
Abstract
Sodium–glucose cotransporter 2 inhibitors (SGLT2i), a new drug class initially designed and approved for treatment of diabetes mellitus, have been shown to exert pleiotropic metabolic and direct cardioprotective and nephroprotective effects that extend beyond their glucose-lowering action. These properties prompted their use in two frequently intertwined conditions, heart failure and chronic kidney disease. Their unique mechanism of action makes SGLT2i an attractive option also to lower the rate of cardiac events and improve overall survival of oncological patients with preexisting cardiovascular risk and/or candidate to receive cardiotoxic therapies. This review will cover biological foundations and clinical evidence for SGLT2i modulating myocardial function and metabolism, with a focus on their possible use as cardioprotective agents in the cardio-oncology settings. Furthermore, we will explore recently emerged SGLT2i effects on hematopoiesis and immune system, carrying the potential of attenuating tumor growth and chemotherapy-induced cytopenias.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sodium‐glucose cotransporter 2 inhibitors (SGLT2i) represent a class of antidiabetic medications with a unique story. Initially approved for the treatment of type 2 diabetes mellitus, thanks to their glycosuric action, subsequently showed cardioprotective and nephroprotective effects that made them one of the most innovative drugs in cardiology and nephrology over the last decades [20, 164].
SGLT2i reduce serum glucose by inhibiting renal tubular glucose reabsorption, thus promoting urinary glucose excretion [143]. Two types of sodium‐glucose cotransporters are present in the nephron, mainly located in the S2 and S3 segments of the proximal tubule, reabsorbing the majority of filtrated urinary glucose [155]. By inhibiting SGLT2, these drugs impair glucose reabsorption in the proximal tubule, determining a reduction of the renal threshold of glycosuria. Canagliflozin, dapagliflozin and empagliflozin are representatives of SGLT2i, also known as gliflozins [97].
An important hemodynamic effect of gliflozins is related to blood pressure reduction, through tubulo-glomerular feedback stimulation reversal, glycosuria and natriuresis. Moreover, SGLT2i may improve blood pressure control by inhibiting the sympathetic nervous system [100]. Of importance, SGLT2i promote weight loss and lipid metabolism shift, inducing visceral fat reduction, as well as increased lipolysis [72, 133]. The ramifications of SGLT2 inhibition in lipid metabolism also enable to understand their atheroprotective effect [8, 133]. In this context, an emerging property of gliflozins is related to their anti-inflammatory action, causing the attenuation of interleukin (IL)-6, tumor necrosis factor (TNF), interferon (IFN)-γ, NF-κβ, toll-like receptor (TLR)-4 and transforming growth factor (TGF)-β [17, 74]. Both in vitro and in vivo models showed that gliflozins may limit inflammation, targeting a wide array of pathways that play a pivotal role in the development of atherosclerosis [107, 128].
In 2015, the EMPA-REG OUTCOME trial was the first study to focus on the cardiovascular outcomes related to treatment with SGLT2i. In 7020 patients with type 2 diabetes mellitus and high cardiovascular risk, randomized to empagliflozin or placebo and evaluated at a median follow-up of 3.1 years, a reduction in heart failure (HF) hospitalization, cardiovascular death and mortality was observed in the empagliflozin arm compared to placebo [13]. These promising results laid the groundwork for successful trials with SGLT2i also in HF patients with either reduced or preserved ejection fraction (HFrEF and HFpEF, respectively) [6, 91, 130], afterwards confirmed by several meta-analyses [19, 162]. Overall, available evidences made SGLT2i one of the “four pillars” of HFrEF treatment and the first recommended medication in HF with mildly-reduced ejection fraction (HFmrEF) and HFpEF, according to most recent updates of guidelines [89, 90].
Growing data also introduced the possibility of adding SGLT2i to the pharmacological armamentarium of cardiovascular disease prevention [33] or treatment of subclinical cardiac dysfunction, including those cases observed in cancer patients exposed to potentially-cardiotoxic agents [21, 95]. At the same time, preclinical data suggested a role of SGLT2i in preventing cancer progression and mitigating cytopenia induced by chemotherapy [95].
This review will, therefore, focus on the role of SGLT2i as modulators of the cardiovascular system, with special emphasis being paid to myocardial function and metabolism pathways. The biological rationale and clinical evidence for bringing SGLT2i at the center of the Cardio-Oncology stage will be presented (Fig. 1). We will also shed light on possible research directions and their implications in cancer patients’ management.

Role of sodium glucose cotransporter 2 (SGLT2) inhibitors across the spectrum of cardiotoxicity, solid tumors and hematologic malignancies
Mechanisms of cardiac protection in cancer patients through SGLT2 inhibition
In preclinical models, SGLT2i have consistently shown to exert direct beneficial effects on the cardiac tissue, especially in the setting of chronic conditions such as HF. These effects reflect SGLT2i modulation of pro-inflammatory and oxidative stress processes, ion transport, myocardial sodium and calcium homeostasis, as well as metabolic/mitochondrial pathways, such as ketone bodies production [25, 96, 120, 164]. Given that these processes are also supposed to be involved in chemotherapy-associated cardiac injury, there is growing interest in the potential role played by SGLT2i in the field of Cardio-Oncology [31].
Biological basis of SGLT2i protection against anthracycline-induced cardiotoxicity
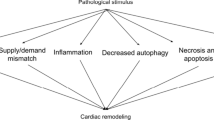
Anthracyclines represent the backbone treatment for several solid and hematological malignancies and are associated with potential dose-dependent cardiotoxic effects, eventually leading to the development of subclinical left ventricular dysfunction, overt HF and malignant arrhythmias [125]. There is a plethora of mechanisms involved in anthracycline-induced cardiotoxicity, among others (i) regulation of macrophage phenotype expression; (ii) generation of highly reactive oxygen species (ROS) inducing membrane lipid peroxidation and causing a direct damage to cardiac myocytes [52,53,54]; (iii) inhibition of cardiac topoisomerase IIβ, leading to transcriptome changes, mitochondrial dysfunction and apoptosis [56, 116, 126, 134, 135, 137, 159]; (iv) downregulation of ferroptosis. Main mechanisms of anthracycline induced cardiotoxicity are sketched in Fig. 2.

Molecular mechanisms of anthracycline-induced cardiomyocyte death. Anthracyclines, in particular doxorubicin, treatment initiates multiple pathways including upregulation of death receptors, calcium overload, disruption in iron homeostasis with lipid peroxidation, generation of ROS with final DNA damage and cell death. AMPK 5ʹ adenosine monophosphate-activated protein kinase, DNA deoxyribonucleic acid, Fe2+ iron, GLC glucose, MyD88 Myeloid differentiation primary response 88, IL-1β R Interleukin-1 beta receptor, NF-kB Nuclear factor kappa-light-chain-enhancer of activated B cells, ROS reactive oxygen species, TLR toll-like receptor
Regulation of macrophage phenotype expression
Macrophages are important actors of immune response, being involved in both activation and resolution of the inflammatory process, and in tissue repair through fibrosis [64]. In brief, the different roles played by macrophages are explained by their phenotype expression, with M1 and M2 macrophages exerting, respectively, induction and resolution of inflammation [85].
M2 macrophages have been shown to influence post-myocardial infarction remodeling in mice models through the action of IL-13, which modulates monocyte infiltration in the wounded cardiac tissue and their polarization [55]. M2 macrophages contribute to the control of the inflammatory process through several actions; in particular, M2a macrophages are associated with arginase production, which is believed to determine collagen deposition and fibrosis development. On the other hand, M2c macrophages, preferentially activated by SGLT2i, are believed to favor IL-10 expression, a suppressor cytokine that promotes the resolution of the fibrotic process [76]. It was in keeping with these notions that four weeks treatment with SGLT2i promoted cardiac repair and reduced pathological remodeling in a rat model of myocardial infarction [76].
SGLT2i also attenuated cardiac fibrosis via regulating the signal transducer and transcription activator STAT3-dependent pathway, consequently reducing infiltration of myofibroblast and collagen accumulation [76]. At the same time, empagliflozin limited cardiac fibrosis by inhibiting the NADPH-oxidase activity following ischemic injury [79].
Although multiple studies investigated the effects of SGLT2i on macrophages and fibrosis, our understanding of the precise role of cardiac macrophages in doxorubicin-induced cardiotoxicity is still limited. The imbalance between macrophage phenotypes and their infiltration in the myocardium seems to be involved also in anthracycline-induced myocardial injury [39]. Studies in mouse models suggest that acute doxorubicin treatment increases M1 macrophage population, while suppressing the M2 counterpart [15, 73]. Moreover, studies of an IL-12p35-KO mouse mode showed that IL-12 and IL-35 deficiency exacerbated doxorubicin-induced myocardial injury through promotion of M1 macrophage differentiation, increase of pro-inflammatory cytokines and reduction of M2 macrophage-related anti-inflammatory cytokines [157]. Other studies demonstrated that IL-22 is a critical regulator of macrophage differentiation in response to cardiac injury [158], while IL-22 deficiency reverses doxorubicin-induced cardiac imbalance of M1 versus M2 macrophages, increasing the M2 population. This effect was associated with reduced cardiomyocyte vacuolization and apoptosis, with concomitant improvement of global cardiac function [158].
Modulation of cardiomyocyte Ca 2+ /Na + homeostasis and downregulation of inflammasome
The inflammasome is a multiprotein complex that plays a role in the activation and perpetuation of the inflammatory process, with nucleotide-binding oligomerization domain (NOD)-like receptor pyrin domain containing protein 3 (NLRP3) being a critical component of the innate immune system. This mediates caspase-1 activation and the secretion of proinflammatory cytokines in response to microbial infection and cellular damage [61]. The inflammasome, in order to exert its function, needs to be activated through three stimulus events: ionic flux, ROS and/or organelle damage.
Indeed, multiple studies investigated the role of the inflammasome in myocardial injury and its interaction with gliflozins. In acute myocardial infarction, the activation of NLRP3 inflammasome is critical, as shown by the reduced extension of infarct size in Nlrp3−/− mice [61, 93], although its role in the ischemia–reperfusion (I/R) damage is still controversial [58, 123]. In mice models of HF, empagliflozin blunts the decline in cardiac function, also reducing NLRP3 inflammasome activation and IL1β secretion [66]. Likewise, empagliflozin attenuates macrophagic NLRP3 inflammasome activation in patients with type 2 diabetes [68]. As well, a series of reports showed that empagliflozin positively regulates cardiomyocyte homeostasis and inflammasome activity. Of note, in preclinical studies, SGLT2i inhibited the Na+/H+ exchanger, leading to lower cytosolic concentration of Na+ and Ca2+ in cardiomyocytes [14, 141]. In diabetic rats, empagliflozin exerts a potential anti-diabetic effect through prevention of Ca2+/Na+ dysregulation in cardiomyocytes and decrease in ROS, leading to improved cardiac function, attenuation of ventricular hypertrophy and correction of prolonged QT interval [75].
In the Cardio-Oncology field, anthracycline exposure increases levels of ROS and Ca2+ in cardiomyocytes, with subsequent mitochondrial dysfunction, inflammasome activation, apoptosis and necrosis [79]. Doxorubicin exposure increases the production of circulating IL-1-β, IL-6 and C reactive-protein (CRP), enhancing through inflammation, the risk of metabolic diseases and cardiovascular manifestations [114]. Indeed, gliflozins counterbalance the various cardiotoxic mechanisms generated by anthracyclines, thus leading to an improvement in dysregulated pathways. Quagliariello et al. demonstrated that incubation of cardiomyocytes from non-diabetic mice with empagliflozin significantly reduced the intracellular Ca2+ content and expression of several pro-inflammatory cytokines. Moreover, the authors demonstrated an improvement in LVEF and radial/longitudinal strain after anthracycline exposure in the empagliflozin mice group [66]. This report also highlighted a role of SGLT2i in the modulation of NLRP3 and myeloid differentiation primary response 88 (MyD88)-related pathways, known to be involved in the cytokine release that characterizes HF [111]. Similarly, the role of empagliflozin in preventing doxorubicin-induced myocardial dysfunction in non-diabetic mice was reported by Sabatino et al. In particular, empagliflozin caused 50% less myocardial fibrosis independently of glycaemic control [121].
Normalization of SGK1 and ENaC myocardial expression
Serum- and Glucocorticoid-Regulated Kinase 1 (SGK1) is highly expressed in the human and murine heart and is upregulated in many pathophysiological settings, including obesity, heart disease and diabetes. SGK1 regulates the expression of a number of ion channels, including epithelial sodium channel (ENaC), which is upregulated in obesity and diabetes [47]. In insulin-resistant female diabetic mice, increased myocardial expression of SGK1 and ENaC caused myocardial fibrosis and eccentric LV hypertrophy, while empagliflozin reduced SGK1 and ENaC levels, as well as related pro-fibrotic signals, leading to an improved diastolic relaxation. These results suggest that the SGLT2i mediated improvement in cardiac function may be modulated, in part, through reduction in SGK1/ENaC activity [47]. In a mouse model, Zhang et al. also demonstrated that the downregulation of SGK1 pathway reduced doxorubicin-mediated cardiotoxicity through the phosphorylation and nuclear translocation of NFκB, leading to reduction of cardiomyocyte inflammation and apoptosis [166]. This would further identify SGK1 downregulation as a possible mechanism of cardiac protection induced by SGLT2i.
Upregulation of ketone body release
Recent in vitro studies suggest that β-hydroxybutyrate (β-OHB), a ketone body that increases during SGLT2i treatment, may protect against anthracyclines by reducing ROS levels and by improving mitochondrial function in cardiomyocytes [102]. On the other hand, ketone serves as an effective source of energy in the failing heart and can mitigate cardiomyocyte damage by reducing ROS and by increasing ATP levels [10]. Modulation of cardiac metabolism might thus represent a rather pivotal mechanism through which SGLT2i could protect against anthracycline cardiotoxicity.
Downregulation of ferroptosis
Ferroptosis is a mechanism of iron-dependent non-apoptotic cell death characterized by iron overload and lipid peroxidation. Ferroptosis occurs in many pathophysiologic conditions and may contribute to acute kidney injury, neuronal death, and cancer cell death as well [36, 62, 81, 82, 154]. Ferroptosis is also involved in cardiovascular diseases such as cardiomyopathies and acute myocardial infarction [32]. In a mouse model of cardiac ischemia–reperfusion injury, the iron chelator desferrioxamine and the glutaminolysis inhibitor compound 968, both with ferroptosis inhibitory activity, reduced myocardial infarct size and improved cardiac function [44, 106].
Canagliflozin mitigates ferroptosis, and by doing so it was shown to improve cardiac function in a preclinical model of HFpEF [83]. This raises interest on SGLT2i as potential cardiac protectant against anthracyclines, especially since ferroptosis is strongly involved in anthracycline cardiotoxicity [167]; however, we are unaware of studies that addressed this hypothesis in detail.
Upregulation of PGC-1α and mitochondrial protection
In preclinical studies, anthracycline-dependent cardiomyocyte damage was fingerprinted by reduced AMP-activated protein kinase (AMPK) expression and double-strand deoxyribonucleic acid (DNA) breaks, which is consistent with anthracyclines intercalating in DNA and blocking the activity of topoisomerase IIβ that is constitutively expressed in cardiomyocytes [76]. As anticipated earlier, topoisomerase IIβ inhibition triggers transcriptome changes that culminate in mitochondrial dysfunction and apoptosis [145]. Doxorubicin would nonetheless induce anthracycline cardiotoxicity by many other mechanisms, which may range from excess of ROS production in mitochondria to suppression of peroxisome proliferator-activated receptor gamma coactivator-1 alpha (PGC-1α), a transcription coactivator that plays a primary role in mitochondrial oxidative phosphorylation and fatty acid oxidation [117, 146, 160]. Studies of ischemic cardiomyopathy in laboratory animals have shown that empagliflozin increases the levels of PGC-1α, leading to mitochondrial protection [161], while other studies showed that similar mechanisms enabled empagliflozin to protect cardiomyocytes against doxorubicin [144].
The molecular mechanisms through which SGLT2i might exert cardioprotection against anthracyclines are summarized in Fig. 3.

Molecular mechanisms through which SGLT2 inhibitors may exert a cardioprotective role in anthracycline-treated patients and in patients developing cancer therapy related cardiac dysfunction. AMPK 5ʹ adenosine monophosphate-activated protein kinase, CKMB Creatine Kinase MB, FGF-21 Fibroblast growth factor-21, NLRP3 nucleotide-binding oligomerization domain-like receptor family pyrin domain containing 3, NTproBNP N-terminal pro B-type natriuretic peptide, MyD88 Myeloid differentiation primary response 88, IL-1β Interleukin-1 beta, NADPH nicotinamide adenine dinucleotide phosphate, IL-10 Interleukin-10, PGC-1 Peroxisome proliferator-activated receptor-gamma coactivator, SIRT-1 Sirtuin-1, STAT-3 Signal transducer and activator of transcription 3
Biological basis of cardioprotection in cardiotoxicity induced by other cancer drugs
Cardiotoxicity complicates the clinical use of many anticancer drugs other than anthracyclines [52, 138]. There are few preclinical studies exploring the cardioprotective role of SGLT2i against these drugs.
Trastuzumab is an anti-Human Epidermal Growth Factor Receptor 2 (HER2) antibody, primarily used in the treatment of HER2 overexpressing breast cancers (approximately 15–20% of all diagnoses). Anti-HER2 agents exert cardiotoxicity by a variety of mechanisms that include, among others, DNA-damage and ferroptosis [80, 131]. In vitro and in vivo studies showed that empagliflozin mitigates DNA damage and ferroptosis induced by trastuzumab [94]. In other preclinical studies, empagliflozin was able to ameliorate cardiomyocyte autophagy induced by sunitinib, an angiogenesis inhibitor. This reflect empagliflozin interactions with the AMP-activated Protein Kinase-Mammalian Target of Rapamycin (AMPK-mTOR) signaling pathway but other mechanisms of protection might well involve a reversal of sunitinib-induced microvascular injury and the consequent increase of coronary flow [63, 115].
Interesting data emerging from murine models and clinical studies suggest the potential protective effect of SGLT2i in mitigating cardiovascular toxicity caused by anti-angiogenic therapies. Luseogliflozin showed to upregulate the expression of vascular endothelial growth factor (VEGF)-α in the kidney after ischemia–reperfusion injury in non-diabetic mice [165]; at the same time, Nikolaou et al. demonstrated the protective role of chronic administration of empagliflozin in the myocardial ischemia–reperfusion injury in diabetic mice, sustained by the upregulation of VEGF pathway [101].
Antimetabolite drugs, such as 5-Fluorouracil (5-FU) and Capecitabine, are also associated with serious cardiovascular toxicities, such as coronary spasm, myocardial infarction, arrhythmias and HF [129]. Evidence regarding antimetabolites cardiotoxicity may reveal a cardioprotective role exerted by SGLT2i. In a preclinical study, the administration of empagliflozin in mice treated with 5-FU ameliorated the cardiotoxic effects by different mechanisms including vasodilating effect, antioxidant and anti-inflammatory properties and the downregulation of tumor necrosis factor alpha (TNFα)/ toll like receptors (TLR)/ nuclear factor-κB (NF-κB) pathway [113].
Another example of how SGLT2i can counteract or prevent cardiovascular toxicity from non-anthracycline drugs involves studies on carfilzomib (CFZ), a proteasome inhibitor, currently approved for relapsed/refractory multiple myeloma. Clinical employment of CFZ is limited by cardiovascular toxicity, mainly due to endothelial dysfunction. Canagliflozin abrogated the apoptotic effects of CFZ in cultured endothelial cells, and again this occurred by an AMPK-dependent mechanism [26].
A detailed analysis should be reserved to immune checkpoints inhibitors (ICIs), currently approved for many solid tumors treatment [4]. Main compounds used in clinical practice are reported in Table 1 [4, 9, 35, 49, 59, 110, 150, 168]. ICIs, inducing a non-specific autoimmune response by activating T cells, may lead to immune-related adverse events (irAEs) occurrence [109]. It has been observed that around 60–80% of patients who undergo ICIs treatment may experience irAEs, with 20% of them suffering from high-grade irAEs [65]. As well, among cancer patients who received a combination scheme (e.g. ipilimumab + nivolumab), more than 90% had at least one irAE, and nearly half of them had severe irAEs [67]. These side effects obviously do not spare the heart. However, ICIs-induced cardiotoxicity is a relatively rare complication and frequently manifests with myocarditis. Recent studies have found that SGLT2i may offer a promising approach to reduce all-cause mortality in cancer patients receiving ICIs therapy. Perelman et al. retrospectively analyzed 119 patients treated with ICIs; of them, 24 (20%) had SGLT2i on board. SGLT2i emerged as an independent predictor for lower all-cause mortality [108]. Noteworthy, another important finding was that the baseline hematocrit (HCT) was higher in the SGLT2i group (37 ± 6% vs. 33 ± 6%, p = 0.003) underling the active role on hematopoiesis played by these medications in cancer patients [38]. The complex mechanisms through which SGLT2i may oppose ICIs cardiotoxic effects are still unknown and translational research is needed.
All these findings identify SGLT2i as potential cardioprotective agents against a wide spectrum of antitumor drugs. Needless to say, further preclinical and clinical studies are much needed to support this hypothesis. However, the translation of data emerged from preclinical studies to clinical practice is limited by the lack of models able to mimic the clinical scenario of cancer patients, often affected by concomitant comorbidities and cardiovascular disease. Indeed, in most studies in vivo models were represented by healthy animals, complicating the transposition of results to elderly, comorbid cancer patients [84, 122]. While assessing the effectiveness of cardioprotective strategies, it is essential to consider the impact of cardiovascular risk factors, such as diabetes or arterial hypertension, and previous cardiovascular events, i.e. acute and chronic coronary syndromes, which frequently affect patients with active cancer or cancer survivors years after drug exposure [171]. Appropriate in vivo models able to integrate the complexity of cancer patients are needed in order to better understand the pathogenesis of cardiovascular toxicity, the relationship between fn cancer and cardiovascular diseases and potential therapeutic options [7, 18].
SGLT2 inhibitors in cardio-oncology: clinical experience
The promising results of preclinical studies paved the way to the evaluation of potential cardioprotective effects of SGLT2i in patients undergoing anthracycline-based chemotherapy (Table 2). A retrospective study recently included 3033 patients with diabetes mellitus and treated with anthracyclines for solid and hematologic malignancies. After matching subjects for age, sex, and anthracycline starting date, the authors could identify a case population of 32 patients who received SGLT2i during chemotherapy and a control population of 96 subjects who did not. The primary outcome was a composite of HF incidence, HF hospitalization, clinically significant arrhythmias, or a > 10% absolute decline in LVEF to a final value < 53%. During a median follow-up period of 1.5 years, when compared to controls, case patients experienced a lower incidence of the composite primary outcome, as well as reduced overall mortality and incidence of a composite of sepsis and neutropenic fever (16% vs 40%; p = 0.013). A non-significant higher incidence of genital infections was observed. The lower incidence of cardiac events was principally driven by a reduction in HF hospitalizations and > 10% decline LVEF to < 53% during chemotherapy [46].
In the same way, Chiang et al. conducted a retrospective propensity score-matched cohort study, involving adult patients with type 2 diabetes mellitus diagnosed with cancer (95% with solid tumors and 5% with haematologic malignancies). From a total cohort of 8640 patients, 878 patients received an SGLT2i [primarily empagliflozin (49%), followed by dapagliflozin (38%) and canagliflozin (14%)] while 7556 did not and served as controls. During a median follow-up of almost 20 months, SGLT2i patients showed a three-fold lower rate of hospitalizations for incident HF compared to controls (2.92 vs 8.95 per 1000 patient-years, p = 0.018). In Cox regression and competing regression analyses, SGLT2i were associated with a 72% reduction in the risk of hospitalization for HF. Overall, SGLT2i was associated with a higher survival (85.3% vs 63.0% at 2 years, p < 0.001), with the risk of serious adverse events (i.e. acute kidney injury and diabetic ketoacidosis) similar in the two groups [24]. Of interest, the use of SGLT2i was associated with a decreased risk of urosepsis, sepsis and hypoglycemia.
Other investigators evaluated a cohort of 933 patients without HF history who were receiving medications for diabetes and underwent anthracycline-based chemotherapy. Of these patients, 99 received an SGLT2i while 834 did not and served as controls. During a median follow-up of 1.6 years, in the SGLT2i population there was a significant lower incidence of HF hospitalizations, while no significant difference in incident HF diagnosis, cardiovascular disease diagnosis or mortality could be detected [1]. Finally, Avula et al. conducted a retrospective investigation on the role of SGLT2i in diabetic patients exposed to antineoplastic agents and developing a decline in left ventricular function. In a cohort of 1280 adult patients without a history of ischemic heart disease but with chemotherapy-induced cardiac dysfunction, patients treated with SGLT2i on top of therapy showed reductions in acute HF exacerbation, all-cause mortality, all-cause hospitalizations, atrial fibrillation/flutter, acute kidney injury and need for renal replacement therapy compared to controls who did not receive SGLT2i [11].
Even if limited by observational design and unmeasured confounders between the populations examined, these studies formed the rationale for randomized controlled trials. The EMPACT (Empagliflozin in the Prevention of Cardiotoxicity in Cancer Patients Undergoing Chemotherapy Based on Anthracyclines; NCT05271162) trial is a multicenter phase III study aiming to assess whether prophylactic empagliflozin may prevent LV dysfunction in patients receiving high-dose anthracycline therapy.
Cardiotoxicity may not only manifest as cardiac dysfunction, but also with arrhythmic events, such as atrial fibrillation (AF) and ventricular tachycardia, generally developing acutely after anticancer treatment administration. In particular, AF is frequent in oncological patients, with an estimated prevalence of 9.77% and an age-related relative risk ratio in patients with cancer compared with no cancer of 10.45 [12]. Pathogenetic factors coming at play are numerous, even though generally involving patients’ related factors, anti-neoplastic therapy performed and/or direct compression/invasion exerted by cancer itself [23, 34, 48, 92, 112]. There is growing evidence about the potential protective role of SGLT2i in arrhythmias and AF [77]. Potential biological mechanisms implicated in antiarrhythmic properties probably involve regulation of Ca2+ and Na+ homeostasis, as well as the anti-fibrotic and anti-inflammatory effects exerted on myocardial cells [43]. In clinical practice, dapagliflozin showed to influence AF or atrial flutter events in the DECLARE-TIMI 58 population, reducing their occurrence by 19%; this effect was consistent regardless of the patient’s previous history of AF, atherosclerotic disease, blood pressure or history of HF [163]. Likewise, Abu-Qaoud et al. studied the effects of SGLT2i in patients with type 2 diabetes previously diagnosed with AF and treated with catheter ablation, demonstrating that the use of these drugs was associated with a lower risk of arrhythmia recurrence and consequently a reduced need for cardioversion, antiarrhythmic therapy or new AF ablation [3]. In the light of this new evidence, SGLT2i may become an important tool in the management of rhythm-related disorders in the oncological population, specifically in patients treated with cancer therapies associated with arrhythmic events.
SGLT2 inhibitors and bone marrow function: implications for cancer patients
In addition to the aforementioned cardioprotective effects, SGLT2i seem to exert unique and complex effects on hematopoiesis and bone marrow (BM) function; accordingly, BM cells represent the extrarenal compartment with the highest level of expression of SGLT2, followed by skeletal muscle and myocardial cells [5].
SGLT2 inhibitors and modulation of erythropoiesis.
The most evident BM effect of SGLT2i is modulation of erythropoiesis. SGLT2i increased hemoglobin count and hematocrit in various randomized controlled trials (summarized in Table 3 along with the suggested mechanisms of action of SGLT2i). The most notable trial to provide such information was the EMPA-REG OUTCOME trial, which recruited 7028 diabetic patients and reported higher hematocrit values in the empagliflozin groups than in the placebo group [170]. While originally ascribed to natriuresis and plasma volume contraction, the increase in hematocrit was later related to an enhanced production of erythropoietin [30, 37, 88]. In fact, an increase in erythropoietin occurred early upon treatment initiation and subsided a new set point for the equilibrium between erythropoietin and haemoglobin levels [153]. SGLT2i improve erythropoietin expression/secretion via at least three distinct mechanisms: modulation of tubulointerstitial hypoxia and hypoxia-induced factor (HIF)‐α expression, mitigation of inflammation-induced functional iron deficiency, and metabolic reprogramming (the latter manifested at a cellular level by an increased production of ketone bodies and a state of starvation mimicry) [104].
The action of SGLT2i to block Na+ reabsorption in the proximal renal tubule leads to increased delivery of sodium to more distal portions of the nephron, where counterregulatory mechanisms of Na+ absorption are recruited to limit the magnitude of natriuresis. It has been speculated that SGLT2i might in this way increase oxygen consumption and predispose to tissue hypoxia in the S3 segment of the nephron, possibly in close proximity to the specialized interstitial fibroblast-like cells belonging to the deep cortex, which could be stimulated to produce erythropoietin [156]. On the other hand, inhibition of glucose reabsorption would alleviate metabolic demands in the proximal tubules and reduce oxygen consumption, thus improving oxygenation in the outer renal cortex and allowing dysfunctional fibroblasts to revert to a phenotype that is able to synthetizes erythropoietin [124].
In addition to the aforesaid considerations, the disease states that pose an indication to SGLT2i, such as HF and kidney impairment, are associated with the development of a typical anemia of chronic disease. This is caused by an inflammatory state characterized by increased levels of two major iron regulatory proteins, hepcidin and ferritin, and by a state of functional iron deficiency [147]. Hepcidin, produced by the liver, blocks the absorption of iron from the duodenum and its release from the reticuloendothelial system. SGLT2i reduced serum hepcidin and ferritin levels in both type 2 diabetes [45] and chronic HF [24, 104], in part by stimulating erythropoiesis and in part by a direct anti-inflammatory effect [103]. Regardless of the ultimate mechanism, decreases in hepcidin and ferritin would lead to an increased release of iron from macrophages and intracellular storages, respectively, fueling red blood cells production. A key role seems to be played also by ketone bodies. Acetoacetic acid, β‐hydroxybutyrate, and acetone are in fact hyperproduced in patients during chronic SGLT2i therapy and experimental evidence suggests that β‐hydroxybutyrate infusion causes a concomitant 30% increase in erythropoietin levels and bone marrow glucose uptake in healthy volunteers [71]. If and how precisely an SGLTi-driven upregulation of ketone bodies would contribute to increasing erythropoiesis will nonetheless require further investigations. Finally, SGLT2i up-regulate sirtuin (SIRT)1 [132], which can then activate HIF-2α directly [28] or through heme oxygenase-1 and can enhance transcription of the erythropoietin gene [139]. Thus, SGLT2i may well augment erythropoietin production in kidney and liver cells also in an oxygen-independent manner.
SGLT2i has also been linked with enhanced development and BM release of pro-angiogenic progenitor cells. In particular, dapagliflozin increased BM resident mature leukocytes and circulating white blood cells in both diabetic and non-diabetic mice, and improved diabetes-associated defects in hematopoietic stem cell mobilization by granulocyte colony-stimulating factor, eventually promoting the migration of BM cells to the sites of vascular injury [5]. In humans, empagliflozin increased circulating pro-angiogenic CD133+ progenitor cells and increased M2 polarized monocytes with anti-inflammatory properties [51]. As anthracycline cardiomyopathy is at least in part due to impaired mobilization and regenerative functions of endothelial cell progenitors [57], SGLT2i could be of potential value to restore the recruitment of endothelial cell progenitors to sites of injury. As a last observation, the aforementioned work by Gongora et al. [46] showed that SGLT2i reduced a composite outcome of sepsis and neutropenic fever, possibly suggesting that these drugs may also exert some effects on the defense mechanisms of cancer patients treated with chemotherapy. SGLT2i effects on lymphocyte and granulocyte functions could be an interesting field for future research.
Implications of SGLT2 inhibitors use in clinical practice.
Anemia is a frequent finding in patients diagnosed with solid cancers and lymphomas or acute leukemias, and is further worsened by chemotherapy treatments. Unsurprisingly, anemia is an important adverse prognostic factor for outcomes in lymphoma patients, worsening the overall and progression free survival independent of bone marrow involvement [98]. The pathogenesis of cancer-induced anaemia (CIA) is complex and multifactorial [2]. Cancer-associated chronic inflammatory is an important contributor, leading to a condition of functional iron deficiency; for example, patients with diffuse large B cells lymphoma are characterized by increased circulating levels of IL-6, hepcidin and ferritin, along with a defective erythropoietin production [136]. Current management of CIA relies on iron replacement therapy, transfusions and erythropoiesis stimulating agents (ESA) [16]. However, CIA correction remains an unmet clinical need, considering that only 1/3 of patients respond to ESA. Moreover, ESA is associated with an increased risk of thromboembolic events and concerns around an accelerated tumor progression have been raised. Transfusions of red blood cells, although in some cases unavoidable, seem to worsen prognosis [2]. In this setting, the multifaceted activities of SGLT2i on reducing inflammation while also safely upregulating erythropoiesis hold promise for a better management of CIA. Nevertheless, it is worth noting that SGLT2i-induced erythrocytosis seems not to be accompanied by an increased risk of thrombosis, which makes premature SGLT2i discontinuation or haematocrit lowering strategies (such as implementation of phlebotomy program) unnecessary precautions [41, 42].
Figure 4 summarizes the role of SGLT2i in the hematologic patient.

The hematological effects of SGLT2 inhibitors. On the right, the effects on the red blood cell in the context of chemotherapy-induced anemia (CIA): in particular, inhibition of SGL2 promotes the increase in hematocrit by stimulating the release of erythropoietin and activating SIRT1; it induces the modulation of interstitial tubule hypoxia by promoting the reduction of ferritin and hepcidin. SGLT2 inhibitors also reduce Ferroptosis favoring a greater amount of circulating iron from macrophages. On white blood cells, in the mouse models, SGLT2 inhibitors promote the increase of CD34+ cells in peripheral blood and the migration of neutrophils at the level of vascular injuries. Furthermore, they favor the polarization of macrophages into the M2 phenotype. The presence of SGLT2 at the level of lymphoblasts and myeloblasts has not yet been fully elucidated. EPO Erythropoietin, HIF hypoxia induced factor, IL6 interleukin 6, SIRT1 Sirtuin-1
Proposed SGLT2 inhibitors effects on solid tumors and hematological malignancies
There are several in vitro studies suggesting that SGLT2i exhibit anti-proliferative activity against some types of solid tumors, including breast, lung, prostate, colon and pancreas. This may help to design strategies that combine SGLT2i with other cancer drugs [60, 69, 70, 169].
The metabolic reprogramming of cancer cells involves changes in the uptake of various substrates and changes in their metabolic pathways. The most striking example of altered metabolism is the Warburg Effect [78], which consists of increased glucose uptake and its conversion to lactate via anaerobic glycolysis. One of the proposed mechanisms through which SGLT2i may limit tumor growth therefore consists in decreasing glucose availability to cancer cells through inhibition of sodium glucose cotransporter, which in fact is overexpressed in pancreatic, lung and prostate adenocarcinomas and in high grade glioblastomas [127, 148].
In this context, emerging anticancer effect of SGLT2i may also be obtained by reducing the activity of Hexokinase II (HK II), an enzyme that has been found to play a significant role in cancer development. It is overexpressed in many types of cancer cells and has been shown to promote tumor growth and survival by increasing the rate of glucose uptake and metabolism. This enzyme is known to interact with mitochondria, serving as facilitator and gatekeeper of malignancy [87]. HK II is also able to suppress the death of cancer cells, increasing their metastatic potential. For these reasons, targeting this key enzyme is currently being investigated as novel cancer therapies [40]. Recent studies have shown that SGLT2i may have anticancer effects by reducing the activity of HK II. As well, by reducing glucose uptake and metabolism in cancer cells, SGLT2i may attenuate tumor growth and improve the effectiveness of cancer treatments [29, 142, 152]. While both Hexokinase II and SGLT2i have been extensively studied in the context of cancer, further research is needed to fully understand their complex roles in the disease and to develop effective treatments that target these pathways.
Programmed cell death-ligand 1 (PD-L1) function on cancer cells appears to be critical in immune escape and tumor development. SGLT2i promotes PD-L1 recycling, allowing the ubiquitination and proteasome-mediated degradation [27]. As well, a number of emerging evidences suggests that SGLT2i may have direct anticancer effects, going beyond metabolic properties. A recent study demonstrated that SGLT2 is overexpressed in osteosarcoma cells and its inhibition can promote treatment efficacy by inducing T cell infiltration [149]. In the same direction, other recent reports found similar efficacy of SGLT2i in the context of liver [50] and cervical carcinoma [151].
Effects of SGLT2i in solid tumors are currently investigated in small clinical trials. The first trial enrolled 15 participants to assess the tolerability and efficacy of dapagliflozin on top of standard chemotherapy (Gemcitabine and nab-Paclitaxel) in patients with metastatic pancreatic cancer. The drug was well tolerated with favorable changes in tumor response and plasma biomarkers [105]. An ongoing phase 1b/2 study (NCT04073680) plans to recruit 60 participants with advanced solid tumors (breast, endometrial, lung, colorectal, and head and neck cancers) treated in combination with serabelisib (a PI3K inhibitor) and canagliflozin. The study rationale is that hampering the glucose/insulin feedback pathway would enhance the efficacy of PI3K inhibition. A phase I trial (NCT04887935) involving 24 participants is planning to assess the tolerability/efficacy of neoadjuvant SGLT2i before radical prostatectomy in high-risk localized prostate cancer. Finally, a phase 1 study (NCT05521984) will probe dapagliflozin in combination with carmustine for treatment of pediatric brain tumors.
Metastasis is a significant challenge in cancer treatment, often resulting in poor prognosis. Recent research has suggested that SGLT2i may play a role in the prevention of metastasis through different mechanisms: reducing the expression of certain molecules that are involved in the adhesion and migration of cancer cells and reducing the activity of signaling pathways involved in the growth and survival of cancer cells. On this line, a recent study from Rogava and colleagues explored new insights into the mechanisms behind liver metastasis in cancer [118]. In particular, they focused on Pip4k2c (phosphatidylinositol-5-phosphate 4-kinase), an actionable target of SGLT2i, and found that the loss of Pip4k2c leads to the activation of the PI3K-AKT pathway, which plays a crucial role in liver metastasis.
Less is known about SGLT2i as therapeutic agents for hematologic malignancies. A meta-analysis of diabetic patients treated SGLT2i or Dipeptidyl Peptidase (DPP)IV inhibitors found a lower risk of hematological malignancies in the SGLT2i arm, which suggests a possible protective role of these agents [119]. In preclinical studies, SGLT2 was shown to be expressed in the human tonsil [22], in regulatory T cells, memory CD4 and CD8 T cells and NK cells [140]. Moreover, SGLT2 was remarkably expressed in leukemic cells from patients with adult T-cell leukemia (ATL) and SGLT2i suppressed the proliferation of two ATL cell lines and of leukemic cells in peripheral blood from patients with ATL. The antiproliferative effect of SGLT2i in these cells was due to reduced glucose uptake and consequent reduced intracellular levels of ATP and NADPH [99].
On the basis of these preclinical studies, more evidence of SGLT2i in the treatment of hematological malignancies, including the broad spectrum of lymphoproliferative disorders, is highly warranted.
Conclusions and future directions
We reviewed the promise that SGLT2i hold as cardiac protectants, potential antiproliferative agents and stimulants of erythropoiesis and immune defense in the cancer patient. Overall, this represents the conceptual pillar to foster clinical research and use of SGLT2i beyond currently recommended indications in HF and diabetes. The need for cardioprotective agents is in fact a notable challenge in contemporary Cardio-Oncology, particularly in view of the avalanche of new drugs and therapies heavily treated patients with relapsed or refractory cancer might be considered candidates for. At the same time, the need for improving quality of life and patient reported outcomes in oncologic settings encourage studies of SGLT2i in cancer pathophysiology, immune system regulation and infection control.
Abbreviations
- AMPK:
-
Adenosine monophosphate-activated protein kinase
- AF:
-
Atrial fibrillation
- AMPK-mTOR:
-
Adenosine Monophosphate-activated Protein Kinase-Mammalian Target of Rapamycin
- ARNI:
-
Angiotensin II receptor blockers-neprilysin inhibitors
- ATL:
-
Adult T-cell leukemia
- β-OHB:
-
Beta hydroxybutyrate
- Ca2+:
-
Calcium ion
- CIA:
-
Cancer-induced anaemia
- CFZ:
-
Carfilzomib
- CKD:
-
Chronic kidney disease
- ENaC:
-
Epithelial sodium channel
- EPO:
-
Erythropoietin
- ESC:
-
European society of cardiology
- HR:
-
Hazard ratio
- HF:
-
Heart failure
- HFmrEF:
-
Heart failure with mildly reduced ejection fraction
- HFpEF:
-
Heart failure with preserved ejection fraction
- HFrEF:
-
Heart failure with reduced ejection fraction
- HNF4:
-
Hepatocyte nuclear factor 4
- hs-CRP:
-
Hypersensitive-C-reactive-protein
- HIF‐α:
-
Hypoxia-inducible factor alpha
- HER2:
-
Human Epidermal Growth Factor Receptor 2
- HK II:
-
Hexokinase II
- IFN:
-
Interferon
- Fe2+:
-
Iron ion
- ICIs:
-
Immune checkpoint inhibitors
- IL:
-
Interleukin
- mmHg:
-
Millimeters of mercury
- MyD88:
-
Myeloid differentiation primary response 88
- NADPH:
-
Nicotinamide Adenine Dinucleotide Phosphate
- NLRP3:
-
NOD-like receptor pyrin domain containing protein 3
- PGC-1:
-
Peroxisome proliferator-activated receptor-gamma coactivator
- PI3K:
-
Phosphoinositide 3-kinase
- Pip4k2c:
-
Phosphatidylinositol-5-phosphate 4-kinase
- PD-L1:
-
Programmed cell death-ligand 1
- ROS:
-
Reactive oxygen species
- RBC:
-
Red blood cells
- SGK1:
-
Serum- and Glucocorticoid-Regulated Kinase 1
- STAT3:
-
Signal transducer and activator of transcription 3
- SIRT1:
-
Sirtuin 1
- Na2+:
-
Sodium ion
- SGLT2:
-
Sodium–glucose cotransporter 2
- TNF:
-
Tumor necrosis factor
- TLR:
-
Toll-like receptor
- TGF:
-
Transforming growth factor
References
Abdel-Qadir H, Carrasco R, Austin PC, Chen Y, Zhou L, Fang J, Su HMH, Lega IC, Kaul P, Neilan TG, Thavendiranathan P (2023) The association of sodium–glucose cotransporter 2 inhibitors with cardiovascular outcomes in anthracycline-treated patients with cancer. JACC CardioOncol 5:318–328. https://doi.org/10.1016/J.JACCAO.2023.03.011
Abdel-Razeq H, Hashem H (2020) Recent update in the pathogenesis and treatment of chemotherapy and cancer induced anemia. Crit Rev Oncol Hematol. https://doi.org/10.1016/J.CRITREVONC.2019.102837
Abu-Qaoud MR, Kumar A, Tarun T, Abraham S, Ahmad J, Khadke S, Husami R, Kulbak G, Sahoo S, Januzzi JL, Neilan TG, Baron SJ, Martin D, Nohria A, Reynolds MR, Kosiborod M, Dani SS, Ganatra S (2023) Impact of SGLT2 inhibitors on AF recurrence after catheter ablation in patients with type 2 diabetes. JACC Clin Electrophysiol 9:2109–2118. https://doi.org/10.1016/J.JACEP.2023.06.008
Akinleye A, Rasool Z (2019) Immune checkpoint inhibitors of PD-L1 as cancer therapeutics. J Hematol Oncol. https://doi.org/10.1186/S13045-019-0779-5
Albiero M, Tedesco S, Amendolagine FI, D’anna M, Migliozzi L, Zuccolotto G, Rosato A, Cappellari R, Avogaro A, Fadini GP (2021) Inhibition of SGLT2 rescues bone marrow cell traffic for vascular repair: role of glucose control and ketogenesis. Diabetes 70:1767–1779. https://doi.org/10.2337/DB20-1045
Anker SD, Butler J, Filippatos G, Ferreira JP, Bocchi E, Böhm M, Brunner-La Rocca H-P, Choi D-J, Chopra V, Chuquiure-Valenzuela E, Giannetti N, Gomez-Mesa JE, Janssens S, Januzzi JL, Gonzalez-Juanatey JR, Merkely B, Nicholls SJ, Perrone SV, Piña IL, Ponikowski P, Senni M, Sim D, Spinar J, Squire I, Taddei S, Tsutsui H, Verma S, Vinereanu D, Zhang J, Carson P, Lam CSP, Marx N, Zeller C, Sattar N, Jamal W, Schnaidt S, Schnee JM, Brueckmann M, Pocock SJ, Zannad F, Packer M (2021) Empagliflozin in heart failure with a preserved ejection fraction. N Engl J Med 385:1451–1461. https://doi.org/10.1056/NEJMOA2107038/SUPPL_FILE/NEJMOA2107038_DATA-SHARING.PDF
Asnani A, Moslehi JJ, Adhikari BB, Baik AH, Beyer AM, De Boer RA, Ghigo A, Grumbach IM, Jain S, Zhu H (2021) Preclinical models of cancer therapy-associated cardiovascular toxicity: a scientific statement from the American Heart Association. Circ Res 129:E21–E34. https://doi.org/10.1161/RES.0000000000000473
Association AD (2021) 9. Pharmacologic approaches to glycemic treatment: standards of medical care in diabetes—2021. Diabetes Care 44:S111–S124. https://doi.org/10.2337/DC21-S009
Atkins MB, Tannir NM (2018) Current and emerging therapies for first-line treatment of metastatic clear cell renal cell carcinoma. Cancer Treat Rev 70:127–137. https://doi.org/10.1016/J.CTRV.2018.07.009
Aubert G, Martin OJ, Horton JL, Lai L, Vega RB, Leone TC, Koves T, Gardell SJ, Krüger M, Hoppel CL, Lewandowski ED, Crawford PA, Muoio DM, Kelly DP (2016) The Failing Heart Relies on Ketone Bodies as a Fuel. Circulation 133:698–705. https://doi.org/10.1161/CIRCULATIONAHA.115.017355
Avula V, Sharma G, Kosiborod MN, Vaduganathan M, Neilan TG, Lopez T, Dent S, Baldassarre L, Scherrer-Crosbie M, Barac A, Liu J, Deswal A, Khadke S, Yang EH, Ky B, Lenihan D, Nohria A, Dani SS, Ganatra S (2023) SGLT2 Inhibitor Use and Risk of Clinical Events in Patients With Cancer Therapy-Related Cardiac Dysfunction. JACC Heart Fail. https://doi.org/10.1016/J.JCHF.2023.08.026
Ay C, Grilz E, Nopp S, Moik F, Königsbrügge O, Klimek P, Thurner S, Posch F, Pabinger I (2022) Atrial fibrillation and cancer: prevalence and relative risk from a nationwide study. Res Pract Thromb Haemost 7. https://doi.org/10.1016/J.RPTH.2022.100026
Bernard Z, Wanner C, Lachin JM, Fitchett D, Bluhmki E, Hantel S, Mattheus M, Devins T, Johansen OE, Woerle HJ, Broed UC, Inzucchi SE (2015) Empagliflozin, cardiovascular outcomes, and mortality in type 2 diabetes. N Engl J Med 373:17–18. https://doi.org/10.1056/NEJMOA1504720
Baartscheer A, Schumacher CA, Wüst RCI, Fiolet JWT, Stienen GJM, Coronel R, Zuurbier CJ (2017) Empagliflozin decreases myocardial cytoplasmic Na+ through inhibition of the cardiac Na+/H+ exchanger in rats and rabbits. Diabetologia 60:568–573. https://doi.org/10.1007/S00125-016-4134-X/FIGURES/2
Bhagat A, Shrestha P, Kleinerman ES (2022) The Innate Immune System in Cardiovascular Diseases and Its Role in Doxorubicin-Induced Cardiotoxicity. Int J Mol Sci 23. https://doi.org/10.3390/IJMS232314649
Bohlius J, Bohlke K, Castelli R, Djulbegovic B, Lustberg MB, Martino M, Mountzios G, Peswani N, Porter L, Tanaka TN, Trifiro G, Yang H, Lazo-Langner A (2019) Management of cancer-associated anemia with erythropoiesis-stimulating agents: ASCO/ASH clinical practice guideline update. Blood Adv 3:1197–1210. https://doi.org/10.1182/BLOODADVANCES.2018030387
Bonnet F, Scheen AJ (2018) Effects of SGLT2 inhibitors on systemic and tissue low-grade inflammation: The potential contribution to diabetes complications and cardiovascular disease. Diabetes Metab 44:457–464. https://doi.org/10.1016/J.DIABET.2018.09.005
Bouleftour W, Mery B, Rowinski E, Rivier C, Daguenet E, Magne N (2021) Cardio-Oncology Preclinical Models: A Comprehensive Review. Anticancer Res 41:5355–5364. https://doi.org/10.21873/ANTICANRES.15348
Camilli M, Lombardi M, Chiabrando JG, Del Buono MG, Montone RA, Biondi-Zoccai G, Crea F, Minotti G (2021) Efficacy of sodium–glucose cotransporter-2 inhibitors in heart failure patients treated with dual angiotensin receptor blocker-neprilysin inhibitor: an updated meta-analysis. Eur Heart J Cardiovasc Pharmacother 7:E74–E76. https://doi.org/10.1093/EHJCVP/PVAB034
Camilli M, Lombardi M, Chiabrando JG, Zito A, Del Buono MG, Vergallo R, Aspromonte N, Lombardo A, Montone RA, Niccoli G, Biondi-Zoccai G, Crea F, Minotti G (2022) Sodium–glucose Cotransporter Inhibitors Reduce Mortality and Morbidity in Patients with Heart Failure: Evidence from a Meta-Analysis of Randomized Trials. Am J Ther 29:E199–E204. https://doi.org/10.1097/MJT.0000000000001452
Cavender MA, Norhammar A, Birkeland KI, Jørgensen ME, Wilding JP, Khunti K, Fu AZ, Bodegård J, Blak BT, Wittbrodt ET, Thuresson M, Fenici P, Hammar N, Kosiborod M, Holl RW, Chen H, Scheerer MF, Surmont F, Nahrebne K, Gulseth HL, Carstensen B, Bollow E, García Rodríguez LA, Cea Soriano L, Cantero OF, Thiel E, Murphy B (2018) SGLT-2 Inhibitors and Cardiovascular Risk: An Analysis of CVD-REAL. J Am Coll Cardiol 71:2497–2506. https://doi.org/10.1016/J.JACC.2018.01.085
Chen J, Williams S, Ho S, Loraine H, Hagan D, Whaley JM, Feder JN (2010) Quantitative PCR tissue expression profiling of the human SGLT2 gene and related family members. Diabetes Therapy 1:57–92. https://doi.org/10.1007/S13300-010-0006-4/FIGURES/28
Cheng WL, Kao YH, Chen SA, Chen YJ (2016) Pathophysiology of cancer therapy-provoked atrial fibrillation. Int J Cardiol 219:186–194. https://doi.org/10.1016/J.IJCARD.2016.06.009
Chiang CH, Chiang CH, Chiang CH, Ma KSK, Peng CY, Hsia YP, Horng CS, Chen CY, Chang YC, See XY, Chen YJ, Wang SS, Suero-Abreu GA, Peterson LR, Thavendiranathan P, Armand P, Peng CM, Shiah HS, Neilan TG (2023) Impact of sodium–glucose cotransporter-2 inhibitors on heart failure and mortality in patients with cancer. Heart 109:470–477. https://doi.org/10.1136/HEARTJNL-2022-321545
Cowie MR, Fisher M (2020) SGLT2 inhibitors: mechanisms of cardiovascular benefit beyond glycaemic control. Nat Rev Cardiol 17:761–772. https://doi.org/10.1038/S41569-020-0406-8
Dabour MS, Abdelgawad IY, Grant MKO, El-Sawaf ES, Zordoky BN (2023) Canagliflozin mitigates carfilzomib-induced endothelial apoptosis via an AMPK-dependent pathway. Biomed Pharmacother 164:114907. https://doi.org/10.1016/J.BIOPHA.2023.114907
Ding L, Chen X, Zhang W, Dai X, Guo H, Pan X, Xu Y, Feng J, Yuan M, Gao X, Wang J, Xu X, Li S, Wu H, Cao J, He Q, Yang B (2023) Canagliflozin primes antitumor immunity by triggering PD-L1 degradation in endocytic recycling. J Clin Invest. https://doi.org/10.1172/JCI154754
Dioum EM, Chen R, Alexander MS, Zhang Q, Hogg RT, Gerard RD, Garcia JA (2009) Regulation of Hypoxia-Inducible Factor 2α Signaling by the Stress-Responsive Deacetylase Sirtuin 1. Science (1979) 324:1289–1293. https://doi.org/10.1126/SCIENCE.1169956/SUPPL_FILE/DIOUM_SOM.PDF
Dutka M, Bobiński R, Francuz T, Garczorz W, Zimmer K, Ilczak T, Ćwiertnia M, Hajduga MB (2022) SGLT-2 inhibitors in cancer treatment-mechanisms of action and emerging new perspectives. Cancers (Basel). https://doi.org/10.3390/CANCERS14235811
Ekanayake P, Mudaliar S (2023) Increase in hematocrit with SGLT-2 inhibitors—hemoconcentration from diuresis or increased erythropoiesis after amelioration of hypoxia? Diabetes Metab Syndr. https://doi.org/10.1016/J.DSX.2022.102702
Faggiano A, Gherbesi E, Cardinale D, Vicenzi M, Carugo S (2023) SGLT2-i prevent left ventricular dysfunction induced by anthracycline in mouse model: a systematic-review and meta-analysis. Vascul Pharmacol 150:107171. https://doi.org/10.1016/j.vph.2023.107171
Fang X, Ardehali H, Min J, Wang F (2023) The molecular and metabolic landscape of iron and ferroptosis in cardiovascular disease. Nat Rev Cardiol 20:7–23. https://doi.org/10.1038/S41569-022-00735-4
Farkouh ME, Verma S (2018) Prevention of heart failure with SGLT-2 inhibition: insights from CVD-real. J Am Coll Cardiol 71:2507–2510. https://doi.org/10.1016/J.JACC.2018.02.078
Farmakis D, Filippatos G (2021) Arrhythmias in cancer: rhythm is gonna get you! Eur J Heart Fail 23:154–156. https://doi.org/10.1002/EJHF.2079
Francisco LM, Sage PT, Sharpe AH (2010) The PD-1 pathway in tolerance and autoimmunity. Immunol Rev 236:219–242. https://doi.org/10.1111/J.1600-065X.2010.00923.X
Friedmann Angeli JP, Schneider M, Proneth B, Tyurina YY, Tyurin VA, Hammond VJ, Herbach N, Aichler M, Walch A, Eggenhofer E, Basavarajappa D, Rådmark O, Kobayashi S, Seibt T, Beck H, Neff F, Esposito I, Wanke R, Förster H, Yefremova O, Heinrichmeyer M, Bornkamm GW, Geissler EK, Thomas SB, Stockwell BR, Odonnell VB, Kagan VE, Schick JA, Conrad M (2014) Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nat Cell Biol 16(12):1180–1191. https://doi.org/10.1038/ncb3064
Fuchs Andersen C, Omar M, Glenthøj A, El Fassi D, Møller HJ, Lindholm Kurtzhals JA, Styrishave B, Kistorp C, Tuxen C, Poulsen MK, Faber J, Køber L, Gustafsson F, Møller JE, Schou M, Jensen J (2023) Effects of empagliflozin on erythropoiesis in heart failure: data from the Empire HF trial. Eur J Heart Fail 25:226–234. https://doi.org/10.1002/EJHF.2735
Fujiwara M, Shimizu M, Okano T, Maejima Y, Shimomura K (2023) Successful treatment of nivolumab and ipilimumab triggered type 1 diabetes by using sodium–glucose transporter 2 inhibitor: a case report and systematic review. Front Public Health. https://doi.org/10.3389/FPUBH.2023.1264056
Gambardella J, Santulli G, Fiordelisi A, Cerasuolo FA, Wang X, Prevete N, Sommella E, Avvisato R, Buonaiuto A, Altobelli GG, Rinaldi L, Chiuso F, Feliciello A, Dal Piaz F, Campiglia P, Ciccarelli M, Morisco C, Sadoshima J, Iaccarino G, Sorriento D (2023) Infiltrating macrophages amplify doxorubicin-induced cardiac damage: role of catecholamines. Cell Mol Life Sci 80:1–14. https://doi.org/10.1007/S00018-023-04922-5/FIGURES/6
Ganapathy-Kanniappan S, Kunjithapatham R, Geschwind JF (2013) Anticancer efficacy of the metabolic blocker 3-bromopyruvate: specific molecular targeting. Anticancer Res 33:13–20
Gangat N, Abdallah M, Szuber N, Saliba A, Alkhateeb H, Al-Kali A, Begna KH, Pardanani A, Tefferi A (2023) Sodium–glucose co-transporter-2 inhibitor use and JAK2 unmutated erythrocytosis in 100 consecutive cases. Am J Hematol. https://doi.org/10.1002/AJH.26933
Gangat N, Szuber N, Alkhateeb H, Al-Kali A, Pardanani A, Tefferi A (2021) JAK2 wild-type erythrocytosis associated with sodium–glucose cotransporter 2 inhibitor therapy. Blood 138:2886–2889. https://doi.org/10.1182/BLOOD.2021013996
Gao J, Xue G, Zhan G, Wang X, Li J, Yang X, Xia Y (2022) Benefits of SGLT2 inhibitors in arrhythmias. Front Cardiovasc Med. https://doi.org/10.3389/FCVM.2022.1011429
Gao M, Monian P, Quadri N, Ramasamy R, Jiang X (2015) Glutaminolysis and transferrin regulate ferroptosis. Mol Cell 59:298–308. https://doi.org/10.1016/j.molcel.2015.06.011
Ghanim H, Abuaysheh S, Hejna J, Green K, Batra M, Makdissi A, Chaudhuri A, Dandona P (2020) Dapagliflozin suppresses hepcidin and increases erythropoiesis. J Clin Endocrinol Metab 105:e1056–e1063. https://doi.org/10.1210/CLINEM/DGAA057
Gongora CA, Drobni ZD, Quinaglia Araujo Costa Silva T, Zafar A, Gong J, Zlotoff DA, Gilman HK, Hartmann SE, Sama S, Nikolaidou S, Suero-Abreu GA, Jacobsen E, Abramson JS, Hochberg E, Barnes J, Armand P, Thavendiranathan P, Nohria A, Neilan TG (2022) Sodium–glucose co-transporter-2 inhibitors and cardiac outcomes among patients treated with anthracyclines. JACC Heart Fail 10:559–567. https://doi.org/10.1016/J.JCHF.2022.03.006
Habibi J, Aroor AR, Sowers JR, Jia G, Hayden MR, Garro M, Barron B, Mayoux E, Rector RS, Whaley-Connell A, DeMarco VG (2017) Sodium glucose transporter 2 (SGLT2) inhibition with empagliflozin improves cardiac diastolic function in a female rodent model of diabetes. Cardiovasc Diabetol 16:1–15. https://doi.org/10.1186/S12933-016-0489-Z/FIGURES/7
Hajjar LA, Fonseca SMR, Machado TIV (2021) Atrial fibrillation and cancer. Front Cardiovasc Med. https://doi.org/10.3389/FCVM.2021.590768
Han Y, Liu D, Li L (2020) PD-1/PD-L1 pathway: current researches in cancer. Am J Cancer Res 10:727–742
Hendryx M, Dong Y, Ndeke JM, Luo J (2022) Sodium–glucose cotransporter 2 (SGLT2) inhibitor initiation and hepatocellular carcinoma prognosis. PLoS ONE. https://doi.org/10.1371/JOURNAL.PONE.0274519
Hess DA, Terenzi DC, Trac JZ, Quan A, Mason T, Al-Omran M, Bhatt DL, Dhingra N, Rotstein OD, Leiter LA, Zinman B, Sabongui S, Yan AT, Teoh H, Mazer CD, Connelly KA, Verma S (2019) SGLT2 inhibition with empagliflozin increases circulating provascular progenitor cells in people with type 2 diabetes mellitus. Cell Metab 30:609–613. https://doi.org/10.1016/J.CMET.2019.08.015
Heusch G (2023) Cardioprotection in cardio-oncology: a case for concern? Cardiovasc Res 119:e144–e145. https://doi.org/10.1093/cvr/cvad111
Heusch G (2024) Myocardial ischemia/reperfusion: translational pathophysiology of ischemic heart disease. Med (N Y) 5:10–31. https://doi.org/10.1016/j.medj.2023.12.007
Heusch G, Andreadou I, Bell R, Bertero E, Botker H-E, Davidson SM, Downey J, Eaton P, Ferdinandy P, Gersh BJ, Giacca M, Hausenloy DJ, Ibanez B, Krieg T, Maack C, Schulz R, Sellke F, Shah AM, Thiele H, Yellon DM, Di Lisa F (2023) Health position paper and redox perspectives on reactive oxygen species as signals and targets of cardioprotection. Redox Biol 67:102894. https://doi.org/10.1016/j.redox.2023.102894
Hofmann U, Knorr S, Vogel B, Weirather J, Frey A, Ertl G, Frantz S (2014) Interleukin-13 deficiency aggravates healing and remodeling in male mice after experimental myocardial infarction. Circ Heart Fail 7:822–830. https://doi.org/10.1161/CIRCHEARTFAILURE.113.001020
Horenstein MS, Vander Heide RS, L’Ecuyer TJ (2000) Molecular basis of anthracycline-induced cardiotoxicity and its prevention. Mol Genet Metab 71:436–444. https://doi.org/10.1006/MGME.2000.3043
Jahn SK, Hennicke T, Kassack MU, Drews L, Reichert AS, Fritz G (2020) Distinct influence of the anthracycline derivative doxorubicin on the differentiation efficacy of mESC-derived endothelial progenitor cells. Biochim Biophys Acta Mol Cell Res. https://doi.org/10.1016/J.BBAMCR.2020.118711
Jong WMC, Leemans JC, Weber NC, Juffermans NP, Schultz MJ, Hollmann MW, Zuurbier CJ (2014) Nlrp3 plays no role in acute cardiac infarction due to low cardiac expression. Int J Cardiol 177:41–43. https://doi.org/10.1016/J.IJCARD.2014.09.148
Joseph RW, Elassaiss-Schaap J, Kefford R, Hwu WJ, Wolchok JD, Joshua AM, Ribas A, Hodi FS, Hamid O, Robert C, Daud A, Dronca R, Hersey P, Weber JS, Patnaik A, De Alwis DP, Perrone A, Zhang J, Kang SP, Ebbinghaus S, Anderson KM, Gangadhar TC (2018) Baseline tumor size is an independent prognostic factor for overall survival in patients with melanoma treated with pembrolizumab. Clin Cancer Res 24:4960–4967. https://doi.org/10.1158/1078-0432.CCR-17-2386
Kaji K, Nishimura N, Seki K, Sato S, Saikawa S, Nakanishi K, Furukawa M, Kawaratani H, Kitade M, Moriya K, Namisaki T, Yoshiji H (2018) Sodium glucose cotransporter 2 inhibitor canagliflozin attenuates liver cancer cell growth and angiogenic activity by inhibiting glucose uptake. Int J Cancer 142:1712–1722. https://doi.org/10.1002/IJC.31193
Kelley N, Jeltema D, Duan Y, He Y (2019) The NLRP3 inflammasome: an overview of mechanisms of activation and regulation. Int J Mol Sci 20:3328. https://doi.org/10.3390/IJMS20133328
Kenny EM, Fidan E, Yang Q, Anthonymuthu TS, New LA, Meyer EA, Wang H, Kochanek PM, Dixon CE, Kagan VE, Bayir H (2019) Ferroptosis contributes to neuronal death and functional outcome after traumatic brain injury∗. Crit Care Med 47:410–418. https://doi.org/10.1097/CCM.0000000000003555
Kerkela R, Woulfe KC, Durand JB, Vagnozzi R, Kramer D, Chu TF, Beahm C, Chen MH, Force T (2009) Sunitinib-induced cardiotoxicity is mediated by off-target inhibition of AMP-activated protein kinase. Clin Transl Sci 2:15–25. https://doi.org/10.1111/J.1752-8062.2008.00090.X
Khoury MK, Yang H, Liu B (2021) Macrophage biology in cardiovascular diseases. Arterioscler Thromb Vasc Biol 41:E77–E81. https://doi.org/10.1161/ATVBAHA.120.313584
Khunger A, Battel L, Wadhawan A, More A, Kapoor A, Agrawal N (2020) New insights into mechanisms of immune checkpoint inhibitor-induced cardiovascular toxicity. Curr Oncol Rep. https://doi.org/10.1007/S11912-020-00925-8
Kim SR, Lee SG, Kim SH, Kim JH, Choi E, Cho W, Rim JH, Hwang I, Lee CJ, Lee M, Oh CM, Jeon JY, Gee HY, Kim JH, Lee BW, Kang ES, Cha BS, Lee MS, Yu JW, Cho JW, Kim JS, Lee YH (2020) SGLT2 inhibition modulates NLRP3 inflammasome activity via ketones and insulin in diabetes with cardiovascular disease. Nat Commun 11:1–11. https://doi.org/10.1038/s41467-020-15983-6
Koelzer VH, Rothschild SI, Zihler D, Wicki A, Willi B, Willi N, Voegeli M, Cathomas G, Zippelius A, Mertz KD (2016) Systemic inflammation in a melanoma patient treated with immune checkpoint inhibitors-an autopsy study. J Immunother Cancer. https://doi.org/10.1186/S40425-016-0117-1
Kounatidis D, Vallianou N, Evangelopoulos A, Vlahodimitris I, Grivakou E, Kotsi E, Dimitriou K, Skourtis A, Mourouzis I (2023) SGLT-2 Inhibitors and the inflammasome: what’s next in the 21st century? Nutrients 15:2294. https://doi.org/10.3390/NU15102294
Kuang H, Liao L, Chen H, Kang Q, Shu X, Wang Y (2017) Therapeutic effect of sodium glucose co-transporter 2 inhibitor dapagliflozin on renal cell carcinoma. Med Sci Monit 23:3737–3745. https://doi.org/10.12659/MSM.902530
Lau KTK, Ng L, Wong JWH, Loong HHF, Chan WWL, Lee CH, Wong CKH (2021) Repurposing sodium–glucose co-transporter 2 inhibitors (SGLT2i) for cancer treatment—a review. Rev Endocr Metab Disord 22:1121–1136. https://doi.org/10.1007/S11154-021-09675-9
Lauritsen KM, Søndergaard E, Svart M, Møller N, Gormsen LC (2018) Ketone body infusion increases circulating erythropoietin and bone marrow glucose uptake. Diabetes Care 41:e152–e154. https://doi.org/10.2337/DC18-1421
Lauritsen KM, Voigt JH, Pedersen SB, Hansen TK, Møller N, Jessen N, Gormsen LC, Søndergaard E (2022) Effects of SGLT2 inhibition on lipid transport in adipose tissue in type 2 diabetes. Endocr Connect. https://doi.org/10.1530/EC-21-0558
Lavine KJ, Pinto AR, Epelman S, Kopecky BJ, Clemente-Casares X, Godwin J, Rosenthal N, Kovacic JC (2018) The macrophage in cardiac homeostasis and disease: JACC macrophage in CVD series (Part 4). J Am Coll Cardiol 72:2213–2230. https://doi.org/10.1016/J.JACC.2018.08.2149
Lee N, Heo YJ, Choi SE, Jeon JY, Han SJ, Kim DJ, Kang Y, Lee KW, Kim HJ (2021) Anti-inflammatory effects of empagliflozin and gemigliptin on LPS-stimulated macrophage via the IKK/NF-κB, MKK7/JNK, and JAK2/STAT1 signalling pathways. J Immunol Res. https://doi.org/10.1155/2021/9944880
Lee TI, Chen YC, Lin YK, Chung CC, Lu YY, Kao YH, Chen YJ (2019) Empagliflozin attenuates myocardial sodium and calcium dysregulation and reverses cardiac remodeling in streptozotocin-induced diabetic rats. Int J Mol Sci 20:1680. https://doi.org/10.3390/IJMS20071680
Lee TM, Chang NC, Lin SZ (2017) Dapagliflozin, a selective SGLT2 Inhibitor, attenuated cardiac fibrosis by regulating the macrophage polarization via STAT3 signaling in infarcted rat hearts. Free Radic Biol Med 104:298–310. https://doi.org/10.1016/J.FREERADBIOMED.2017.01.035
Li J, Yu Y, Sun Y, Yu B, Tan X, Wang B, Lu Y, Wang N (2023) SGLT2 inhibition, circulating metabolites, and atrial fibrillation: a mendelian randomization study. Cardiovasc Diabetol 22:278. https://doi.org/10.1186/s12933-023-02019-8
Liberti MV, Locasale JW (2016) The Warburg effect: how does it benefit cancer cells? Trends Biochem Sci 41:211–218. https://doi.org/10.1016/j.tibs.2015.12.001
Lin B, Koibuchi N, Hasegawa Y, Sueta D, Toyama K, Uekawa K, Ma MJ, Nakagawa T, Kusaka H, Kim-Mitsuyama S (2014) Glycemic control with empagliflozin, a novel selective SGLT2 inhibitor, ameliorates cardiovascular injury and cognitive dysfunction in obese and type 2 diabetic mice. Cardiovasc Diabetol. https://doi.org/10.1186/S12933-014-0148-1
Lin M, Xiong W, Wang S, Li Y, Hou C, Li C, Li G (2022) The Research progress of trastuzumab-induced cardiotoxicity in HER-2-positive breast cancer treatment. Front Cardiovasc Med 8:821663. https://doi.org/10.3389/FCVM.2021.821663
Linkermann A, Skouta R, Himmerkus N, Mulay SR, Dewitz C, De Zen F, Prokai A, Zuchtriegel G, Krombach F, Welz PS, Weinlich R, Vanden BT, Vandenabeele P, Pasparakis M, Bleich M, Weinberg JM, Reichel CA, Bräsen JH, Kunzendorf U, Anders HJ, Stockwell BR, Green DR, Krautwald S (2014) Synchronized renal tubular cell death involves ferroptosis. Proc Natl Acad Sci U S A 111:16836–16841. https://doi.org/10.1073/PNAS.1415518111/SUPPL_FILE/PNAS.1415518111.SM04.AVI
Lőrincz T, Jemnitz K, Kardon T, Mandl J, Szarka A (2015) Ferroptosis is involved in acetaminophen induced cell death. Pathol Oncol Res 21:1115–1121. https://doi.org/10.1007/S12253-015-9946-3
Ma S, He LL, Zhang GR, Zuo QJ, Wang ZL, Zhai JL, Zhang TT, Wang Y, Ma HJ, Guo YF (2022) Canagliflozin mitigates ferroptosis and ameliorates heart failure in rats with preserved ejection fraction. Naunyn Schmiedebergs Arch Pharmacol 395:945–962. https://doi.org/10.1007/S00210-022-02243-1/FIGURES/9
Madonna R, Cadeddu C, Deidda M, Mele D, Monte I, Novo G, Pagliaro P, Pepe A, Spallarossa P, Tocchetti CG, Zito C, Mercuro G (2015) Improving the preclinical models for the study of chemotherapy-induced cardiotoxicity: a position paper of the Italian Working Group on Drug Cardiotoxicity and Cardioprotection. Heart Fail Rev 20:621–631. https://doi.org/10.1007/S10741-015-9497-4
Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M (2004) The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol 25:677–686. https://doi.org/10.1016/j.it.2004.09.015
Mascolo A, Scavone C, Ferrajolo C et al (2021) Immune checkpoint inhibitors and cardiotoxicity: an analysis of spontaneous reports in eudravigilance. Drug Saf 44:957–971
Mathupala SP, Ko YH, Pedersen PL (2006) Hexokinase II: cancer’s double-edged sword acting as both facilitator and gatekeeper of malignancy when bound to mitochondria. Oncogene 25:4777–4786. https://doi.org/10.1038/SJ.ONC.1209603
Mazer CD, Hare GMT, Connelly PW, Gilbert RE, Shehata N, Quan A, Teoh H, Leiter LA, Zinman B, Jüni P, Zuo F, Mistry N, Thorpe KE, Goldenberg RM, Yan AT, Connelly KA, Verma S (2020) Effect of empagliflozin on erythropoietin levels, iron stores, and red blood cell morphology in patients with type 2 diabetes mellitus and coronary artery disease. Circulation 141:704–707. https://doi.org/10.1161/CIRCULATIONAHA.119.044235
McDonagh TA, Metra M, Adamo M, Baumbach A, Böhm M, Burri H, Čelutkiene J, Chioncel O, Cleland JGF, Coats AJS, Crespo-Leiro MG, Farmakis D, Gardner RS, Gilard M, Heymans S, Hoes AW, Jaarsma T, Jankowska EA, Lainscak M, Lam CSP, Lyon AR, McMurray JJV, Mebazaa A, Mindham R, Muneretto C, Piepoli MF, Price S, Rosano GMC, Ruschitzka F, Skibelund AK, de Boer RA, Schulze PC, Abdelhamid M, Aboyans V, Adamopoulos S, Anker SD, Arbelo E, Asteggiano R, Bauersachs J, Bayes-Genis A, Borger MA, Budts W, Cikes M, Damman K, Delgado V, Dendale P, Dilaveris P, Drexel H, Ezekowitz J, Falk V, Fauchier L, Filippatos G, Fraser A, Frey N, Gale CP, Gustafsson F, Harris J, Iung B, Janssens S, Jessup M, Konradi A, Kotecha D, Lambrinou E, Lancellotti P, Landmesser U, Leclercq C, Lewis BS, Leyva F, Linhart A, Løchen ML, Lund LH, Mancini D, Masip J, Milicic D, Mueller C, Nef H, Nielsen JC, Neubeck L, Noutsias M, Petersen SE, Petronio AS, Ponikowski P, Prescott E, Rakisheva A, Richter D, Schlyakhto E, Seferovic P, Senni M, Sitges M, Sousa-Uva M, Tocchetti CG, Touyz R, Tschoepe C, Waltenberger J, Krim M, Hayrapetyan H, Moertl D, Mustafayev I, Kurlianskaya A, Depauw M, Kušljugić Z, Gatzov P, Agathangelou P, Melenovský V, Løgstrup BB, Mostafa AM, Uuetoa T, Lassus J, Logeart D, Kipiani Z, Chrysohoou C, Sepp R, Ingimarsdóttir IJ, O’Neill J, Gotsman I, Iacoviello M, Bajraktari G, Lunegova O, Kamzola G, Massih TA, Benlamin H, Žaliaduonyte D, Noppe S, Moore A, Vataman E, Boskovic A, Bennis A, Manintveld OC, Kostovska ES, Gulati G, Straburzyńska-Migaj E, Silva-Cardoso J, Rimbaş RC, Lopatin Y, Foscoli M, Stojkovic S, Goncalvesova E, Fras Z, Segovia J, Lindmark K, Maeder MT, Bsata W, Abid L, Altay H, Voronkov L, Davies C, Abdullaev T, Baigent CN, Antoniou S, Collet JP, Halvorsen S, Koskinas KC (2021) 2021 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure. Eur Heart J 42:3599–3726. https://doi.org/10.1093/EURHEARTJ/EHAB368
McDonagh TA, Metra M, Adamo M, Gardner RS, Baumbach A, Böhm M, Burri H, Butler J, Čelutkienė J, Chioncel O, Cleland JGF, Crespo-Leiro MG, Farmakis D, Gilard M, Heymans S, Hoes AW, Jaarsma T, Jankowska EA, Lainscak M, Lam CSP, Lyon AR, McMurray JJV, Mebazaa A, Mindham R, Muneretto C, Francesco Piepoli M, Price S, Rosano GMC, Ruschitzka F, Skibelund AK, de Boer RA, Schulze PC, Arbelo E, Bartunek J, Bauersachs J, Borger MA, Buccheri S, Cerbai E, Donal E, Edelmann F, Färber G, Heidecker B, Ibanez B, James S, Køber L, Koskinas KC, Masip J, McEvoy JW, Mentz R, Mihaylova B, Møller JE, Mullens W, Neubeck L, Nielsen JC, Pasquet AA, Ponikowski P, Prescott E, Rakisheva A, Rocca B, Rossello X, Sade LE, Schaubroeck H, Tessitore E, Tokmakova M, van der Meer P, Van Gelder IC, Van Heetvelde M, Vrints C, Wilhelm M, Witkowski A, Zeppenfeld K, Shuka N, Chettibi M, Hayrapetyan H, Pavo N, Islamli A, Pouleur A-C, Kusljugic Z, Tokmakova M, Milicic D, Christodoulides T, Malek F, Køber L, Koriem MAG, Põder P, Lassus J, Roubille F, Agladze V, Frantz S, Stavrati A, Kosztin A, Ingimarsdóttir IJ, Campbell P, Hasin T, Oliva F, Aidargaliyeva N, Bajraktari G, Mirrakhimov E, Kamzola G, El Neihoum AM, Zaliaduonyte D, Moore A, Vataman E, Boskovic A, Alami M, Manintveld O, Kostovska ES, Broch K, Nessler J, Franco F, Popescu BA, Foscoli M, Milosavljevic AS, Goncalvesova E, Fras Z, Gonzalez-Costello J, Lindmark K, Paul M, Oudeh A, Zakhama L, Celik A, Voronkov L, Clark A, Abdullaev T, Prescott E, James S, Arbelo E, Baigent C, Borger MA, Buccheri S, Ibanez B, Køber L, Koskinas KC, McEvoy JW, Mihaylova B, Mindham R, Neubeck L, Nielsen JC, Pasquet AA, Rakisheva A, Rocca B, Rossello X, Vaartjes I, Vrints C, Witkowski A, Zeppenfeld K (2023) 2023 focused update of the 2021 ESC guidelines for the diagnosis and treatment of acute and chronic heart failure. Eur Heart J. https://doi.org/10.1093/EURHEARTJ/EHAD195
McMurray JJV, Solomon SD, Inzucchi SE, Køber L, Kosiborod MN, Martinez FA, Ponikowski P, Sabatine MS, Anand IS, Bělohlávek J, Böhm M, Chiang C-E, Chopra VK, de Boer RA, Desai AS, Diez M, Drozdz J, Dukát A, Ge J, Howlett JG, Katova T, Kitakaze M, Ljungman CEA, Merkely B, Nicolau JC, O’Meara E, Petrie MC, Vinh PN, Schou M, Tereshchenko S, Verma S, Held C, DeMets DL, Docherty KF, Jhund PS, Bengtsson O, Sjöstrand M, Langkilde A-M (2019) Dapagliflozin in patients with heart failure and reduced ejection fraction. N Engl J Med 381:1995–2008. https://doi.org/10.1056/NEJMOA1911303
Melloni C, Shrader P, Carver J, Piccini JP, Thomas L, Fonarow GC, Ansell J, Gersh B, Go AS, Hylek E, Herling IM, Mahaffey KW, Yu AF, Peterson ED, Kowey PR (2017) Management and outcomes of patients with atrial fibrillation and a history of cancer: the ORBIT-AF registry. Eur Heart J Qual Care Clin Outcomes 3:192–197. https://doi.org/10.1093/EHJQCCO/QCX004
Mezzaroma E, Toldo S, Farkas D, Seropian IM, Van Tassell BW, Salloum FN, Kannan HR, Menna AC, Voelkel NF, Abbate A (2011) The inflammasome promotes adverse cardiac remodeling following acute myocardial infarction in the mouse. Proc Natl Acad Sci U S A 108:19725–19730. https://doi.org/10.1073/PNAS.1108586108/SUPPL_FILE/PNAS.201108586SI.PDF
Min J, Wu L, Liu Y, Song G, Deng Q, Jin W, Yu W, Miyesaier A, Pei Z, Ren J (2023) Empagliflozin attenuates trastuzumab-induced cardiotoxicity through suppression of DNA damage and ferroptosis. Life Sci 312:121207. https://doi.org/10.1016/J.LFS.2022.121207
Minotti G, Menna P, Camilli M, Salvatorelli E, Levi R (2022) Beyond hypertension: diastolic dysfunction associated with cancer treatment in the era of cardio-oncology. Adv Pharmacol 94:365–409. https://doi.org/10.1016/BS.APHA.2022.02.002
Minotti G, Reggiardo G, Camilli M, Salvatorelli E, Menna P (2022) From cardiac anthracycline accumulation to real-life risk for early diastolic dysfunction: a translational approach. Cardio Oncology 4:139–140. https://doi.org/10.1016/J.JACCAO.2021.12.002
Monica Reddy RP, Inzucchi SE (2016) SGLT2 inhibitors in the management of type 2 diabetes. Endocrine 53:364–372. https://doi.org/10.1007/S12020-016-0943-4
Moullet I, Salles G, Ketterer N, Dumontet C, Bouafia F, Neidhart-Berard EM, Thieblemont C, Felman P, Coiffier B (1998) Frequency and significance of anemia in non-Hodgkin’s lymphoma patients. Ann Oncol 9:1109–1115. https://doi.org/10.1023/A:1008498705032
Nakachi S, Okamoto S, Tamaki K, Nomura I, Tomihama M, Nishi Y, Fukushima T, Tanaka Y, Morishima S, Imamura M, Maeda S, Tsutsui M, Matsushita M, Masuzaki H (2022) Impact of anti-diabetic sodium–glucose cotransporter 2 inhibitors on tumor growth of intractable hematological malignancy in humans. Biomed Pharmacother. https://doi.org/10.1016/J.BIOPHA.2022.112864
Neal B, Perkovic V, Mahaffey KW, de Zeeuw D, Fulcher G, Erondu N, Shaw W, Law G, Desai M, Matthews DR (2017) Canagliflozin and cardiovascular and renal events in type 2 diabetes. N Engl J Med 377:644–657. https://doi.org/10.1056/NEJMOA1611925/SUPPL_FILE/NEJMOA1611925_DISCLOSURES.PDF
Nikolaou PE, Efentakis P, Abu Qourah F, Femminò S, Makridakis M, Kanaki Z, Varela A, Tsoumani M, Davos CH, Dimitriou CA, Tasouli A, Dimitriadis G, Kostomitsopoulos N, Zuurbier CJ, Vlahou A, Klinakis A, Brizzi MF, Iliodromitis EK, Andreadou I (2021) Chronic empagliflozin treatment reduces myocardial infarct size in nondiabetic mice through STAT-3-mediated protection on microvascular endothelial cells and reduction of oxidative stress. Antioxid Redox Signal 34:551–571. https://doi.org/10.1089/ARS.2019.7923
Oh CM, Cho S, Jang JY, Kim H, Chun S, Choi M, Park S, Ko YG (2019) Cardioprotective potential of an SGLT2 inhibitor against doxorubicin-induced heart failure. Korean Circ J 49:1183–1195. https://doi.org/10.4070/KCJ.2019.0180
Packer M (2022) How can sodium–glucose cotransporter 2 inhibitors stimulate erythrocytosis in patients who are iron-deficient? Implications for understanding iron homeostasis in heart failure. Eur J Heart Fail 24:2287–2296. https://doi.org/10.1002/EJHF.2731
Packer M (2023) Mechanisms of enhanced renal and hepatic erythropoietin synthesis by sodium–glucose cotransporter 2 inhibitors. Eur Heart J. https://doi.org/10.1093/EURHEARTJ/EHAD235
Park LK, Lim K-H, Volkman J, Abdiannia M, Johnston H, Nigogosyan Z, Siegel MJ, McGill JB, McKee AM, Salam M, Zhang RM, Ma D, Popuri K, Chow VTY, Beg MF, Hawkins WG, Peterson LR (2023) Safety, tolerability, and effectiveness of the sodium–glucose cotransporter 2 inhibitor (SGLT2i) dapagliflozin in combination with standard chemotherapy for patients with advanced, inoperable pancreatic adenocarcinoma: a phase 1b observational study. Cancer Metab 11:1–12. https://doi.org/10.1186/S40170-023-00306-2
Park TJ, Park JH, Lee GS, Lee JY, Shin JH, Kim MW, Kim YS, Kim JY, Oh KJ, Han BS, Kim WK, Ahn Y, Moon JH, Song J, Bae KH, Kim DH, Lee EW, Lee SC (2019) Quantitative proteomic analyses reveal that GPX4 downregulation during myocardial infarction contributes to ferroptosis in cardiomyocytes. Cell Death Dis 10:835. https://doi.org/10.1038/S41419-019-2061-8
Pennig J, Scherrer P, Gissler MC, Anto-Michel N, Hoppe N, Füner L, Härdtner C, Stachon P, Wolf D, Hilgendorf I, Mullick A, Bode C, Zirlik A, Goldberg IJ, Willecke F (2019) Glucose lowering by SGLT2-inhibitor empagliflozin accelerates atherosclerosis regression in hyperglycemic STZ-diabetic mice. Sci Rep 9:1–12. https://doi.org/10.1038/s41598-019-54224-9
Perelman MG, Brzezinski RY, Waissengrin B, Leshem Y, Bainhoren O, Rubinstein TA, Perelman M, Rozenbaum Z, Havakuk O, Topilsky Y, Banai S, Wolf I, Laufer-Perl M (2024) Sodium–glucose co-transporter-2 inhibitors in patients treated with immune checkpoint inhibitors. Cardiooncology. https://doi.org/10.1186/S40959-023-00199-6
Postow MA, Sidlow R, Hellmann MD (2018) Immune-related adverse events associated with immune checkpoint blockade. N Engl J Med 378:158–168. https://doi.org/10.1056/NEJMRA1703481
Qin W, Hu L, Zhang X, Jiang S, Li J, Zhang Z, Wang X (2019) The diverse function of PD-1/PD-L pathway beyond cancer. Front Immunol. https://doi.org/10.3389/FIMMU.2019.02298
Quagliariello V, De Laurentiis M, Rea D, Barbieri A, Monti MG, Carbone A, Paccone A, Altucci L, Conte M, Canale ML, Botti G, Maurea N (2021) The SGLT-2 inhibitor empagliflozin improves myocardial strain, reduces cardiac fibrosis and pro-inflammatory cytokines in non-diabetic mice treated with doxorubicin. Cardiovasc Diabetol. https://doi.org/10.1186/S12933-021-01346-Y
Quartermaine C, Ghazi SM, Yasin A, Awan FT, Fradley M, Wiczer T, Kalathoor S, Ferdousi M, Krishan S, Habib A, Shaaban A, Kola-Kehinde O, Kittai AS, Rogers KA, Grever M, Ruz P, Bhat S, Dickerson T, Byrd JC, Woyach J, Addison D (2023) Cardiovascular toxicities of BTK inhibitors in chronic lymphocytic leukemia: JACC: cardiooncology state-of-the-art review. JACC CardioOncol 5:570–590. https://doi.org/10.1016/J.JACCAO.2023.09.002
Refaie MMM, Shehata S, El-Hussieny M, Fawzy MA, Ahmed NZM, Marey H, Hishmat AM, Alkully T, El RESMA (2023) Molecular mechanism of empagliflozin cardioprotection in 5-fluorouracil (5-FU)-induced cardiotoxicity via modulation of SGLT2 and TNFα/TLR/NF-κB signaling pathway in rats. Toxicol Res 40:139–151. https://doi.org/10.1007/S43188-023-00204-1
Reis-Mendes A, Padrão AI, Duarte JA, Gonçalves-Monteiro S, Duarte-Araújo M, Remião F, Carvalho F, Sousa E, Bastos ML, Costa VM (2021) Role of inflammation and redox status on doxorubicin-induced cardiotoxicity in infant and adult CD-1 male mice. Biomolecules. https://doi.org/10.3390/BIOM11111725
Ren C, Sun K, Zhang Y, Hu Y, Hu B, Zhao J, He Z, Ding R, Wang W, Liang C (2021) Sodium–glucose cotransporter-2 inhibitor empagliflozin ameliorates sunitinib-induced cardiac dysfunction via regulation of AMPK-mTOR signaling pathway-mediated autophagy. Front Pharmacol. https://doi.org/10.3389/FPHAR.2021.664181
Renu K, Abilash VG, Tirupathi TP, Arunachalam S (2017) Molecular mechanism of doxorubicin-induced cardiomyopathy—an update. Eur J Pharmacol 818:241–253. https://doi.org/10.1016/J.EJPHAR.2017.10.043
Riehle C, Abel ED (2012) PGC-1 proteins and heart failure. Trends Cardiovasc Med 22:98–105. https://doi.org/10.1016/J.TCM.2012.07.003
Rogava M, Aprati TJ, Chi WY, Melms JC, Hug C, Davis SH, Earlie EM, Chung C, Deshmukh SK, Wu S, Sledge G, Tang S, Ho P, Amin AD, Caprio L, Gurjao C, Tagore S, Ngo B, Lee MJ, Zanetti G, Wang Y, Chen S, Ge W, Melo LMN, Allies G, Rösler J, Gibney GT, Schmitz OJ, Sykes M, Creusot RJ, Tüting T, Schadendorf D, Röcken M, Eigentler TK, Molotkov A, Mintz A, Bakhoum SF, Beyaz S, Cantley LC, Sorger PK, Meckelmann SW, Tasdogan A, Liu D, Laughney AM, Izar B (2024) Loss of Pip4k2c confers liver-metastatic organotropism through insulin-dependent PI3K-AKT pathway activation. Nat Cancer. https://doi.org/10.1038/S43018-023-00704-X
Rokszin G, Kiss Z, Sütő G, Kempler P, Jermendy G, Fábián I, Szekanecz Z, Poór G, Wittmann I, Molnár GA (2021) Sodium–glucose co-transporter 2 inhibitors may change the development of urinary tract and hematological malignancies as compared with dipeptidyl peptidase-4 inhibitors: data of the post-hoc analysis of a nationwide study. Front Oncol. https://doi.org/10.3389/FONC.2021.725465
Ryaboshapkina M, Ye R, Ye Y, Birnbaum Y (2023) Effects of dapagliflozin on myocardial gene expression in btbr mice with type 2 diabetes. Cardiovasc Drugs Ther. https://doi.org/10.1007/s10557-023-07517-1
Sabatino J, De Rosa S, Tammè L, Iaconetti C, Sorrentino S, Polimeni A, Mignogna C, Amorosi A, Spaccarotella C, Yasuda M, Indolfi C (2020) Empagliflozin prevents doxorubicin-induced myocardial dysfunction. Cardiovasc Diabetol 19:66. https://doi.org/10.1186/S12933-020-01040-5
Salloum FN, Tocchetti CG, Ameri P, Ardehali H, Asnani A, de Boer RA, Burridge P, Cabrera JÁ, de Castro J, Córdoba R, Costa A, Dent S, Engelbertsen D, Fernández-Velasco M, Fradley M, Fuster JJ, Galán-Arriola C, García-Lunar I, Ghigo A, González-Neira A, Hirsch E, Ibáñez B, Kitsis RN, Konety S, Lyon AR, Martin P, Mauro AG, Mazo Vega MM, Meijers WC, Neilan TG, Rassaf T, Ricke-Hoch M, Sepulveda P, Thavendiranathan P, van der Meer P, Fuster V, Ky B, López-Fernández T (2023) Priorities in cardio-oncology basic and translational science: GCOS 2023 symposium proceedings: JACC: cardiooncology state-of-the-art review. JACC CardioOncol 5:715–731. https://doi.org/10.1016/J.JACCAO.2023.08.003
Sandanger GE, Ranheim T, Bliksøen M, Kaasbøll OJ, Alfsnes K, Nymo SH, Rashidi A, Ohm IK, Attramadal H, Aukrust P, Vinge LE, Yndestad A (2016) NLRP3 inflammasome activation during myocardial ischemia reperfusion is cardioprotective. Biochem Biophys Res Commun 469:1012–1020. https://doi.org/10.1016/J.BBRC.2015.12.051
Sano M, Goto S (2019) Possible mechanism of hematocrit elevation by sodium glucose cotransporter 2 inhibitors and associated beneficial renal and cardiovascular effects. Circulation 139:1985–1987. https://doi.org/10.1161/CIRCULATIONAHA.118.038881
Sawyer DB (2013) Anthracyclines and heart failure. N Engl J Med 368:1154–1156. https://doi.org/10.1056/NEJMCIBR1214975/SUPPL_FILE/NEJMCIBR1214975_DISCLOSURES.PDF
Sawyer DB, Fukazawa R, Arstall MA, Kelly RA (1999) Daunorubicin-induced apoptosis in rat cardiac myocytes is inhibited by dexrazoxane. Circ Res 84:257–265. https://doi.org/10.1161/01.RES.84.3.257
Scafoglio C, Hirayama BA, Kepe V, Liu J, Ghezzi C, Satyamurthy N, Moatamed NA, Huang J, Koepsell H, Barrio JR, Wright EM (2015) Functional expression of sodium–glucose transporters in cancer. Proc Natl Acad Sci U S A 112:E4111–E4119. https://doi.org/10.1073/PNAS.1511698112
Scisciola L, Cataldo V, Taktaz F, Fontanella RA, Pesapane A, Ghosh P, Franzese M, Puocci A, De Angelis A, Sportiello L, Marfella R, Barbieri M (2022) Anti-inflammatory role of SGLT2 inhibitors as part of their anti-atherosclerotic activity: data from basic science and clinical trials. Front Cardiovasc Med. https://doi.org/10.3389/FCVM.2022.1008922
Shiga T, Hiraide M (2020) Cardiotoxicities of 5-fluorouracil and other fluoropyrimidines. Curr Treat Options Oncol. https://doi.org/10.1007/S11864-020-0719-1
Solomon SD, McMurray JJV, Claggett B, de Boer RA, DeMets D, Hernandez AF, Inzucchi SE, Kosiborod MN, Lam CSP, Martinez F, Shah SJ, Desai AS, Jhund PS, Belohlavek J, Chiang C-E, Borleffs CJW, Comin-Colet J, Dobreanu D, Drozdz J, Fang JC, Alcocer-Gamba MA, Al Habeeb W, Han Y, Cabrera Honorio JW, Janssens SP, Katova T, Kitakaze M, Merkely B, O’Meara E, Saraiva JFK, Tereshchenko SN, Thierer J, Vaduganathan M, Vardeny O, Verma S, Pham VN, Wilderäng U, Zaozerska N, Bachus E, Lindholm D, Petersson M, Langkilde AM (2022) Dapagliflozin in heart failure with mildly reduced or preserved ejection fraction. N Engl J Med 387:1089–1098. https://doi.org/10.1056/NEJMOA2206286/SUPPL_FILE/NEJMOA2206286_DATA-SHARING.PDF
Sun L, Wang H, Yu S, Zhang L, Jiang J, Zhou Q (2022) Herceptin induces ferroptosis and mitochondrial dysfunction in H9c2 cells. Int J Mol Med 49:1–8. https://doi.org/10.3892/IJMM.2021.5072/HTML
Swe MT, Thongnak L, Jaikumkao K, Pongchaidecha A, Chatsudthipong V, Lungkaphin A (2019) Dapagliflozin not only improves hepatic injury and pancreatic endoplasmic reticulum stress, but also induces hepatic gluconeogenic enzymes expression in obese rats. Clin Sci 133:2415–2430. https://doi.org/10.1042/CS20190863
Szekeres Z, Toth K, Szabados E (2021) The effects of SGLT2 inhibitors on lipid metabolism. Metabolites 11:1–9. https://doi.org/10.3390/METABO11020087
Tan TC, Neilan TG, Francis S, Plana JC, Scherrer-Crosbie M (2015) Anthracycline-induced cardiomyopathy in adults. Compr Physiol 5:1517–1540. https://doi.org/10.1002/CPHY.C140059
Tewey KM, Chenb GL, Nelsonf EM, Lius LF (1984) Intercalative antitumor drugs interfere with the breakage-reunion reaction of mammalian DNA topoisomerase 11. J Biol Chem 259:9182–9187. https://doi.org/10.1016/S0021-9258(17)47282-6
Tisi MC, Bozzoli V, Giachelia M, Massini G, Ricerca BM, Maiolo E, D’Alo F, Larocca LM, Piciocchi A, Tjalsma H, Swinkels DW, Voso MT, Leone G, Hohaus S (2014) Anemia in diffuse large B-cell non-Hodgkin lymphoma: the role of interleukin-6, hepcidin and erythropoietin. Leuk Lymphoma 55:270–275. https://doi.org/10.3109/10428194.2013.802314
Tokarska-Schlattner M, Zaugg M, Zuppinger C, Wallimann T, Schlattner U (2006) New insights into doxorubicin-induced cardiotoxicity: the critical role of cellular energetics. J Mol Cell Cardiol 41:389–405. https://doi.org/10.1016/J.YJMCC.2006.06.009
Totzeck M, Schuler M, Stuschke M, Heusch G, Rassaf T (2019) Cardio-oncology - strategies for management of cancer-therapy related cardiovascular disease. Int J Cardiol 280:163–175. https://doi.org/10.1016/j.ijcard.2019.01.038
Tsuchiya T, Kominato Y, Ueda M (2002) Human hypoxic signal transduction through a signature motif in hepatocyte nuclear factor 4. J Biochem 132:37–44. https://doi.org/10.1093/OXFORDJOURNALS.JBCHEM.A003196
Uhlén M, Fagerberg L, Hallström BM, Lindskog C, Oksvold P, Mardinoglu A, Sivertsson Å, Kampf C, Sjöstedt E, Asplund A, Olsson IM, Edlund K, Lundberg E, Navani S, Szigyarto CAK, Odeberg J, Djureinovic D, Takanen JO, Hober S, Alm T, Edqvist PH, Berling H, Tegel H, Mulder J, Rockberg J, Nilsson P, Schwenk JM, Hamsten M, Von Feilitzen K, Forsberg M, Persson L, Johansson F, Zwahlen M, Von Heijne G, Nielsen J, Pontén F (2015) Tissue-based map of the human proteome. Science 1979:347. https://doi.org/10.1126/SCIENCE.1260419/SUPPL_FILE/1260419_UHLEN.SM.PDF
Uthman L, Baartscheer A, Bleijlevens B, Schumacher CA, Fiolet JWT, Koeman A, Jancev M, Hollmann MW, Weber NC, Coronel R, Zuurbier CJ (2018) Class effects of SGLT2 inhibitors in mouse cardiomyocytes and hearts: inhibition of Na+/H+ exchanger, lowering of cytosolic Na+ and vasodilation. Diabetologia 61:722–726. https://doi.org/10.1007/S00125-017-4509-7
Uthman L, Kuschma M, Römer G, Boomsma M, Kessler J, Hermanides J, Hollmann MW, Preckel B, Zuurbier CJ, Weber NC (2021) Novel anti-inflammatory effects of canagliflozin involving hexokinase ii in lipopolysaccharide-stimulated human coronary artery endothelial cells. Cardiovasc Drugs Ther 35:1083–1094. https://doi.org/10.1007/S10557-020-07083-W
Vallon V, Thomson SC (2017) Targeting renal glucose reabsorption to treat hyperglycaemia: the pleiotropic effects of SGLT2 inhibition. Diabetologia 60:215–225. https://doi.org/10.1007/S00125-016-4157-3
Vaziri Z, Saleki K, Aram C, Alijanizadeh P, Pourahmad R, Azadmehr A, Ziaei N (2023) Empagliflozin treatment of cardiotoxicity: a comprehensive review of clinical, immunobiological, neuroimmune, and therapeutic implications. Biomed Pharmacother 168:115686. https://doi.org/10.1016/J.BIOPHA.2023.115686
Vejpongsa P, Yeh ETH (2014) Topoisomerase 2β: a promising molecular target for primary prevention of anthracycline-induced cardiotoxicity. Clin Pharmacol Ther 95:45–52. https://doi.org/10.1038/CLPT.2013.201
Villeneuve C, Guilbeau-Frugier C, Sicard P, Lairez O, Ordener C, Duparc T, De Paulis D, Couderc B, Spreux-Varoquaux O, Tortosa F, Garnier A, Knauf C, Valet P, Borchi E, Nediani C, Gharib A, Ovize M, Delisle MB, Parini A, Mialet-Perez J (2012) p53-PGC-1α pathway mediates oxidative mitochondrial damage and cardiomyocyte necrosis induced by monoamine oxidase-A upregulation: role in chronic left ventricular dysfunction in mice. Antioxid Redox Signal 18:5–18. https://doi.org/10.1089/ARS.2011.4373
Weiss G, Goodnough LT (2005) Anemia of chronic disease. N Engl J Med 352:1011–1023. https://doi.org/10.1056/NEJMRA041809
Wright EM (2020) SGLT2 and cancer. Pflugers Arch 472:1407–1414. https://doi.org/10.1007/S00424-020-02448-4
Wu W, Zhang Z, Jing D, Huang X, Ren D, Shao Z, Zhang Z (2022) SGLT2 inhibitor activates the STING/IRF3/IFN-β pathway and induces immune infiltration in osteosarcoma. Cell Death Dis. https://doi.org/10.1038/S41419-022-04980-W
Wu X, Gu Z, Chen Y, Chen B, Chen W, Weng L, Liu X (2019) Application of PD-1 blockade in cancer immunotherapy. Comput Struct Biotechnol J 17:661–674. https://doi.org/10.1016/J.CSBJ.2019.03.006
Xie Z, Wang F, Lin L, Duan S, Liu X, Li X, Li T, Xue M, Cheng Y, Ren H, Zhu Y (2020) An SGLT2 inhibitor modulates SHH expression by activating AMPK to inhibit the migration and induce the apoptosis of cervical carcinoma cells. Cancer Lett 495:200–210. https://doi.org/10.1016/J.CANLET.2020.09.005
Xu C, Wang W, Zhong J, Lei F, Xu N, Zhang Y, Xie W (2018) Canagliflozin exerts anti-inflammatory effects by inhibiting intracellular glucose metabolism and promoting autophagy in immune cells. Biochem Pharmacol 152:45–59. https://doi.org/10.1016/J.BCP.2018.03.013
Yamada T, Sakaguchi K, Okada Y, Miura H, Otowa-Suematsu N, So A, Komada H, Hirota Y, Ohara T, Kuroki Y, Hara K, Matsuda T, Kishi M, Takeda A, Yokota K, Tamori Y, Ogawa W (2021) Analysis of time-dependent alterations of parameters related to erythrocytes after ipragliflozin initiation. Diabetol Int 12:197–206. https://doi.org/10.1007/S13340-020-00474-2/FIGURES/5
Yang WS, Sriramaratnam R, Welsch ME, Shimada K, Skouta R, Viswanathan VS, Cheah JH, Clemons PA, Shamji AF, Clish CB, Brown LM, Girotti AW, Cornish VW, Schreiber SL, Stockwell BR (2014) Regulation of ferroptotic cancer cell death by GPX4. Cell 156:317–331. https://doi.org/10.1016/j.cell.2013.12.010
Yaribeygi H, Atkin SL, Butler AE, Sahebkar A (2019) Sodium–glucose cotransporter inhibitors and oxidative stress: an update. J Cell Physiol 234:3231–3237. https://doi.org/10.1002/JCP.26760
Yaribeygi H, Maleki M, Nasimi F, Butler AE, Jamialahmadi T, Sahebkar A (2022) Sodium–glucose co-transporter 2 inhibitors and hematopoiesis. J Cell Physiol 237:3778–3787. https://doi.org/10.1002/JCP.30851
Ye J, Huang Y, Que B, Chang C, Liu W, Hu H, Liu L, Shi Y, Wang Y, Wang M, Zeng T, Zhen W, Xu Y, Shi L, Liu J, Jiang H, Ye D, Lin Y, Wan J, Ji Q (2018) Interleukin-12p35 knock out aggravates doxorubicin-induced cardiac injury and dysfunction by aggravating the inflammatory response, oxidative stress, apoptosis and autophagy in mice. EBioMedicine 35:29–39. https://doi.org/10.1016/J.EBIOM.2018.06.009
Ye J, Wang Y, Xu Y, Wang Z, Liu L, Wang M, Ye D, Zhang J, Yang Z, Lin Y, Ji Q, Wan J (2020) Interleukin-22 deficiency alleviates doxorubicin-induced oxidative stress and cardiac injury via the p38 MAPK/macrophage/Fizz3 axis in mice. Redox Biol 36:101636. https://doi.org/10.1016/J.REDOX.2020.101636
Yen HC, Oberley TD, Vichitbandha S, Ho YS, St. Clair DK (1996) The protective role of manganese superoxide dismutase against adriamycin-induced acute cardiac toxicity in transgenic mice. J Clin Invest 98:1253–1260. https://doi.org/10.1172/JCI118909
Yin J, Guo J, Zhang Q, Cui L, Zhang L, Zhang T, Zhao J, Li J, Middleton A, Carmichael PL, Peng S (2018) Doxorubicin-induced mitophagy and mitochondrial damage is associated with dysregulation of the PINK1/parkin pathway. Toxicol In Vitro 51:1–10. https://doi.org/10.1016/J.TIV.2018.05.001
Yurista SR, Silljé HHW, Oberdorf-Maass SU, Schouten EM, Pavez Giani MG, Hillebrands JL, van Goor H, van Veldhuisen DJ, de Boer RA, Westenbrink BD (2019) Sodium–glucose co-transporter 2 inhibition with empagliflozin improves cardiac function in non-diabetic rats with left ventricular dysfunction after myocardial infarction. Eur J Heart Fail 21:862–873. https://doi.org/10.1002/EJHF.1473
Zannad F, Ferreira JP, Pocock SJ, Anker SD, Butler J, Filippatos G, Brueckmann M, Ofstad AP, Pfarr E, Jamal W, Packer M (2020) SGLT2 inhibitors in patients with heart failure with reduced ejection fraction: a meta-analysis of the EMPEROR-reduced and DAPA-HF trials. The Lancet 396:819–829. https://doi.org/10.1016/S0140-6736(20)31824-9
Zelniker TA, Bonaca MP, Furtado RHM, Mosenzon O, Kuder JF, Murphy SA, Bhatt DL, Leiter LA, McGuire DK, Wilding JPH, Budaj A, Kiss RG, Padilla F, Gause-Nilsson I, Langkilde AM, Raz I, Sabatine MS, Wiviott SD (2020) Effect of dapagliflozin on atrial fibrillation in patients with type 2 diabetes mellitus: insights from the DECLARE-TIMI 58 trial. Circulation 141:1227–1234. https://doi.org/10.1161/CIRCULATIONAHA.119.044183
Zelniker TA, Braunwald E (2020) Mechanisms of cardiorenal effects of sodium–glucose cotransporter 2 inhibitors: JACC state-of-the-art review. J Am Coll Cardiol 75:422–434. https://doi.org/10.1016/J.JACC.2019.11.031
Zhang Y, Nakano D, Guan Y, Hitomi H, Uemura A, Masaki T, Kobara H, Sugaya T, Nishiyama A (2018) A sodium–glucose cotransporter 2 inhibitor attenuates renal capillary injury and fibrosis by a vascular endothelial growth factor-dependent pathway after renal injury in mice. Kidney Int 94:524–535. https://doi.org/10.1016/J.KINT.2018.05.002
Zhang Y, Zhang D, Meng T, Tian P, Chen J, Liu A, Zheng Y, Su G (2023) SGK1 is involved in doxorubicin-induced chronic cardiotoxicity and dysfunction through activation of the NFκB pathway. Int Immunopharmacol 125:111151. https://doi.org/10.1016/J.INTIMP.2023.111151
Zhao J, Zhang N, Ma X, Li M, Feng H (2023) The dual role of ferroptosis in anthracycline-based chemotherapy includes reducing resistance and increasing toxicity. Cell Death Discov 9:1–12. https://doi.org/10.1038/s41420-023-01483-1
Zhao L, Yang Y, Gao Q (2019) Efficacy and safety of nivolumab plus apatinib in advanced liver carcinosarcoma: a case report. Immunotherapy 11:651–656. https://doi.org/10.2217/IMT-2018-0214
Zhou J, Zhu J, Yu SJ, Ma HL, Chen J, Ding XF, Chen G, Liang Y, Zhang Q (2020) Sodium–glucose co-transporter-2 (SGLT-2) inhibition reduces glucose uptake to induce breast cancer cell growth arrest through AMPK/mTOR pathway. Biomed Pharmacother. https://doi.org/10.1016/J.BIOPHA.2020.110821
Zinman B, Wanner C, Lachin JM, Fitchett D, Bluhmki E, Hantel S, Mattheus M, Devins T, Johansen OE, Woerle HJ, Broedl UC, Inzucchi SE (2015) Empagliflozin, cardiovascular outcomes, and mortality in type 2 diabetes. N Engl J Med 373:2117–2128. https://doi.org/10.1056/NEJMoa1504720
Zullig LL, Sung AD, Khouri MG, Jazowski S, Shah NP, Sitlinger A, Blalock DV, Whitney C, Kikuchi R, Bosworth HB, Crowley MJ, Goldstein KM, Klem I, Oeffinger KC, Dent S (2022) Cardiometabolic comorbidities in cancer survivors: JACC: CardioOncology State-of-the-Art Review. JACC CardioOncol 4:149–165. https://doi.org/10.1016/J.JACCAO.2022.03.005
Acknowledgements
Figures created with Biorender.com.
Funding
Open access funding provided by Università Cattolica del Sacro Cuore within the CRUI-CARE Agreement. This research received no external funding.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests
The authors declare no conflict of interest.
Ethical approval
The manuscript does not contain clinical studies or patient data.
Additional information
This article is part of the special issue “Cardio-Oncology”.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Camilli, M., Viscovo, M., Maggio, L. et al. Sodium–glucose cotransporter 2 inhibitors and the cancer patient: from diabetes to cardioprotection and beyond. Basic Res Cardiol (2024). https://doi.org/10.1007/s00395-024-01059-9
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00395-024-01059-9