
Abstract
Cardiac output (Q̇) is a primary determinant of blood pressure and O2 delivery and is critical in the maintenance of homeostasis, particularly during environmental stress. Cardiac output can be determined invasively in patients; however, indirect methods are required for other situations. Soluble gas techniques are widely used to determine Q̇. Historically, measurements during a breathhold, prolonged expiration and rebreathing to CO2 equilibrium have been used; however, with limitations, especially during stress. Farhi and co-workers developed a single-step CO2 rebreathing method, which was subsequently revised by his group, and has been shown to be valid (compared to direct measures) and reliable. Carbon dioxide output (V̇CO2), partial pressure of arterial CO2 (PaCO2), and partial pressure of mixed venous CO2 (PvCO2) are determined during 12–25 s of rebreathing, using the appropriate tidal volume, and Q̇ is calculated. This method has the utility to provide accurate data in laboratory and field experiments during exercise, increased and micro-gravity, water immersion, lower body pressure, head-down tilt, and changes in gas composition and pressure. Utilizing the Buffalo CO2 rebreathing method it has been shown that the Q̇ can adjust to a wide range of changes in environments maintaining blood pressure and O2 delivery at rest and during exercise.








Similar content being viewed by others


References
Arieli R, Boutellier U, Farhi LE (1985) Effect of water immersion on cardiopulmonary physiology at high gravity (+Gz). J Appl Physiol 61:1686–1692
Beltrami E (1987) Mathematics for dynamic modeling. Academia, San Diego, pp 257–261
Boutellier U, Farhi LE (1986a) Influence of breathing frequency and tidal volume on cardiac output. Respir Physiol 66:123–133
Boutellier U, Farhi LE (1986b) A fundamental problem in determining functional residual capacity or residual volume. J Appl Physiol 60:1810–1813
Boutellier U, Arieli R, Farhi LE (1985) Ventilation and CO2 response during +Gz acceleration. Respir Physiol 62:141–151
Buderer MC, Rummel JA, Savin C, Mauldin DG (1973) Use of single-breath method of estimating cardiac output during exercise-stress testing. Aerospace Med 44(7):756–760
Campbell EJM, Howell JBL (1960) Simple rapid methods of estimating arterial and mixed venous PCO2. Br Med J 1:458–462
Cassuto Y, Farhi LE (1979) Circulatory responses to arterial hypoxia. J Appl Physiol 46:973–977
Cerretelli P (1976) Limiting factors to oxygen transport on Mount Everest. J Appl Physiol 40:658–667
Cerretelli P, Cruz JC, Farhi LE, Rahn H (1966a) Determination of mixed venous O2 and CO2 tensions and cardiac output by a re-breathing method. Respir Physiol 1:258–264
Cerretelli P, Sikand R, Farhi LE (1966b) Readjustments in cardiac output during onset of exercise and recovery. J Appl Physiol 21(4):1345–1350
Cerretelli P, Di Prampero PE, Rennie DW (1970) Measurement of mixed venous oxygen tension by a modified re-breathing procedure. J Appl Physiol 28:707–711
Cerretelli PC, Marconi C, Dériaz O, Pendergast DR (1984) After effects of chronic hypoxia on cardiac output and muscle flow at rest and exercise. Eur J Appl Physiol 53:92–96
Chang C-M, Cassuto Y, Pendergast DR, Farhi LE (1994) Cardiorespiratory responses to lower body negative pressure. Aviat Space Environ Med 65:615–620
Christiansen J, Douglas CG, Haldane JS (1914) The absorption and dissociation of carbon dioxide by human blood. J Physiol (Lond) 48:244–271
Collier CR (1956) Determination of mixed venous CO2 tensions by re-breathing. J Appl Physiol 9:25–29
Cruz JC, Cerretelli P, Farhi LE (1967) Role of ventilation in maintaining cardiac output under positive-pressure breathing. J Appl Physiol 22:900–904
Cruz JC, Rahn H, Farhi LE (1969) Mixed venous PO2, (PCO2,) pH and cardiac output during exercise in trained subjects. J Appl Physiol 27:431–4 SAM-TR-70–5. Tech Report Sam-Tr:110–16
Davies CTM, di Prampero PE, Cerretelli P (1972) Kinetics of cardiac output and respiratory gas exchange during exercise and recovery. J Appl Physiol 35:618–625
DeFares JG (1958) Determination of PvCO2 from the exponential rise of CO2 during re-breathing. J Appl Physiol 13:159–164
Farhi LE (1982) Dilution methods for measurement of cardiac output: a review. In: Loeppky JA Riedesel ML (eds) Oxygen transport to human tissues. Elsevier, Amsterdam, pp 37–49
Farhi LE, Linnarsson D (1977) Cardiopulmonary readjustments during graded immersion in water at 35 degrees C. Respir Physiol 30:35–50
Farhi LE, Nesarajah MS, Olszowka AJ, Metildi LA, Elllis AK (1976) Cardiac output determination by a simple one-step re-breathing technique. Respir Physiol 28:141–159
Fenn WO, Dejours P (1954) Composition of alveolar air during breathholding with and without prior inhalation of oxygen and carbon dioxide. J Appl Physiol 7:313–319
Ferretti G (2001) Extreme human breath-hold Diving. Eur J Appl Physiol 84:254–271
Ferretti G, Boutellier U, Pendergast DR, Moia C, Minetti AE, Howald H, di Prampero PE (1990) Oxygen transport system before and after exposure to chronic hypoxia. Int J Sports Med 11 [Suppl. 1]:S15–S20
Ferretti G, Costa M, Ferrigno M, Grassi B, Marconi C, Lundgren CEG, Cerretelli P (1991) Alveolar gas composition and exchange during deep breath-hold diving and dry breath holds in elite dives. J Appl Physiol 70:794–802
Ferrigno M, Hickey D, Liner MH, Lundgren C (1987) CEG Simulated breath-hold diving to 20 meters: cardiac performance. J Appl Physiol 62:2160–2167
Ferrigno M, Grassi B, Ferretti G, Costa M, Marconi C, Cerretelli P, Lundgren C (1991) Electrocardiogram during deep breath-hold dives by elite divers. Undersea Biomed Res 18:81–91
Ferrigno M, Ferretti G, Ellis A, Warkander D, Costa M, Cerretelli P, Lundgren CEG (1997) Cardiovascular changes during deep breath-hold dives in a pressure chamber. J Appl Physiol 83:1282–1290
Gilbert R, Auchincloss JH (1971) Comparison of cardiovascular responses to steady- and unsteady-state exercise. J Appl Physiol 30:388–393
Grassi B, Ferretti G, Costa M, Ferrigno M, Panzacchi A, Lundgren CEG, Marconi C, Cerretelli P (1994) Ventilatory responses to hypercapnia and hypoxia in elite breath-hold divers. Respir Physiol 97:323–332
Haab P, Held DR, Ernst H, Farhi LE (1969) Ventilation-perfusion relationships during high-altitude adaptation. J Appl Physiol 26:77–81
Hlastala MP, Wranne B, Lenfant CJ (1972) Single breath method for measuring cardiac output – a reevaluation. J Appl Physiol 33:846–848
Hong SK, Ceretelli P, Cruz JC, Rahn H (1969) Mechanics of respiration during submersion in water. J Appl Physiol 27:535–538
Hook C, Meyer M (1982) Pulmonary blood flow, diffusing capacity, and tissue volume by rebreathing: theory. Respir Physiol 48:255–279
Hornbeck RW (1975) Numerical methods. Quantum, London, pp 194–195
Jansen RJ (1995) The thermal dilution method for the clinical assessment of cardiac output. Intensive Care Med 21:691–697
Jones NL, Campbell EJ, Edwards RH, Wilkoff WG (1969) Alveolar-to-blood PCO2 difference during rebreathing in exercise. J Appl Physiol 27:356–360
Knowles JH, Newman W, Fenn WO (1960) Determination of oxygenated mixed venous blood CO2 tension by a breathholding method. J Appl Physiol 15:225–228
Kim TS, Rahn H, Farhi LE (1966) Estimation of true venous and arterial PCO2 by gas analysis of a single breath. J Appl Physiol 21:1338–1344
Kurss DI, Lundgren CEG, AJ Pasche (1981) The effect of water temperature on vital capacity during head-out immersion. In: Bachrach AJ (ed) Underwater physiology, vol VII. Undersea Medicine Society, Kensington, Md., pp 297–301
Marquardt DW (1963) An algorithm for least squares estimating of nonlinear parameters. J Soc Ind Appl Math 11:431–444
Matalon SV, Farhi LE (1979) Cardiopulmonary readjustments in passive tilt. J Appl Physiol 47:503–507
Matalon SV, Dashkoff N, Nesarajah MS, Klocke FJ, Fahri LE (1982) Effects of hyperventilation on pulmonary blood flow and recirculation time of humans J Appl Physiol 52:1161–1166
Meade F, Pearl N, Saunders MJ (1967) Distribution of lung function (V̇ A/Q̇) in normal subjects deduced from changes in alveolar gas tensions during expiration. Scand J Respir Dis 48:354–365
Meyer M, Schuster KD, Schultz A, Mohr M, Piiper J (1990) Pulmonary diffusing capacities for nitric oxide and carbon monoxide determined by rebreathing in dogs. J Appl Physiol 68:2344–2357
Olszowka A, Farhi LE (1968) A system of digital computer subroutines for blood gas calculations. Respir Physiol 4:270–280
Pawelczyk JA, Levine BD, Prisk GK, Shykoff BM, Ellitt A, Rosow E (1995) Accuracy and precision of flight system for determination of cardiac output by soluble gas re-breathing. American Institute for Aeronautical and Astronomy meeting, Dallas, Tex.
Pendergast DR (1988) The effect of body cooling on oxygen transport during exercise. Med Sci Sports Exerc Suppl 20(5):S171–S176
Pendergast DR, Shindell D, Cerretelli PC, Rennie DW (1980) Role of central and peripheral circulatory adjustments in oxygen transport at the onset of exercise. Int J Sports Med 1(4):160–170
Pendergast DR, Olszowka AJ, Rokitka MA, Farhi LE (1988) Biomedical support of man in space. Acta Astronautica 17(2):187–193
Plewes JL, Olszowka AJ, Farhi LE (1976) Amount and rates of CO2 storage in lung tissue. Respir Physiol 28:359–369
Shykoff B, Farhi LE, Olszowka AJ, Pendergast DR, Rokitka MA, Eisenhardt CG, Morin RA (1996) Cardiovascular responses to submaximal exercise in sustained microgravity. J Appl Physiol 81(1):26–32
Sikand R, Cerretelli P, Farhi LE (1966) Effects of V̇ A, Q̇, and V̇ A/ Q̇ distribution and time on the alveolar plateau. J Appl Physiol 21:1331–1337
Szlyk PC, Evans KC, Sils IV (1988) Validation of a modified one-step re-breathing technique for measuring exercise cardiac output. Aviat Space Environ Med 59:1193–1197
Vogel JA, Hartley H, Cruz J (1974) Cardiac output during exercise in altitude natives at se levels and high altitude. J Appl Physiol 36:173–176
Acknowledgements
The experiments described in this paper were approved by the Institutional Review Board and all rules for the ethical treatment of subjects were followed. We wish to thank all of our co-investigators cited in the references and all the subjects that participated in the studies sited. The studies were funded by grants from the Dr. Henry C. and Bertha H. Buswell Fellowship Grant, National Institute of Health, National Aeronautic and Space Administration, Office of Naval Research, Naval Medical Research Command, Department of the Army, US Air Force, Air Force System Command, Aerospace Medical Division, National Oceanographic and Atmospheric Administration Office of Sea Grant.
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix A
Gas exchange relationships during a prolonged expiration
Let V̇ E represent the expired flow rate, V A the alveolar gas volume, and ELTV the volume of the lung CO2 stores. During an expiration, for CO2:
Based on observations that O2 stores are negligible Eq. A1 can be expressed for O2:
Taking into account the alveolar equations, dividing Eq. A1 by Eq. A2, and setting q=(V A+ELTV)/V A, after converting fractions to pressures, we get:
and then solving for R yields:
In practice q is set to 1, i.e. the effects of the lung CO2 stores are assumed to be insubstantial.
Appendix B
One-step rebreathing method
The analysis of the PCO2 versus time data obtained during the rebreathing involves a number of steps:
-
1.
Each breath starting with the last expiration preceding the respiratory maneuver is broken into four segments: a horizontal inspiratory plateau AB, a rising limb BC, an alveolar plateau CD, and a descending limb DA (Fig. 9). The course of end capillary PCO2 during inspiration is determined by joining each point D to point C on the following expiration. The rational for this step was based on the analysis of a computer simulation of the rebreathing maneuver (Plewes et al. 1976).
Fig. 9A, B. 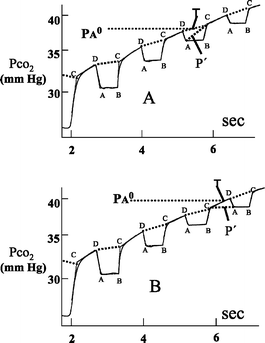
Analysis of tracings obtained during the one-step rebreathing method. The first part of the maneuver is not shown. PCO2 0 is obtained as shown in Fig. 1. A Time T occurs during inspiration and P′ is obtained by interpolation along a line drawn between point A and point C. B Time T occurs during expiration and P′ is obtained by interpolation along a line drawn between point C and point A
-
2.
Time T is the time at which PCO2 reaches PA0, the end tidal PCO2 of the last steady-state breath.
-
3.
Cc̄CO2 is obtained by determining the CCO2 (Olszowka and Farhi 1968) corresponding to the mean PCO2 on each segment between points D-C and C-D, by calculating the average of these values weighted by the duration of the segments.
-
4.
Computation of ΔVCO2 is somewhat complicated because T rarely occurs at FRC. P′ is the virtual partial pressure that would result if the difference in lung volumes at times T and 0 could be added to and mixed with the gas in the rebreathing bag. Because ΔVCO2 does not necessarily equal ΔVO2 the resulting volume does not have to equal VRB0, the initial bag volume. To compute P′ one would have to know the bag volume at time T as well as the lung volumes at times 0 and T. Instead P′ is obtained by interpolating along a line between points A and C or between points C and A depending on where T occurs in the respiratory cycle (Fig. 9).
CvCO2 is obtained from the asymptote of the PCO2 tracing vs. time. Analysis of changes in inspired (i.e. bag) and alveolar composition during rebreathing (see Appendix D and Fig. 7) predict that after the first four or five breaths the data can be fitted to the equations:
$$ {P{\rm{ACO}}_{{\rm{2}}} = P{\rm{eqCO}}_{{\rm{2}}} - A \cdot {\rm{e}}^{{ - \lambda \cdot t}} \;{\rm{and}}\;P{\rm{ACO2}} \cdot = P{\rm{eqCO}}_{{\rm{2}}} - B \cdot {\rm{e}}^{{ - \lambda \cdot t}} } $$(B1)

Previously the standard assumption had been that PeqCO2 equaled \( P{\text{vCO}}^{{{\text{ox}}}}_{{\text{2}}} \), the oxygenated venous CO2 tension. However, at equilibrium the continued uptake of oxygen causes some continued shrinkage in volume, which implies that at PeqCO2 must be greater than \( P{\text{vCO}}^{{{\text{ox}}}}_{{\text{2}}} \). Taking this into consideration and with suitable assumptions and algebraic manipulations the final expression of Eq. 8 in the text became:
in which, in addition to the terms already defined, CeqCO2 is the oxygenated blood CO2 concentration corresponding to PeqCO2, P b and t are the ambient pressure and temperature, and P H2O at time t the water vapor pressure at t.
Appendix C
Modified one-step rebreathing method
In general \( \dot{Q} \cdot {\left( {C{\text{vCO}}_{2} - C{\text{\={c}CO}}^{{\text{k}}}_{2} } \right)} \cdot \Delta T_{{\text{k}}} \) represents the CO2 excreted by the blood over the time interval ending at point C of the kth breath. For the breath that is closest to T, the time at which the PCO2 equals the PA0 as defined in Appendix B, the interval starts at the beginning of the rebreathing maneuver. For the subsequent breaths it starts at point C of the previous breath. See Fig. 10.

Analysis of tracings obtained during the modified one-step rebreathing method. Shown are examples of intervals in which CO2 exchange between blood, tissue and gas are computed
If the bag is completely emptied on each inspiration, FCO2 k, the CO2 fraction at point C of the breath, represents the CO2 fraction of all the gas in the system. For the breath closest to time T
represents the CO2 added to the gas during the period up to point C of that breath, while for subsequent breaths
represents CO2 added to the gas during the period between point C of breath k−1 and point C of breath k.
The difference between CO2 excreted by the blood and CO2 added to the gas phase represents the amount of CO2 added to the stores. Thus we may write:
For the breath closest to T, \( {\Delta F{\rm{CO}}^{{\rm{k}}}_{2} = F{\rm{CO}}^{{\rm{k}}}_{2} - F{\rm{A}}^{{\rm{0}}} } \) and for subsequent breaths \( {\Delta F{\rm{CO}}^{{\rm{k}}}_{2} = F{\rm{CO}}^{{\rm{k}}}_{2} - F{\rm{CO}}^{{{\rm{k}} - 1}}_{2} } \)
To obtain values for the gas volumes, the rebreathing bag initially is filled with a mixture of argon, nitrogen, and oxygen. Applying the ideas developed by Boutellier and Farhi (1986b) to compute FRC, the following equations express the gas exchange for those two gases during the time interval \( {T^{{\rm{k}}}_{{\rm{C}}} } \) starting at the beginning of the rebreathing maneuver and ending at point C each breath k:
These form a set of equations which with standard regression methods can yield the gas volume at point C of each breath. In these expressions \( {{\rm{ }}F{\rm{\bar{c}}}^{{\rm{k}}}_{{{\rm{N}}_{2} }} \;{\rm{and}}\;{\rm{ }}F{\rm{\bar{c}}}^{{\rm{k}}}_{{{\rm{Ar}}}} } \) are proportional to the mean blood concentration of each gas in the period up to the beginning of the kth expiration and are computed using the method used to obtain the mean blood CO2 concentration. Since the solubility coefficients αN2 and αAr are quite small, the first term of each equation is ignored to get a first estimate of the gas volumes needed for Eq. C4 below.
The remaining unknowns to be determined are Q̇, CvCO2, and ELTV. Since we have sufficient number of breaths, these can be obtained by a least-squares technique. To facilitate this computation the following form of Eq. C1 is used.
where 1/Q̇ instead of Q̇ is one of the unknowns to be determined. To further facilitate the computation one could lump the product of ELTV times 1/Q̇ into a single variable. Instead what is done is to assign a sequence of values to ELTV, computing for each ELTV a value for Q̇ and the corresponding least sum of squared errors, and then use the minimum of this set of least sum of squared errors to determine Q̇ and ELTV.
As noted above, in the initial sequence of computations the inert gas exchange is ignored. However once this yields a value for Q̇, that value can be inserted into Eqs. C2 and C3 and the computations repeated to improve the estimates of Q̇, CvCO2, and ELTV.
Appendix D
Estimating mixed venous CO2 tension by rebreathing
The following set of differential equations describing gas exchange between a homogenous lung and a bag during a rebreathing maneuver are similar to those published by Hook and Meyer (1982) and Meyer and coworkers (1990) to measure pulmonary blood flow as well as the diffusing capacity of nitric oxide and carbon monoxide. In the development that follows X′′ represents dX/dt, the rate of change of variable X per unit time.
For the alveoli:
where V L includes the effective lung tissue volume – ELTV – as well as the volume of gas present. V′ represents the rate of alveolar volume change and is non-zero since uptake of O2 does not equal the release of CO2.V̇ eff represents the effective ventilation between the lung and bag, and because of the dead space is less than the product of tidal volume and breathing frequency.
Assuming a linear dissociation curve with slope α over the sampled range of data and equilibrium between blood and gas, we have:
Substituting the above in Eq. D1, multiplying through it by P b−47, and rearranging terms:
Defining
and dividing through by V L, Eq. D2 takes the form:
Except for the inclusion of the term representing the rate of change of volume that occurs when the CO2 excretion does not equal O2 absorption, the above formulation is similar to that proposed by previous investigators. Note in particular that when the rate of volume change is ignored \( {P{\rm{eqCO}}_{{\rm{2}}} } \) equals \( P{\text{vCO}}^{{{\text{ox}}}}_{{\text{2}}} \).
Assuming that the net volume change for the gas in the system occurs only in the lungs, the formulation of the CO2 exchange in the bag is exactly like that proposed previously by Hook and Meyer (1982). Thus, for the rebreathing bag:
Determinations of the total gas volume changes during rebreathing indicate that a simplifying assumption of a constant rate of volume change is reasonable- see Fig. 8. Another simplification is that since the rebreathing period is short the gas volumes are constant. If rebreathing ventilation is considered continuous, we can treat the above pair of equations as a linear set of equations that can be solved by vector analysis (Beltrami 1987).
The general solutions are:
and
where
After the first four or five breaths the first exponential term becomes very small and the lung and bag concentration data can be fitted to the equations:
where λ=β2 and B/A=U 2.
As an aside, the above equations predict that when one plots \( {{\left( {P{\rm{eqCO}}_{{\rm{2}}} - P{\rm{ACO}}_{{\rm{2}}} } \right)}\;{\rm{and}}\;{\left( {P{\rm{eqCO}}_{{\rm{2}}} - P{\rm{BCO}}_{{\rm{2}}} } \right)}} \) versus time on a semi-log scale, after the first four or five breaths the bag and alveolar data will fall along two parallel lines whose slope is λ. See Fig. 7 in text. We now introduce a new parameter H which is set equal to B/A.
From the first part of Eq. D7:
From the second part of Eq. D7:
Substituting the above in Eq. D8 and rearranging terms yields:
The first two terms of the this expression for computing pulmonary flow are equivalent to formulations derived by Hook and Meyer (1982), but avoid the problem of how to partition the total gas volume into a lung and bag volume. The third term represents a correction term, which would be relatively small at low metabolic rates, for the volume changes that occur during the rebreathing period.
Rights and permissions
About this article
Cite this article
Olszowka, A.J., Shykoff, B.E., Pendergast, D.R. et al. Cardiac output: a view from Buffalo. Eur J Appl Physiol 90, 292–304 (2003). https://doi.org/10.1007/s00421-003-0921-4
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-003-0921-4