
Abstract
The Y chromosome is male-specific and is important for spermatogenesis and male fertility. However, the Y chromosome is poorly characterized due to massive palindromes and inverted repeats, which increase the likelihood of genomic rearrangements, resulting in short tandem repeats on the Y chromosome or long fragment deletions. The present study reports a large-scale (2.573~2.648 Mb) deletion in the Yp11.2 region in a Chinese population based on the analysis of 34 selected Y-specific sequence-tagged sites and subsequent sequencing of the breakpoint junctions on the Y chromosome from 5,068,482–5,142,391 bp to 7,715,462–7,716,695 bp. The results of sequence analysis indicated that the deleted region included part or all of the following five genes: PCDH11Y, TSPY, AMELY, TBL1Y, and RKY. These genes are associated with spermatogenesis or amelogenesis and various other processes; however, specific physiological functions and molecular mechanisms of these genes remain unclear. Notably, individuals with this deletion pattern did not have an obvious pathological phenotype but manifested some degree of amelogenesis imperfecta.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The human genome contains many genetic variants. Two decades of the studies of human Y chromosome variability have determined a number of aspects of population histories and male-biased behaviors. The Y chromosome contains the PAR (pseudoautosomal region) and MSY (male-specific region of the Y chromosome). During meiosis, the PARs at both ends can recombine with the X chromosome, and the MSY does not recombine, resulting in paternal haploid inheritance [1]. This inheritance pattern provides for unique advantages of the use of the STR loci located in the MSY region in forensic science. The human MSY was fully sequenced in 2003 [2]; however, the complexity of the Y chromosome sequences, which are rich in segmental duplications and repeats, makes it almost impossible to accurately assemble these sequences using short-read sequencing technologies.
Human MSY sequences can be classified into the following three major classes: the X-degenerate (XDG) region with variable degrees of sequence similarity to the X chromosome; the ampliconic segment composed of sequences with high similarity to other sequences in the MSY and containing a large number of palindromes; and the X-transposed region (XTR) transferred from the long arm of the X chromosome (Fig. 1a).

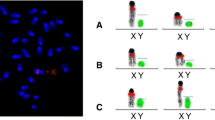
The genotyping schedule of STS loci and the deletion map in this study. a Ideogram of the human Y chromosome. b Expanded view of human MSY euchromatin showing palindromes and genes as arrowheads pointing in the direction of transcription. The location of Y-STRs in the MSY region is shown below. c Scheme of low-resolution mapping based on known STSs. d Scheme of high-resolution mapping based on newly selected loci
The MSY contains a large number of highly homologous repetitive sequences, resulting in a highly unstable structure in this region susceptible to NAHR (nonallelic homologous recombination) within the chromosome, which leads to structural rearrangements in the Y chromosome, such as deletions, inversions, or repetitions. The MSY has a considerable impact on the forensic identification of Y-STR (STR of the Y chromosome) dependence. For example, AMELY (amelogenin Y-linked)-negative male samples can be incorrectly genotyped as females [3]. During evolution, the Y chromosome has acquired many testis-specific genes responsible for spermatogenesis [4]. Alterations in these genes are associated with several male fertility-specific traits. For example, the lack of the AZF (azoospermia factor) region can alter spermatogenesis and may result in male infertility, which is primary manifested as azoospermia and oligospermia [4].
The Y chromosome is important for spermatogenesis and male fertility and may be involved in schizophrenia and other diseases [5,6,7]. Moreover, males may play a larger role than females in the demic diffusion model based on the analysis of the Y chromosome [8]. The suppression of recombination causes MSY degeneration, intrachromosomal rearrangements, evolution of ampliconic repeat regions, and the accumulation of male gametogenesis genes. The loss of the Y chromosome in the peripheral blood was recently shown to be associated with increased risk for all-cause mortality and diseases, such as various forms of cancer, Alzheimer’s disease, and other conditions in aging men.
The frequencies of the deletions of Y-STRs located in the MSY region, such as DYS393 and DYS19, have been reported to increase in various populations; thus, the types of identified deletions in the Yp11.2 region have been gradually diversified [9,10,11]. However, Y-STRs are scattered on the Y chromosome, and the intermediate space is large. Deletions of the corresponding regions of the Y chromosome represented by invalid amplification of one or several Y-STRs are complex and diverse. The detection of other genetic markers in these regions is required to detect the presence of the STS sites and to define the deletions of these regions of the Y chromosome in detail [12].
The present Yp11.2 deletion study was initiated based on the observation of Amelogenin Y and Y-STR (AMELY-DYS570-DYS576) null alleles in China in forensic studies. The present study identified 34 STS loci that can be used for the detection of Yp11.2 region deletions by screening the STS sites in the Yp11.2 region. The data obtained based on these STSs were used to accurately determine the location of the deletion junction in the Yp11.2 region of two Chinese males from Yp 5,068,482–5,142,391 bp to Yp 7,715,462–7,716,695 bp.
Materials and methods
Samples and DNA extraction
Yp11.2 deletion, positive, and negative samples were provided by the center for forensic science of Jining Medical University. DNA was extracted by a genomic DNA extraction kit (Tiangen Biotech, China) from fresh peripheral blood according to the manufacturer’s instructions.
STS loci mapping
Locations of the SRY and STS loci in the Y chromosome were obtained from the UCSC Genome database (http://genome.ucsc.edu/cgi-bin/hgTracks), and the sequence of the Y chromosome was obtained from the NCBI database (https://www.ncbi.nlm.nih.gov/nuccore/NC_000024.10?from=2686000&to=7912000&report=genbank&strand=true). The primers were designed by using Lasergene and BLAST on the NCBI website. The primer sequences are shown in Table 1.
Gradient PCR was used to determine suitable annealing temperature for each primer pair. Annealing temperatures are shown in Table 1. Genomic DNA extracted from the blood sample was used as a template for PCR amplification. PCR amplification was carried out using 10 μL of 2×Taq PCR Master Mix II (TIANGEN, China), 0.5 μL of each of the primers (10 μmol/L), 2 μL of DNA template (200~400 ng/μL), and 7 μL of nuclease-free water in a total volume of 20 μL. Thermal cycling conditions were as follows: 95 °C for 5 min, 32 cycles at 94 °C for 30 s, annealing temperature for 30 s, and 72 °C for 40 s, and a final extension at 72 °C for 10 min. Amplified PCR products were separated by 1.0% agarose gel electrophoresis and visualized under a UV light source.
Long PCR amplification
Long PCR amplification was performed using a KOD FX Neo kit (Toyobo Life Science, Japan) according to the manufacturer’s instruction. The samples contained 10 μL of 2× PCR buffer for KOD FX Neo, 0.5 μL of each of the primers (10 μmol/L), 0.5 μL of dNTP (2 mM each), 1 μL of KOD FX Neo (1.0 unit/μL), 2 μL of DNA template (200~400 ng/μL), and 4 μL of nuclease-free water in a total volume of 20 μL. The thermal cycling conditions were as follows: 95 °C for 5 min, 32 cycles at 94 °C for 30 s, 62 °C for 30 s, and 72 °C for 40 min, and a final extension at 72 °C for 1 h. Amplified PCR products were separated by 0.7% agarose gel electrophoresis and visualized under a UV light source.
Results
Two samples from paternity testing were from males, and all autosomal STRs were successfully genotyped; we were unable to detect AMELY by two different amplification kits. The results of Y-STR genotyping showed that DYS570 and DYS576 were missing; however, DYS456, DYS458, and other Y-STRs were successfully genotyped (Fig. 1b). Therefore, the breakpoint junctions were identified as DYS456-AMELY and DYS576-DYS458.
Sixteen known Y chromosome-specific STS loci located in these two breakpoint regions were used for low-resolution mapping (Fig. 1c). Six STS loci, comprising DYS399, BV703954, sY2207, sY2220, DYS463A18, and DYS288, were present, and ten loci (BV703904, BV703923, sY2137, sY716, DYS256, DYS267, sY2232, sY2233, DYS261, and DYS260) were deleted in both males (Fig. 1c, top). The results of the STS loci amplification are shown in Table 2.
The data indicated that the distal breakpoint was located between BV703954 (5.080 Mb) and BV703904 (5.193 Mb). The proximal breakpoint was located between DYS260 (7.715 Mb) and sY2220 (7.726 Mb) (Fig. 1c, bottom); additional closely spaced STSs (~20 kb) were used for high-resolution mapping of the breakpoint junctions. The distal BV54-04-1, BV54-04-2, BV54-04-3, BV54-04-4, and BV54-04-5 breakpoints were positive, and the BV54-04-10, BV54-04-11, BV54-04-12, BV54-04-13, BV54-04-14, and BV54-04-15 breakpoints were negative. We were unable to determine the presence of the BV54-04-6, BV54-04-7, BV54-04-8, and BV54-04-9 loci due to poor specificity of the binding on the Y chromosome. The proximal sY20-08-1, sY20-08-2, sY20-08-3, sY20-08-4, sY20-08-5, sY20-08-6, and sY20-08-7 loci were positive.
Finally, BV54-04-5, BV54-04-6, BV54-04-7, BV54-04-8, and BV54-04-9 were used as forward primers and sY20-08-1 was used as a reverse primer for long PCR amplification to detect the deletion junction sequences; unfortunately, all results of the long PCR amplifications were negative.
Discussion
The nulls of the AMELY and Y-STR loci in the Yp11.2 region have been frequently reported in the Chinese population. The present study demonstrated that the Y-STRs of two Chinese males contained DYS570 and DYS576 combined with the AMELY deletion, and other Y-STRs were normally amplified. We used a three-stage strategy of combined low-resolution STS mapping, high-resolution STS mapping, and long PCR amplification [8]. A total of 34 STSs between DYS456 and DYS458 (DYS399, BV703954, BV54-04-1, BV54-04-2, BV54-04-3, BV54-04-4, BV54-04-5, BV54-04-10, BV54-04-11, BV54-04-12, BV54-04-13, BV54-04-14, BV54-04-15, BV703904, BV703923, sY2137, sY716, DYS256, DYS267, sY2232, sY2233, DYS261, DYS260, sY20-08-1, sY20-08-2, sY20-08-3, sY20-08-4, sY20-08-5, sY20-08-6, sY20-08-7, sY2207, sY2220, DYS463A18, and DYS288) were mapped to this region. The deletion junctions of the two Chinese males we tested based on these STS loci starting from 5,068,482–5,142,391 to 7,715,462–7,716,695, and the size of the deletion region was 2.573~2.648 Mb. The breakpoint regions were too long (>40 kb) or/and complex to amplify; thus, long PCR amplification did not obtain any long fragments, and all results were negative, suggesting that the length of the missing fragments was close to 2.573 Mb.
The design of STSs for high-resolution mapping in the breakpoint region is difficult because this region has extremely high homology with the X, 3, and 5 chromosomes; thus, it was not possible to precisely locate the deletion site.
Sequence analysis indicated that the missing region contained PCDH11Y (protocadherin 11 Y-linked), TSPY (testis-specific protein Y-linked 1), AMELY, TBL1Y (transducin beta-like 1 Y-linked), and PRKY (protein kinase Y-linked). PCDH11Y plays a role in cell-cell recognition during central nervous system development, belongs to the protocadherin family, and is very closely related to its paralog on the X chromosome. PCDH11Y is essential for spermatogonial differentiation and initiation of meiosis [13] and may also be a candidate marker for susceptibility to psychiatric disorders [14]. TSPY is expressed only in testicular tissue and may be involved in spermatogenesis. Previous studies demonstrated that TSPY1 has physiological functions in the proliferation and differentiation of spermatogonia during spermatogenesis [15]. Remarkably, TSPY1 is also involved in the initiation and development of many tumors by activating the PI3K/AKT and RAS signaling pathways by suppressing IGFBP3 expression [16]. Amelogenins are useful for sex identification and are involved in biomineralization during tooth enamel development [17]. Mutations in a related gene on chromosome X cause X-linked amelogenesis imperfecta. Amelogenin can also be used to monitor bone marrow implantation in patients with sex-mismatched bone marrow transplantation. TBL1Y is a Y-linked homolog of TBL1X that was considered a novel candidate for hereditary hearing loss acting via the Wnt signaling pathway [18]. The expression pattern and function of TBL1Y are unknown; however, this gene may play some other roles in maleness due to specific chromatin binding and transcription corepressor activity. PRKY is similar to the protein kinase X-linked gene in the PAR and is classified as a transcribed pseudogene. However, abnormal recombination between this gene and a related gene on the chromosome X is a frequent cause of XX males and XY females [19]. Meyfour et al. reported alterations in the mRNA and protein levels of TBL1Y, PCDH11Y, ZFY, KDM5D, USP9Y, RPS4Y1, DDX3Y, PRY, XKRY, BCORP1, RBMY, HSFY, and UTY accompanied by changes in intracellular localization during cardiac differentiation of human embryonic stem cells (hESCs) [20]. In some cases, repeated mutations of the AMELY, TBL1Y, and PRKY regions have been detected without obvious deformities [21].
The results of the present study suggest that the C-terminus could have been deleted from the full-length PCDH11Y, even though two males did not show any symptoms of infertility or mental illness. The Y chromosome contains numerous TSPY repeats. Changes in the copy number of TSPY can change the tumorigenic ability of prostate cancer cells in nude mice and may lead to prostate cancer in men. However, deletion of a single copy of TSPY did not result in a phenotype in the present study. Family investigation showed that males of this family generally have poor teeth conditions, such as amelogenesis imperfecta (Fig. S1), with no obvious pathological manifestations. In addition, the effects of this deletion on physiological conditions of the two male subjects cannot be confirmed without systematic health examinations.
The effects of the gene and protein functions on human physiology and pathology require comparative studies. Male infertility is related not only to a certain gene or region in the Y chromosome. Many autosomal proteins loss-of-function mutations of autosomal proteins can also cause male infertility, including the function of the Golgi matrix protein GM130 in male-specific germ cells [22]. Knockout of GM130 resulted in the absence of acrosomes in mice and caused male infertility [23]. Analysis of the Yp11.2 region deletion and individual phenotypes can provide information for future basic studies on male infertility and other related diseases, such as enamel hypoplasia and deafness, and the data can be used in the studies of gene expression and functions of the corresponding proteins.
The majority of the studies on Y chromosome deletions related to male infertility involved the AZF region; however, the relationship between other regions outside the AZF region and male infertility is unknown. The lack of data in these areas is mainly due to the lack of convenient and fast commercial kits. We can optimize the STS primer system used in the present study to construct a multiple amplification system, which can be used for specific amplification of the Yp11.2 region and will be suitable for large-scale screening. Moreover, the present study provides theoretical data and technical support for investigations of the Y chromosome deletions in Chinese men. The system used in the present study is suitable for rapid clinical diagnostic and genetic screening of infertility in men.
The missing regions and related pathological phenotype reported in this study have been determined. The Yp11.2 deletion region identified in the present study can be used as a negative reference to analyze whether the cause of the infertility is associated with microdeletions in the Y chromosome, to exclude the genetic cause and thus confirm the actual pathogenesis of male infertility, and to provide a more accurate reference for the targeted and special treatments. Moreover, identification of the cause of infertility will facilitate early diagnosis and treatment of reproductive disorders in the offspring.
The present study detected 38 STSs in the Yp11.2 region between DYS456 and DYS458, confirming the deletion junction in the two Chinese males from 5,068,482–5,142,391 to 7,715,462–7,716,695, and the size of the deletion region was 2.573~2.648 Mb. A total of 34 chromosome Y-specific STSs were confirmed to be suitable for the deletion mapping in this region. Molecular analysis demonstrated that the missing region contained five genes: PCDH11Y, TSPY, AMELY, TBL1Y, and PRKY. Two males presented with certain signs of amelogenesis imperfecta with no obvious pathological manifestations. The data of the present study can provide theoretical and technical support for the investigations of gene functions and Yp11.2 region deletion mapping and for the development of multiple amplification systems.
Abbreviations
- AMEL:
-
Amelogenin
- AMELX:
-
Amelogenin X-linked
- AMELY:
-
Amelogenin Y-linked
- AZF:
-
Azoospermia factor
- MSY:
-
Male-specific region of Y chromosome
- PAR:
-
Pseudoautosomal region
- PCDH11Y:
-
Protocadherin 11 Y-linked
- PRKY:
-
Protein kinase Y-linked
- SRY:
-
Sex-determining region Y
- STR:
-
Short tandem repeat
- STS:
-
Sequence-tagged site
- TBL1Y:
-
Transducin beta-like 1 Y-linked
- TSPY:
-
Testis-specific protein Y-linked
- Y-STR:
-
Y chromosome STR
References
Hallast P, Jobling MA (2017) The Y chromosomes of the great apes. Hum Genet 136:511–528. https://doi.org/10.1007/s00439-017-1769-8
Skaletsky H, Kuroda-Kawaguchi T, Minx PJ, Cordum HS, Hillier L, Brown LG, Repping S, Pyntikova T, Ali J, Bieri T, Chinwalla A, Delehaunty A, Delehaunty K, du H, Fewell G, Fulton L, Fulton R, Graves T, Hou SF, Latrielle P, Leonard S, Mardis E, Maupin R, McPherson J, Miner T, Nash W, Nguyen C, Ozersky P, Pepin K, Rock S, Rohlfing T, Scott K, Schultz B, Strong C, Tin-Wollam A, Yang SP, Waterston RH, Wilson RK, Rozen S, Page DC (2003) The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 423:825–837. https://doi.org/10.1038/nature01722
Santos FR, Pandya A, Tyler-Smith C (1998) Reliability of DNA-based sex tests. Nat Genet 18:103–103. https://doi.org/10.1038/ng0298-103
Krausz C, Casamonti E (2017) Spermatogenic failure and the Y chromosome. Hum Genet 136:637–655. https://doi.org/10.1007/s00439-017-1793-8
Hirata T, Hishimoto A, Otsuka I, Okazaki S, Boku S, Kimura A, Horai T, Sora I (2018) Investigation of chromosome y loss in men with schizophrenia. Neuropsychiatr Dis Treat 14:2115–2122
Kimura A, Hishimoto A, Otsuka I, Okazaki S, Boku S, Horai T, Izumi T, Takahashi M, Ueno Y, Shirakawa O, Sora I (2018) Loss of chromosome Y in blood, but not in brain, of suicide completers. PLoS One 13:e0190667. https://doi.org/10.1371/journal.pone.0190667
Forsberg LA (2017) Loss of chromosome Y (LOY) in blood cells is associated with increased risk for disease and mortality in aging men. Hum Genet 136:657–663. https://doi.org/10.1007/s00439-017-1799-2
Wen B, Li H, Lu D, Song X, Zhang F, He Y, Li F, Gao Y, Mao X, Zhang L, Qian J, Tan JZ, Jin JZ, Huang W, Deka R, Su B, Chakraborty R, Jin L (2004) Genetic evidence supports demic diffusion of Han culture. Nature 431:302–305. https://doi.org/10.1038/nature02878
Ma Y, Kuang JZ, Zhang J, Wang GM, Wang YJ, Jin WM, Hou YP (2012) Y chromosome interstitial deletion induced Y-STR allele dropout in AMELY-negative individuals. Int J Legal Med 126:713–724. https://doi.org/10.1007/s00414-012-0720-8
Balaresque P, Parkin EJ, Roewer L, Carvalho-Silva DR, Mitchell RJ, van Oorschot RA, Henke J, Stoneking M, Nasidze I, Wetton J et al (2009) Genomic complexity of the Y-STR DYS19: inversions, deletions and founder lineages carrying duplications. Int J Legal Med 123:15–23. https://doi.org/10.1007/s00414-008-0253-3
Borovko S, Shyla A, Korban V, Borovko A (2015) Amelogenin test abnormalities revealed in Belarusian population during forensic DNA analysis. Forensic Sci Int-Genet 15:98–104. https://doi.org/10.1016/j.fsigen.2014.10.014
Yong RY, Gan LS, Chang YM, Yap EP (2007) Molecular characterization of a polymorphic 3-Mb deletion at chromosome Yp11.2 containing the AMELY locus in Singapore and Malaysia populations. Hum Genet 122:237–249. https://doi.org/10.1007/s00439-007-0389-0
Anilkumar TR, Devi AN, Pillai SM, Jayakrishnan CK, Oommen OV, Kumar PG (2017) Expression of protocadherin 11Yb (PCDH11Yb) in seminal germ cells is correlated with fertility status in men. Reprod Fertil Dev 29:2100–2111. https://doi.org/10.1071/RD16478_AC
Durand CM, Kappeler C, Betancur C, Delorme R, Quach H, Goubran-Botros H, Melke J, Nygren G, Chabane N, Bellivier F, Szoke A, Schurhoff F, Rastam M, Anckarsater H, Gillberg C, Leboyer M (2006) Expression and genetic variability of PCDH11Y, a gene specific to Homo sapiens and candidate for susceptibility to psychiatric disorders. Am J Med Genet 141B:67–70. https://doi.org/10.1002/ajmg.b.30229
Zhang GW, Wu Y, Luo Z, Guan J, Wang L, Luo X, Zuo F (2019) Comparison of Y-chromosome-linked TSPY, TSPY2, and PRAMEY genes in Taurus cattle, yaks, and interspecific hybrid bulls. J Dairy Sci 102:6263–6275. https://doi.org/10.3168/jds.2018-15680
Tu W, Yang B, Leng X, Pei X, Xu J, Liu M, Dong Q, Tao D, Lu Y, Liu Y, Yuan Y (2019) Testis-specific protein, Y-linked 1 activates PI3K/AKT and RAS signaling pathways through suppressing IGFBP3 expression during tumor progression. Cancer Sci 110:1573–1586. https://doi.org/10.1111/cas.13984
Gibson CW, Yuan ZA, Hall B, Longenecker G, Chen E, Thyagarajan T, Sreenath T, Wright JT, Decker S, Piddington R, Harrison G, Kulkarni AB (2001) Amelogenin-deficient mice display an amelogenesis imperfecta phenotype. J Biol Chem 276:31871–31875. https://doi.org/10.1074/jbc.M104624200
Di Stazio M, Collesi C, Vozzi D, Liu W, Myers M, Morgan A, PA DA, Girotto G, Rubinato E, Giacca M, Gasparini P (2019) TBL1Y: a new gene involved in syndromic hearing loss. Eur J Hum Genet 27:466–474. https://doi.org/10.1038/s41431-018-0282-4
Nakashima S, Watanabe Y, Okada J, Ono H, Nagata E, Fukami M, Ogata T (2013) Critical role of Yp inversion in PRKX/PRKY-mediated Xp; Yp translocation in a patient with 45, X testicular disorder of sex development. Endocr J 60:1329–1334. https://doi.org/10.1507/endocrj.ej13-0334
Meyfour A, Ansari H, Pahlavan S, Mirshahvaladi S, Rezaei-Tavirani M, Gourabi H, Baharvand H, Salekdeh GH (2017) Y chromosome missing protein, TBL1Y, may play an important role in cardiac differentiation. J Proteome Res 16:4391–4402. https://doi.org/10.1021/acs.jproteome.7b00391
Murphy KM, Cohen JS, Goodrich A, Long PP, Griffin CA (2007) Constitutional duplication of a region of chromosome Yp encoding AMELY, PRKY, and TBL1Y: implications for sex chromosome analysis and bone marrow engraftment analysis. J Mol Diagn 9:408–413. https://doi.org/10.2353/jmoldx.2007.060198
Han F, Liu C, Zhang L, Chen M, Zhou Y, Qin Y, Wang Y, Chen M, Duo S, Cui X, Bao S, Gao F (2017) Globozoospermia and lack of acrosome formation in GM130-deficient mice. Cell Death Dis 8:e2532. https://doi.org/10.1038/cddis.2016.414
Jiang Y, Liu Y, Han F, Zhou J, Zhang X, Xu J, Yu Z, Zhao S, Gao F, Zhao H (2020) Loss of GM130 does not impair oocyte meiosis and embryo development in mice. Biochem Bioph Res Co 532:336–340. https://doi.org/10.1016/j.bbrc.2020.08.05
Acknowledgements
We are grateful to the AMELY-negative individual and other participants for their cooperation in this study. We also want to thank the AJE editorial team for language editing.
Funding
This work was supported by the Natural Science Foundation of Shandong Province, China (Grant ZR2019PH076), the Open Project of Forensic Medicine Key Laboratory of Shanxi Province, China (SFM2019001), and the Research Support Fund for Young Teachers of Jining Medical University (JY2017FY002).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Bloodstains and fresh peripheral blood samples were acquired from individuals in the Chinese Han population of Jining City. All procedures performed in the present study were approved by the Medical Ethics Committee of Jining Medical School in accordance with the 2013 Helsinki Declaration.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(PNG 2556 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pang, Q., Lin, Q., Wang, D. et al. Molecular characterization of the Yp11.2 region deletion in the Chinese Han population. Int J Legal Med 135, 1351–1358 (2021). https://doi.org/10.1007/s00414-021-02596-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00414-021-02596-x