
Abstract
Crown-of-thorns seastar (CoTS, Acanthaster cf. solaris) outbreaks remain a significant cause of coral loss on the Great Barrier Reef (GBR) and across the West-Pacific Ocean. Previous outbreaks on the GBR have only been discovered once fully established, which constrains opportunities for effective control. Early detection of outbreaks would provide an important opportunity for early intervention and increase understanding of outbreak cause(s). Here, we assess the utility of environmental DNA (eDNA) monitoring to detect the initiation of a population outbreak at Lizard Island over five years (2019–2023), compared with density estimates obtained using Scooter-Assisted Large Area Diver-based (SALAD) surveys. At each of the five eDNA sampling sites, 30 replicate samples were collected annually and analysed with CoTS-specific primer sets and digital droplet PCR. Both methods detected distinct increases in CoTS densities from 2020/21 onwards, indicating the start of a new population outbreak. A large part of the observed variation in eDNA (expressed as the percentage of positive samples) was explained by changes in recorded CoTS density, confirming that eDNA data provide a quantitative estimate for adult CoTS abundance. SALAD surveys and eDNA are new and complementary monitoring methods that facilitate early detection of CoTS outbreaks, which will enable more effective management intervention.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Acanthaster spp. are corallivorous seastar species distributed throughout the tropical Indo-Pacific (Uthicke et al. 2024). Several of these species exhibit episodic population explosions (‘outbreaks’, ‘irruptions’) leading to significant coral loss. Population outbreaks of the western Pacific CoTS, Acanthaster cf. solaris, have occurred in French Polynesia (Kayal et al. 2012), Indonesia (Baird et al. 2013), Australia’s Great Barrier Reef (Moran 1986; Pratchett 2005), and Okinawa, Japan (Nakamura et al. 2014). Similarly, outbreaks of A. planci and A. mauritiensis have caused substantial coral loss in the Indian Ocean (summarised in Uthicke et al. 2024).
The Great Barrier Reef (GBR) has been subjected to four distinct waves of outbreaks of A. cf. solaris since the 1960s (Pratchett et al. 2014, 2017), with early stages of a 5th outbreak recently reported (Chandler et al. 2023) in the northern sections of the GBR. Outbreak dynamics and causes are still poorly understood (Pratchett et al. 2017), partly because outbreaks are detected only when CoTS densities are already at elevated levels (Babcock et al. 2020). The standard method used to monitor densities of CoTS on the GBR is manta tows (Miller et al. 2019), where a snorkeler records the number of CoTS sighted while being towed behind a tender vessel. While useful for detecting the occurrence of established outbreaks (Vanhatalo et al. 2017), manta tows have limited detectability, particularly at low to moderate densities, due to the often cryptic behaviour of juveniles and adults (Fernandes et al. 1990). Effective surveillance methods which are capable of monitoring all stages of an outbreak, in particular the earliest stages of population build-up, are critical for understanding the cause(s) of outbreaks and will allow for early intervention (Pratchett et al. 2017; Chandler et al. 2023).
Environmental DNA (eDNA) monitoring represents an alternative method for detecting low densities of organisms in aquatic environments (Ficetola et al. 2008), defined as genetic material obtained from environmental samples (i.e. soil, sediment, water column) (Taberlet et al. 2012). Environmental DNA approaches can be divided in two methods. First, metabarcoding using general primers can describe species presence and diversity in different communities. Second, species specific primers can be used to detect and to some extent quantify individual species. In freshwater environments, eDNA was applied to detect introduced species (e.g. Jerde et al. 2013; Rees et al. 2014; Fukumoto et al. 2015; Hunter et al. 2017) or to describe community composition of plants (Espinosa Prieto et al. 2023), fishes (Ito et al. 2023) or invertebrates (Coghlan et al. 2021).
Quantifying eDNA in marine environments is challenging due to dilution factors and environmental conditions, including currents (Uthicke et al. 2022). However, an increasing number of studies have effectively applied eDNA-based methods to the marine realm. Metabarcoding has been used in temperate and arctic seas (Thomsen et al. 2016; Yamamoto et al. 2017; Jeunen et al. 2019), but also in coral reef environments (Dugal et al. 2022, 2023; Marwayana et al. 2022). Species-specific primer approaches in marine environments have also now been used for several marine taxa, including crustaceans (Forsström and Vasemägi 2016), molluscs (Mauvisseau 2017), echinoderms (Uthicke et al. 2018; Ellis et al. 2021), cnidarians (Minamoto et al. 2017) and cubozoans (Morrissey et al. 2022).
CoTS specific PCR primers were initially developed to quantify planktonic larvae (Uthicke et al. 2015; Doyle et al. 2017). The same primers have also been employed to detect and quantify post-settlement CoTS from high to very low densities using digital PCR (Uthicke et al. 2018, 2022; Kwong et al. 2021) or lateral flow assays (Doyle and Uthicke 2020). In addition, these DNA-based methods have recently been tested to detect CoTS recruits in artificial settlement collectors (Doll et al. 2021).
The purpose of this study was to assess the utility of eDNA to detect changes in local densities of CoTS within reef environments, which may provide early warning of the onset of new or renewed population outbreaks. We conducted eDNA sampling over five consecutive years (2019—2023) to test for inter-annual changes in densities of CoTS at Lizard Island in the Northern GBR (14.7°S, 145.5°E). Lizard Island is located within the putative “initiation box” of CoTS outbreaks and has been a key location for previous studies on the population dynamics and impacts of CoTS (Pratchett 2005, 2010; Caballes et al. 2021). To test how changes in eDNA compare to direct density estimates, we also undertook extensive in-water surveys at corresponding sites, using a new method designed to provide high-resolution abundance data, even at low to moderate density levels (Chandler et al. 2023). Here, we directly compare the eDNA data with density estimates from the in-water surveys to corroborate the application of eDNA for effective CoTS population monitoring in a temporal context.
Methods
Field collection
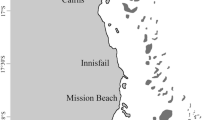
Environmental DNA sampling was undertaken at five sites around Lizard Island between August and September in each year from 2019 until 2023 (Table 1). Sites were selected to allow access in most weather conditions from the Lizard Island Research Station (Fig. 1, Table 1). To account for any possible minor effect of water residence or tide (Uthicke et al. 2022), on most occasions, two collection events separated by 1–5 d were undertaken for each site/year combination. A total of 30 replicates (15 per collection event) were conducted at each site/year combination with the exception being in 2020 when 31 and 29 samples were collected from Big Vicki’s Reef and the Lagoon site, respectively. A subset of samples from the first two sampling years were included in a previous publication (Uthicke et al. 2022).

Map of North-East Australian and the Great Barrier Reef. Insert: Location of eDNA sampling sites (red dots) and complementary SALAD survey areas (red dashed boxes) at Lizard Island, northern Great Barrier Reef
Scooter-Assisted Large Area Diver-based (SALAD) surveys were conducted as described in Chandler et al. (2023). Data presented here were collected over 5 consecutive years (2019–2023) at complementary study sites to allow direct comparison to eDNA data. The mean annual location-wide density was estimated using SALAD data from across the north-western side of Lizard Island (Mermaid Cove to Palfrey Island), while corresponding site-level density estimates were calculated for each sampling year using data from the vicinity of eDNA sites (Big Vicki’s Reef, Casuarina Beach, Clam Gardens and Mermaid Cove). No comparative SALAD data exist for the eDNA Lagoon site. SALAD data collected in 2019–2022 were included in Chandler et al. (2023). Data presented here are based on actual observations of CoTS (i.e., recorded densities), as opposed to inferred density estimates that include distinct sets of feeding scars where the CoTS was not detected.
DNA extraction and digital droplet PCR assay
Water samples for eDNA analyses were collected as previously described (Uthicke et al. 2022). Briefly, 2.5lt water was pumped directly from the ocean through an eDNA housing (Smith-Root, USA) containing a 1.2 mm mixed cellulose ester membrane using an eDNA sampling device (Grover-Pro™, Grover Scientific, Townsville Australia). Membrane filters were removed from the housings within 2 h of collection, folded carefully into eighths using sterilised tweezers and placed in a 1.5-ml screw cap tube containing 540 µl of Qiagen buffer ATL as a preservative (Majaneva et al. 2018). All sampling equipment was bleach-cleaned with a concentrated chlorine solution (100 g dichloroisocyanuric acid per 20 L, equivalent to 0.275% w/v available chlorine). Filters were extracted using a Qiagen Blood and Tissue DNeasy kit on a Qiacube automated nucleic extraction instrument following the method described in Doyle and Uthicke (2020). All samples were eluted in 50 µl of a 10 × diluted TE buffer (TE0.1) except for the 2019 samples which were eluted in 2 × 50 µl TE0.1. Digital droplet PCR (ddPCR) was conducted as per Uthicke et al. (2018) in the absence of restriction enzyme. For 2019 and 2020 samples, a manual QX200 droplet generator was used, whereas for subsequent samples, an automated droplet generator was employed (Bio-Rad, QX200 AutoDG). Accurate calling of positive detection in ddPCR includes quality control of each ddPCR reaction. This includes running positive and negative controls (see below) to confirm expected distribution of droplet fluorescence for each microplate run, rejecting a ddPCR reaction if the accepted droplet count is less than 10,000 (https://www.bio-rad.com/webroot/web/pdf/lsr/literature/Bulletin_6407.pdf) to ensure consistent quantification (independent of droplet generation method), and consistent thresholding for determination of positive and negative droplets. Controls were included at all levels of sample handling (Table 2), namely ‘No Template Controls’ (NTC), laboratory extraction controls (blank filters extracted alongside samples) and field controls (MilliQ water filtered on site). No positive detections were observed in any NTC, or other controls from 2019, 2022 to 2023. A small number of false positive droplets were observed in 2020 and 2021 (Table 2). Although the small proportion of false positives was unlikely to affect the outcome, we randomly removed positive droplets from positive PCR replicates in those years, based on the proportion of false positives in the pooled field and extraction controls.
Statistical analyses
For CoTS eDNA, we previously analysed two separate metrics (Uthicke et al. 2022). The actual DNA concentration (Acanthaster cf. solaris mitochondrial DNA copy numbers) works at intermediate to high concentrations and was not suitable to track the build-up of low-density populations in the present dataset. Hence, these data were not formally analysed, but a summary graph is presented. The second metric is binomial, scoring each individual sample as positive or negative for the presence of CoTS eDNA. Binomial results in Uthicke et al. (2022) were analysed using an occupancy approach. Here, we simplified this method and analyse these data as the proportion of positive samples to allow for more appropriate accounting of error structure in this larger dataset. Thus, we used a binomial model with ‘Year’ and ‘Site’ as fixed factors. The ‘Sampling Occasion’ (15 filters at 2 occasions per year at each site) was included as a random variable during initial model runs but explained low amounts of variance and models did not converge. This factor was thus excluded from the model. The density data obtained from SALAD surveys were analysed in a linear model including ‘Year’ and ‘Site’ as fixed factors. eDNA and SALAD samples were compared using a linear model with ‘CoTS density’ (SALAD estimates, log transformed) and ‘Site’ as factors explaining variation in the proportion of samples positive for CoTS eDNA. The interaction term in the latter model was insignificant and removed from the final model. All models fitted (using the lme4 library) complied well with assumptions, as tested in the DHArma library. All analysis and graphs were done in the R environment (R-Core-Team 2018).
Results
A total 750 eDNA samples were collected over five years (2019–2023) at five sites around Lizard Island, of which, 364 samples were positive (i.e. above detection limit) for CoTS mtDNA. A binomial linear model of the presence–absence data exhibited significant effects of both year (χ2 = 126.21, DF = 4, p < 0.0001) and sampling site (χ2 = 125.83, DF = 4, p < 0.0001). However, the interaction of both factors was also significant (χ2 = 57.22, DF = 16, p < 0.0001), suggesting that trends were not parallel across all sites.
Averaged across all sites, the proportion of positive samples distinctly increased from year to year (Fig. 2A, factor Year). Low proportions of positive samples (p < 0.30) were observed in the first two sampling years, but the proportion of positive samples increased past 0.50 in 2021. Values in the final year were the highest observed and significantly (based on non-overlapping confidence intervals) above those of all other years. Similarly, the average mitochondrial DNA copy numbers (not formerly analysed) for all sites were close to zero within the first 2–3 y of this study, with averages distinctly increasing from 2021 onwards at most sites (Fig. 3).

The proportion of positive samples for Acanthaster cf. solaris eDNA on Lizard Island, A averaged for five years (Factor “Year”, N = 150) and B separated by sampling site for each year (Factor “Year x Site”, N = 30). Coloured dots represent model fit and vertical bars represent the lower and upper bounds of the 95% confidence interval

Average copy number of Acanthaster cf. solaris eDNA on Lizard Island, separate for five sampling sites. Vertical bars represent the lower and upper bounds of the 95% confidence interval, based on bootstrapping
Overall, there were several distinct differences in the proportion of positive samples over the years among sampling sites (Year x Site interaction). The Lagoon site generally exhibited the lowest values, with the proportion of positive samples only slightly increasing in 2023 (Fig. 2B). Casuarina Beach, Clam Garden and Mermaid Cove follow the same trend as the Island-wide averages with continuously increasing proportion of positive samples between 2019 and 2023. The proportion of positive samples at Big Vicki’s Reef was already around 0.50 at the beginning of the study and increased further past 2021.
CoTS densities recorded using SALAD surveys, averaged over the North-Western sites (Big Vicki’s Reef, Casuarina Beach, Clam Gardens, Mermaid Cove) of Lizard Island, differed among sampling years (χ2 = 64.047, DF = 4, p = < 0.0001; Fig. 4A). Differences in CoTS densities among sites were also significant (linear model, χ2 = 8.020, DF = 3, p = 0.0456), but the interaction term was not significant (χ2 = 15.455, DF = 12, p = 0.2175). Densities recorded were relatively low (< 3 ha−1) in 2019 and 2020. Similar to the eDNA data, average CoTS densities increased substantially from 2020 to 2021 and were distinctly higher again (near 10 Ind. ha−1) in 2022 and 2023 (Fig. 4A). Significant differences between sites were likely driven by somewhat higher overall values at Clam Gardens (Fig. 4B).

Densities of Acanthaster cf. solaris recorded with SALAD surveys A at Lizard Island over 5 sampling years (2019–2023) and B at four sampling locations, averaged across years. Coloured dots represent model fit (factor ‘Year’ and factor ‘Site’) and vertical bars represent the lower and upper bounds of the 95% confidence interval
Linear model analysis indicated a significant effect of recorded CoTS density on measured eDNA, with the overall model explaining 51% of the variation (Tab. 3). Thus, increasing proportions of positive eDNA samples were well explained by (log transformed) CoTS densities detected using the SALAD surveys (Fig. 5).

Linear model fit (Table 3) of the proportion of positive eDNA samples versus recorded densities of A. cf. solaris using the SALAD method. The blue shaded area depicts the 95% confidence interval, the black line the model fit. Red dots are average observations per site and year
Discussion
The primary objective of this study was to assess whether eDNA sampling can resolve inter-annual changes at low densities of CoTS. This is a critical component for achieving early detection and has previously only been possible with high-resolution in-water surveys (Chandler et al. 2023). The proportion of samples with positive detections for CoTS eDNA increased across all sites over the 5y study period and corresponds well with variation in CoTS densities recorded using in-water SALAD surveys. Over the same time, manta tow surveys (traditional monitoring method) conducted around Lizard Island detected no such temporal variation and generally reported low CoTS densities, below the relevant threshold of an outbreak, in each of the sampling years (≤ 0.01 CoTS per tow; AIMS LTMP, available at: https://apps.aims.gov.au/reef-monitoring/reef/Lizard%20Isles/manta). In contrast, both sets of monitoring data presented here report substantial and rapid (densities more than tripled and the proportion of positive eDNA samples more than doubled between 2 years) population growth at this key location, indicating the onset of a 5th CoTS outbreak on the GBR since the 1960s. The results from the present study clearly demonstrate the utility and necessity for novel sampling methods to detect the early onset of population outbreaks of CoTS.
Based on seemingly consistent periodicity of CoTS outbreaks on the GBR since the 1960s, renewed outbreaks were expected in 2025–2027 (Babcock et al. 2020). Our data suggest that these anticipated population outbreaks have already commenced, with clear and consistent increases in densities of CoTS at Lizard Island since 2020 (see also Chandler et al. 2023). However, the methods presented herein provide unprecedented capacity to detect the initiation of population outbreaks and protracted build-up of CoTS densities over several years. The initiation of past outbreaks has been reported based on detection of highly elevated CoTS densities (e.g. Wooldridge and Brodie 2015), which largely fails to recognise or detect the protracted build-up in CoTS populations that may often precede the initial establishment of outbreaks (Pratchett 2005). It appears the build-up of the new outbreak occurs, while the previous outbreak is still underway in parts of the central and southern GBR. Thus, an alternative explanation for the marked temporal patterns observed in this study may also be that elevated densities at Lizard Island represent a flare-up of the 4th outbreak in the northern GBR. At present it is unresolved if current strong anthropogenic intervention (culling) could cause a flare up or create conditions for permanent outbreaks.
The broad size range of CoTS at Lizard Island, presented in Chandler et al. (2023), suggests that increasing densities resulted from the accumulation of seastar over several successive years of successful recruitment. Notably, CoTS densities (< 3 Ind. ha−1) recorded in the first years of the present study (2019 and 2020) were below those expected to contribute to accelerated reproductive success (Rogers et al. 2017). It is uncertain if these relatively low densities constitute natural baseline levels at this location or were already an indication of CoTS population build-up. In addition, density estimates based solely on the direct observation of CoTS specimens during SALAD surveys are likely still an underestimate of CoTS abundance. Inferred densities, including observations of distinct sets of feeding scars where the CoTS was not detected, are generally higher and present a more realistic estimate of population size (Chandler et al. 2023). It is also unknown to what extent sustained and ongoing recruitment may be driven by continual input of larvae from reefs to the north (Luick et al. 2007) and/or retention of larvae produced by increasing local populations. Nevertheless, considering the substantial increase in CoTS densities documented here from 2021 onwards, the reproductive capacity of this population is rapidly building and may reinforce population growth around Lizard Island and contribute to the establishment and spread of population outbreaks.
The eDNA analysis presented here also detected some spatial differences among sampling sites at Lizard Island. Particularly low proportions of positive eDNA samples were collected from the Lagoon site, which was not included in the SALAD surveys but had reported low CoTS densities during a previous outbreak (Pratchett 2005). Another difference among sites in the eDNA dataset was the relatively high proportion of positive samples from Big Vicki’s Reef, even before this population build-up was detected in the SALAD surveys. It is possible that eDNA at that location was influenced by currents and other environmental factors. However, water residence time, sea temperature and water depth explained only a small amount of variance in CoTS eDNA concentration (Uthicke et al. 2022).
eDNA detection corresponded well with SALAD-based estimates along this low to moderate density gradient, with the overall model explaining more than 50% of the total variation. These insights further corroborate and compliment previous demonstrations of eDNA sampling in areas of higher density populations, compared to 200m2 belt transects (Uthicke et al. 2018, 2022). Similar to Uthicke et al. (2022), we found that a simplified metric (proportion of positive samples) effectively represents changing densities of CoTS better than average mitochondrial DNA copy numbers, although an increase in the latter metric was also detectable in the last years of the present study.
Both SALAD surveys and eDNA detect CoTS at low and moderate densities during the early stages of outbreaks, and given inherent differences in methodological approaches, requirements and scalability, these two methods are complementary tools for early warning and intervention efforts. Key advantages of the eDNA method are that field sampling is fast (e.g. < 2 h are needed to survey a reef on three sites, Uthicke, Doyle, Gomez Cabrera, unpublished data) and overcomes some of the inherent time, bias, risk and training limitations associated with in-water surveys, underlining its application for wide-sweeping monitoring of potentially hundreds of reefs. In addition, eDNA sampling is particularly useful for sampling in locations where in-water sampling is not safe (e.g. due to the presence of saltwater crocodiles) or practicable. The laboratory-based processing of numerous samples can take several weeks; however, eDNA samples can be securely stored and subsequently analysed for other species groups or biodiversity assessments. An additional disadvantage is that the sensitivity of ddPCR does come at higher price per sample compared to standard PCR or even real-time PCR. While there may be cost-prohibitive aspects of the current eDNA method, we have demonstrated proof-of-concept alternatives that, with appropriate development, may provide an entry point for less developed nations to adopt a similar monitoring program (Doyle and Uthicke 2020). Although eDNA monitoring will add cost to existing surveys (in the range of 1-2 k A$ per reef), ship time is usually up to 1 order of magnitude more expensive, hence the relative increase of costs is small. In addition, field methods for eDNA collection are easy and rangers and ‘citizen scientists’ can be trained to collect samples. This will overall reduce costs for eDNA surveys by using ships of opportunity for sampling.
By contrast, SALAD surveys take much longer to complete, but data are instantaneously available. In addition, those surveys also yield fine-scale data on additional variables such behaviour and population structure of CoTS (all recorded CoTS are measured) and provide environmental context by recording benthic data and other disturbances (Chandler et al. 2023). Thus, an effective and efficient use of both methods would be to conduct large-scale eDNA monitoring over many reefs and focus SALAD surveys on ecologically important locations and selected reefs that recently reported high and/or increasing proportions in positive eDNA samples.
Conclusions
The eDNA and SALAD methods utilised here are important, and complementary, new tools enabling early detection of CoTS population increases, thereby allowing for early and effective intervention through targeted population control. Moreover, early detection of population outbreaks and the location of these is crucial to improve understanding of the factor(s) that contribute to the initiation of recurrent outbreaks on the GBR. For instance, to evaluate whether recruitment enhancement through increased runoff (Birkeland 1982; Brodie et al. 2005) or oceanographic events (Wooldridge and Brodie 2015) play a significant role in the initial population build-up, it is important to pinpoint the start of these outbreaks in time and space. Similarly, to understand the role of predation on juvenile (Balu et al. 2021; Desbiens et al. 2023) and adult CoTS (Cowan et al. 2020; Caballes et al. 2022), it is necessary to quantify predator densities and understand predation pressure on reefs where CoTS populations begin and continue to build, as opposed to reefs with well-established populations of adults.
References
Babcock RC, Plaganyi EE, Condie SA, Westcott DA, Fletcher CS, Bonin MC, Cameron D (2020) Suppressing the next crown-of-thorns outbreak on the Great Barrier Reef. Coral Reefs 39:1233–1244. https://doi.org/10.1007/s00338-020-01978-8
Baird A, Pratchett M, Hoey A, Herdiana Y, Campbell S (2013) Acanthaster planci is a major cause of coral mortality in Indonesia. Coral Reefs 32:803–812
Balu V, Messmer V, Logan M, Hayashida-Boyles AL, Uthicke S (2021) Is predation of juvenile crown-of-thorns seastars (Acanthaster cf. solaris) by peppermint shrimp (Lysmata vittata) dependent on age, size, or diet? Coral Reefs 40:641–649
Birkeland C (1982) Terrestrial runoff as a cause of outbreaks of Acanthaster planci (Echinodermata: Asteroidea). Mar Biol 69:175–185
Brodie J, Fabricius K, De’ath G, Okaji K (2005) Are increased nutrient inputs responsible for more outbreaks of crown-of-thorns starfish? An appraisal of the evidence. Mar Pollut Bull 51:266–278
Caballes CF, Byrne M, Messmer V, Pratchett MS (2021) Temporal variability in gametogenesis and spawning patterns of crown-of-thorns starfish within the outbreak initiation zone in the northern Great Barrier Reef. Mar Biol 168:13. https://doi.org/10.1007/s00227-020-03818-3
Caballes CF, Messmer V, Raymundo ML, Pratchett MS (2022) Prevalence and severity of sublethal injuries in crown-of-thorns starfish relative to marine reserves in the Great Barrier Reef. Aquat Conserv-Mar Freshw Ecosyst 32:993–1004. https://doi.org/10.1002/aqc.3762
Chandler JF, Burn D, Caballes CF, Doll PC, Kwong SL, Lang BJ, Pacey KI, Pratchett MS (2023) Increasing densities of Pacific crown-of-thorns starfish (Acanthaster cf. solaris) at Lizard Island, northern Great Barrier Reef, resolved using a novel survey method. Sci Rep 13:19306
Coghlan SA, Currier CA, Freeland J, Morris TJ, Wilson CC (2021) Community eDNA metabarcoding as a detection tool for documenting freshwater mussel (Unionidae) species assemblages. Environ DNA 3:1172–1191
Cowan Z-L, Ling SD, Caballes CF, Dworjanyn SA, Pratchett MS (2020) Crown-of-thorns starfish larvae are vulnerable to predation even in the presence of alternative prey. Coral Reefs 39:293–303. https://doi.org/10.1007/s00338-019-01890-w
Desbiens AA, Mumby PJ, Dworjanyn S, Plagányi ÉE, Uthicke S, Wolfe K (2023) Novel rubble-dwelling predators of herbivorous juvenile crown-of-thorns starfish (Acanthaster sp.). Coral Reefs 42:579–591
Doll PC, Messmer V, Uthicke S, Doyle JR, Caballes CF, Pratchett MS (2021) DNA-based detection and patterns of larval settlement of the corallivorous crown-of-thorns sea star (Acanthaster sp.). Biol Bull 241:271–285
Doyle J, Uthicke S (2020) Sensitive environmental DNA detection via lateral flow assay (dipstick)—A case study on corallivorous crown-of-thorns sea star (Acanthaster cf. solaris) detection. Environmental DNA 3:323–342
Doyle JR, McKinnon AD, Uthicke S (2017) Quantifying larvae of the coralivorous seastar Acanthaster cf. solaris on the Great Barrier Reef using qPCR. Mar Biol 164:176. https://doi.org/10.1007/s00227-017-3206-x
Dugal L, Thomas L, Wilkinson SP, Richards ZT, Alexander JB, Adam AA, Kennington WJ, Jarman S, Ryan NM, Bunce M (2022) Coral monitoring in northwest Australia with environmental DNA metabarcoding using a curated reference database for optimized detection. Environmental DNA 4:63–76
Dugal L, Thomas L, Meenakshisundaram A, Simpson T, Lines R, Colquhoun J, Jarman S, Meekan M (2023) Distinct coral reef habitat communities characterized by environmental DNA metabarcoding. Coral Reefs 42:17–30
Ellis MR, Clark ZS, Treml EA, Brown MS, Matthews TG, Pocklington JB, Stafford-Bell RE, Bott NJ, Nai YH, Miller AD (2021) Detecting marine pests using environmental DNA and biophysical models. Sci Tot Environ 816:151666
Espinosa Prieto A, Beisel JN, Verschuren P, Hardion L (2023) Toward freshwater plant diversity surveys with eDNA barcoding and metabarcoding. Environ DNA 5(4):648–670
Fernandes L, Marsh H, Moran P, Sinclair D (1990) Bias in manta tow surveys of Acanthaster planci. Coral Reefs 9:155–160
Ficetola GF, Miaud C, Pompanon F, Taberlet P (2008) Species detection using environmental DNA from water samples. Biol Let 4:423–425
Forsström T, Vasemägi A (2016) Can environmental DNA (eDNA) be used for detection and monitoring of introduced crab species in the Baltic Sea? Mar Pollut Bull 109:350–355. https://doi.org/10.1016/j.marpolbul.2016.05.054
Fukumoto S, Ushimaru A, Minamoto T (2015) A basin-scale application of environmental DNA assessment for rare endemic species and closely related exotic species in rivers: a case study of giant salamanders in Japan. J Appl Ecol 52:358–365
Hunter ME, Dorazio RM, Butterfield JS, Meigs-Friend G, Nico LG, Ferrante JA (2017) Detection limits of quantitative and digital PCR assays and their influence in presence–absence surveys of environmental DNA. Mol Ecol Resour 17:221–229
Ito G, Yamauchi H, Shigeyoshi M, Ashino K, Yonashiro C, Asami M, Goto Y, Duda JJ, Yamanaka H (2023) Using eDNA metabarcoding to establish targets for freshwater fish composition following river restoration. Global Ecol Conserv 43:e02448
Jerde CL, Chadderton WL, Mahon AR, Renshaw MA, Corush J, Budny ML, Mysorekar S, Lodge DM (2013) Detection of Asian carp DNA as part of a Great Lakes basin-wide surveillance program. Can J Fish Aquat Sci 70:522–526
Jeunen GJ, Knapp M, Spencer HG, Lamare MD, Taylor HR, Stat M, Bunce M, Gemmell NJ (2019) Environmental DNA (eDNA) metabarcoding reveals strong discrimination among diverse marine habitats connected by water movement. Mol Ecol Resour 19:426–438
Kayal M, Vercelloni J, Lison de Loma T, Bosserelle P, Chancerelle Y, Geoffroy S, Stievenart C, Michonneau F, Penin L, Planes S, Adjeroud M (2012) Predator crown-of-thorns starfish (Acanthaster planci) outbreak, mass mortality of corals, and cascading effects on reef fish and benthic communities. PLoS ONE 7:e47363. https://doi.org/10.1371/journal.pone.0047363
Kwong SLT, Villacorta-Rath C, Doyle J, Uthicke S (2021) Quantifying shedding and degradation rates of environmental DNA (eDNA) from Pacific crown-of-thorns seastar (Acanthaster cf. solaris). Mar Biol 168:85. https://doi.org/10.1007/s00227-021-03896-x
Luick JL, Mason L, Hardy T, Furnas MJ (2007) Circulation in the Great Barrier Reef Lagoon using numerical tracers and in situ data. Cont Shelf Res 27:757–778
Majaneva M, Diserud OH, Eagle SH, Boström E, Hajibabaei M, Ekrem T (2018) Environmental DNA filtration techniques affect recovered biodiversity. Sci Rep 8:1–11
Marwayana ON, Gold Z, Meyer CP, Barber PH (2022) Environmental DNA in a global biodiversity hotspot: lessons from coral reef fish diversity across the Indonesian archipelago. Environmental DNA 4:222–238
Mauvisseau Q (2017) On the way for detecting and quantifying elusive species in the sea: the Octopus vulgaris case study. Fish Res 191:41–48. https://doi.org/10.1016/j.fishres.2017.02.023
Miller IR, Jonker M, Coleman G (2019) Crown-of-thorns starfish and coral surveys using the manta tow technique. Long-term monitoring of the Great Barrier Reef standard operational procedure 9:1–41
Minamoto T, Fukuda M, Katsuhara KR, Fujiwara A, Hidaka S, Yamamoto S, Takahashi K, Masuda R (2017) Environmental DNA reflects spatial and temporal jellyfish distribution. PLoS ONE 12:e0173073
Moran P (1986) The Acanthaster phenomenon. Oceanogr Mar Biol Ann Rev 24:379–480
Morrissey SJ, Jerry DR, Kingsford MJ (2022) Genetic detection and a method to study the ecology of deadly cubozoan jellyfish. Diversity 14:1139
Nakamura M, Okaji K, Higa Y, Yamakawa E, Mitarai S (2014) Spatial and temporal population dynamics of the crown-of-thorns starfish, Acanthaster planci, over a 24-year period along the central west coast of Okinawa Island, Japan. Mar Biol 161:2521–2530
Pratchett MS (2005) Dynamics of an outbreak population of Acanthaster planci at Lizard Island, northern Great Barrier Reef (1995–1999). Coral Reefs 24:453–462
Pratchett MS (2010) Changes in coral assemblages during an outbreak of Acanthaster planci at Lizard Island, northern Great Barrier Reef (1995–1999). Coral Reefs 29:717–725. https://doi.org/10.1007/s00338-010-0602-9
Pratchett M, Caballes CF, Rivera-Posada J, Sweatman H (2014) Limits to understanding and managing outbreaks of crown-of-thorns starfish (Acanthaster spp.). Oceanogr Mar Biol Ann Rev 52:133–200
Pratchett M, Caballes C, Wilmes J, Matthews S, Mellin C, Sweatman H, Nadler L, Brodie J, Thompson C, Hoey J, Bos A, Byrne M, Messmer V, Fortunato S, Chen C, Buck A, Babcock R, Uthicke S (2017) Thirty years of research on crown-of-thorns starfish (1986–2016): Scientific advances and emerging opportunities. Diversity 9(4):41
R-Core-Team (2018) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Rees HC, Maddison BC, Middleditch DJ, Patmore JR, Gough KC (2014) The detection of aquatic animal species using environmental DNA–a review of eDNA as a survey tool in ecology. J Appl Ecol 51:1450–1459
Rogers JGD, Pláganyi ÉE, Babcock RC (2017) Aggregation, Allee effects and critical thresholds for the management of the crown-of-thorns starfish Acanthaster planci. Mar Ecol Prog Ser 578:99–114
Taberlet P, Coissac E, Hajibabaei M, Rieseberg LH (2012) Environmental DNA. Mol Ecol 21:1789–1793
Thomsen PF, Møller PR, Sigsgaard EE, Knudsen SW, Jørgensen OA, Willerslev E (2016) Environmental DNA from seawater samples correlate with trawl catches of subarctic, deepwater fishes. PLoS ONE 11:e0165252
Uthicke S, Doyle J, Duggan S, Yasuda N, McKinnon AD (2015) Outbreak of coral-eating Crown-of-Thorns creates continuous cloud of larvae over 320 km of the Great Barrier Reef. Sci Rep 5:16885. https://doi.org/10.1038/srep16885
Uthicke S, Lamare M, Doyle JR (2018) eDNA detection of corallivorous seastar (Acanthaster cf. solaris) outbreaks on the Great Barrier Reef using digital droplet PCR. Coral Reefs 37:1229–1239. https://doi.org/10.1007/s00338-018-1734-6
Uthicke S, Pratchett MS, Bronstein O, Alvarado JJ, Wörheide G (2024) The crown-of-thorns seastar species complex: knowledge on the biology and ecology of five corallivorous Acanthaster species. Mar Biol 171:32. https://doi.org/10.1007/s00227-023-04355-5
Uthicke S, Robson B, Doyle JR, Logan M, Pratchett MS, Lamare M (2022) Developing an effective marine eDNA monitoring: eDNA detection at pre-outbreak densities of corallivorous seastar (Acanthaster cf. solaris). Sci Tot Environ 851:158143
Vanhatalo J, Hosack GR, Sweatman H (2017) Spatio-temporal modelling of crown-of-thorns starfish outbreaks on the Great Barrier Reef to inform control strategies. Journal of Applied Ecology 54(1):188–197
Wooldridge SA, Brodie JE (2015) Environmental triggers for primary outbreaks of crown-of-thorns starfish on the Great Barrier Reef, Australia. Mar Pollut Bull 101:805–815. https://doi.org/10.1016/j.marpolbul.2015.08.049
Yamamoto S, Masuda R, Sato Y, Sado T, Araki H, Kondoh M, Minamoto T, Miya M (2017) Environmental DNA metabarcoding reveals local fish communities in a species-rich coastal sea. Sci Rep 7:40368
Acknowledgements
We are grateful to Drs. Emma Lawrence and Scott Foster for discussions on statistical models. This research was funded by the Ian Potter Foundation 50th Anniversary Commemorative Grants Scheme through the Australian Museum’s Lizard Island Research Station, and the CoTS Control Innovation Program (CCIP) administered by the Great Barrier Reef Foundation. We are grateful for ongoing support by Drs. Anne Hoggett and Lyle Vail, Directors of Lizard Island Research Station.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
Morgan Pratchett currently serves as Editor-in-Chief for Coral Reefs, but is recused from any role in the peer-review process for this manuscript. The authors declare no other conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Uthicke, S., Doyle, J.R., Gomez Cabrera, M. et al. eDNA monitoring detects new outbreak wave of corallivorous seastar (Acanthaster cf. solaris) at Lizard Island, Great Barrier Reef. Coral Reefs (2024). https://doi.org/10.1007/s00338-024-02506-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00338-024-02506-8