
Abstract
Aim
Montmorency cherries are rich in polyphenols that possess antioxidant, anti-inflammatory and vasoactive properties. We investigated whether 7-day Montmorency cherry powder supplementation improved cycling time-trial (TT) performance.
Methods
8 trained male cyclists (\(\dot {V}{{\text{O}}_{2{\text{peak}}}}\): 62.3 ± 10.1 ml kg−1 min−1) completed 10-min steady-state (SS) cycling at ~ 65% \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\) followed by a 15-km TT on two occasions. Participants consumed 6 pills per day (Montmorency cherry powder, MC; anthocyanin 257 mg day−1 or dextrose powder, PL) for a 7-day period, 3 pills in the morning and evening. Capillary blood [lactate] was measured at baseline, post SS and post TT. Pulmonary gas exchange and tissue oxygenation index (TOI) of m. vastus lateralis via near-infrared spectroscopy, were measured throughout.
Results
TT completion time was 4.6 ± 2.9% faster following MC (1506 ± 86 s) supplementation compared to PL (1580 ± 102 s; P = 0.004). Blood [lactate] was significantly higher in MC after SS (PL: 4.4 ± 2.1 vs. MC: 6.7 ± 3.3 mM, P = 0.017) alongside an elevated baseline TOI (PL: 68.7 ± 2.1 vs. MC: 70.4 ± 2.3%, P = 0.018).
Discussion
Montmorency cherry supplementation improved 15-km cycling TT performance. This improvement in exercise performance was accompanied by enhanced muscle oxygenation suggesting that the vasoactive properties of the Montmorency cherry polyphenols may underpin the ergogenic effects.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Reactive oxygen species (ROS) are continuously generated during repetitive muscular action from a variety of sources including enzymes such as NADPH oxidase and xanthine oxidase (Reid 2016a) in an intensity-dependent fashion (Bailey et al. 2007). Reactive oxygen species act as important signalling molecules and have been implicated in contraction-mediated increase in muscle glucose uptake (Merry and McConell 2009) and control of skeletal muscle blood flow (Trinity et al. 2016). It appears that under conditions of low oxidative stress and redox balance, ROS promote optimal vasodilation and hyperaemia in exercising muscle (Durand et al. 2015). However, under conditions of oxidative stress or already disturbed redox balance, ROS generation during exercise impairs blood flow and vasodilatory capacity (Donato et al. 2010).
Ryanodine receptors are the major calcium release channel in sarcoplasmic reticulum and due to the high number of cysteine residues in this protein, it is redox sensitive. Excess ROS generation has been shown to impair calcium handling and sensitivity, resulting in reduced contractile force development, thus impairing exercise performance (Reid 2016a). It is, therefore, plausible that elevated muscle antioxidant capacity may counteract fatigue and enhance performance during high intensity or prolonged exercise, by minimising disturbance of redox balance (Reid 2016b).
As a consequence, there is growing interest in the efficacy of fruit-derived polyphenol supplements, which possess antioxidant and anti-inflammatory properties, in improving exercise performance and/or tolerance both acutely (Cases et al. 2017; Deley et al. 2017; Oh et al. 2010; Trexler et al. 2014) and chronically (Braakhuis et al. 2014; Braakhuis and Hopkins 2015; Cook et al. 2015; Kang et al. 2012; MacRae and Mefferd 2006; Murphy et al. 2017; Sadowska-Krępa et al. 2008). Montmorency cherry polyphenols have been shown to enhance recovery of muscle strength following a bout of muscle-damaging exercise (Bowtell et al. 2011; Howatson et al. 2010). This enhanced exercise performance and functional recovery following muscle damage is likely to be mediated through the observed reduction in serum markers of oxidative damage after Montmorency cherry supplementation. These effects are suggested to be mediated via inhibition of superoxide producing enzymes such as NADPH oxidase or xanthine oxidase (Rodriguez-Mateos et al. 2013) or enhanced endogenous antioxidant capacity induced via nrf2 signalling (Huang et al. 2015). The resulting attenuation in superoxide exposure would reduce conversion of nitric oxide (NO) to peroxynitrite so preserving NO bioavailability during prolonged intense exercise (Benjamin et al. 1994; Cosby et al. 2003), and thus blood flow and tissue perfusion.
Polyphenol supplementation has also been implicated in increasing nitric oxide (NO) availability directly (Stoclet et al. 2004) by increasing the conversion of nitrite to NO (Rocha et al. 2009) as well as upregulating nitric oxide synthase (NOS, for review Galleano et al. 2010). Indeed, acute (Rodriguez-Mateos et al. 2013) and chronic (Khan et al. 2014) polyphenol supplementation has been linked to endothelium-dependent vasodilation. A meta-analysis found that supplementation with a mix of flavonoids increased flow-mediated dilatation (FMD) by 2.3% (based on 18 acute supplementation studies) and by 0.7% with chronic supplementation (based on 14 studies, Kay et al. 2012). This is likely to have a significant impact on exercise performance, during whole body exercise, where blood flow is considered to be a critical limiting factor to perfusion (Mortensen et al. 2008). Increased perfusion would result in increased tissue oxygen saturation and improved efflux of metabolic waste products such as lactate during exercise and subsequently enhance muscle function (Jacobs et al. 2011).
However, despite the potential for ergogenic effects of Montmorency cherry supplementation on exercise performance, very few studies have directly tested this hypothesis. Specifically, Montmorency cherry supplementation has recently been shown to enhance end-sprint cycling (Keane et al. 2018), aerobic running (Levers et al. 2016), and the recovery from prolonged, intermittent running performance (Bell et al. 2016) following a single, acute dose, of 7 or 8 days of supplementation, respectively.
The purpose of this study was to investigate the effect of 7-day Montmorency cherry supplementation on cycling time-trial (TT) performance. It was hypothesised that, compared to placebo, 7-day Montmorency cherry supplementation would: (1) enhance cycling TT performance, measured as a reduced time-to-complete the 15-km TT; (2) enhance tissue oxygenation during exercise, measured via the tissue oxygenation index (TOI); and (3) increase end-exercise capillary blood [lactate].
Materials and methods
Participants
Eight trained male competitive (> 250 miles/week) cyclists (mean ± SD: age: 19.7 ± 1.6 years, height: 1.79 ± 0.69 m, body mass: 75.0 ± 9.6 kg, \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\): 62.3 ± 10.1 ml kg−1 min−1, power output at \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\): 401 ± 38 W) volunteered and gave written informed consent to participate in this double blind crossover study, which had been approved by the University of Exeter Research Ethics Committee. A power analysis with an α error = 0.05, power = 0.95, and effect size = 2.89, was performed using the G × Power 3.1 analysis software (Heinrich Hein University, Duesseldorf, Germany), based on the effects of 7 days of blackcurrant supplementation on 16.1-km time-trial performance (Cook et al. 2015). This produced a minimum sample size of 4 participants. A total of eight participants were recruited to account for possible variation in the effects of Montmorency cherries and blackcurrants, and to maximise the statistical power for the secondary outcomes of TOI and other measures that may provide insight into the mechanisms of action. Participants reported to all testing sessions well-hydrated, having avoided strenuous exercise and caffeine ingestion for 24 and 3 h prior to testing, respectively. Participants were also instructed to consume their habitual diet and continue normal training activities for the first 5 days of the supplementation period but to refrain from strenuous physical activity for 48 h prior to the intensive exercise protocol. Participants recorded their diet and physical activity for 7 days prior to the cycling exercise trial (as described below) and then replicated this diet for the second, cross-over, trial. Testing was performed at the same time of day (± 2 h) for each subject.
Experimental design
Participants visited the laboratory on 4 occasions during a 4-week period. All participants completed: (i) an incremental test to exhaustion; (ii) two familiarisation 15-km cycling TTs; and (iii) two 15-km cycling TTs following 7-day supplementation of placebo and Montmorency cherry supplementation. Experimental tests were randomised and counter-balanced, separated by a minimum 2-week wash-out period and performed on a customised TT bike (Planet X, Sheffield, UK) that replicated the set-up (namely seat post and handlebars) of their own competition bike to maximise ecological validity. The bike was then loaded onto a static trainer to complete TT simulations within the lab (Kinetic Magnetic, Minneapolis, USA). Power output and work done (and distance) were measured via a mobile power meter integrated into the rear wheel (PowerTap G3 Hub, Madison, USA) connected wirelessly to a data logger (PowerTap Joule, Madison, WI, USA). The PowerTap G3 device was zeroed before each test. All laboratory-based tests were performed in similar environmental conditions (temperature, 18–20 °C; relative humidity, 45–55%). Participants were provided with feedback regarding the elapsed work done and distance completed as well as the work and distance remaining during the laboratory TTs at 5-km intervals.
Incremental test
On the first laboratory visit, participants performed a ramp-incremental cycling test for the determination of peak oxygen uptake (\(\dot {V}{{\text{O}}_{2{\text{peak}}}}\)) and calculation of a heavy-intensity work load for steady-state (SS) exercise on an electronically braked cycle ergometer (Corival, Lode BV, Groningen, The Netherlands). The test was preceded by 3 min of ‘baseline’ cycling at 20 W after which the workload increased to 75 W. The work load then continued to increase at a rate of 30 W min−1 until the limit of exercise tolerance. Participants pedalled at a self-selected cadence between 80 and 100 rpm. The test was terminated when cadence dropped by more than 10 rpm from the selected cadence for more than 10 s despite strong verbal encouragement. The \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\) was calculated as the highest 30-s average value attained before volitional exhaustion.
Experimental trial
Following familiarisation, participants completed two trials under the experimental conditions, placebo (PL) and Montmorency cherry supplementation (MC). To assess differences in muscle tissue oxygenation, initially, participants completed 3 min of baseline cycling (i.e., 20 W) before an abrupt increase to the work rate which corresponded to ~ 65% \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\) (steady-state exercise) for 10 min on a lode cycle ergometer. Following a 5-min period of rest, participants then completed a 15-km TT in the shortest time possible on a TT bike (as described above). Muscle tissue oxygen saturation, blood lactate (BLa) and pulmonary gas exchange (detailed below) were measured throughout.
Muscle oxygenation
For all experimental visits, muscle oxygenation status of the m. vastus lateralis of the right leg was monitored using near-infrared spectroscopy (Portamon, Artinis medical systems, The Netherlands). The CV of the device in our lab during unloaded cycling was recorded at 1.3 ± 0.5%. On arrival at the laboratory, the skin area underneath the near-infrared spectroscopy device was shaved, then exfoliated and cleaned with alcohol to minimise skin impedance. The sensor was placed at the midpoint between the lateral epicondyle of the femur and the femoral head. Adhesive tape and a hypoallergenic medical tape were used to ensure the sensor stability. An elastic bandage was wrapped around the participants’ leg and secured with adhesive tape to ensure the sensor did not move during exercise as well as minimising potential of extraneous light influencing the signal. The transmitted light was recorded at 10 Hz but down-sampled and exported at 1 Hz into proprietary software (Oxysoft, Artinis medical systems, The Netherlands). Tissue oxygenation index (i.e., TOI) was calculated as the percentage of total haemoglobin and myoglobin that was oxygenated. During SS exercise, TOI was averaged across the final 5 min of the 10-min bout. TOI was averaged in three work periods corresponding to 5-km intervals during TT exercise (i.e., 0–5 , 5–10 , 10–15 km) (Fig. 1).

Exemplar plot of TOI during steady-state (a) and time-trial (b) exercise. Tissue oxygenation index (i.e., TOI) was calculated as the percentage of total haemoglobin and myoglobin that was oxygenated. During steady-state exercise, TOI was averaged across the final 5 min of the 10-min bout. TOI was averaged in three work periods corresponding to 5-km intervals during TT exercise
Measurements
Throughout all tests, pulmonary gas exchange and ventilation was measured using an online gas analyser (Cortex Metalyzer 2R, Leipzig, Germany) with a CV of < 2% (Macfarlane and Wong 2012). A volume transducer was securely attached to a facemask and a capillary line was connected to the mask allowing inspired and expired gas volume and gas concentration to be collected. Known gas concentrations were used to calibrate gas analysers in line with manufacturer’s recommendations. The turbine volume transducer was calibrated using a 3-L syringe (Hans Rudolph, Kansas City, MO). Fingertip capillary blood samples were collected at baseline, post SS cycling and immediately after the completion of TT for the assessment of BLa using a portable lactate analyser (Lactate Pro, Arkray Inc, Japan). The CV using this device has been shown to be < 3.6% (Bonaventura et al. 2015).
Nutritional supplements
Each cherry capsule (CherryActive® Ltd, MC) consisted of finely powdered freeze-dried Montmorency cherry surrounded by a gelatine shell containing 0.3 g CHO and provided a total of 1.3 kCal. Tart cherry powder was analysed for total polyphenol content (Folin-Denis method) and anthocyanin content (high performance liquid chromatography, Atlas Bioscience Inc., Tucson, AZ, USA). Total polyphenol content provided by the 6 capsules, consumed daily (3 in the morning and 3 in the evening), was 462.8 mg day−1 and total anthocyanin content was 256.8 mg day−1. On day 7 (day of experimental testing), participants consumed 3 pills 60 min prior to the first experimental test. The optimal dose identified in a recent meta-analysis (Kay et al. 2012) was 500 mg per day total flavonoids or 300 mg per day of procyanidins, and the dose provided in the present study was based on this work. The placebo was made from dextrose powder inserted into gelatine capsules designed to have a similar appearance to cherry capsules but without the phytochemical content. To ensure compliance, in addition to recording diet and physical activity, participants were required to complete a 7-day supplementation record. This was then repeated in the crossover trial. For all participants, compliance was recorded at 100%.
Statistical analysis
Paired samples t test was used to compare mean \(\dot {V}{{\text{O}}_2}\), respiratory exchange ratio and TOI for the final 5 min of SS exercise between PL and MC. In addition, paired samples t test was used to assess the differences in completion times of the 15-km TT between conditions. Data normality assumptions were assessed using Kolmogorov–Smirnov test. Pearson’s product–moment correlation was used to examine the inter-relationship between TOI and the percentage of \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\) during SS exercise. Specifically, the correlation between the difference in TOI between conditions and the percentage of \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\) during SS exercise was calculated. Profiles of TT performance split times, \(\dot {V}{{\text{O}}_2}\) and TOI were analysed using two-way ANOVAs with repeated measures (condition [placebo vs. MC] × distance: [first, middle and final 5-km averages]). BLa was analysed using a two-way repeated-measures ANOVA with two timepoints (baseline and end-exercise). Where the ANOVA revealed a significant interaction effect, a post hoc t test was conducted using a Bonferroni correction. For calculation of effect size, partial eta squared (η2) was used for omnibus tests and Cohen’s d was used to calculate the effect size for paired t tests. In addition, Cohen’s d was corrected for the paired t test for dependence. Where sphericity was violated, a Greenhouse-Geisser correction factor was used. For all tests, results were considered statistically significant when P < 0.05. Data are presented as means ± SD unless otherwise indicated. All statistical analyses were conducted using IBM SPSS Statistics version 24.
Results
Physiological responses to steady-state (SS) exercise
The work rate required to elicit ~ 65% \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\) during SS exercise, determined during the preliminary incremental test, was 235 ± 38 W, and 141 ± 23 kJ of work was completed during SS exercise. The physiological responses to SS exercise for MC and PL can be seen in Table 1. Baseline TOI was significantly higher in MC (70.4 ± 2.3%) compared to PL (68.7 ± 2.1%, P = 0.02, d = 0.76, Table 1). In addition, whilst there was not a statistically significant difference in mean TOI during SS exercise following MC supplementation (57.4 ± 1.7 vs. 54.4 ± 6.9%, P = 0.06, d = 0.52), the difference in TOI between PL and MC trials was negatively correlated with SS relative exercise intensity (i.e., percentage of \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\), r = − 0.79, P = 0.02, Fig. 2). Baseline BLa was similar between trials (PL: 1.7 ± 0.4, MC: 1.5 ± 0.4 mM, P = 0.31, d = 0.38 Table 1). However, end-exercise BLa was significantly higher in MC (PL: 4.4 ± 2.1, MC: 6.7 ± 3.3 mM, P = 0.02, d = 0.86, Table 1). There was no difference in mean V̇O2 (PL: 3.1 ± 0.5 vs. MC: 3.1 ± 0.6 l min−1, P = 0.23, d = 0.10) or respiratory exchange ratio (PL: 0.96 ± 0.06, MC: 0.93 ± 0.07, P = 0.19, d = 0.49) between conditions.

Difference in tissue oxygenation index (TOI) between cherry and placebo trials during steady-state exercise for each participant. A significant correlation was found between the difference in TOI between trials and SS percentage of \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\) (r = − 0.79, r2 = 0.62, P < 0.05) with the cherry supplementation shown to yield larger changes in TOI compared to placebo trial at lower relative \(\dot {V}{{\text{O}}_{\text{2}}}\)
15-km cycling TT performance
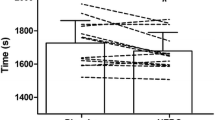
Mean completion time for the 15-km TT was 4.6 ± 2.9% (74 ± 50 s) faster in MC (1506 ± 86 s) compared to PL (1580 ± 102 s, P < 0.01, d = 0.78, Fig. 3). In addition, there was a significant interaction effect (distance × condition, P < 0.0001, η2 = 0.75) and the main effects of distance (P < 0.0001, η2 = 0.99) and condition (P = 0.02, η2 = 0.58) on time taken to complete each 5 km block of the TT, indicating improved performance following MC supplementation. Post hoc t test confirmed that there was a difference between conditions during the middle (i.e., 5–10 km) and final 5-km intervals (all P < 0.0001, Fig. 4). This improvement in mean group performance was consistent in all 8 participants, ranging from a 0.6 to 8.9% reduction in time to complete the 15-km TT (Fig. 3).

Mean completion time for the 15-km TT following placebo (dark grey) and cherry supplementation (grey). Completion time was significantly decreased following cherry supplementation (P < 0.05). Of the 8 participants, all 8 completed the TT in a quicker time following cherry supplementation compared to placebo, ranging from a 9 s (0.6%) improvement to a 155 s (8.9%) improvement. Asterisk significantly different to placebo (P < 0.05)

Mean ± SD completion time for the 15-km TT following placebo (dark grey) and cherry supplementation (grey) in 5-km intervals. Completion time was significantly decreased following cherry supplementation (P < 0.05). TT time was significantly different at 10- and 15-km time points. Asterisk significantly different to placebo (P < 0.05)
15-km cycling physiological responses
BLa was significantly increased after the TT (P < 0.0001, η2 = 0.90, Table 1). However, no interaction effect or main effect of condition were observed between MC (12.1 ± 4.2 mM) and PL (12.4 ± 3.9 mM, P = 0.82) immediately post TT. There was also no interaction effect or main effect of condition on TOI between MC (55.1 ± 7.7%) and PL (53.7 ± 7.2%, P = 0.70), despite observing a significant decrease in TOI during the TT (main time effect: P < 0.0001, η2 = 0.71). Similarly, analysis of the V̇O2 profile revealed no interaction effect or main effect of condition (mean \(\dot {V}{{\text{O}}_{\text{2}}}\) PL: 3.4 ± 0.6 l min−1 MC: 3.5 ± 0.5 l min−1P = 0.70). A main effect of time on \(\dot {V}{{\text{O}}_{\text{2}}}\) was observed during the TT (main time effect: P < 0.0001, η2 = 0.74), reflecting increase in oxygen consumption as the TT progressed. Asterisk significantly different from placebo (P < 0.05)
Discussion
The main novel finding of this study was that, consistent with our primary hypothesis, 15-km TT performance was improved following 7-day MC supplementation compared with PL in a group of trained male cyclists. This improvement in exercise performance was accompanied by a significant increase in resting (‘baseline’) m. vastus lateralis oxygenation.
In the current study, 15-km cycling TT performance was improved by ~ 4.6% following MC relative to PL. MC supplementation has been shown to decrease markers of inflammation and oxidative stress (Bell et al. 2014), as well as enhance recovery from muscle damage (Bowtell et al. 2011; Howatson et al. 2010). The magnitude of improvement in TT performance in our study is larger than that proposed as the “smallest worthwhile change” for road TT cycling (Paton and Hopkins 2006) and are similar to acute nitrate supplementation of similar TT duration (Lansley et al. 2011). However, it is pertinent to note that whilst the magnitude of improvement in mean group TT performance was larger than that proposed as the smallest worthwhile change, only 6 of 8 subjects experienced improvements in TT performance that exceeded the smallest worthwhile change. Thus, whilst all 8 subjects experienced an ‘improvement’ in TT performance, the ‘differences’ observed within 2 participants were within the day-to-day variability of the measurement. As a consequence, we only observed a medium effect size of supplementation on TT performance (d = 0.78).
However, our findings support recent observations on the potential of MC supplementation to improve exercise performance (Keane et al. 2018; Levers et al. 2016). Specifically, Keane et al. (2018) found a single acute dose of Montmorency cherry concentrate (60 ml, providing 73 mg cyanidin-3-glucoside per L) to increase peak power by 9.5% in trained endurance cyclists. Whereas, Levers et al. (2016), demonstrated a freeze-dried Montmorency cherry supplement (480 mg daily, ~ 990 mg polyphenols including ~ 66 mg anthocyanins, 7 days before, the day of, and 2 days after completing a half-marathon) to attenuate post-run markers of muscle catabolism and physiological stress (dampening of the inflammatory response and better maintenance of redox balance) in trained individuals. However, it should be noted that this study randomised participants into independent groups matched for reported race pace and thus did not perform a crossover design. In addition, a ‘non-significant trend’ (P = 0.09) was observed for an improvement in running performance (~ 13%) when compared to predicted race pace. In addition, many studies, including Keane et al. (2018), ask participants to adhere to a restricted diet in the days preceding the trials, highlighting the need for further work in investigating the potential synergetic effects of MC supplementation within habitual dietary practices. This is particularly important for athletes who typically have a diet high in polyphenols.
Our findings are in agreement with Cook et al. (2015) who reported enhanced 16.1-km cycling TT performance by ~ 2.4% following 7-day blackcurrant supplementation (providing 105 mg day−1 anthocyanins). A number of other studies have also observed enhanced exercise performance after acute polyphenol supplementation during, for example, treadmill running (ecklonia cava, Oh et al. 2010) and repeated all-out cycling (grape, pomegranate and green tea blend, Cases et al. 2017; pomegranate extract; Trexler et al. 2014). Chronic (7 day) blackcurrant polyphenol (105 mg anthocyanin, Cook et al. 2015; Perkins et al. 2015; Murphy et al. 2017) and 2-day epigallocatechin (EGCG, Richards et al. 2010) supplementation have also been shown to enhance exercise performance.
In contrast, a previous study has reported no improvement in 20-km TT performance in moderately trained individuals following MC supplementation despite a 3-day ingestion of dried Montmorency cherries which provided 216 mg of polyphenol of which the final dose was administered 2–3 h prior to the cycling trial (Clifford et al. 2013). In addition, not all studies have found ergogenic effects of polyphenol ingestion (Crum et al. 2017; Labonté et al. 2013). The absence of effect in these studies may relate to differences in dosing (i.e., 462 vs. 216 mg; 7 vs. 3 days, Clifford et al. 2013), training status (i.e., novice vs. athlete, Crum et al. 2017; Labonté et al. 2013), environmental conditions (i.e., high altitude, Crum et al. 2017), current antioxidant status (i.e., high vs. low, Green et al. 2004) as well as the timing of intake (i.e., 60 min vs. 2–3 h prior to exercise, Clifford et al. 2013) which may have marked effects on the ergogenic effects of polyphenol supplementation. The current investigation used MC powdered capsules in trained cyclists, using a TT scenario and a high polyphenol dose (462 mg polyphenols day−1) for 7 days, combined with a maintenance of habitual diet (i.e., with no polyphenol restrictions), thereby potentially explaining some of the differences observed between studies and maximising the ecological validity of our findings, respectively. In addition, the last supplement dose on day 7 was taken 60 min prior to exercise, since it has been observed that the endothelial-dependent vasodilatation response peaks approximately 60 min after ingestion of blueberry polyphenols (Rodriguez-Mateos et al. 2013). However, other studies have timed the polyphenol dose to coincide with peak plasma anthocyanin metabolite concentration which occurs 90–120 min following consumption (Keane et al. 2018).
We also observed a significant increase in baseline muscle oxygenation (i.e., TOI) with a medium effect size (d = 0.76), suggesting increased perfusion following MC supplementation. However, there was no statistically significant effect of MC on TOI during SS exercise (d = 0.52), which may indicate that the study was underpowered to detect an effect on TOI during exercise. There was a significant negative correlation in the difference in TOI between conditions with the percentage of \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\) suggesting that the increase in TOI after MC supplementation was more pronounced at lower exercise intensities. In agreement with this observation of increased perfusion with MC supplementation, enhanced vascular function has been implicated in other studies following polyphenol supplementation, which paralleled the ergogenic effects for performance (blackcurrant, Cook et al. 2015; pomegranate; Roelofs et al. 2017; pomegranate; Trexler et al. 2014). This suggestion of increased perfusion is corroborated by the elevated BLa in the cherry trial, which is unlikely to be due to increased lactate production, since oxygen consumption and the amount of work completed during 10-min SS cycle exercise at 65% \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\) were identical between trials. Rather, the elevated BLa is most likely due to increased lactate efflux from the muscle as a consequence of higher perfusion (Richardson et al. 1998). These observations are consistent with previous findings of reduced blood pressure following MC supplementation (Keane et al. 2018), providing further support that the enhanced performance might be mediated through the vasodilatory properties of polyphenol-rich MC.
These vasoactive properties of fruit-derived polyphenols have been also been demonstrated in resting conditions, with enhanced FMD evident after acute blueberry (Rodriguez-Mateos et al. 2013) and chronic blackcurrant (Khan et al. 2014) supplementation. The improvement in FMD is, by definition, a result of increased NO bioavailability, since FMD is NO dependent (Pyke and Tschakovsky 2005). It is likely that similar mechanisms underpin the enhanced muscle oxygenation observed in the present study and the improved arterio-venous difference postulated by others (Richards et al. 2010). However, Keane et al. (2018) recently demonstrated an improvement in end-sprint cycling performance despite no differences in plasma nitrite concentration between MC and PL suggesting that the improvements in performance with MC supplementation appear to be independent of NO-mediated signalling (and thus perfusion) and likely due to a reduction in ROS production. Although plasma nitrite may not be a sufficiently sensitive measure of NO in muscle (see Bryan and Grisham 2007 for review). However, our results do corroborate Keane et al. (2018) as, during all-out cycling where the participant has an ability to manipulate work output, we observed no differences in TOI.
During high-intensity exercise, excessive ROS production can lead to cellular damage and oxidative stress (Powers et al. 2004; MacRae and Mefferd 2006). There is evidence that acute (dark chocolate, Davison et al. 2012; black grape, raspberry and redcurrant polyphenol blend; Morillas-Ruiz et al. 2006; curcumin; Takahashi et al. 2014) and chronic (blueberry, McAnulty et al. 2011) polyphenol supplementation protects against endurance exercise-induced oxidative damage, but unfortunately these studies did not include an assessment of exercise performance. Whereas in the present study, whilst performance was enhanced, no measures of oxidative damage are available. However, MC concentrate has been shown to reduce oxidative damage after intense exercise (i.e., Bowtell et al. 2011; Howatson et al. 2010).
Previous literature has reported that the baseline antioxidant profile of an individual is an important determinant of the ergogenic effectiveness of an antioxidant intervention (Paschalis et al. 2018). In contrast to some other studies, no dietary restrictions to reduce polyphenol intake were imposed in the present study and ergogenic effects were nonetheless evident. The improvement in cycling performance in the current study could prove beneficial in elite sporting performance where athletes are attempting to find small but significant improvements in performance. Our findings are especially pertinent for highly trained individuals who demonstrate optimal NOS expression and high habitual dietary intakes of antioxidants to combat oxidative stress (Green et al. 2004).
In conclusion, 7-day Montmorency cherry powder supplementation enhanced 15-km cycling TT performance. This improvement in exercise performance seems to involve enhanced muscle perfusion as evidenced by increased muscle oxygenation presumably due to the vasoactive and anti-oxidative effects of the phytochemicals within the Montmorency cherries. The results of this study suggest that supplementation with MC concentrate might represent, a practical, non-pharmacological, dietary intervention to reduce enhance cycling performance in trained individuals. However, further research is required to investigate the dose–response between MC supplementation and cycling performance as well as the precise mechanisms responsible for this ergogenic potential, especially in the presence of a diet high in polyphenols.
Abbreviations
- BLa:
-
Blood lactate
- CV:
-
Coefficient of variation
- FMD:
-
Flow-mediated dilatation
- MC:
-
Montmorency cherry
- NO:
-
Nitric oxide
- PL:
-
Placebo
- ROS:
-
Reactive oxygen species
- SS:
-
Steady-state exercise
- TOI:
-
Tissue oxygenation index
- TT:
-
Time-trial
- \(\dot {V}{{\text{O}}_{2{\text{peak}}}}\) :
-
Peak oxygen uptake
References
Bailey DM, Lawrenson L, McEneny J, Young IS, James PE, Jackson SK, Henry R, Mathieu-Costello O, Richardson RS (2007) Direct evidence for exercise-induced free radical accumulation in human skeletal muscle. Free Radic Res 41:182–190. https://doi.org/10.1080/10715760601028867
Bell PG, Mchugh MP, Stevenson E, Howatson G (2014) The role of cherries in exercise and health. Scand J Med Sci Sports 24(3):477–490. https://doi.org/10.1111/sms.12085
Bell PG, Stevenson E, Davison GW, Howatson G (2016) The effects of montmorency tart cherry concentrate supplementation on recovery following prolonged, intermittent exercise. Nutr. https://doi.org/10.3390/nu8070441
Benjamin N, O’Driscoll F, Dougall H, Duncan C, Smith L, Golden M, McKenzie H (1994) Stomach NO synthesis. Nature 368(6471):502. https://doi.org/10.1038/368502a0
Bonaventura JM, Sharpe K, Knight E, Fuller KL, Tanner RK, Gore CJ (2015) Reliability and accuracy of six hand-held blood lactate analysers. J Sports Sci Med 14(1):203–214
Bowtell JL, Sumners DP, Dyer A, Fox P, Mileva KN (2011) Montmorency cherry juice reduces muscle damage caused by intensive strength exercise. Med Sci Sports Exerc 43(8):1544–1551. https://doi.org/10.1249/MSS.0b013e31820e5adc
Braakhuis AJ, Hopkins WG (2015) Impact of dietary antioxidants on sport performance: a review. Sports Med (Auckland NZ) 45(7):939–995. https://doi.org/10.1007/s40279-015-0323-x
Braakhuis AJ, Hopkins WG, Lowe TE (2014) Effects of dietary antioxidants on training and performance in female runners. Eur J Sport Sci 14(2):160–168. https://doi.org/10.1080/17461391.2013.785597
Bryan NS, Grisham MB (2007) Methods to detect nitric oxide and its metabolites in biological samples. Free Radic Biol Med 43:645–657
Cases J, Romain C, Marín-Pagán C, Chung LH, Rubio-Pérez JM, Laurent C, Gaillet S, Prost-Camus E, Prost M, Alcaraz PE (2017) Supplementation with a polyphenol-rich extract, PerfLoad®, improves physical performance during high-intensity exercise: a randomized, double blind, crossover trial. Nutr 9(4):421. https://doi.org/10.3390/nu9040421
Clifford T, Scott A, Mitchell N (2013) The influence of different sources of polyphenols on submaximal cycling and time trial performance. Int J Sport Nutr Exerc Metab 2(6):S10
Cook MD, Myers SD, Blacker SD, Willems MET (2015) New Zealand blackcurrant extract improves cycling performance and fat oxidation in cyclists. Eur J Appl Physiol 115(11):2357–2365. https://doi.org/10.1007/s00421-015-3215-8
Cosby K, Partovi KS, Crawford JH, Patel RP, Reiter CD, Martyr S et al Gladwin MT (2003) Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nat Med 9:1498–1505. https://doi.org/10.1038/nm954
Crum EM, Che Muhamed AM, Barnes M, Stannard SR (2017) The effect of acute pomegranate extract supplementation on oxygen uptake in highly-trained cyclists during high-intensity exercise in a high altitude environment. J Int Soc Sports Nutr. https://doi.org/10.1186/s12970-017-0172-0
Davison G, Callister R, Williamson G, Cooper KA, Gleeson M (2012) The effect of acute pre-exercise dark chocolate consumption on plasma antioxidant status, oxidative stress and immunoendocrine responses to prolonged exercise. Eur J Nutr 51(1):69–79. https://doi.org/10.1007/s00394-011-0193-4
Deley G, Guillemet D, Allaert F, Babault N (2017) An acute dose of specific grape and apple polyphenols improves endurance performance: a randomized, crossover, double-blind versus placebo controlled study. Nutr 9(8):917. https://doi.org/10.3390/nu9080917
Donato AJ, Uberoi A, Bailey DM, Wray DW, Richardson RS (2010) Exercise-induced brachial artery vasodilation: effects of antioxidants and exercise training in elderly men. Am J Physiol Heart Circ Physiol 298(2):H671–H678
Durand MJ, Dharmashankar K, Bian JT, Das E, Vidovich M, Gutterman DD, Phillips SA (2015) Acute exertion elicits a H2O2-dependent vasodilator mechanism in the microvasculature of exercise-trained but not sedentary adults. Hypertension 65:140–145. https://doi.org/10.1161/HYPERTENSIONAHA.114.04540
Galleano M, Pechanova O, Fraga G, C (2010) Hypertension, nitric oxide, oxidants, and dietary plant polyphenols. Curr Pharm Biotechnol 11(8):837–848. https://doi.org/10.2174/138920110793262114
Green DJ, Maiorana A, O’Driscoll G, Taylor R (2004) Effect of exercise training on endothelium-derived nitric oxide function in humans. J Physiol 561(1):1–25. https://doi.org/10.1112/jphysiol.2004.068197
Howatson G, McHugh MP, Hill JA, Brouner J, Jewell AP, Van Someren KA, Shave RE, Howatson SA (2010) Influence of tart cherry juice on indices of recovery following marathon running. Scand J Med Sci Sports 20(6):843–852. https://doi.org/10.1111/j.1600-0838.2009.01005.x
Huang Y, Li W, Su Z, yuan, Kong ANT (2015) The complexity of the Nrf2 pathway: Beyond the antioxidant response. J Nutr Biochem 26:1401–1413. https://doi.org/10.1016/j.jnutbio.2015.08.001
Jacobs RA, Rasmussen P, Siebenmann C, Díaz V, Gassmann M, Pesta D, Gnaiger E, Nordsborg NB, Robach P, Lundby C (2011) Determinants of time trial performance and maximal incremental exercise in highly trained endurance athletes. J Appl Physiol 111(5):1422–1430. https://doi.org/10.1152/japplphysiol.00625.2011
Jones AM (2014) Dietary nitrate supplementation and exercise performance. Sports Med (Auckland, NZ) 44(1): S35-45. https://doi.org/10.1007/s40279-014-0149-y
Kang SW, Hahn S, Kim JK, Yang SM, Park BJ, Chul LS (2012) Oligomerized lychee fruit extract (OLFE) and a mixture of vitamin C and vitamin E for endurance capacity in a double blind randomized controlled trial. J Clin Biochem Nutr 50(2):106–113. https://doi.org/10.3164/jcbn.11-46
Kay CD, Hooper L, Kroon PA, Rimm EB, Cassidy A (2012) Relative impact of flavonoid composition, dose and structure on vascular function: a systematic review of randomised controlled trials of flavonoid-rich food products. Mol Nutr Food Res 56(11):1605–1616. https://doi.org/10.1002/mnfr.201200363
Keane KM, Bailey SJ, Vanhatalo A, Jones AM, Howatson G (2018) Effects of Montmorency tart cherry (L. Prunus Cerasus) consumption on nitric oxide biomarkers and exercise performance. Scand J Med Sci Sports. https://doi.org/10.1111/sms.13088 (in press)
Khan F, Ray S, Craigie AM, Kennedy G, Hill A, Barton KL, Broughton J, Belch JJF (2014) Lowering of oxidative stress improves endothelial function in healthy subjects with habitually low intake of fruit and vegetables: a randomized controlled trial of antioxidant- and polyphenol-rich blackcurrant juice. Free Radic Biol Med 72:232–237. https://doi.org/10.1016/j.freeradbiomed.2014.04.006
Labonté K, Couillard C, Motard-Bélanger A, Paradis M-E, Couture P, Lamarche B (2013) Acute effects of polyphenols from cranberries and grape seeds on endothelial function and performance in elite athletes. Sports 1(3):55–68. https://doi.org/10.3390/sports1030055
Lansley KE, Winyard PG, Bailey SJ, Vanhatalo A, Wilkerson DP, Blackwell JR, Gilchrist M, Benjamin N, Jones AM (2011) Acute dietary nitrate supplementation improves cycling time trial performance. Med Sci Sports Exerc 43(6):1125–1131. https://doi.org/10.1249/MSS.0b013e31821597b4
Levers K, Dalton R, Galvan E, O’Connor A, Goodenough C, Simbo S, Mertens-Talcott SU, Rasmussen C, Greenwood M, Riechman S, Crouse S (2016) Effects of powdered Montmorency tart cherry supplementation on acute endurance exercise performance in aerobically trained individuals. J Int Soc Sports Nutr 13(1):22. https://doi.org/10.1186/s12970-016-0133-z
Macfarlane DJ, Wong P (2012) Validity, reliability and stability of the portable Cortex Metamax 3B gas analysis system. Eur J Appl Physiol 112(7): 2539–2547. https://doi.org/10.1007/s00421-011-2230-7
MacRae HSH, Mefferd KM (2006) Dietary antioxidant supplementation combined with quercetin improves cycling time trial performance. Int J Sport Nutr Exerc Metab 16:405–419
McAnulty LS, Nieman DC, Dumke CL, Shooter LA, Henson DA, Utter AC, Milne G, McAnulty SR (2011) Effect of blueberry ingestion on natural killer cell counts, oxidative stress, and inflammation prior to and after 2.5 h of running. Appl Physiol Nutr Metab 36(6):976–984. https://doi.org/10.1139/h11-120
Merry TL, McConell GK (2009) Skeletal muscle glucose uptake during exercise: a focus on reactive oxygen species and nitric oxide signaling. IUBMB Life 61:479–484. https://doi.org/10.1002/iub.179
Morillas-Ruiz JM, García V, López JA, Vidal-Guevara FJ, M. L., and Zafrilla P (2006) Effects of polyphenolic antioxidants on exercise-induced oxidative stress. Clin Nutr 25(3):444–453. https://doi.org/10.1016/j.clnu.2005.11.007
Mortensen SP, Damsgaard R, Dawson EA, Secher NH, González-Alonso J (2008) Restrictions in systemic and locomotor skeletal muscle perfusion, oxygen supply and V̇O2 during high-intensity whole-body exercise in humans. J Physiol 586(10):2621–2635. https://doi.org/10.1113/jphysiol.2007.149401
Murphy CA, Cook M, Willems ME (2017) Effect of New Zealand blackcurrant extract on repeated cycling time trial performance. Sports 5(25):2075–4663
Oh JK, Shin YO, Yoon JH, Kim SH, Shin HC, Hwang HJ (2010) Effect of supplementation with Ecklonia cava polyphenol on endurance performance of college students. Int J Sport Nutr Exerc Metab 20(1):72–79
Paschalis V, Theodorou AA, Margaritelis NV, Kyparos A, Nikolaidis MG (2018) N-acetylcysteine supplementation increases exercise performance and reduces oxidative stress only in individuals with low levels of glutathione. Free Radic Biol Med 1(115):288–297
Paton CD, Hopkins WG (2006) Ergometer error and biological variation in power output in a performance test with three cycle ergometers. Int J Sports Med 27(6):444–447. https://doi.org/10.1055/s-2005-865781
Perkins IC, Vine SA, Blacker SD, Willems MET (2015) New Zealand blackcurrant extract improves high-intensity intermittent running. Int J Sport Nutr Exerc Metab 25(5):487–493. https://doi.org/10.1123/ijsnem.2015-0020
Powers SK, Deruisseau KC, Quindry J, Hamilton KL (2004) Dietary AOXs and exercise. J Sports Sci 22:81–94
Pyke KE, Tschakovsky ME (2005) The relationship between shear stress and flow-mediated dilatation: Implications for the assessment of endothelial function. J Physiol 568:357–369
Reid MB (2016a) Reactive oxygen species as agents of fatigue. Med Sci Sports Exerc 48(11):2239–2246. https://doi.org/10.1249/MSS.0000000000001006
Reid MB (2016b) Redox interventions to increase exercise performance. J Physiol 594(18):5125–5133
Richards JC, Lonac MC, Johnson TK, Schweder MM, Bell C (2010) Epigallocatechin-3-gallate increases maximal oxygen uptake in adult humans. Med Sci Sports Exerc 42(4):739–744. https://doi.org/10.1249/MSS.0b013e3181bcab6c
Richardson RS, Noyszewski E, Leigh JS, Wagner PD (1998) Lactate efflux from exercising human skeletal muscle: role of intracellular PO2. J Appl Physiol 85:627–634
Rocha BS, Gago B, Barbosa RM, Laranjinha J (2009) Dietary polyphenols generate nitric oxide from nitrite in the stomach and induce smooth muscle relaxation. Toxicol 265(1–2):41–48. https://doi.org/10.1016/j.tox.2009.09.008
Rodriguez-Mateos A, Rendeiro C, Bergillos-Meca T, Tabatabaee S, George TW, Heiss C, Spencer JPE (2013) Intake and time dependence of blueberry flavonoid-induced improvements in vascular function: a randomized, controlled, double-blind, crossover intervention study with mechanistic insights into biological activity. Am J Clin Nutr 98:1179–1191. https://doi.org/10.3945/ajcn.113.066639
Roelofs EJ, Smith-Ryan AE, Trexler ET, Hirsch KR, Mock MG (2017) Effects of pomegranate extract on blood flow and vessel diameter after high-intensity exercise in young, healthy adults. Eur J Sport Sci 17(3):317–325. https://doi.org/10.1021/jf403757g
Sadowska-Krępa E, Kłapcińska B, Kimsa E, Karpińsk R (2008) Effects of supplemetation with red grape skin polyphenolic extract and interval swimming test on the blood antioxidant status in healthy men. Med Sport 12(1):1–7
Stoclet JC, Chataigneau T, Ndiaye M, Oak MH, El Bedoui J, Chataigneau M, Schini-Kerth VB (2004) Vascular protection by dietary polyphenols. Eur J Pharmacol 500:299–313
Takahashi M, Suzuki K, Kim HK, Otsuka Y, Imaizumi A, Miyashita M, Sakamoto S (2014) Effects of curcumin supplementation on exercise-induced oxidative stress in humans. Int J Sports Med 35(6):469–475. https://doi.org/10.1055/s-0033-1357185
Trexler ET, Smith-Ryan AE, Melvin MN, Roelofs EJ, Wingfield HL (2014) Effects of pomegranate extract on blood flow and running time to exhaustion. Appl Physiol Nutr Metab 39(9):1038–1042. https://doi.org/10.1139/apnm-2014-0137
Trinity JD, Broxterman RM, Richardson RS (2016) Regulation of exercise blood flow: role of free radicals. Free Radic Biol Med 98:90–102. https://doi.org/10.1016/j.freeradbiomed.2016.01.017
Acknowledgements
The supplements were provided gratis by Active Edge Ltd. We would like to express our gratitude to Dr Mary O’Leary for support with the production of figures.
Funding
This research was not supported by any funding body external to University of Exeter.
Author information
Authors and Affiliations
Contributions
MJB and JLB conceived and designed the research. MJB conducted all experiments. JB provided assistance with pilot testing prior to experimental data collection as well as supporting data analysis. P.M wrote the manuscript and supported data analysis. All authors contributed to the interpretation of results and read, edited and approved the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors state no conflict of interest in the publication of this manuscript.
Additional information
Communicated by William J. Kraemer.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
OpenAccess This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Morgan, P.T., Barton, M.J. & Bowtell, J.L. Montmorency cherry supplementation improves 15-km cycling time-trial performance. Eur J Appl Physiol 119, 675–684 (2019). https://doi.org/10.1007/s00421-018-04058-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-018-04058-6