as simple as assigning the product \(w_i\Big (\mathrm{SF}_j\Big )\cdot \mathrm{SF}_i\) to the effective senor mode, \(\mathrm{SF}_{eff}\). The act of computing weights and using them to create an effective representation of the sensory environment is also analogous to one population of neurons inhibiting or exciting another.
The controller contribution from each effective sensor mode can be computed as
where \(\dot{{\varLambda }_i}\) is a function that maps the ith effective sensor mode to an appropriate control value. Once the controller contributions are computed, Eq. 4 can be used to compute the total command contribution to the agent. In this way, anything in the external environment is represented by elliptical spike rate distributions. The data encoded by these distributions are combined and processed to yield some type of motor command.
D Bicoordinate algorithm
1.1 D.1 Determining weights and effective distributions
To implement the proposed strategy, weights need to be selected that will scale the raw effective spike rate ellipses and circles based on current sensor feedback. To do this, first recall Fig. 3. Denote the contour lines in Fig. 3a by \(\gamma \), and those in Fig. 3b by \(\beta \). For the bicoordinate strategy, the magnetic \(\gamma \) and \(\beta \) sensor distributions are used at the current time and a previous time to determine whether the agent is moving North/South, or East/West. Define \(\mathrm{SF}_{\gamma , t-l} = \mathrm{SF}(\gamma , t-l)\), and \(\mathrm{SF}_{\beta , t-l} = \mathrm{SF}(\beta , t-l)\), where t is the current time step, and \(t-l\) is l time steps before the current time step. With this definition, North/South and East/West motion can be determined via the sign of the following differences.
The mean function 1) provides a single number from the difference of the two measurement distributions at times t and \(t-l\), and 2) ensures that in cases where noise is present, the resulting differences can be positive or negative. With the environmental formulation of Fig. 3 and Appendix 1, the measurement \(sign(\big (mean(\gamma _t-\gamma _{t-l})\big )>0\) implies the agent is moving North, while the measurement \(sign\big (mean(\beta _t-\beta _{t-l})\big )>0\) implies the agent is moving East. This occurs because the values of each environment contour map increase toward the North and East, respectively.
With an estimated direction of motion, an effective neural output for generating motor commands needs to be determined. A distribution of the following form was sought:
Here, for notational convenience, \(\rho \) is a placeholder for \(\gamma \) and \(\beta \). Also, \(\mathrm{SF}_{\rho , t-l} = \mathrm{SF}(\rho , t-l)\), where t is the current time step, and \(t-l\) is l time steps before the current time step. \(w_{m}\) is a weight that affects the magnetic spike rate ellipses, \(\mathrm{SF}_{\rho t}\) is a magnetic spike rate ellipse for a specific magnetic property at the current time step, and \(\mathrm{SF}_{\rho t}^w\) is the weighted magnetic spike rate for a specific magnetic property at the current time step. Based on the goals outlined in section 2.5, the following properties were sought for the weight:
-
1.
Wind speeds below a threshold should allow the magnetic spike rate generated by the magnetic field to be used as the spike rate that generates command values. This means that \((w_{m})(\mathrm{SF}_{m} )\approx \mathrm{SF}_{m}\)
-
2.
Wind speeds above a threshold should force the magnetic spike rate generated by the magnetic field to induce no behavioral changes
-
3.
Weights vary linearly with wind speeds that are between thresholds.
To satisfy the first two conditions, one can define a ratio that compares a reference spike rate to the current spike rate
Here, \(\mathrm{SF}_{\rho - Ref}\) is the spike rate at which the agent should exhibit zero turn rate due to the \(\rho \)th magnetic sensor contribution, and \(\mathrm{SF}_{\rho t}\) is the current spike rate. With this parameter defined, conditions 1 and 2 can be satisfied in the following way
-
1.
If \(w_{m}=1\), then \((w_{m} )(\mathrm{SF}_{m} )=\mathrm{SF}_{m}\)
-
2.
If \(w_{m}= \rho _{ratio}\), then \((w_{m} )(\mathrm{SF}_{m} ) = \frac{\mathrm{SF}_{\rho - Ref}}{\text {mean}(\mathrm{SF}_{\rho , t} )} (\mathrm{SF}_{m} )=\mathrm{SF}_{\rho - Ref})\)
This provides two points that can be used to generate the linear requirement of condition 3 (illustrated in Fig. 16).

Piecewise linear wind-gated magnetic weighting. Here, U is the wind speed, and \(\mathrm{SF}_{WS}\) is a spike rate due to the wind speed. \(\mathrm{SF}^*_\mathrm{min}\) and \(\mathrm{SF}^*_\mathrm{max}\) are the minimum and maximum wind speed spike rates, respectively. \(w_m\) is the magnetic weighting
In Fig. 16, the x-axis is the spike rate from the wind, and the y-axis is the magnetic weight. Defining the wind speed as U, \(U_\mathrm{max}\) is the wind speed at which wind-driven behavior should dominate, and magnetically driven behavior should be suppressed. \(U_\mathrm{min}\) is the wind speed at which magnetically driven behavior should still be present, and the minimum wind speed at which wind-driven behavior should appear. Consequently, \(\mathrm{SF}_{WS}^* (U)_\mathrm{max}\) is the wind speed spike rate at which magnetic behavior should be suppressed, and \(\mathrm{SF}_{WS}^*(U)_\mathrm{min}\) is the minimum wind speed spike rate at which wind-driven behavior should appear. Using the point–slope formula, the point \((\mathrm{SF}_{WS}^*(U)_\mathrm{min}, 1)\), and denoting wind speed by WS one can create the following piecewise linear relationship
Note that \(\mathrm{SF}_{WS}^* = \mathrm{max}\Big (\mathrm{SF}_{WS} \Big )\), where \(\mathrm{max}(\mathrm{SF}_{WS})\) is the maximum value of the currently sensed wind speed spike ellipse. With this relationship, Eq. 22 can be satisfied.
1.2 D.2 Mapping Spike rate distributions to commands
With weighting and effective spike rate values determined, a function that maps the spike rate into a turn rate is needed. A linear function was chosen for simplicity. With the goal of having the reference spike rate \(\mathrm{SF}_{\rho -Ref}\) produce zero turn rate, Fig. 8A can be sketched. The difference from the zero-turn-rate spike rate \(\mathrm{SF}_{\rho -Ref}\) is computed. This difference is then added or subtracted to \(\mathrm{SF}_{\rho -Ref}\). This difference, denoted here as \({\varDelta }\) can be computed as
Using North/South tracking as an example, \(|{\varDelta }|\) becomes
To determine the turn direction, the logic of Fig. 9 was used.
Suppose that there is no wind, so that the weighting is unity. Based on the logic illustrated in Fig. 9, tracking North/South gives the following
-
If South of goal \((\text {mean}(\mathrm{SF}_{\gamma , t} )- \mathrm{SF}_{\gamma -Ref}<0)\)
-
If moving West: \(\text {mean}(\mathrm{SF}_{\beta , t} )- \mathrm{SF}_{\beta , t - l}<0 \Rightarrow \dot{{\varLambda }}<0\)
-
If moving East: \(\text {mean}(\mathrm{SF}_{\beta , t} )- \mathrm{SF}_{\beta , t - l}>0 \Rightarrow \dot{{\varLambda }}>0\)
-
-
If North of goal \((\text {mean}(\mathrm{SF}_{\gamma , t} )- \mathrm{SF}_{\gamma -Ref}>0)\)
-
If moving West: \(\text {mean}(\mathrm{SF}_{\beta , t} )- \mathrm{SF}_{\beta , t - l}<0 \Rightarrow \dot{{\varLambda }}>0\)
-
If moving East: \(\text {mean}(\mathrm{SF}_{\beta , t} )- \mathrm{SF}_{\beta , t - l}>0 \Rightarrow \dot{{\varLambda }}<0\)
-
Based on this logic, if the agent is moving West, the sign that \(sign(\mathrm{SF}_{\gamma , t} - \mathrm{SF}_{\gamma -Ref})\) would give is needed, whereas if the agent is moving East, then the sign of \((\mathrm{SF}_{\gamma , t} - \mathrm{SF}_{\gamma -Ref})\) needs to be reversed. This leads to the following way to appropriately determine the sign on \({\varDelta }\) for tracking streamlines
Putting all of this together yields the following effective magnetic spike rate for tracking North/South
The same approach for tracking East/West can be used
-
If East of goal \((mean(\mathrm{SF}_{\beta , t} )- \mathrm{SF}_{\beta -Ref}<0)\)
-
If moving North: \(\text {mean}(\mathrm{SF}_{\gamma , t} )- \mathrm{SF}_{\gamma , t - l}<0 \Rightarrow \dot{{\varLambda }}>0\)
-
If moving South: \(\text {mean}(\mathrm{SF}_{\gamma , t} )- \mathrm{SF}_{\gamma , t - l}>0 \Rightarrow \dot{{\varLambda }} <0\)
-
-
If West of goal \((\text {mean}(\mathrm{SF}_{\beta , t} )- \mathrm{SF}_{\beta -Ref}<0)\)
-
If moving North: \(\text {mean}(\mathrm{SF}_{\gamma , t} )- \mathrm{SF}_{\gamma , t - l}<0 \Rightarrow \dot{{\varLambda }}<0\)
-
If moving South: \(\text {mean}(\mathrm{SF}_{\gamma , t} )- \mathrm{SF}_{\gamma , t - l}>0 \Rightarrow \dot{{\varLambda }} >0\)
-
Making the same kinds of arguments as was done for the North/South case, one can obtain the following sign requirement for \(|{\varDelta }|\).
This leads to the following effective magnetic spike rate for tracking East/West
A linear function is used to transform the effective spike rate into the turn rate. Using Fig. 8 as a guide, with reference points at the minimum spike rate and the reference spike rate for a slope, one can write the following
E Determining wind direction using ellipses and circles

Illustration of fixed coordinates (X, Y), and the agent’s (1) body coordinates (x, y), (2) nominally stimulated distribution (blue ellipse), (3) actual stimulated distribution (purple ellipse), and (4) angle difference between distributions. The purple arrow represents, in this case, a wind velocity vector
To determine the wind angle, an ellipse based on Eq. 1 that represents the wind vector’s magnitude (related to the parameters A and B) and direction (i.e., \(\phi \)) was computed. The wind angle \(\zeta \) was computed according to the following equation based on Fig. 17
\(\delta \) is the angle between the fixed X axis and the major axis of the stimulated distribution, and \({\varPsi }\) is the angle between the fixed X axis, and the body coordinate x axis (Fig. 17). The minus sign is used for the purposes of this particular implementation. Next, the parameters A and B are computed according to the wind speed based on Eqs. 2 and 3. Defining the wind vector as \(\mathbf {U}_{\infty }\), and the wind speed as \(U_{\infty } = ||\mathbf {U}_{\infty }||\), this gives the following.
With the angle and wind speed-based constants determined, a spike rate ellipse based on Eq. 1 can be computed.
With the wind vector determined, a circular distribution was used to determine the wind angle. A circular distribution was used because 1) the agent does not actually know its orientation \(\phi \) in the environment, and 2) it was desired to represent all of the sensory information from the environment with elliptical spike rate distributions, which is analogous to how an animal would have to encode sensory information (i.e., an animal encodes all information about the environment through its nervous system). First, the parameters for the circular distribution are computed.
With the angle and wind speed-based constants determined, a wind angle spike rate ellipse based on Eq. 1 can be computed.
Setting \(a_A = a_B\), and the minimum and maximum values of \(A_{\phi }\) and \(B_{\phi }\) to be the same, yields a circular distribution whose radius is nominally \(A_{\phi }\).With these two distributions determined, functions can now be constructed that 1) compute an effective representation of the sensed wind by using an appropriate weighting, and 2) map the effective wind representation to an appropriate turn rate. The goal is to turn into the wind when it is relatively strong. This means that the behavioral response to the wind (i.e., the turn rate) is based on the sensed wind angle, while the behavioral importance of the wind (i.e., the weighting) is determined by wind speed. A piecewise linear weight with the following form was chosen, similar to that used for the magnetic weighting.

\(\varepsilon \) is a value that is chosen so that if the total wind spike rate is the same as the spike rate at which wind becomes behaviorally important, then there is a nonzero effective spike rate, though for now, an offset of zero is used for simplicity. \(\mathrm{SF}_{\text {TW}}|_\mathrm{max}\) is the maximum spike rate that can be generated by the Total Wind distribution. \(\mathrm{SF}_{WS}^*(U)_\mathrm{min}\) is the spike rate generated by the minimum wind speed necessary to elicit a behavioral response. The effective wind spike rate is then given as
With this in mind, a sigmoidal turn rate response can be constructed (Fig. 8B). The curves are generated by the following
\(\text {sign}(\phi )\) determines whether the wind sensors on the agent’s left (i.e., port) or right (i.e., starboard) side are being stimulated.
F Final implementation
An overall system command is generated by adding the turn rate due to the wind, \(\dot{{\varLambda }}_{\zeta }\) (Eq. 40), to the turn rate due to the magnetic field, \(\dot{{\varLambda }}_{m}\) (Eq. 30). This is the final step depicted in Fig. 7.
Rights and permissions
About this article

Cite this article
Taylor, B.K. Bioinspired magnetic reception and multimodal sensing. Biol Cybern 111, 287–308 (2017). https://doi.org/10.1007/s00422-017-0720-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00422-017-0720-3