
Abstract
Plant ascorbate and glutathione metabolism counteracts oxidative stress mediated, for example, by excess light. In this review, we discuss the properties of immunocytochemistry and transmission electron microscopy, redox-sensitive dyes or probes and bright-field microscopy, confocal microscopy or fluorescence microscopy for the visualization and quantification of glutathione at the cellular or subcellular level in plants and the quantification of glutathione from isolated organelles. In previous studies, we showed that subcellular ascorbate and glutathione levels in Arabidopsis are affected by high light stress. The use of light-emitting diodes (LEDs) is gaining increasing importance in growing indoor crops and ornamental plants. A combination of different LED types allows custom-made combinations of wavelengths and prevents damage related to high photon flux rates. In this review we provide an overview on how different light spectra and light intensities affect glutathione metabolism at the cellular and subcellular levels in plants. Findings obtained in our most recent study demonstrate that both light intensity and spectrum significantly affected glutathione metabolism in wheat at the transcriptional level and caused genotype-specific reactions in the investigated Arabidopsis lines.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The increasing number of sudden drought, flooding or late frost events is challenging for current and future human food supply. Maize, rice and wheat are the most important staple foods worldwide (Pariona 2019). In 2019, 15% of the world’s wheat production areas did not receive enough irrigation water (Trnka et al. 2019). By the end of the twenty-first century, these authors extrapolated a high risk for simultaneous water shortages in up to 60% of the world’s wheat production areas, with the degree of risk dependent on mankind’s efforts to alleviate climate change.
Wheat, the model plant Arabidopsis thaliana and other C3 photosynthesis-type plants of temperate origin are highly susceptible to drought, which causes substantial yield losses in these plants due to the effects of osmotic and oxidative stress (Gill and Tuteja 2010). Plant organelles are differentially affected by oxidative stress. Excess levels of reactive oxygen species (ROS) as a consequence of intense and/or persistent abiotic or biotic stress frequently occur in the vicinity of electron transport chains. Consequently, in plants, thylakoid membranes in chloroplasts (Noctor et al. 2018) and the mitochondrial inner membranes (Huang et al. 2016), which are involved in intermediate steps of photosynthesis and respiration, respectively, are particularly prone to oxidative damage. At the cellular and subcellular levels, persistent oxidative stress damages RNA, DNA, pigments and proteins and causes the disintegration of membranes through lipid peroxidation, which eventually leads to de-compartmentation and cell death (Gill and Tuteja 2010; Karuppanapandian et al. 2011; Dorion et al. 2021). The oxidative stress response in plants is therefore also cell compartment specific (Zechmann 2014, 2020). In this context, it is highly important to gain a better understanding of the complex and finely tuned regulatory and signaling networks functioning in plant stress response at the subcellular level.
The redox state in plants results from a combined effect of environmental conditions and genetic regulation. Plants can adapt their growth and development to mild environmental changes and/or seasonal or annual alterations, whereas sudden and/or strong environmental changes over an extended period of time may lead to tissue damage and death (Kocsy et al. 2013). The growth and development of plants is driven by photosynthesis, which is most efficient within the blue and red spectrum of light. During photosynthesis, electrons are transferred from water via an electron transfer chain to nicotinamide adenine dinucleotide phosphate (NADP+), using energy absorbed from sunlight. The electrochemical gradient between the thylakoid lumen and the stroma initiates a proton gradient that facilitates the phosphorylation of adenosine diphosphate (ADP). The products of this phosphorylation, adenosine triphosphate (ATP) and reduced NADP+ (NADPH), are used for carbon fixation and the incorporation of carbon into organic compounds. The accumulation of ROS depends on a functional redox-homeostasis in the chloroplast and other cell compartments. For example, superoxide radicals (O2−) are generated by the Mehler reaction from oxygen and two O2− radicals, and are subsequently dismutated by superoxide dismutase (SOD; EC 1.15.1.1) to hydrogen peroxide (H2O2) and oxygen. If the light energy absorbed exceeds the scavenging capacity for ROS, irreversible damage from singlet oxygen (1O2) in photosystem II (PS II) results in photoinhibition. Carotenoids are involved in quenching 1O2 and xanthophyll carotenoids facilitate the dissipation of excess absorbed light energy as heat (non-photochemical quenching). H2O2 and other ROS detoxification in chloroplasts is mediated by the ascorbate–glutathione (or Foyer-Halliwell-Asada) cycle or by redox transmitters (peroxiredoxins).
Due to their sessile nature, plants are required to develop a fast and versatile response system to fluctuating and/or excess light intensity (Scheibe and Dietz 2012; Ding et al. 2016; Mullineaux et al. 2018; Turkan et al. 2018). Interestingly, in one study, stress signaling transcripts strongly reacted to a single excess light trigger at 1000 µmol m−2 s−1 photon flux density in non-primed Arabidopsis plants, while the same transcripts in primed plants (repeatedly pre-treated with excess light at 1000 µmol m−2 s−1) remained largely unaffected. Ganguly et al. (2019) concluded that primed Arabidopsis plants were more tolerant to excess light compared to non-primed plants, due to adjustments in the efficiency of photosynthetic electron transport and because priming was independent from functioning DNA methylation. Sudden fluctuations between full sunlight and shade interferes with the excitation state (and thus redox-homoeostasis) of the two photosystems and causes ROS accumulation along the electron transport chain. Plants are able to re-arrange the antenna structure of the photosystems (state transition) in chloroplasts within minutes. Given that chlorophyll and carotenoids primarily absorb in the red spectrum, but not in the far-red light spectrum, understory plants or leaves of lower nodes on a plant need long-term adaptation strategies for efficient electron transport during photosynthesis. Long-term (hours to days) response to low red/far-red (shade) conditions involves adaptations in photosystem stoichiometry (Dietzel et al. 2008). In Phaseolus vulgaris plants cultivated under a low red/far-red ratio (0.2 instead of the normal 1.1), which resembles light availability to ‘shade’ leaves, leaf ascorbate (Asc) and glutathione contents, the activity of antioxidant enzymes and respiration rates indeed were lower compared to the control white light regime (Bartoli et al. 2009). Excess ROS may, however, be formed in seedlings grown under continuous far-red light because the reduction of protochlorophyllide to chlorophyll is impaired by the limited ability of protochlorophyllide to absorb far-red light and by the transcriptional repression (mediated by phytochrome A [phyA]) of the oxidoreductase involved (Runge et al. 1996; Sheerin and Hiltbrunner 2017).
Light is also involved in the signal transduction of morphogenesis (plant elongation, leaf expansion), stomatal opening, the circadian clock and flowering, all of which are sensed by wavelength-specific photoreceptors. The following photreceptors have been identified in plants: for the UV/A-blue wavelengths, the cryptochromes (cry), phototropins and members of the Zeitlupe family; for the red and far-red wavelengths, phytochromes; and for UV/B, UV RESISTANCE LOCUS8, a UV-B photoreceptor, which also reacts to short wavelength UV/A (Rai et al. 2021; Kami et al. 2010; Chen et al. 2004). Far-red light is sensed by phyA and is involved in the regulation of plant development and growth by inducing phyA or repressing phytochrome B, whereas changes in the ratio of red/far-red light are primarily sensed by phytochrome B, which mediates shade response or ‘shade avoidance syndrome’ (Sheerin and Hiltbrunner 2017; Viczián et al. 2017). Phytochromes and cryptochromes are associated with phytochrome-interacting factors that mediate downstream gene regulation (Kianianmomeni 2014; Pedmale et al. 2016; Pham et al. 2018). The subcellular localization of photoreceptors is light dependent. While most of the Arabidopsis phytochromes have been localized in the cytosol in the dark, under light conditons they are translocated into the nucleus. One exception is cry1, which is localized in the nucleus in the dark, but mainly in the cytosol in the light (Chen et al. 2004) and activated by light-dependent changes in the redox state of a cofactor (flavin) (Bouly et al. 2007).
Prevention of oxidative damage in plant organelles during osmotic stress
On hot, dry days C3-photosynthesis-type plants like wheat, rice or Arabiopsis close their stomata to prevent water loss. Under sunny conditions, photons are highly abundant, but closing the stomata limits gas exchange and hence the availability of the electron acceptors ADP+ and NADP+ from the Calvin–Benson cycle. To prevent thylakoid electron transport chain overload, photorespiration is induced, which on the one hand supplies ADP+ and NADP+ and on the other hand regenerates 3-phosphoglycerate from 2-phosphoglycolate. However, the photorespiratory cycle in peroxisomes, which allows the regeneration of glycolate from photorespiration for primary metabolism, produces H2O2, which serves a signaling function at low concentrations, but is toxic at high concentrations and is balanced via Asc, glutathione and the enzyme catalase (CAT; EC 1.11.1.6). In mitochondria, ROS, in particular O2−, accumulate under conditions of limited ADP. However, under excess light, the respiratory rate increases, which in turn aids photochemical quenching, and an alternative oxidase isoform (AOX) is expressed, which limits the energy efficiency, thereby avoiding intermediate ROS formation (Gill and Tuteja 2010; Exposito-Rodriguez et al. 2017; Mullineaux et al. 2018).
Plants are able to counteract oxidative stress, with redox changes triggering and modifying physiological processes. ROS, reactive nitrogen and reactive sulfur species together with antioxidants play a pivotal regulatory role in these adaptation and defense mechanisms at the transcriptome, proteome and metabolome levels. Asc and glutathione are the most important antioxidants in plants, given their functions as coenzyme and posttranslational modifier, respectively (Foyer and Noctor 2005; Noctor et al. 2012; Kocsy et al. 2013; Olson 2020). In plants, the concerted action of Asc, glutathione and other non-proteinaceous (NADP+/H) and proteinaceous (ascorbate peroxidase [APX], EC 1.11.1.11; monodehydroascorbate reductase [EC 1.6.5.4]; dehydroascorbate reductase [EC 1.8.5.1; glutathione reductase [EC 1.8.1.7]) components of the ascorbate–glutathione cycle (or Foyer-Halliwell-Asada cycle) allows the detoxification of excess H2O2 (Foyer and Halliwell 1976; Asada 1999; Foyer and Noctor 2011). The pool of reduced glutathione (GSH) is fueled by glutathione reductase (GR) activity, which catalyzes the reduction of glutathione disulfide (GSSG) to two molecules of GSH by electron transfer from NADPH. Compartment-specific GRs are found in the cytosol, chloroplasts, mitochondria and peroxisomes (reviewed by Csiszár et al. 2016). Arabidopsis GR2, located in the chloroplast and mitochondria, helps to maintain the function of PS II under excess light conditions by increasing the level of glutathione (Ding et al. 2016). The lack of reducing power in GR2 null mutants can be partially complemented by the ATP-binding cassette transporter and thioredoxin system in mitochondria, but not in chloroplasts (Marty et al. 2019). Similarly, GR1 knockout lines of the model moss Physcomitrella patens are not able to maintain a full reducing environment in the chloroplast stroma under excess light (Müller-Schüssele et al. 2020). An increase in the amount and ratio of GSSG and GSH with increasing light intensity has been reported for Arabidopsis (Heyneke et al. 2013; Choudhury et al. 2018; Gasperl et al. 2021) and wheat (Gasperl et al. 2021). Excess light at 500 µmol m−2 s−1 was found to induce cysteine and glutathione synthesis and glutathione reduction in wheat at the transcriptional level (Monostori et al. 2018; Gyugos et al. 2019; Toldi et al. 2019; Gasperl et al. 2021). Glutathione synthesis is ATP-consuming and depends on the availability of the amino acids cysteine, glutamine and glycine and requires two subsequent reactions, in which γ-glutamylcysteine synthetase (γ-ECS or GSH1, EC 6.3.2.2) first catalyzes the formation of γ-glutamylcysteine (γ-EC) from cysteine and glutamine and glutathione synthetase (GSHS or GSHS2, EC 6.3.2.3), then catalyzes the formation of glutathione from γ-EC and glycine. While the first step of glutathione synthesis is restricted to chloroplasts in Arabidopsis and wheat, the second step is primarily confined to the cytosol (Noctor et al. 2012). Koffler et al. (2011) found glutathione synthesis in Arabidopsis to be limited to the availability of γ-EC in chloroplasts and the cytosol. In a drought-tolerant wheat variety (Plainsman), the recovery photosynthesis rate and recovery growth rate improved with increasing light intensity, which may have partly been due to a higher cysteine availability for sufficient glutathione synthesis after drought stress, evident from the increase in cysteine and glutathione levels and expression of the glutathione synthetase gene GSHS2 (Gyugos et al. 2019). Interestingly, shade pre-treated (low red/far-red light ratio of 0.4 ~ 0.6) soybean seedlings likewise coped better with drought by an improved ROS scavenging system (Asghar et al. 2020). Glutathione further plays an important role in the detoxification of xeno- or endobiotics, where glutathione transferases (GTs, EC 2.5.1.18) catalyze the conjugation of glutathione to electrophilic compounds, which facilitates transport into the vacuole for metabolization (Labrou et al. 2015). GTs in plants are grouped into the cytosolic, mitochondrial and microsomal super families. Certain types of GTs have antioxidant properties, such as dehydroascorbate reductase or glutathione peroxidases, and others are involved in hormone signaling (Noctor et al. 2012; Csiszár et al. 2016). Darkness, low light or shade mostly reduce the activity of GTs, while high light intensity elevates the activity and gene expression of GTs (Gallé et al. 2018).
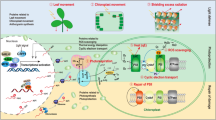
Subcellular detoxification model for excess H2O2 in Arabidopsis under excess light or lower red/far-red ratio and regulation of antioxidants by light intensity and spectrum composition
Increased H2O2 formation was sensed in chloroplast stroma, the cytosol and nuclei of genetically modified tobacco HyPer2 (fluorescent H2O2 biosensor) plants cultivated in excess light at 1000 µmol m−2 s−1 for 1 h. Given that chloroplast-derived H2O2 is detected in nuclei, chloroplast-derived H2O2 may facilitate a fast response to changes in light intensity at the gene expression level (Exposito-Rodriguez et al. 2017). It was proposed that excess H2O2 from overstrained electron transport chains in Arabidopsis chloroplasts and from photorespiration in peroxisomes may leak into the cytosol and vacuole with increasing light intensity. Excess H2O2 can be detoxified by Asc, GSH, CAT and APX activity in peroxisomes and the cytosol, by Asc, GSH and APX activity in chloroplasts, while in vacuoles only Asc is available as a reductant and guaiacol-type peroxidase (EC 1.11.1.7) activity aids in scavenging excess H2O2 (Fig. 1) (Takahama 2004; Heyneke et al. 2013; Zipor and Oren-Shamir 2013; Gasperl et al. 2021).

Adapted model of subcellular reactive oxygen species (ROS) accumulation and detoxification by antioxidants and catalase in plants under conditions of excess light or white light with lower red/far-red ratio (graph reproduced using Corel® Photo Paint 2019 [Corel Corp., Ottawa, ON, Canada] with modifications from Heyneke et al. 2013). Line drawing proposing a model of the effects of high light stress or of a decreased red/far-red ratio on the subcellular accumulation of ROS in Arabidopsis thaliana and wheat with special focus on the compartment-specific detoxification of hydrogen peroxide (H2O2) by ascorbate (Asc), catalase (Cat) and reduced glutathione (GSH). Excess light stress (indicated by white thunderbolt) induces the generation of ROS and H2O2 in chloroplasts (C) and in peroxisomes (Px) by overstraining the electron transport chain in thylakoids (green ovals inside the chloroplast) and through photorespiration, respectively. Asc, Cat and GSH detoxify and suppress the accumulation of ROS and H2O2 in these cell compartments. Accumulation of H2O2 (detected by cerium chloride) and subcellular antioxidants (detected via immunolabeling) by excess light in Arabidopsis and wheat is indicated by a white upwards arrow. White light with decreased red/far-red ratio (indicated by pink thunderbolt) induces the generation of ROS and H2O2 in chloroplasts of seedlings compared to seedlings grown in white light with normal red/far-red ratio by inefficient chlorophyll biosynthesis (impaired reduction of protochlorophyllide to chlorophyllide by protochlorophyllide oxidoreductase [POR] activity). With increasing stress, including high light intensities or shade (limited reducing power and assimilates), H2O2 leaks from chloroplasts and peroxisomes into the cytosol and eventually into vacuoles. In shaded leaves (lower red/far-red ratio), an imbalance in the excitation of photosystems II and I induces ROS and H2O2 formation in chloroplasts. Accumulation of subcellular glutathione (detected via immunolabeling) by a decreased red/far-red ratio in the Arabidopsis pad2-1 mutant and wheat is indicated by a pink upwards arrow. Arabidopsis wild-type and vtc2-1 mutant cell compartments were largely unaffected by a decreased red/far-red ratio. Whereas Asc, Cat and GSH detoxify H2O2 also in the cytosol, only Asc is involved in the detoxification of H2O2 in vacuoles (V), where it helps to reduce phenoxyl radicals created by oxidation of phenols by H2O2. H2O2 was detected after short-term (4 h) exposure to high light stress, but was successfully detoxified in the long term (2 weeks). H2O2 and Asc were not analyzed (n.a.) under a decreased red/far-red ratio (Runge et al. 1996; Takahama 2004; Dietzel et al. 2008; Scheibe and Dietz 2012; Kim and Apel 2013; Sheerin and Hiltbrunner 2017; Gasperl et al. 2021)
Indeed, short-term (4 h) excess light at 1500 µmol m−2 s−1 led to the accumulation of H2O2 in the cytosol and along the tonoplast, but was absent in cell compartments after 2 weeks of excess light treatment (Heyneke et al. 2013). This detoxification model for excess ROS and H2O2 in Arabidopsis is based on findings derived from high light intensity treatments using compact fluorescent lamps (Plug and Grow, 6400 K, white/blue spectrum; Agriculture Trading AG, Walenstadt, Switzerland), at 1500 µmol m−2 s−1 for 4 h, which resulted in the accumulation of H2O2 in the cytosol and along the tonoplast in vacuoles, increased density of immunolabeled Asc and total glutathione (= sum of GSH and GSSG) in chloroplasts, peroxisomes and the cytosol, degradation of thylakoid membranes (plastoglobuli formation) and an overall increase in H2O2 level and CAT activity in the leaves after 14 days of treatment. Interestingly, the Arabidopsis lines showed a genotype-specific adaptation to 14 days of excess light. The glutathione-deficient Arabidopsis mutant pad2-1 ( seemed to compensate its lower level of total glutathione by accumulating more Asc in peroxisomes, whereas the opposite was true for the Asc-deficient Arabidopsis mutant vtc2-1, which showed an increase in chloroplast and nuclei total glutathione content (Heyneke et al. 2013). In our most recent studies (Monostori et al. 2018; Toldi et al. 2019; Gasperl et al. 2021), we used continuous wide-spectrum light-emitting diodes (LEDs) to manipulate the light intensity (Lumileds LXZ-5790y; Philips, Amsterdam, The Netherlands) and spectrum composition (narrow-spectrum LEDs with dominant wavelengths of 448 nm [Lumileds LXZ1-PR01; Philips], 665 nm [Lumileds LXZ1-PA01; Philips] and 750 nm [2ER101FX0000001; Edison Edixeon, Moers, Germany). Under white light with lower red/far-red ratio, we propose an increase in ROS and H2O2 in chloroplasts followed by an increase in these molecules in other subcellular compartments due to inefficient chlorophyll biosynthesis in seedlings after the transition to white light. In mature shaded leaves, an imbalance in the excitation states of the two photosystems may increase ROS and H2O2 in chloroplasts. With increasing stress from a limited supply of reducing power (NADPH, ATP) and assimilates, H2O2 may subsequently accumulate in other cell compartments. These compartment-specific redox changes induce the accumulation and redistribution of antioxidants and associated enzyme activities (see Fig. 1; Runge et al. 1996; Dietzel et al. 2008; Scheibe and Dietz 2012; Kim and Apel 2013; Sheerin and Hiltbrunner 2017; Gasperl et al. 2021). LEDs have gained more and more importance as energy-efficient plant growth light sources during the last years and have been used to study the involvement of light, with special focus on red and blue spectra, in the regulation of growth and development of the model plant Arabidopsis thaliana and various crop plants; in this context, the enhanced accumulation of specific metabolites is also of great interest (Li et al. 2017; Monostori et al. 2018; Kong and Zheng 2020; Jenkins 2021; Samuolienė et al. 2020; Appolloni et al. 2022).
Effect of light intensity and spectrum on genes encoding compartment-specific enzymes of glutathione metabolism
Changes in light intensity or spectrum may influence the level of total glutathione and its reduction state (GSSG/GSH) through modification of the activity of the proteins and the expression of the genes associated with glutathione metabolism and transport in the individual organelles. Regarding glutathione synthesis, its first step, the formation of γ-EC takes place in the chloroplasts and its transcriptional regulation by light intensity (higher expression under conditions of low light intensity than under normal light intensity) and far-red light (lower expression under white light with decreased red/far-red ratio) was shown in wheat (Gasperl et al. 2021). The expression of the two genes encoding the enzyme of the second step of glutathione synthesis (GSHS) in the cytosol and chloroplast increased with increasing light intensity, and this change was accompanied by an increase in total glutathione contents in these cell compartments. The expression of the gene encoding the chloroplastic form of this enzyme was one magnitude lower than that of the gene of the cytoplasmic form. Among the genes associated with GSSG reduction, the expression of the gene of cytosolic GR was also increased under conditions of high light intensity, but not in far-red light conditions, in wheat (Gasperl et al. 2021). These observations indicate that light conditions may also control the glutathione-dependent subcellular redox environment at the transcriptional level.
The tissue- and cell compartment-specific enzyme activity and gene expression of GTs is also influenced by both light intensity and quality (Chen et al. 2007; Gallé et al. 2018). A tau class GT, the FIN219-interacting protein 1 (FIP1), which interacts with Far-Red Insensitive 219 (FIN219), is involved in the phytochrome A-mediated signaling in Arabidopsis (Chen et al. 2007). This process in turn regulates cell elongation and flowering in response to light. FIP1gene transcription was induced by far-red light and the FIP1 protein was localized in the nucleus and cytoplasm. Thus, light spectrum-dependent changes in its activity may influence glutathione content in these compartments. Another GT gene, AtGSTU17, also participates in the signaling process associated with phytochrome A and controls the size and redox state of the glutathione pool in Arabidopsis, which in turn affect development (Jiang et al. 2010). The effect of blue and red light on the expression of GT and other genes encoding antioxidant enzymes was also shown in leaf tissues of maize seedlings (Liu and Zhang 2021). Glutathione contents and GR activities increased after long-term exposure to a combination of continuous red and blue light in lettuce (Zha et al. 2019). Unfortunately, in the latter studies, the effect of spectrum on genes encoding the isozymes in various organelles was not investigated, but their specific and different regulation can be assumed based on the observations in Arabidopsis and wheat (Chen et al. 2007; Gasperl et al. 2021).
Quantification of glutathione in plant cell compartments
Methods for quantifying glutathione in plant cell compartments were concisely evaluated by Zechmann in 2014. Each of the approaches evaluated here has its own specific advantages and limitations, which are summarized in Table 1. Additional details on targets for redox-sensitive green fluorescence protein (roGFP) are included from Bratt et al. (2016), as are additional details on glutathione quantification from isolated organelles from Hajdinák et al. (2019) and Pradedova et al. (2019). Briefly, subcellular glutathione can be bound in situ or in vivo to glutathione-specific antibodies (total glutathione = sum of GSH and GSSG), redox-sensitive dyes (monobromobimane [mbrB] or monochlorobimane [mchlB]; GSH) or roGFP probes (GSH, GSSG), visualized by transmission electron microscopy (TEM), bright-field microscopy, confocal-laser scanning microscopy (CLSM) or fluorescence microscopy, respectively, and quantified by software-assisted image analysis. Total glutathione localization by use of a specific antibody (henceforth referred to as immunolabeling) on ultrathin sections (80 nm) of embedded samples preserves the current physiological state with respect to subcellular total glutathione distribution and labeling density in situ. Immunolabeling of total glutathione is possible in all plant cell compartments in which glutathione either plays a major role as an antioxidant (chloroplasts, the cytosol, mitochondria, peroxisomes, nuclei) and/or where glutathione is synthesized (chloroplasts, the cytosol) and/or reduced (chloroplasts, the cytosol, mitochondria, peroxisomes). Tolin et al. (2013) verified the accumulation of glutathione with this method in the apoplast of the Arabidopsis ggt-1 mutant (GGT-1 knockout), which lacks glutathione degradation (by gamma-glutamyl transferase/transpeptidase [GGT-1]) in the apoplast. In the study of Queval et al. (2011), the accumulation of glutathione was detected in vacuoles of the catalase-deficient Arabidopsis cat2 mutant. These results indicate that in Arabidopsis wildtype plants, glutathione levels are either not present or the level is too low to be detected with immunolabeling methods in vacuoles and the apoplast.
Further, deep cell layers, such as the mesophyll or vascular tissue, are accessible and the high resolution (~ 0.2 nm) of a TEM allows the visualization of even local, subcompartmental changes. The resolution limit for immunolabeling of total glutathione, however, is > 10 nm due to the size of the complex formed by glutathione, the primary (anti-glutathione rabbit polyclonal immunoglobulin G [IgG]) antibody and the secondary (goat anti-rabbit IgG) antibody which is conjugated to a gold particle of 10 nm in diameter (Zechmann et al. 2008a). Tissue samples of 1.5 mm2 are of sufficient size for study, thus saving plant material for additional analyses (such as studies on gene expression and metabolite composition). However, the immunolabeling approach requires careful sample fixation and is momentarily limited to a primary antibody which cannot distinguish between the GSH and GSSG. The use of redox-sensitive mbrB or mchlB and visualization of reduced glutathione via bright-field, fluorescence microscopy or CLSM, by contrast, allows insights into the in vivo situation within the nucleus and cytosol. Other plant cell compartments and the glutathione distribution in the mesophyll or vascular tissue are not accessible due to the limited infiltration potential of redox-sensitive dyes and/or a microscope resolution of approx. 200 nm. Sample preparation and microscope properties are potential stress sources and can thus influence glutathione redox state and distribution within the compartments. Cell compartment-specific, roGFPs, visualized via a fluorescence microscope or CLSM, indicate the in vivo redox state of glutathione in mitochondria, chloroplasts, the cytosol, peroxisomes (primarily GSH) and the endoplasmic reticulum (ER; primarily GSSG). Again, examinations are limited to thin organs/tissues and outer cell layers, and sample preparation or microscope properties are potential stress sources. Probes are mainly available for Arabidopsis and require genetical modifications of the target plant.
Alternatively, glutathione can be quantified after reduction of GSSG in the sample with dithiotreitol (DTT) and subsequent derivatization of GSH with mbrB or mchlB via a reverse-phase high-performance liquid chromatography (HPLC) system equipped with a fluorescent or UV detector, or spectrophotometrically after a GR-mediated glutathione recycling assay (total glutathione) from cell organelles after isolation or fractionation. To assess the GSSG fraction, first, thiol groups of GSH are blocked with N-ethylmaleimide (NEM) in a subsample, followed by removal of excess NEM with toluene prior to reduction of GSSG to GSH with DTT and derivatization via mbrB (Kranner 1998; Kranner and Grill 1993; Roach et al. 2018) or mchlB (Hajdinák et al. 2019). Similarly, NEM or vinylpyridine is added to the glutathione recycling assay reaction mix, and the GSH content can be calculated by the subtraction of GSSG from total glutathione. Hajdinák et al. (2019) found that the use of mchlB and HPLC analysis yielded more exact results compared to the glutathione recycling assay when applied to Arabidopsis suspension cell cultures, and mitochondrial, microsomal (ER) and cytosol fractions. HPLC with UV detection facilitates the quantification of GSH and GSSG without the need of derivatization. Such an approach has recently been applied to leucoplasts and vacuoles isolated from storage parenchyma cells of red beet (Beta vulgaris L.) (Pradedova et al. 2019). Generally, great care has to be taken during sample preparation to avoid auto-oxidation of GSH, washing out of and/or redistributing of glutathione between cell compartments. Such cross-contamination or additional stress to the sample may occur by applying too much pressure during tissue infiltration or due to microscope properties (strong light source, high temperature, lack of oxygen, water stress), but likewise during organelle isolation or fractionation (Zechmann 2014). According to Hajdinák et al. (2019), the addition of 1 mM mchlB to organelle isolation buffer is recommended to prevent auto-oxidation of GSH during the fractionation process.
Glutathione concentrations in individual cell compartments, calculated from a combination of data on the volume of cell compartments, with total glutathione immunolabeling density and glutathione concentrations in whole Arabidopsis leaf tissue (Queval et al. 2011; Han et al. 2013; Koffler et al. 2013; Zechmann 2020; Dorion et al. 2021) are in the millimolar range (mitochondria [15 mM], > nuclei [6.4 mM], > cytosol [4.5 mM], > peroxisomes [4.4 mM], > chloroplasts [1.2 mM], > vacuole [0.08 mM]). Similar glutathione concentrations have been found in earlier studies in animal tissue or Arabidopsis for mitochondria (Wahlländer et al. 1979; García-Ruiz et al. 1994; Meyer et al. 2001; Krueger et al. 2009), whereas the glutathione concentration was higher in animal cytosol (Wahlländer et al. 1979; García-Ruiz et al. 1994), and calculated cytosol levels were lower in young wheat leaves (Noctor et al. 2002). The reported chloroplast and vacuole glutathione concentration differs markedly among plant species (0.5–5 mM and 0.08–0.7 mM, respectively), which might be due to redistribution of glutathione via compartment-specific glutathione transporters (identified transporters summarized by Dorion et al. 2021) and/or sample handling during organelle isolation (discussed in detail by Noctor et al. 2002; Krueger et al. 2009; Hajdinák et al. 2019). Traces of 0.003 mM glutathione were detected in barley apoplast extract (Vanacker et al. 1998). Although this very low glutathione concentration in the apoplast may not be sufficient to act as antioxidant, such as during biotic stress (fungal infection), glutathione and/or its redox state may alternatively have a defense-signaling function in this cell compartment (Zechmann 2020). Given that the total glutathione concentration of different C3-type plants is highest in mitochondria (Zechmann et al. 2008a; Zechmann and Müller 2010; Heyneke et al. 2013; Koffler et al. 2013; Müller et al. 2014; Vidović et al. 2016; Gasperl et al. 2021) and increases with leaf age in Arabidopsis (Koffler et al. 2013), which is probably induced by higher ROS accumulation in older leaves, this high and comparatively stable glutathione level seems to be crucial for mitochondria and cell functioning and viability under non-stress and stress conditions (Zechmann 2017, 2020). Although it seems surprising at first that chloroplasts of non-stressed plants have comparatively low glutathione levels, their ability to synthesize glutathione rapidly upon high light stress, such as within 90 s when Arabidopsis plants were transferred from 50 to 1000 µmol m−2 s−1 (Choudhury et al. 2018), seems to be sufficient for redox-homeostasis in this compartment and even to supply glutathione for redox-balancing of the cytosol (Maughan et al. 2010).
A high glutathione concentration in the nucleus protects nucleic acids, proteins and lipids from oxidation. Nuclear glutathione further plays a role in redox adjustments during the cell cycle (Diaz Vivancos et al. 2010b, 2010a). Redox-homeostasis in the nucleus is maintained by GSH and GSSG exchange with the cytosol via the nuclear pores and by nuclear GR activity (Dorion et al. 2021; Müller-Schüssele et al. 2021). The increased GSH demand of the nucleus, triggered by either salicylic acid or excess light treatment, upregulated glutathione synthesis at the transcriptional level in wheat and Arabidopsis (Diaz Vivancos et al. 2010a; Gasperl et al. 2021). For redox adjustments to excess light, a high nuclear glutathione demand seems to be important, whereas a relocation of glutathione to peroxisomes seems to facilitate adaptations to far-red light under Asc deficiency (Heyneke et al. 2013; Gasperl et al. 2021). Peroxisomes require glutathione mainly to counteract H2O2 from SOD- or glycolate oxidase-associated reactions (Noctor et al. 2018). As mentioned above, glutathione in peroxisomes of stressed plants scavenges excess H2O2 from photorespiration under excess light or osmotic stress (Zechmann 2014; Gasperl et al. 2021).
Studies using Arabidopsis roGFP reporter lines that targeted specific cell compartments showed that in most of these compartments, glutathione is maintained in a reduced redox state under non-stress conditions. Exceptions are the ER lumen, the vacuole and the apoplast, where GR, to regenerate GSH from GSSG, is absent (Noctor et al. 2012; Müller-Schüssele et al. 2021). A more positive reduction potential (Eh) and evidence for the presence of GT activity, which may use GSSG in the vacuole to reduce organic hydroperoxides (Öztetik 2008), was recently detected in vacuoles of dormant red beet taproot cells (Pradedova et al. 2019). The distinct glutathione (and Asc) level and its redox state in individual cell compartments allow control of the local subcellular redox environment and gradient, which affects many redox-dependent metabolic processes and is therefore considered a valuable marker for the plant stress response (Foyer and Noctor 2009; Noctor et al. 2012, 2013; Müller et al. 2014; Zechmann 2020; Dorion et al. 2021).
Modifications in compartment-specific glutathione distribution and redox-state by light intensity and spectrum composition
Subcellular total glutathione levels changed with the diurnal rhythm in Arabidopsis (except for vacuoles), reaching a maximum after 3 h of light (150 µmol m−2 s−1), followed by a strong decrease within the next 1–2 h and a minimum at the end of the night. A much lower glutathione concentration seems to result from limited availability of the glutathione precursors glycine (from reduced photorespiration) and cysteine (from reduced sulfur uptake and incorporation) during darkness (Buwalda et al. 1990; Noctor et al. 1997, 1999; Huseby et al. 2013). The depleted glutathione level was restored when plants were exposed to light (150 µmol m−2 s−1) and fed with glycine or cysteine, respectively (Zechmann et al. 2007, 2008b; Höller et al. 2010; Király et al. 2012). Accordingly, excess light induced cysteine supply, glutathione synthesis (chloroplastic and more prominently cytosolic GSHS) and reduction (cytosolic GR) in wheat (Gasperl et al. 2021). A gradual increase in the chl-roGFP oxidation state (chl-EGSH) was recently reported in a chloroplast-targeted redox-sensitive (chl-roGFP2) potato reporter line to positively correlate with gradually increasing light intensity from 200 up to 720 and 1250 µmol m−2 s−1, respectively. The oxidizing effect was reversible by returning to a light intensity of 200 µmol m−2 s−1 after 14 h. A particularly strong effect on the potato chl-EGSH in older leaves was found when the higher light intensities were combined with cold treatment (3 °C). Increased chl-EGSH did not recover when the light intensity was lowered to 200 µmol m−2 s−1 after 10 h. The effect was, however, dependent on the developmental stage of the leaves, as newly formed upper leaves showed no increase in chl-roGFP oxidation state. This discrepancy may be explained by the comparatively low photosynthetic capacity of young leaves and by the induction of photo-protective mechanisms (non-photochemical quenching, photorespiration) (Hipsch et al. 2021). In accordance with the previous findings, organelle-specific responses to changes in light intensity were detected by redox-sensitive Arabidopsis reporter lines (roGFP2). The redox state shifted towards oxidation in peroxisomes, organelles associated with the detoxification of ROS and H2O2 from photorespiration, during 22 h of darkness, whereas in chloroplasts, which are organelles associated with glutathione synthesis, the redox state was specifically affected by short-term excess light (3 h at 600 µmol m−2 s−1) (Bratt et al. 2016). Haber and Rosenwasser (2020) reported immediate oxidation of Arabidopsis chl-roGFP after transition from darkness to light, which was more severe at light intensities of 750–1700 µmol m−2 s−1 than at 220–650 µmol m−2 s−1, and followed by gradual reduction of the chl-roGFP, which impressively demonstrates the versatility of the plant subcellular redox environment in response to changes in light.
Low light intensity (50 µmol m−2 s−1) for a few hours (4 h) did not alter the total glutathione level in the organelles of Arabidopsis wildtype plants. However, it was lower compared than that at the control light intensity (150 µmol m−2 s−1) in peroxisomes of the Asc-deficient Arabidopsis vtc2-1 mutant (Heyneke et al. 2013). Similarly, in peroxisomes, nuclei and the cytosol of wheat cultivated at the same low light intensity for several days, decreased total glutathione levels were detected compared to plants grown at 250 µmol m−2 s−1 (Gasperl et al. 2021). Arabidopsis vtc2-1 accumulates only 10 to 30% of the wildtype Asc level, which results from a mutation in the GDP-l-galactose phosphorylase 1 gene, the enzyme catalyzing an intermediate step in Asc synthesis (Müller-Moulé et al. 2004; Linster et al. 2007). Excess light (300, 700, 1500 µmol m−2 s−1) for a few hours stimulated total glutathione accumulation in the Arabidopsis wildtype, particularly in organelles associated with glutathione synthesis (chloroplasts and the cytosol) and detoxification of ROS and H2O2 from photorespiration (peroxisomes), whereas in the glutathione-deficient Arabidopsis pad2-1 mutant, total glutathione increased in mitochondria, where ROS are frequently formed during respiration (Heyneke et al. 2013). Arabidopsis pad2-1 accumulates only approximately 20% of the wildtype glutathione level (except for mitochondria), which results from a mutation in the γ-ECS (GSH1) gene, the enzyme catalyzing the rate-limiting step in glutathione synthesis (Parisy et al. 2007; Zechmann and Müller 2010; Koffler et al. 2011). Arabidopsis wildtype, the pad2-1 and vtc2-1 mutants and the moderately frost-sensitive wheat variety Chinese Spring adapted to excess light at 500 µmol m−2 s−1 (control light intensity: 250 µmol m−2 s−1) for several days by increasing glutathione concentrations. At the subcellular level, excess light stimulated total glutathione accumulation, particularly in chloroplasts, the cytosol and peroxisomes, in the Arabidopsis pad2-1 mutant and wheat (Gasperl et al. 2021).
Long-term excess light at 300 and 700 µmol m−2 s−1 for 2 weeks enhanced total glutathione (and Asc) accumulation in the stroma of Arabidopsis wildtype chloroplasts. Only plants grown at 1500 µmol m−2 s−1 accumulated glutathione (and Asc) in the stroma and inside the thylakoid lumen. Arabidopsis wildtype chloroplasts were able to counteract oxidative stress from 14 days of exposure to excess light at up to 1500 µmol m−2 s−1 in the mesophyll of the leaf center, as H2O2 concentration did not alter in this area (in contrast to short-term excess light; see above), and total glutathione (and Asc) level increased markedly; this local increase in antioxidants was accompanied by structural and ultrastructural adaptations, namely reduction of chloroplast number and thylakoids and accumulation of plastoglobuli, respectively (Heyneke et al. 2013). Plastoglobuli are in close contact with thylakoid membranes and serve as biosynthesis and storage subcompartments, for example for carotenoids and xanthophylls (Austin et al. 2006). Plastoglobuli accumulate during thylakoid membrane re-organization and degradation in response to stress and thus play an important role in the adjustment of chloroplasts to stress (Espinoza-Corral et al. 2021; Zechmann et al. 2021). Excess light for 5 days at 500 µmol m−2 s−1 (control light intensity: 120 µmol m−2 s−1) likewise led to plastoglobuli accumulation and thylakoid membrane degradation/re-organization and increased carotenoid content in plastoglobuli in Arabidopsis wildtype plants and induced proteins associated with leaf senescence and jasmonic acid biosynthesis (Espinoza-Corral et al. 2021). Specifically, additional adjustments to 14 days of exposure to excess light at 1500 µmol m−2 s−1 were found in the glutathione (pad2-1)- and Asc (vtc2-1)-deficient mutants of Arabidopsis. In comparison to the wildtype and vtc2-1 mutant, the pad2-1 mutant seems to compensate its glutathione deficiency by maintaining comparatively high levels of Asc in peroxisomes and by forming an additional layer of palisade cells (Heyneke et al. 2013). Thickened leaves as a response to high light intensity are also known from other plant species (Björkman 1981; Pearcy 1998). Total glutathione increased in most compartments of the vtc2-1 mutant, but particularly in the chloroplasts and nuclei (Heyneke et al. 2013). The depletion of cytosolic glutathione by import into the nucleus induces glutathione synthesis, which accelerates the accumulation of GSH and allows the maintenance of a reducing redox environment under stress conditions (Diaz Vivancos et al. 2010a). It would appear that the vtc2-1 mutant seems to compensate its lack of Asc by a combination of glutathione re-location and synthesis. At the whole leaf level o thef Arabidopsis wildtype, increase in photo-protective anthocyanins was visible in plants grown at 300 µmol m−2 s−1, which was more pronounced, but also accompanied by small necrotic lesions at leaf edges at 700 µmol m−2 s−1, and intensified even more at 1500 µmol m−2 s−1, while the photosynthesis-associated pigments (chlorophyll and carotenoids) were reduced (Heyneke et al. 2013).
White light with a lower red/far-red ratio induces cysteine and glutathione metabolism in wheat, but not at the transcriptional level (Monostori et al. 2018; Toldi et al. 2019; Gasperl et al. 2021). Lower red/far-red ratio (blue/red 1:5; red/far-red: 10:1) at 250 µmol m−2 s−1 for several days raised GSH and GSSG concentrations compared to white light (blue/red 1:2; red/far-red: 15:1) of the same intensity; GSH increased, however, to a greater extent. Thus, the GSSG/GSH ratio was at the same level (~ 0.1) as in wheat leaves grown under white light of the same intensity. Leaves of the Arabidopsis wildtype and vtc2-1 and pad2-1 mutations maintained largely the same GSH and GSSG concentrations and GSSG/GSH ratio (0.1) when grown under lower red/far-red ratio or white light spectrum, respectively. Total glutathione labeling density in the Arabidopsis wildtype and the vtc2-1 mutant, except for peroxisomes (1.8-fold increase) in the vtc2-1 mutant, was likewise largely unaffected by the decreased red/far-red ratio. In the Arabidopsis pad2-1 mutant, total glutathione labeling density increased in nuclei by twofold, and in peroxisomes and the cytosol by onefold; in wheat, it decreased in cytosol by 1.8-fold, in chloroplasts by 1.5-fold and in nuclei and peroxisomes, by onefold respectively, in comparison to white light at the same intensity. Similar to excess light response, cysteine, GSSG and GSH in wheat leaf extracts increased under the lower red/far-red ratio. However, in contrast to excess light, GSH supply either from de novo synthesis and/or reduction was not transcriptionally upregulated (Gasperl et al. 2021). For this experimental setting and from work involving even higher light intensities, we can therefore conclude that Arabidopsis and wheat plants are able to adjust their subcellular redox environment to different light intensities (Heyneke et al. 2013; Gasperl et al. 2021) and to a lower red/far-red ratio, and that the adaptations in subcellular glutathione distribution are similar under excess light and a lower red/far-red ratio in the Arabidopsis pad2-1 mutant and wheat (Gasperl et al. 2021).
Conclusion
The increase in ROS and H2O2 along the photosynthetic electron transport chain in chloroplast thylakoids and the subsequent transfer of excess ROS and H2O2 from photorespiration in peroxisomes into the cytosol and vacuoles under excess light initiates the accumulation and redistribution of antioxidants (glutathione, ascorbate) in plant cell compartments (Fig. 1). Under a lower red/far-red ratio, we propose that an increase in ROS and H2O2 in chloroplasts arises from inefficient chlorophyll biosynthesis in seedlings after transition to white light or in mature shaded leaves from an imbalance in the excitation state of the two photosystems (short term) and limited availability of reductants and assimilates (long term), which again initiates the accumulation and redistribution of glutathione (and ascorbate) in plant cell compartments (Fig. 1). Choosing the most suitable method for glutathione localization and quantification studies in subcellular compartments of plants depends on the cell layers (epidermis or mesophyll, vascular tissue), cell compartments and form of glutathione (total glutathione or redox state of glutathione) that studies target (Table 1). Limiting the auto-oxidation and/or redistribution of glutathione during analyses is generally crucial for obtaining reliable results. Light intensity-mediated redox shifts in plant cells are compartment specific, time dependent and reversible under non-stress conditions. Light intensity signaling by phyA and possibly by ROS into the nucleus induces glutathione synthesis and reduction at the transcriptional level in wheat. Differences between Arabidopsis and wheat in response to moderate excess light regimes at the subcellular level may arise from the different light intensity optima (Arabidopsis: 100–150 µmol m−2 s−1, wheat: 250 µmol m−2 s−1) of both plants, and Arabidopsis may require thus faster signaling and adaptation. Differences between Arabidopsis and wheat in terms of their response to a lower red/far-red ratio at the subcellular level may arise, because in Arabidopsis, leaves of lower nodes are partially shaded by younger leaves of the same plant, while wheat is grown in dense rows and leaves of lower nodes are shaded by younger leaves of the same plant and by neighboring plants.
Improved drought tolerance in a wheat variety and in soybean after excess light or shade pre-treatment is at least partly linked to antioxidants and associated enzyme activities. Moderate excess light or far-red light may in the future allow the priming of crop seedlings against certain stress conditions prior to transplanting them into the field.
Abbreviations
- 1O2 :
-
Singlet oxygen
- ADP:
-
Adenosine diphosphate
- AOX:
-
Alternative oxidase
- APX:
-
Ascorbate peroxidase
- Asc:
-
Ascorbate
- ATP:
-
Adenosine triphosphate
- cat :
-
Catalase knockout
- CAT:
-
Catalase
- chl-E GSH :
-
Chl-roGFP oxidation state
- chl-roGFP2:
-
Chloroplast targeted roGFP
- CLSM:
-
Confocal-laser scanning microscopy
- cry:
-
Cryptochrome
- DTT:
-
Dithiotreitol
- Eh :
-
Reduction potential
- ER:
-
Endoplasmic reticulum
- FIN219:
-
Far-Red Insensitive 219
- FIP1:
-
FIN219-interacting protein 1
- GGT-1:
-
Gamma-glutamyl transferase/transpeptidase 1
- ggt :
-
GGT-1 knockout
- GR:
-
Glutathione reductase
- GSH:
-
Reduced glutathione
- GSHS:
-
Glutathione synthetase
- GSSG:
-
Oxidized glutathione (= glutathione disulfide)
- GSSG/GSH:
-
Reduction state of glutathione
- GT:
-
Glutathione transferase
- H2O2 :
-
Hydrogen peroxide
- HPLC:
-
High performance liquid chromatography
- HyPer2:
-
Fluorescent H2O2 biosensor
- IgG:
-
Immunoglobulin G
- immunolabeling:
-
Total glutathione localization by use of a specific antibody
- LEDs:
-
Light-emitting diodes
- mbrB:
-
Monobromobimane
- mchlB:
-
Monochlorobimane
- NADP+ :
-
Nicotinamide adenine dinucleotide phosphate
- NADPH:
-
Reduced form of NADP+
- NEM:
-
N-ethylmaleimide
- O2 − :
-
Superoxide radicals
- pad :
-
Phytoalexin deficient
- phyA:
-
Phytochrome A
- PS II:
-
Photosystem II
- roGFP:
-
Redox-sensitive green fluorescence protein
- ROS:
-
Reactive oxygen species
- SOD:
-
Superoxide dismutase
- TEM:
-
Transmission electron microscope
- Total glutathione:
-
Sum of reduced (GSH) and oxidized (GSSG) glutathione, detected via immuno-labeling
- vtc :
-
Vitamin C1 gene
- γ-EC:
-
γ-Glutamylcysteine
- γ-ECS:
-
γ-Glutamylcysteine synthetase
References
Appolloni E, Pennisi G, Zauli I, Carotti L, Paucek I, Quaini S, Orsini F, Gianquinto G (2022) Beyond vegetables: effects of indoor LED light on specialized metabolite biosynthesis in medicinal and aromatic plants, edible flowers, and microgreens. J Sci Food Agric 102(2):472–487. https://doi.org/10.1002/jsfa.11513
Asada K (1999) The water-water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Annu Rev Plant Physiol Plant Mol Biol 50:601–639. https://doi.org/10.1146/annurev.arplant.50.1.601
Asghar MA, Du J, Jiang H, Li Y, Sun X, Shang J, Liu J, Liu W, Imran S, Iqbal N, Ahmad B, Hussain S, Yu L, Liu C, Yang W (2020) Shade pretreatment enhanced drought resistance of soybean. Environ Exp Bot 171:103952. https://doi.org/10.1016/j.envexpbot.2019.103952
Austin JR, Frost E, Vidi P-A, Kessler F, Staehelin LA (2006) Plastoglobules are lipoprotein subcompartments of the chloroplast that are permanently coupled to thylakoid membranes and contain biosynthetic enzymes. Plant Cell 18(7):1693–1703. https://doi.org/10.1105/tpc.105.039859
Bartoli CG, Tambussi EA, Diego F, Foyer CH (2009) Control of ascorbic acid synthesis and accumulation and glutathione by the incident light red/far red ratio in Phaseolus vulgaris leaves. FEBS Lett 583(1):188–122
Björkman O (1981) Responses to different quantum flux densities. In: Lange OL, Nobel PS, Osmond CB et al (eds) Physiological plant ecology I. Encyclopedia of plant physiology, vol 12. Springer Heidelberg, Berlin, pp 57–107
Bouly J-P, Schleicher E, Dionisio-Sese M, Vandenbussche F, van der Straeten D, Bakrim N, Meier S, Batschauer A, Galland P, Bittl R, Ahmad M (2007) Cryptochrome blue light photoreceptors are activated through interconversion of flavin redox states. J Biol Chem 282(13):9383–9391. https://doi.org/10.1074/jbc.M609842200
Bratt A, Rosenwasser S, Meyer A, Fluhr R (2016) Organelle redox autonomy during environmental stress. Plant Cell Environ 39(9):1909–1919. https://doi.org/10.1111/pce.12746
Buwalda F, Stulen I, Kok LJ, Kuiper PJC (1990) Cysteine, gamma-glutamyl-cysteine and glutathione contents of spinach leaves as affected by darkness and application of excess sulfur. II. Glutathione accumulation in detached leaves exposed to H2S in the absence of light is stimulated by the supply of glycine to the petiole. Physiol Plant 80(2):196–204. https://doi.org/10.1111/j.1399-3054.1990.tb04396.x
Chen M, Chory J, Fankhauser C (2004) Light signal transduction in higher plants. Annu Rev Genet 38:87–117. https://doi.org/10.1146/annurev.genet.38.072902.092259
Chen I-C, Huang I-C, Liu M-J, Wang Z-G, Chung S-S, Hsieh H-L (2007) Glutathione S-transferase interacting with far-red insensitive 219 is involved in phytochrome A-mediated signaling in Arabidopsis. Plant Physiol 143(3):1189–1202. https://doi.org/10.1104/pp.106.094185
Choudhury FK, Devireddy AR, Azad RK, Shulaev V, Mittler R (2018) Rapid accumulation of glutathione during light stress in Arabidopsis. Plant Cell Physiol 59(9):1817–1826. https://doi.org/10.1093/pcp/pcy101
Csiszár J, Horváth E, Bela K, Gallé Á (2016) Glutathione-related enzyme system: Glutathione reductase (GR), glutathione transferases (GSTs) and glutathione reroxidases (GPXs). In: Gupta DK, Palma JM, Corpas FJ (eds) Redox state as a central regulator of plant-cell stress responses. Springer International Publishing, Cham, pp 137–158
Diaz Vivancos P, Wolff T, Markovic J, Pallardó FV, Foyer CH (2010a) A nuclear glutathione cycle within the cell cycle. Biochem J 431(2):169–178. https://doi.org/10.1042/BJ20100409
Diaz Vivancos P, Dong Y, Ziegler K, Markovic J, Pallardó FV, Pellny TK, Verrier PJ, Foyer CH (2010b) Recruitment of glutathione into the nucleus during cell proliferation adjusts whole-cell redox homeostasis in Arabidopsis thaliana and lowers the oxidative defence shield. Plant J 64(5):825–838. https://doi.org/10.1111/j.1365-313X.2010.04371.x
Dietzel L, Bräutigam K, Pfannschmidt T (2008) Photosynthetic acclimation: state transitions and adjustment of photosystem stoichiometry—functional relationships between short-term and long-term light quality acclimation in plants. FEBS J 275(6):1080–1088. https://doi.org/10.1111/j.1742-4658.2008.06264.x
Ding S, Jiang R, Lu Q, Wen X, Lu C (2016) Glutathione reductase 2 maintains the function of photosystem II in Arabidopsis under excess light. Biochim Biophys Acta Bioenerg 6:665–677. https://doi.org/10.1016/j.bbabio.2016.02.011
Dorion S, Ouellet JC, Rivoal J (2021) Glutathione metabolism in plants under stress: beyond reactive oxygen species detoxification. Metabolites 11(9):641. https://doi.org/10.3390/metabo11090641
Espinoza-Corral R, Schwenkert S, Lundquist PK (2021) Molecular changes of Arabidopsis thaliana plastoglobules facilitate thylakoid membrane remodeling under high light stress. Plant J 106(6):1571–1587. https://doi.org/10.1111/tpj.15253
Exposito-Rodriguez M, Laissue PP, Yvon-Durocher G, Smirnoff N, Mullineaux PM (2017) Photosynthesis-dependent H2O2 transfer from chloroplasts to nuclei provides a high-light signalling mechanism. Nat Commun 8(1):49. https://doi.org/10.1038/s41467-017-00074-w
Foyer CH, Halliwell B (1976) The presence of glutathione and glutathione reductase in chloroplasts: a proposed role in ascorbic acid metabolism. Planta 133(1):21–25. https://doi.org/10.1007/BF00386001
Foyer CH, Noctor G (2005) Redox homeostasis and antioxidant signaling: a metabolic interface between stress perception and physiological responses. Plant Cell 17(7):1866–1875. https://doi.org/10.1105/tpc.105.033589
Foyer CH, Noctor G (2009) Redox regulation in photosynthetic organisms: signaling, acclimation, and practical implications. Antioxid Redox Signal 11(4):861–905. https://doi.org/10.1089/ars.2008.2177
Foyer CH, Noctor G (2011) Ascorbate and glutathione: the heart of the redox hub. Plant Physiol 155(1):2–18. https://doi.org/10.1104/pp.110.167569
Gallé Á, Czékus Z, Bela K, Horváth E, Ördög A, Csiszár J, Poór P (2018) Plant glutathione transferases and light. Front Plant Sci 9:1944. https://doi.org/10.3389/fpls.2018.01944
Ganguly DR, Stone BAB, Bowerman AF, Eichten SR, Pogson BJ (2019) Excess light priming in Arabidopsis thaliana genotypes with altered DNA methylomes. G3 (Bethesda) 9(11):3611–3621
García-Ruiz C, Morales A, Ballesta A, Rodés J, Kaplowitz N, Fernández-Checa JC (1994) Effect of chronic ethanol feeding on glutathione and functional integrity of mitochondria in periportal and perivenous rat hepatocytes. J Clin Investig 94(1):193–201. https://doi.org/10.1172/JCI117306
Gasperl A, Balogh E, Boldizsár Á, Kemeter N, Pirklbauer R, Möstl S, Kalapos B, Szalai G, Müller M, Zellnig G, Kocsy G (2021) Comparison of light condition-dependent differences in the accumulation and subcellular localization of glutathione in Arabidopsis and wheat. IJMS 22(2):607. https://doi.org/10.3390/ijms22020607
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48(12):909–930. https://doi.org/10.1016/j.plaphy.2010.08.016
Gyugos M, Ahres M, Gulyás Z, Szalai G, Darkó É, Végh B, Boldizsár Á, Mednyánszky Z, Kar RK, Dey N, Simon-Sarkadi L, Galiba G, Kocsy G (2019) Role of light-intensity-dependent changes in thiol and amino acid metabolism in the adaptation of wheat to drought. J Agro Crop Sci 16(247):562–570. https://doi.org/10.1111/jac.12358
Haber Z, Rosenwasser S (2020) Resolving the dynamics of photosynthetically produced ROS by high-resolution monitoring of chloroplastic EGSH in Arabidopsis. BioRxiv. https://doi.org/10.1101/2020.03.04.976092
Hajdinák P, Czobor Á, Lőrincz T, Szarka A (2019) The problem of glutathione determination: a comparative study on the measurement of glutathione from plant cells. Period Polytech Chem Eng 63(1):1–10. https://doi.org/10.3311/PPch.11785
Han Y, Chaouch S, Mhamdi A, Queval G, Zechmann B, Noctor G (2013) Functional analysis of Arabidopsis mutants points to novel roles for glutathione in coupling H2O2 to activation of salicylic acid accumulation and signaling. Antioxid Redox Signal 18(16):2106–2121. https://doi.org/10.1089/ars.2012.5052
Heyneke E, Luschin-Ebengreuth N, Krajcer I, Wolkinger V, Müller M, Zechmann B (2013) Dynamic compartment specific changes in glutathione and ascorbate levels in Arabidopsis plants exposed to different light intensities. BMC Plant Biol 13:104. https://doi.org/10.1186/1471-2229-13-104
Hipsch M, Lampl N, Zelinger E, Barda O, Waiger D, Rosenwasser S (2021) Sensing stress responses in potato with whole-plant redox imaging. Plant Physiol 187(2):618–631. https://doi.org/10.1093/plphys/kiab159
Höller K, Király L, Künstler A, Müller M, Gullner G, Fattinger M, Zechmann B (2010) Enhanced glutathione metabolism is correlated with sulfur-induced resistance in Tobacco mosaic virus-infected genetically susceptible Nicotiana tabacum plants. Mol Plant Microbe Interact 23(11):1448–1459. https://doi.org/10.1094/MPMI-05-10-0117
Huang S, van Aken O, Schwarzländer M, Belt K, Millar AH (2016) The roles of mitochondrial reactive oxygen species in cellular signaling and stress response in plants. Plant Physiol 171(3):1551–1559. https://doi.org/10.1104/pp.16.00166
Huseby S, Koprivova A, Lee B-R, Saha S, Mithen R, Wold A-B, Bengtsson GB, Kopriva S (2013) Diurnal and light regulation of sulphur assimilation and glucosinolate biosynthesis in Arabidopsis. J Exp Bot 64(4):1039–1048. https://doi.org/10.1093/jxb/ers378
Jenkins MW (2021) Cannabis sativa L. response to narrow bandwidth uv and the combination of blue and red light during the final stages of flowering on leaf level gas-exchange parameters, secondary metabolite production, and yield. Agric Sci 12(12):1414–1432. https://doi.org/10.4236/as.2021.1212090
Jiang H-W, Liu M-J, Chen I-C, Huang C-H, Chao L-Y, Hsieh H-L (2010) A glutathione S-transferase regulated by light and hormones participates in the modulation of Arabidopsis seedling development. Plant Physiol 154(4):1646–1658. https://doi.org/10.1104/pp.110.159152
Kami C, Lorrain S, Hornitschek P, Fankhauser C (2010) Light-regulated plant growth and development. In: Timmermans MCP (ed) Plant development, vol 91. Elsevier Academic, Amsterdam, pp 29–66
Karuppanapandian T, Moon J-C, Kim C, Manoharan K, Kim W (2011) Reactive oxygen species in plants: their generation, signal transduction, and scavenging mechanisms. Aust J Crop Sci 5(6):709–725
Kianianmomeni A (2014) More light behind gene expression. Trends Plant Sci 19(8):488–490. https://doi.org/10.1016/j.tplants.2014.05.004
Kim C, Apel K (2013) Singlet oxygen-mediated signaling in plants: moving from flu to wild type reveals an increasing complexity. Photosynth Res 116(2–3):455–464. https://doi.org/10.1007/s11120-013-9876-4
Király L, Künstler A, Höller K, Fattinger M, Juhász C, Müller M, Gullner G, Zechmann B (2012) Sulfate supply influences compartment specific glutathione metabolism and confers enhanced resistance to Tobacco mosaic virus during a hypersensitive response. Plant Physiol Biochem 59:44–54. https://doi.org/10.1016/j.plaphy.2011.10.020
Kocsy G, Tari I, Vanková R, Zechmann B, Gulyás Z, Poór P, Galiba G (2013) Redox control of plant growth and development. Plant Sci 211:77–91. https://doi.org/10.1016/j.plantsci.2013.07.004
Koffler BE, Maier R, Zechmann B (2011) Subcellular distribution of glutathione precursors in Arabidopsis thaliana. J Integr Plant Biol 53(12):930–941. https://doi.org/10.1111/j.1744-7909.2011.01085.x
Koffler BE, Bloem E, Zellnig G, Zechmann B (2013) High resolution imaging of subcellular glutathione concentrations by quantitative immunoelectron microscopy in different leaf areas of Arabidopsis. Micron 45:119–128. https://doi.org/10.1016/j.micron.2012.11.006
Kong Y, Zheng Y (2020) Phototropin is partly involved in blue-light-mediated stem elongation, flower initiation, and leaf expansion: A comparison of phenotypic responses between wild Arabidopsis and its phototropin mutants. Environ Exp Bot 171:103967. https://doi.org/10.1016/j.envexpbot.2019.103967
Kranner I (1998) Determination of glutathione, glutathione disulphine and two related enzymes, glutathione reductase and glucose-6-phoshate dehydrogenase, in fungal and plant cells. In: Varma A (ed) Mycorrhiza manual. Springer, Heidelberg, pp 227–241
Kranner I, Grill D (1993) Content of low-molecular-weight thiols during the imbibition of pea seeds. Physiol Plant 88(4):557–562. https://doi.org/10.1111/j.1399-3054.1993.tb01371.x
Krueger S, Niel A, Lopez Martin MC, Steinhauser D, Donath A, Hildebrandt T, Romero LC, Hoefgen R, Gotor C, Hesse H (2009) Analysis of cytosolic and plastidic serine acetyltransferase mutants and subcellular metabolite distributions suggests interplay of the cellular compartments for cysteine biosynthesis in Arabidopsis. Plant Cell Environ 32(4):349–367. https://doi.org/10.1111/j.1365-3040.2009.01928.x
Labrou NE, Papageorgiou AC, Pavli O, Flemetakis E (2015) Plant GSTome: structure and functional role in xenome network and plant stress response. Curr Opin Biotechnol 32:186–194. https://doi.org/10.1016/j.copbio.2014.12.024
Li C-X, Xu Z-G, Dong R-Q, Chang S-X, Wang L-Z, Khalil-Ur-Rehman M, Tao J-M (2017) An RNA-Seq analysis of grape plantlets grown in vitro reveals different responses to blue, green, red LED light, and white fluorescent light. Front Plant Sci 8:78. https://doi.org/10.3389/fpls.2017.00078
Linster CL, Gomez TA, Christensen KC, Adler LN, Young BD, Brenner C, Clarke SG (2007) Arabidopsis VTC2 encodes a GDP-L-galactose phosphorylase, the last unknown enzyme in the Smirnoff-Wheeler pathway to ascorbic acid in plants. J Biol Chem 282(26):18879–18885. https://doi.org/10.1074/jbc.M702094200
Liu T, Zhang X (2021) Comparative transcriptome and metabolome analysis reveal glutathione metabolic network and functional genes underlying blue and red-light mediation in maize seedling leaf. BMC Plant Biol 21(1):593. https://doi.org/10.1186/s12870-021-03376-w
Marty L, Bausewein D, Müller C, Bangash SAK, Moseler A, Schwarzländer M, Müller-Schüssele SJ, Zechmann B, Riondet C, Balk J, Wirtz M, Hell R, Reichheld J-P, Meyer AJ (2019) Arabidopsis glutathione reductase 2 is indispensable in plastids, while mitochondrial glutathione is safeguarded by additional reduction and transport systems. New Phytol 224(4):1569–1584. https://doi.org/10.1111/nph.16086
Maughan SC, Pasternak M, Cairns N, Kiddle G, Brach T, Jarvis R, Haas F, Nieuwland J, Lim B, Müller C, Salcedo-Sora E, Kruse C, Orsel M, Hell R, Miller AJ, Bray P, Foyer CH, Murray JAH, Meyer AJ, Cobbett CS (2010) Plant homologs of the Plasmodium falciparum chloroquine-resistance transporter, PfCRT, are required for glutathione homeostasis and stress responses. Proc Natl Acad Sci USA 107(5):2331–2336. https://doi.org/10.1073/pnas.0913689107
Meyer AJ, May MJ, Fricker M (2001) Quantitative in vivo measurement of glutathione in Arabidopsis cells. Plant J 27(1):67–78. https://doi.org/10.1046/j.1365-313x.2001.01071.x
Monostori I, Heilmann M, Kocsy G, Rakszegi M, Ahres M, Altenbach SB, Szalai G, Pál M, Toldi D, Simon-Sarkadi L, Harnos N, Galiba G, Darko É (2018) LED lighting - modification of growth, metabolism, yield and flour composition in wheat by spectral quality and intensity. Front Plant Sci 9:605. https://doi.org/10.3389/fpls.2018.00605
Müller M, Koffler BE, Zechmann B (2014) High resolution imaging of subcellular ascorbate and glutathione contents in plants. Phyton (Horn) 54(2):353–368
Müller-Moulé P, Golan T, Niyogi KK (2004) Ascorbate-deficient mutants of Arabidopsis grow in high light despite chronic photooxidative stress. Plant Physiol 134(3):1163–1172. https://doi.org/10.1104/pp.103.032375
Müller-Schüssele SJ, Wang R, Gütle DD, Romer J, Rodriguez-Franco M, Scholz M, Buchert F, Lüth VM, Kopriva S, Dörmann P, Schwarzländer M, Reski R, Hippler M, Meyer AJ (2020) Chloroplasts require glutathione reductase to balance reactive oxygen species and maintain efficient photosynthesis. Plant J 103(3):1140–1154. https://doi.org/10.1093/plphys/kiab019
Müller-Schüssele SJ, Schwarzländer M, Meyer AJ (2021) Live monitoring of plant redox and energy physiology with genetically encoded biosensors. Plant Physiol 186(1):93–109. https://doi.org/10.1093/plphys/kiab019
Mullineaux PM, Exposito-Rodriguez M, Laissue PP, Smirnoff N (2018) ROS-dependent signalling pathways in plants and algae exposed to high light: Comparisons with other eukaryotes. Free Radic Biol Med 122:52–64. https://doi.org/10.1016/j.freeradbiomed.2018.01.033
Noctor G, Arisi A-CM, Jouanin L, Valadier M-H, Roux Y, Foyer CH (1997) Light-dependent modulation of foliar glutathione synthesis and associated amino acid metabolism in poplar overexpressing γ-glutamylcysteine synthetase. Planta 202(3):357–369. https://doi.org/10.1007/s004250050138
Noctor G, Arisi A-CM, Jouanin L, Foyer CH (1999) Photorespiratory glycine enhances glutathione accumulation in both the chloroplastic and cytosolic compartments. J Exp Bot 50(336):1157–1167. https://doi.org/10.1093/jxb/50.336.1157
Noctor G, Gomez L, Vanacker H, Foyer CH (2002) Interactions between biosynthesis, compartmentation and transport in the control of glutathione homeostasis and signalling. J Exp Bot 53(372):1283–1304. https://doi.org/10.1093/jexbot/53.372.1283
Noctor G, Mhamdi A, Chaouch S, Han Y, Neukermans J, Marquez-Garcia B, Queval G, Foyer CH (2012) Glutathione in plants: an integrated overview. Plant Cell Environ 35(2):454–484. https://doi.org/10.1111/j.1365-3040.2011.02400.x
Noctor G, Mhamdi A, Queval G, Foyer CH (2013) Regulating the redox gatekeeper: vacuolar sequestration puts glutathione disulfide in its place. Plant Physiol 163(2):665–671. https://doi.org/10.1104/pp.113.223545
Noctor G, Reichheld J-P, Foyer CH (2018) ROS-related redox regulation and signaling in plants. Sem Cell Dev Biol 80:3–12. https://doi.org/10.1016/j.semcdb.2017.07.013
Olson KR (2020) Reactive oxygen species or reactive sulfur species: why we should consider the latter. J Exp Biol 223(Pt 4):jeb196352. https://doi.org/10.1242/jeb.196352
Öztetik E (2008) A tale of plant glutathione s-transferases: since 1970. Bot Rev 74(3):419–437. https://doi.org/10.1016/j.semcdb.2017.07.013
Pariona A (2019) What are the world’s most important staple foods? https://www.worldatlas.com/articles/most-important-staple-foods-in-the-world.html. Accessed 10 December 2021
Parisy V, Poinssot B, Owsianowski L, Buchala A, Glazebrook J, Mauch F (2007) Identification of PAD2 as a gamma-glutamylcysteine synthetase highlights the importance of glutathione in disease resistance of Arabidopsis. Plant J 49(1):159–172. https://doi.org/10.1111/j.1365-313X.2006.02938.x
Pearcy RW (1998) Acclimation to sun and shade. In: Raghavendra AS (ed) Photosynthesis: a comprehensive treatise. Cambridge University Press, Cambridge, pp 250–263
Pedmale UV, Huang S-SC, Zander M, Cole BJ, Hetzel J, Ljung K, Reis PAB, Sridevi P, Nito K, Nery JR, Ecker JR, Chory J (2016) Cryptochromes interact directly with PIFs to control plant growth in limiting blue light. Cell 164(1–2):233–245. https://doi.org/10.1016/j.cell.2015.12.018
Pham VN, Kathare PK, Huq E (2018) Phytochromes and phytochrome interacting factors. Plant Physiol 176(2):1025–1038. https://doi.org/10.1104/pp.17.01384
Pradedova EV, Nimaeva OD, Rakevich AL, Salyaev RK (2019) Comparative analyses of glutathione system of vacuoles and leucoplasts isolated from the storage parenchyma cells of dormant red beetroots (Beta vulgaris L.). Plant Physiol Biochem 145:52–63
Queval G, Jaillard D, Zechmann B, Noctor G (2011) Increased intracellular H2O2 availability preferentially drives glutathione accumulation in vacuoles and chloroplasts. Plant Cell Environ 34(1):21–32. https://doi.org/10.1111/j.1365-3040.2010.02222.x
Rai N, Morales LO, Aphalo PJ (2021) Perception of solar UV radiation by plants: photoreceptors and mechanisms. Plant Physiol 186(3):1382–1396. https://doi.org/10.1093/plphys/kiab162
Roach T, Stöggl W, Baur T, Kranner I (2018) Distress and eustress of reactive electrophiles and relevance to light stress acclimation via stimulation of thiol/disulphide-based redox defences. Free Radic Biol Med 122:65–73. https://doi.org/10.1016/j.freeradbiomed.2018.03.030
Runge S, Sperling U, Frick G, Apel K, Armstrong GA (1996) Distinct roles for light-dependent NADPH:protochlorophyllide oxidoreductases (POR) A and B during greening in higher plants. Plant J 9(4):513–523. https://doi.org/10.1046/j.1365-313X.1996.09040513.x
Samuolienė G, Viršilė A, Miliauskienė J, Haimi PJ, Laužikė K, Brazaitytė A, Duchovskis P (2020) The physiological response of lettuce to red and blue light dynamics over different photoperiods. Front Plant Sci 11:610174. https://doi.org/10.3389/fpls.2020.610174
Scheibe R, Dietz K-J (2012) Reduction-oxidation network for flexible adjustment of cellular metabolism in photoautotrophic cells. Plant Cell Environ 35(2):202–216. https://doi.org/10.1111/j.1365-3040.2011.02319.x
Sheerin DJ, Hiltbrunner A (2017) Molecular mechanisms and ecological function of far-red light signalling. Plant Cell Environ 40(11):2509–2529. https://doi.org/10.1111/pce.12915
Takahama U (2004) Oxidation of vacuolar and apoplastic phenolic substrates by peroxidase: physiological significance of the oxidation reactions. Phytochem Rev 3(1–2):207–219. https://doi.org/10.1023/B:PHYT.0000047805.08470.e3
Toldi D, Gyugos M, Darkó É, Szalai G, Gulyás Z, Gierczik K, Székely A, Boldizsár Á, Galiba G, Müller M, Simon-Sarkadi L, Kocsy G (2019) Light intensity and spectrum affect metabolism of glutathione and amino acids at transcriptional level. PLoS ONE 14(12):e0227271. https://doi.org/10.1371/journal.pone.0227271
Tolin S, Arrigoni G, Trentin AR, Vljovic-Jovanovic S, Pavato M, Zechmann B, Masi A (2013) Biochemical and quantitative proteomics investigations in Arabidopsis ggt1 mutant leaves reveal a role for the gamma-glutamyl cycle in plant’s adaptation to environment. Proteomics 13(12–13):2031–2045. https://doi.org/10.1002/pmic.201200479
Trnka M, Feng S, Semenov MA, Olesen JE, Kersebaum KC, Rötter RP, Semerádová D, Klem K, Huang W, Ruiz-Ramos M, Hlavinka P, Meitner J, Balek J, Havlík P, Büntgen U (2019) Mitigation efforts will not fully alleviate the increase in water scarcity occurrence probability in wheat-producing areas. Sci Adv 5(9):eaau2406. https://doi.org/10.1126/sciadv.aau2406
Turkan I, Uzilday B, Dietz K-J, Bräutigam A, Ozgur R (2018) Reactive oxygen species and redox regulation in mesophyll and bundle sheath cells of C4 plants. J Exp Bot 69(14):3321–3331. https://doi.org/10.1093/jxb/ery064
Vanacker H, Carver TLW, Foyer CH (1998) Pathogen-induced changes in the antioxidant status of the apoplast in barley leaves. Plant Physiol 117(3):1103–1114. https://doi.org/10.1104/pp.117.3.1103
Viczián A, Klose C, Ádám É, Nagy F (2017) New insights of red light-induced development. Plant Cell Environ 40(11):2457–2468. https://doi.org/10.1111/pce.12880
Vidović M, Morina F, Milić-Komić S, Vuleta A, Zechmann B, Prokić L, Veljović Jovanović S (2016) Characterisation of antioxidants in photosynthetic and non-photosynthetic leaf tissues of variegated Pelargonium zonale plants. Plant Biol 18(4):669–680. https://doi.org/10.1111/plb.12429
Wahlländer A, Soboll S, Sies H, Ingrid L, Monika M (1979) Hepatic mitochondrial and cytosolic glutathione content and the subcellular distribution of GSH- S -transferases. FEBS Lett 97(1):138–140. https://doi.org/10.1016/0014-5793(79)80069-1
Zechmann B (2014) Compartment-specific importance of glutathione during abiotic and biotic stress. Front Plant Sci 5:566. https://doi.org/10.3389/fpls.2014.00566
Zechmann B (2017) Diurnal changes of subcellular glutathione content in Arabidopsis thaliana. Biol Plant 61(4):791–796. https://doi.org/10.1007/s10535-017-0729-4
Zechmann B (2020) Subcellular roles of glutathione in mediating plant defense during biotic stress. Plants 9(9):1067. https://doi.org/10.3390/plants9091067
Zechmann B, Müller M (2010) Subcellular compartmentation of glutathione in dicotyledonous plants. Protoplasma 246(1–4):15–24. https://doi.org/10.1007/s00709-010-0111-2
Zechmann B, Zellnig G, Urbanek-Krajnc A, Müller M (2007) Artificial elevation of glutathione affects symptom development in ZYMV-infected Cucurbita pepo L. plants. Arch Virol 152(4):747–762. https://doi.org/10.1007/s00705-006-0880-2
Zechmann B, Mauch F, Sticher L, Müller M (2008a) Subcellular immunocytochemical analysis detects the highest concentrations of glutathione in mitochondria and not in plastids. J Exp Bot 59(14):4017–4027. https://doi.org/10.1093/jxb/ern243
Zechmann B, Müller M, Zellnig G (2008) Modified levels of cysteine affect glutathione metabolism in plant cells. In: Khan NA, Singh S, Umar S (eds) Sulfur assimilation and abiotic stress in plants. Springer, Berlin Heidelberg, pp 193–206
Zechmann B, Müller M, Möstl S, Zellnig G (2021) Three-dimensional quantitative imaging of Tobacco mosaic virus and Zucchini yellow mosaic virus induced ultrastructural changes. Protoplasma 258:1201–1211. https://doi.org/10.1007/s00709-021-01626-0
Zha L, Zhang Y, Liu W (2019) Dynamic responses of ascorbate pool and metabolism in lettuce to long-term continuous light provided by red and blue LEDs. Environ Exp Bot 163:15–23. https://doi.org/10.1016/j.envexpbot.2019.04.003
Zipor G, Oren-Shamir M (2013) Do vacuolar peroxidases act as plant caretakers? Plant Sci 199–200:41–47. https://doi.org/10.1016/j.plantsci.2012.09.018
Funding
Open access funding provided by University of Graz. This work was supported by the Hungarian National Research, Development and Innovation Office (Grant K131638).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest. The funders had no role in the design of the review; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visithttp://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gasperl, A., Zellnig, G., Kocsy, G. et al. Organelle-specific localization of glutathione in plants grown under different light intensities and spectra. Histochem Cell Biol 158, 213–227 (2022). https://doi.org/10.1007/s00418-022-02103-2
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00418-022-02103-2