
Abstract
Gorgonians are a diverse and conspicuous component of coral reef ecosystems, providing habitat structure that supports unique assemblages of fishes and invertebrates. Evaluating their overall importance as ecological engineers requires an understanding of their spatial patterns of distribution, abundance and assemblage composition, and the biophysical factors that drive these patterns. No baseline data are available on the spatial patterns of distribution of gorgonians for the Great Barrier Reef. In this study, we quantified the abundance, genera richness, and composition of gorgonian assemblages using video surveys at three depths (5, 10, and 15 m) at 16 locations at the Palm Islands, an inshore island group in the central Great Barrier Reef. We compared gorgonian abundance and genera richness between depths and assessed the role of benthic habitat in structuring gorgonian communities. We also conducted a preliminary investigation of the potential role of water currents in driving gorgonian spatial patterns in the Palm Islands, using in situ current meters. Gorgonian abundance and genera richness consistently increased with depth, although the magnitude of the depth effect varied among locations. Abundance increased with increasing percent cover of rubble and conversely declined with increasing cover of hard corals. The composition of gorgonian assemblages also varied among depths, with whip (Junceella, Viminella) and fan (Acanthogorgia, Anthogorgia, Annella) growth forms being dominant at depths of 5 and 10 m, and branching (Dichotella, Icilogorgia) and candelabrum (Ctenocella) forms being dominant at 15 m. The shallow gorgonian assemblage was associated with high coral cover, whilst the deeper assemblage was associated with high per cent cover of rubble, turf and/or macroalgae. This study highlighted that the abundance, diversity, and composition of gorgonian assemblages on coral reefs in the Palm Islands are determined by a range of biophysical factors linked to depth. Further work is required to isolate the primary drivers of these depth-related effects and evaluate their relative importance.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Understanding the physical and biological drivers of the spatial patterns of key habitat-forming organisms on coral reefs is essential to evaluate their significance to reef ecosystems as a whole (Bellwood et al. 2019). Naturally, most attention has been given to scleractinian corals, as they are the building blocks of coral reefs, and in recent years, widespread declines have been reported as a result of multiple stressors (Pandolfi et al. 2003; Bellwood et al. 2019; Williams et al. 2019). Scleractinian corals are important ecosystem engineers, supporting many other reef-associated species such as fishes and invertebrates (Jones et al. 1994, 1997; Graham and Nash 2013). However, there are numerous other habitat-forming benthic organisms on coral reefs, including sponges, soft corals, and gorgonians, which also promote habitat diversity and spatial heterogeneity (Sponaugle 1991; Evans et al. 2011; Abeytia et al. 2013; Calcinai et al. 2013; Maggioni et al. 2020). These organisms can dominate the substratum on some coral reefs, contribute to the complex, three-dimensional structure of reef habitats, and support a broad range of unique fish and invertebrate species (Dinesen 1983; Jones et al. 1997; Diaz and Rützler 2001; Buhl-Mortensen et al. 2010; Grinyó et al. 2016). However, despite the potential importance of these taxa as ecosystem engineers, their spatial patterns in abundance, genera richness, and community structure, and the physical and biological drivers of these patterns have received much less attention (Dinesen 1983).
Gorgonians (phylum Cnidaria and subphylum Anthozoa) are part of the Octocorallia subclass and the Holaxonia–Alcyoniina clade (McFadden et al. 2022). They are a conspicuous and diverse component of coral reef habitats, with multiple growth forms, including candelabrum, fan, whip, branched, and bushy growth forms (Fabricius and Alderslade 2001). Their complex structures create a diverse range of microhabitats that provide resources for a large variety of fish and invertebrate species (Graham and Nash 2013). As passive suspension feeders, extracting nutrients from the water column, hydrodynamic environmental variables have significant influence on nutrient availability, in turn affecting multiple aspects of gorgonian biology, morphology, and distribution (Muzik and Wainwright 1977; Chang-Feng and Ming-Chao 1993; Al-Marayati and Edmunds 2018). For example, faster currents deliver larger quantities of nutrients to gorgonian colonies, enhancing feeding and growth rates (Rowley 2018). The ability to withstand high currents, however, varies among growth forms, and morphological adaptations have developed to allow colonies to benefit from increased nutrient supply. The influence of hydrodynamics is particularly notable for gorgonians with a fan morphology. These colonies are flexible and passively oriented, where water flow controls their orientation to enhance feeding and minimise hydrodynamic forces (Riedl and Forstner 1968; Wainwright and Dillon 1969; Leversee 1976; Lin et al. 2002; Rodríguez-Lanetty et al. 2003).
The direct link between nutrient availability and current speeds means that gradients in hydrodynamic energy strongly define gorgonian abundance and distribution. For example, across the distinct habitat zonation on coral reefs, most gorgonians are often found on the shallow, seaward reef crest, where there is highest hydrodynamic energy (Bayer 1961; Sánchez et al. 1997). Although surface-driven hydrodynamic energy decreases with depth, vertical patterns of gorgonian distribution with increasing depth are more variable. Although current speeds and water motion may be lower at greater depth, many gorgonians are found at higher abundances here and communities are often composed of larger species like fan or tree morphologies (Rodríguez-Lanetty et al. 2003; Quintanilla et al. 2019). This is often explained by the distinction between zooxanthellate or azooxanthellate species among gorgonians, as light availability at greater depths can restrict the distribution of some zooxanthellate species (Kinzie 1973; Fabricius and McCorry 2006; Haapkylä et al. 2007; Rowley 2018).
Hydrodynamic patterns also vary between locations on both regional and local scales where the relative influence of hydrodynamics will vary given differences in reef morphology, exposure and proximity to land (Galbraith et al. 2022). This includes multiple metrics of water quality including turbidity, freshwater input from rivers, terrestrial run-off, wave energy gradients, and the availability of nutrients (Perez-Ruzafa et al. 2019). These factors contribute to differences in environmental conditions between habitat zones on coral reefs and depths and directly influence benthic community composition (Done 1983). In turn, ecological interactions and the availability of suitable substrata also contribute to location specific gorgonian communities (Goh and Chou 1994). For example, lower richness of other benthic organisms, particularly competitors like some scleractinian corals, has been linked to higher gorgonian abundance (Goh and Chou 1994) and increased topographic complexity can increase habitat area available for settlement (Mortensen and Buhl-Mortensen 2004). The composition of the surrounding benthic community therefore can strongly influence the distribution and structure of gorgonian communities at a given location. Although many studies have examined patterns in gorgonian abundance and distribution and the influence of environmental variables, particularly current speeds, these are proportionally fewer in the Indo-Pacific and specifically the Great Barrier Reef (but see Rowley 2018; Kupfner Johnson and Hallock 2020).
This study is the first to investigate the spatial patterns of gorgonians around the Palm Island group, inshore Central Great Barrier Reef. Specifically, we evaluated spatial variation in the abundance, genera richness, and assemblage composition of gorgonians among depths and locations at the Palm Islands on the central Great Barrier Reef. We quantified benthic cover to assess the role of surrounding benthic habitat in driving spatial patterns in gorgonian communities. In addition, we conducted a preliminary investigation of site-specific hydrodynamics by comparing short term current speed averages between different depths. The following questions were addressed: (1) Does gorgonian abundance and genera richness vary among depths and locations in the Palm Islands? (2) Are gorgonian abundance and richness related to benthic cover? (3) How does gorgonian assemblage composition vary along the depth gradient? If so, does the relative abundance of azooxanthellate and zooxanthellate genera, and the different growth forms, vary with depth? (4) Are there differences in current speeds at the depths surveyed at the Palm Islands? We hypothesised that depth would be an important factor in explaining gorgonian distribution, but that it may not influence all gorgonian morphologies in the same way. We also hypothesised that benthic cover would be an important driver of abundance and assemblage composition as the surrounding substrate significantly affects gorgonian attachment and growth. Finally, whilst the scope of this study was not sufficient to attain long-term average current speed data, we also tested whether current speeds differ between depths at the Palm Islands.
Materials and methods
Study area
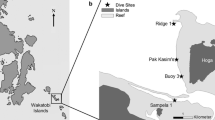
This study was carried out in the Palm Islands on the central Great Barrier Reef (GBR), Australia, based at Orpheus Island Research Station (− 18.7345° S, 146.5794° E). This group of inshore islands comprises 16 islands located within 20 km from the nearest mainland and has extensive areas of fringing reef. To explore the extent of spatial variation in gorgonian abundance, genera richness, and composition, surveys were carried out at 16 locations spread across four of the larger islands, including Orpheus, Pelorus, Fantome, and Curacoa (Fig. 1). The islands surveyed in this study are largely unpopulated, other than the presence of a research station and a resort on Orpheus Island. There is a permanent small community on Great Palm Island, and to avoid any potential effects of human impact, this island was excluded from our surveys. Locations were randomly chosen across the four islands, to explore natural spatial variation in gorgonian abundance and assemblage composition at this scale but were somewhat constrained by logistical considerations during the course of the study. Although nutrient availability from land-based run-off can affect the distribution of benthic organisms on coral reefs (Fabricius 2005), there are no major freshwater sources or rivers on any of the islands surveyed and all sites are approximately equidistant from the nearest mainland and river catchment. The prevailing winds in the Palm Islands come from the South East, so whilst some sites are located on the leeward side of the islands we surveyed, Great Palm Island and the outer Great Barrier Reef afford significant shelter to these locations.

Study locations (1–16) of gorgonian surveys at four islands, Orpheus Is. (Goolboddi), Pelorus Is. (Yanooa), Fantome Is. (Eumilli), and Curacoa Is. (Noogoo), in the Palm Island group on the central Great Barrier Reef
Gorgonian surveys and video analysis
At each location, four replicate 30-m belt transects were conducted at each of three depths (5, 10, and 15 m), summing to a total of 192 transects. Transects were laid out along the depth contour, parallel to the reef slope, with 3–5 m between each transect. Gorgonian abundance was quantified by diver-based surveys using a stereo-diver-operated video (DOV) system (SeaGis, Australia). Our DOV method followed the recommendations of Goetze et al. (2019), with two GoPro Hero 5 cameras set to 1080p resolution and 60fps wide setting. Video surveys were chosen as they facilitate accurate identification after surveys have been conducted, provide a permanent record, and allow for rapid data collection within a limited timeframe. We used EventMeasure software (SeaGis, Australia) to measure a 2.5-m field of view either side of each transect which provided a known belt width of 5 m. Videos were then played back in a standard mediaplayer (QuickTime Player). All gorgonian colonies within each transect were identified to genus level (as per Fabricius and Alderslade 2001) and their growth forms recorded, providing estimates of abundance and genera richness per 150m2. Sea whips were often found in large clumps and were difficult to count individually; therefore, estimates of their abundance were approximate.
Environmental variables: current speeds and habitat substrata
To quantify current speeds at different locations and depths during the course of the study, we deployed Marotte HS Drag-Tilt Current Meters (Marine Geophysics Laboratory, James Cook University, Australia) at the same three depths that were surveyed at each location for a period of 6 days. Whilst quantifying long-term averages was not possible for this study, these preliminary data are some of the first fine-scale hydrodynamic data available for the Palm Islands. Each current meter was deployed at the start of the first of the four transects at each depth, approximately 1 m next to it. Current meters were tethered to a weight in an upright position to ensure free movement of the instrument. All instruments were calibrated prior to deployment and set to record current speed (m s−1) and direction (degrees from North) every 10 s. After deployment, data were processed using the specialist software Marotte HS Config (Marine Geophysics Laboratory, James Cook University). Current speeds were averaged over the 6-day period for each depth at each site.
Benthic habitat surveys were carried out using a random point count method, with 300 points per video transect. Each video transect was stopped at 60 random times, and in each frame, the benthic organism or substratum beneath each of 5 random points was recorded. Benthic organisms and substrata were placed into the following categories: sand stable (not rippled), sand mobile (rippled), rubble, turf rock (i.e. rock covered in turf algae), macroalgae, sponges, soft corals, and hard corals of branching, encrusting and massive morphologies.
Statistical analysis
All statistical analyses were performed using RStudio (version 1.4.1717, R Core Team 2022). Generalised linear models (GLM) were used to test for differences in mean abundance and genera richness among locations and depths, and the interaction between location and depth. Both abundance and richness models used a negative binomial error family with log link and were fitted in the “MASS” package (Venables and Ripley 2002). Adjusted Tukey’s tests were used to identify pairwise differences between depth bands using the package “emmeans” (Russell et al. 2022).
We used GLMMs to test the effect of the percentage cover of each benthic substratum on gorgonian abundance. Each of these models used a negative binomial error family with log link and was structured with the percentage cover of each benthic organism/substratum as a fixed factor and location as a random factor. As a preliminary investigation of potential differences in hydrodynamic conditions with depth around the Palm Islands, we compared 6-day average current speeds between depth bands using GLMMs. The current speed GLMM used a Gaussian error family and identity link and were structured with depth as fixed factors and location as a random factor. All GLMMs were constructed using the package “glmmTMB” (Bolker 2019) which reports coefficient estimates with 95% confidence intervals. Marginal and conditional R2 values for GLMMs were calculated using the r.squared GLMM function of the “MuMIn” package (Barton 2020). Marginal R2 provides the variance explained only by the fixed effects, and conditional R2 provides the variance explained by the entire model. For all models, hypothesis tests of fixed effects were calculated as Type II Wald Chi-square tests conducted using the “anova” function from the package “car”. The goodness of fit for all models was assessed using the “DHARMa” package (Hartig 2021).
As assemblages may be affected by the simultaneous effects of environmental variables, a multivariate approach was used to assess the interactions between them. First, to visualise differences in gorgonian assemblage structure we performed non-metric multidimensional scaling (NMDS) using the function metaMDS, part of the “vegan” package (Oksanen 2020). Data were square root transformed, followed by a Wisconsin double standardisation and fit on a Bray–Curtis distance matrix. Fitted correlation vector scores for genera were extracted using the function “envfit” and genera with the strongest correlations (p < 0.001) plotted as a biplot using “ggplot2” (Wickham 2016). To then assess the role of depth in driving assemblage differences, we conducted permutational analysis of variance (PERMANOVA) using the adonis2 function in the “vegan” package. PERMANOVA was run on 999 permutations and a Bray–Curtis distance matrix. An analysis of multivariate homogeneity (PERMDISP) was carried out using the “betadisper” function to assess within group dispersion (Anderson and Walsh 2013).
Finally, to assess relationships among benthic organisms/substrata and gorgonian assemblage structure, we performed a distance-based redundancy analysis (db-RDA) on a Bray–Curtis distance matrix of square root transformed data. We used the functions “dbrda” and “envfit”, part of the “vegan” package. Only significant (p < 0.05) environmental variables were plotted as a vector biplot with the db-RDA axes (Paliy and Shankar 2016).
Results
General patterns
In the surveys, we recorded 4,610 gorgonian individuals, belonging to the suborders Calcaxonia, Holaxonia and Scleraxonia. This included 8 families and 23 genera (Table 1). The majority were azooxanthellate forms, except for Hiksonella, Isis, and Pteronisis. The genera included branching (9 genera), fan (7 genera), bushy (4 genera), whip (2 genera), and candelabrum (1 genus) growth forms (Fig. 2). In terms of relative abundance of the different genera, the fauna was dominated by two whip genera (Junceella and Viminella), two branching genera (Dichotella and Ellisella), and the candelabrum genera (Ctenocella), all belonging to the family Elisellidae (Fig. 2, Table 1). The other genera recorded were relatively rare.

Mean abundance of 23 gorgonian genera colour-coded by their growth forms: branched, bushy, fan, candelabrum, and whip
How does gorgonian abundance and genera richness vary among depths and locations?
There were significant differences in the abundance (pooled across all genera) of gorgonians among depths (\({\mathrm{GLM}, \chi }^{2}\) = 109.72, df = 2, p < 0.05) and among locations (\({\mathrm{GLM}, \chi }^{2}\) = 178.43, df = 15, p < 0.05). Abundance generally increased with increasing depth, with numbers at 15 m about five times higher than at 5 m (Fig. 3a). However, the increase in abundance with depth was not consistent among locations, as indicated by a significant interaction between depth and location (GLM, \({\chi }^{2}\) = 194.62, df = 30, p < 0.05). Pairwise comparisons between depths found that abundance at 5 m was significantly different to that at 10 m and 15 m, but did not differ between 10 and 15 m (Table 2a).

Mean gorgonian abundance a and genera richness b at each of the three depths. Error bars are 95% confidence intervals
Genera richness also varied among depths (\({\mathrm{GLM}, \chi }^{2}\) = 119.363, df = 2, p < 0.05) and among locations (\({\mathrm{GLM}, \chi }^{2}\) = 85.769, df = 15, p < 0.05), with richness increasing with increasing depth (Fig. 2b). This increase in richness with depth was not consistent among locations (GLM, \({\chi }^{2}\) = 76.548, df = 30, p < 0.05), but on average, genera richness at 15 m was three times higher than at 5 m (Fig. 3b). Pairwise comparisons showed significant differences in genera richness among all depths (Table 2b).
How are gorgonian abundance and richness related to benthic habitat cover?
The effects of benthic organisms or substrata on gorgonian abundance varied, with some positive and some negative trends detected (Fig. 4). The per cent cover of rubble was a significant positive predictor of abundance (\({\mathrm{GLMM}, \chi }^{2}=5.82,\) df = 1, p = 0.02), with a 4.92 increase in gorgonian abundance every 1 per cent increase in cover of rubble (95% CI 1.00–1.03) (Fig. 4a). As with current speed, the relationship was weak, explaining just 5.4% of the variation in gorgonian abundance.

Relationships between gorgonian abundance a and current speed, and genera richness and current speed b, with 95% confidence interval from GLMMs. Colours indicate the three depths surveyed (5, 10, 15 m). Points have been jittered

Relationships between gorgonian abundance and the per cent cover of rubble a, branching hard corals b, with gorgonian abundance pooled across the three depths, with 95% confidence interval from GLMMs
There was a significant negative relationship between gorgonian genera richness and branching hard coral cover (\({\mathrm{GLMM} ,\chi }^{2}=7.6302, \mathrm{d}f=1, p=0.005)\), with genera richness halving with an increase of 1.96 per cent cover of branching hard coral. There were no significant relationships between gorgonian richness and any of the other benthic organisms or substrata.
How does gorgonian assemblage structure vary along the depth gradient?
The PERMANOVA showed that assemblage structure varied significantly among the three depths (R2 = 0.45, pseudo-F = 21, p = 0.001; betadisper, pseudo-F = 27.201, p = 0.001). The NMDS shows that the assemblage structure of gorgonian genera at 5 m is distinct from that at 10 and 15 m, which reflects the very low genera richness and absence of many genera at this depth. The NMDS also shows a gradual transition in genera composition from 10 to 15 m (Fig. 5a). Nine of the 23 genera recorded were significant in explaining the overall depth patterns in assemblage structure (Fig. 5b). All of these gorgonian genera were azooxanthellate (Table 1). However, there was a transition from a dominance of whip (Junceella, Viminella) and fan (Acanthogorgia, Anthogorgia, Annella) growth forms at 10 m to branching (Dichotella, Icilogorgia) and candelabrum (Ctenocella) growth forms at 15 m.
What is the role of habitat structure in explaining spatial variation in the structure of gorgonian assemblages?
The distance-based redundancy analysis found that the benthic organisms/substrata explained 37.7% of variation in gorgonian community structure, which clearly varied along the depth gradient (permutation test, pseudo-F = 9.91, p = 0.001, 999 permutations) (Fig. 6). Benthic organisms/substrata had significant effects in the constrained ordination, with the exception of mobile sand and sponges (Table 3). The first axis of the db-RDA accounted for 84.58% of fitted variation (23.72% of the total variation) and strongly correlated with branching hard coral (Fig. 6b). This reflects high coral cover and low gorgonian abundance in shallow water. Vector overlays show that greater depths (10 and 15 m) were associated with higher per cent cover of rubble, turf rock, and macroalgae. High macroalgal cover was also associated with higher current speed. Both of these habitat variables, turf rock and macroalgae, were more aligned with the second axis of the db-RDA which explained 11.90% of fitted variation 3.34% of total variation.

Non-metric multidimensional scaling (NMDS) plot a showing gorgonian assemblage structure across three depths (5, 10, and 15 m). Points represent gorgonians, grouped by assemblage structures across depths using circles. NMDS plot b with arrows showing the nine most common genera of gorgonians across the assemblage structure, including only genera with p < 0.002
Differences in current speeds between depths
Over the 6-day study period, current speeds were significantly different between depths \(({\mathrm{GLMM} ,\chi }^{2}=71.13, \mathrm{d}f=1, p<0.01)\) (Fig. 7A). Highest average currents were recorded at 15 m (0.12 m s−2, 95%CI [0.10–0.14 m s−2) and were lower at 10 m (0.09 m s−2, 95% CI [0.07–0.11 m s−2) and lowest at 5 m (0.08 m s−2, 95% CI [0.01–0.09 m s−2). Whilst there was no difference in current speeds between the two shallowest depths (contrast estimate 5–10 m = − 0.01, 95% CI [− 0.02–0.00]), currents at 15 m were significantly stronger than both 10 and 15 m (Fig. 7B, Table 4).

A. Mean current speeds (m s−1) recorded at each depth band throughout the 6-day sampling period and B. Pairwise contrast estimates based on adjusted Tukey’s tests. The difference is considered significant if the confidence intervals do not contain zero. Error bars in both plots represent 95% confidence intervals
Discussion
This study showed that gorgonians are a diverse component of the benthic communities on fringing reefs of the Palm Island Group, Great Barrier Reef. We recorded a total of 8 families and 23 genera; however, the gorgonian assemblage was numerically dominated by five genera from the family Elisellidae, which made up 43% of the total abundance of gorgonians. There were an increase in the abundance and genera richness of gorgonians in deeper waters and a pronounced change in the structure of gorgonian assemblages along the depth gradient. These patterns were not consistent across locations, despite considerable variation in abundance, species richness, and assemblage structure among locations. The depth-related patterns in assemblage structure were evident at the level of growth form, with whip (Junceella, Viminella) and fan (Acanthogorgia, Anthogorgia, Annella) growth forms in shallow water being replaced by branching (Dichotella, Icilogorgia) and candelabrum (Ctenocella) growth forms at greater depths. Benthic cover was also important, with gorgonian abundance being positively related to the per cent cover of rubble and negatively related to branching hard coral cover. Gorgonian communities were depauperate in shallow water in most of the locations surveyed, where branching hard coral cover was high (e.g. North Orpheus Island), and they were more diverse in deeper water where the per cent cover of turf, rubble and macroalgal cover was high. Relationships with all of these drivers were generally weak, and it is likely that gorgonian assemblages at the Palm Islands are influenced by a large range of biophysical factors that favour different growth forms at different depths. Further work at the species level will be required to evaluate the relative importance of these factors.
Depth-related patterns in gorgonian assemblages
Most benthic organisms on coral reefs exhibit distinct patterns in abundance, diversity, and assemblage structure along depth gradients, and gorgonians are clearly no exception (Rowley 2018). Our study showed gorgonian abundance and genera richness increased with depth over a relatively small depth range (5–15 m). Similar patterns have been previously described for Indonesia (Rowley 2018) and in the Southern Mexican Pacific coast (Abeytia et al. 2013), with a substantial increase in abundance with depth. In Singapore, gorgonian genera richness was higher in deep water (~ 25 m), where light availability was low; however, gorgonian richness was also high in relatively shallow areas (~ 10 m) where there were low abundances of competitors (e.g. hard corals and sponges) and low light availability due to high turbidity, usually typical of deeper areas (Goh and Chou 1994).
Depth-related patterns in gorgonian abundance, diversity, and assemblage structure are linked to a range of biophysical factors. For example, high sedimentation loads at shallow depths can impact gorgonian abundance negatively (Yoshioka and Yoshioka 1989), nutrient enrichment can have a negative impact on gorgonians but not as severe like for hard corals (McCauley and Goulet 2019), and variability in benthic habitat structure can influence their compositions (Rowley et al. 2015). A study conducted in Indonesia by Rowley (2018) showed that azooxanthellate gorgonians had higher abundances and diversity in deeper areas where there is lower light. Similarly, this study showed that whilst the abundance of most gorgonian genera increased with depth at the Palm Islands, the gorgonian assemblages at all depths were dominated by azooxanthellate genera.
The dominance of azooxanthellate gorgonian species in the Pacific region is thought to be a product of adaptive radiation after the closure of the Isthmus of Panama (Sánchez 2016). This event drastically altered the oceanographic conditions either side of the Isthmus, where the Caribbean turned oligotrophic with reduced plankton availability (Coates et al. 1992) and frequent upwelling in the Pacific enhanced productivity and plankton availability (Fernández-Álamo and Färber-Lorda 2006). Azooxanthellate gorgonian corals were able to access new feeding resources as suspension feeders, which likely drove rapid evolutionary speciation (Schluter 2016). In contrast, Caribbean gorgonians are primarily zooxanthellate, residing in clear shallow waters. A laboratory study by Kinzie (1973) investigated the ability of Caribbean gorgonians to withstand low light, by placing zooxanthellate and azooxanthellate gorgonians in a dark aquarium for 83 days. Most of the zooxanthellate forms died, presumably due to low light, which may explain their absence in deep water. Differently, in areas such as Hong Kong, where sediment and turbidity levels were high, and water quality low, the octocoral community was mainly composed by azooxanthellate taxa, more resistant high turbidity and low light availability (Fabricius and McCorry 2006). Therefore, waters surrounding the Palm Islands are characterised by high turbidity, which may explain the dominance of azooxanthellate gorgonians even in shallow water.
This study shows that changes in assemblage composition along a depth gradient are likely linked to changes in the performance of different growth forms at different depths. Genera with whip and fan growth forms were more prevalent at 5 and 10, whilst branched, bushy, and candelabrum growth forms dominated at 15 m. Our preliminary analysis found that daily average current speeds were highest at the deepest depths during the course of the 6-day sampling period. Multiple other studies have shown the importance of current speeds in driving gorgonian abundance and community structure (Muzik and Wainwright 1977; Rodríguez-Lanetty et al. 2003; Quintanilla et al. 2019), and we suspect that differences among genera in current speed requirements and tolerance to strong currents are a major determinant of the depth-related trends at the Palm islands. It has been suggested previously that deep water may be a refuge for genera that are less tolerant to physical disturbance (Costantini et al. 2016). Gorgonian colonies in shallow water are more susceptible to breakage due to higher wave action (West et al. 1993). Morphological features of gorgonians can also alter the intake of light, with different light quantities captured through the bushy and branching structures than fans (Rodríguez-Lanetty et al. 2003; Rowley et al. 2015). At the Palm Islands, the overall gorgonian fauna presented nine genera with branching growth forms (Dichotella, Ellisella, Icilogorgia, Menella, Astrogorgia, Mopsella, Paraplexaura, Pseudopterogorgia, Subergorgia) and seven with fan growth forms (Annella, Acanthogorgia, Anthogorgia, Meliathaea, Euplexaura, Muricella, Villogorgia). The Caribbean gorgonian fan, Gorgonia ventalina, resides in shallow water, and its structure is presumed to help to capture light (Baker et al. 2015). Our study shows that gorgonians with tree-like morphologies, such as candelabrum (Ctenocella sp.) and branched (Dichotella, Elisella sp.), can live in areas with relatively high current flows, which is consistent with the findings of other studies (Sánchez et al. 1997; Rodríguez-Lanetty et al. 2003). Ultimately, the prevalence of whips and fans at 5–10 m, and complex branching, bushy and candelabrum forms at 15 m, requires further investigation. Additionally, as filter feeders, currents are important determinants of gorgonian community structure and distribution (Sponaugle 1991). Specifically, gorgonians are known to be more abundant in areas of high current speed, where a greater abundance of food is available (Chang-Feng and Ming-Chao 1993; Muzik and Wainwright 1977; Quintanilla et al. 2019). However, our preliminary current speed data are not sufficient to assess causative relationships with gorgonian abundance or richness but do suggest significant differences in hydrodynamic conditions with depth at the Palm Islands. This hypothesis, however, requires long-term averages of current speeds and other hydrodynamic data (e.g. temperature, prevailing current direction) to test fully.
The importance of benthic cover
Our study also confirms that benthic cover can play an important role in shaping gorgonian assemblages (Dahlgren 1989; Sánchez et al. 1997). We found overall abundance was positively related to the per cent cover of rubble, but negatively related to the per cent cover of branching hard coral (Goh and Chou 1994). Almost all of the benthic organisms/substrates explained spatial variation in assemblage structure, which likely represents genera-specific responses to the availability of preferred habitat. Low gorgonian genera richness in shallow water was associated with higher hard and soft coral cover, whilst greater genera richness in deeper water was associated with higher rubble, turf rock, and/or macroalgal cover, each of which may favour different growth forms. Despite the shallow coral reef habitat having higher structural complexity, with branching hard corals and a wide variety of soft corals, there was lower gorgonian abundance and genera richness, which may be due to higher competition for space.
Habitat structure has been shown to be important in other investigations into gorgonian distributions, but the factors that are important vary among studies. Topographic complexity plays a critical role in shaping many ecological patterns on coral reefs, and strong positive relationships have been found between topographic complexity and gorgonian abundance and diversity (Al-Marayati and Edmunds 2018). Similarly, but in contrast to our findings, areas in the Caribbean show a high abundance of gorgonians in complex habitats dominated by branching hard corals and other benthic structure-forming organisms like sponges (Sánchez et al. 1997; Steiner et al. 2018). Clearly, numerous benthic organisms contribute to the overall complexity of a given habitat, yet some may negatively influence gorgonian communities as a result of direct ecological interactions rather than beneficially provide complex habitat structure. Sponges, for example, increase habitat complexity on coral reefs (Maldonado et al 2015) but are also known to negatively influence the distribution of gorgonians through competition for settlement space and allelopathic overgrowth (Dahlgren 1989; Slattery and Lesser 2021). Low complexity habitats with greater proportions of solid substrata are therefore preferred habitat types for some octocorals (Sánchez et al. 1997; Steiner et al. 2018), where bare areas of substrate may facilitate settlement and growth as there is low abundance of such competitors (Goh and Chou 1994). Gorgonian assemblages also follow spatial patterns corresponding to changes in benthic habitat zonation which are not always directly related to the absolute complexity of the habitat (Goldberg 1973; Kinzie 1973; Dahlgren 1989). In Western Australia, for example, a cross shelf-gradient and resulting differences in benthic habitat composition between inshore versus offshore locations was found to be a stronger driver of octocoral assemblage composition than habitat complexity per se (Bryce et al. 2018).
Ecological importance and further work
Our study adds to a diverse range of patterns in the distribution and abundance of gorgonians and suggests we are not yet at a stage where general conclusions can be made. Nevertheless, we provide important baseline information at the genus level on the spatial distribution of gorgonians at the Palm islands and have generated a number of hypotheses concerning the roles habitat structure in explaining depth distributions. Given that multiple biophysical factors are responsible for shaping gorgonian assemblages and vary in their importance among species, further work is needed on this often-neglected group. Although all sites are equidistant from the nearest mainland and largest river catchments, further data on water quality (turbidity, nutrient content, etc.) would also be valuable to characterise hydrodynamic conditions in the Palm Islands. At the most fundamental scale, species-level taxonomy is currently uncertain and will need to be resolved to for patterns to be fully described. The depth patterns are likely linked to a host of physical (turbidity and light penetration, wave action, current speed, substratum stability, topography) and biological factors (food availability, competing species), all of which should be investigated further. In particular, the tolerance of different growth forms to different depths and current speeds is an interesting avenue for future research in the Palm Islands.
References
Abeytia R, Guzmán HM, Breedy O (2013) Composición de especies y distribución batimétrica de gorgonios (Anthozoa: Octocorallia) en la costa sur del pacífico Mexicano. Rev Biol Trop 61:1157–1166
Al-Marayati M, Edmunds PJ (2018) Octocoral distribution is associated with substratum orientation on coral reefs in St. John, U.S. Virgin Islands. J Exp Mar Bio Ecol 500:55–62
Anderson MJ, Walsh DCI (2013) PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecol Monogr 83:557–574
Baker DM, Freeman CJ, Knowlton N, Thacker RW, Kim K, Fogel ML (2015) Productivity links morphology, symbiont specificity and bleaching in the evolution of Caribbean octocoral symbioses. ISME J 9:2620–2629
Barton K (2020) MuMIn: multi-model inference. R package version 1.43.17. Version 1:18
Bayer FM (1961) The shallow-water Octocorallia of the West Indian region. A manual for marine biologists. Stud Fauna Curacao Caribbean Islands 12:1–373
Bellwood DR, Pratchett MS, Morrison TH, Gurney GG, Hughes TP, Álvarez-Romero JG, Day JC, Grantham R, Grech A, Hoey AS, Jones GP, Pandolfi JM, Tebbett SB, Techera E, Weeks R, Cumming GS (2019) Coral reef conservation in the Anthropocene: confronting spatial mismatches and prioritizing functions. Biol Conserv 236:604–615
Bolker B (2019) Getting started with the glmmTMB package. CranR-Project Vignette 9
Bryce M, Radford B, Fabricius K (2018) Soft coral and sea fan (Octocorallia) biodiversity and distribution from a multitaxon survey (2009–2014) of the shallow tropical Kimberley, Western Australia. Rec West Aust Museum 85:45
Buhl-Mortensen L, Vanreusel A, Gooday AJ, Levin LA, Priede IG, Buhl-Mortensen P, Gheerardyn H, King NJ, Raes M (2010) Biological structures as a source of habitat heterogeneity and biodiversity on the deep ocean margins. Mar Ecol 31:21–50
Calcinai B, Bavestrello G, Bertolino M, Pica D, Wagner D, Cerrano C (2013) Sponges associated with octocorals in the Indo-Pacific, with the description of four new species. Zootaxa 3617:1–61
Chang-Feng D, Ming-Chao L (1993) The effects of flow on feeding of three gorgonians from southern Taiwan. J Exp Mar Bio Ecol 173:57–69
Coates AG, Jackson JB, Collins LS, Cronin TM, Dowsett HJ, Bybell LM (1992) Closure of the Isthmus of Panama: the near-shore marine record of Costa Rica and western Panama. Geol Soc Am Bull 104(7):814–828
Costantini F, Gori A, Lopez-González P, Bramanti L, Rossi S, Gili JM, Abbiati M (2016) Limited genetic connectivity between gorgonian morphotypes along a depth gradient. PLoS ONE 11:1–20
Dahlgren EJ (1989) Gorgonian community structure and reef zonation patterns on Yucatan coral reefs. Bull Mar Sci 45:678–696
Diaz MC, Rützler K (2001) Sponges: an essential component of Caribbean coral reefs. Bull Mar Sci 69:535–546
Dinesen ZD (1983) Patterns in the distribution of soft corals across the central Great Barrier Reef. Coral Reefs 1:229–236
Done TJ (1983) Coral zonation, its nature and significance. Perspectives on coral reefs. Australian Institute of Marine Science (AIMS), pp 107–147
Evans A, Steer M, Belle E (2011) The Alcyonacea (soft corals and sea fans) of Antsiranana Bay, northern Madagascar. Madagascar Conserv Dev 6:29–36
Fabricius KE (2005) Effects of terrestrial runoff on the ecology of corals and coral reefs: review and synthesis. Mar Pollut Bull 50(2):125–146
Fabricius KE, McCorry D (2006) Changes in octocoral communities and benthic cover along a water quality gradient in the reefs of Hong Kong. Mar Pollut Bull 52:22–33
Fabricius KE, Alderslade AP (2001) Soft corals and sea fans: a comprehensive guide to the tropical shallow water genera of the central-west Pacific, the Indian Ocean and the Red Sea. Australian Institute of Marine Science
Fernández-Álamo MA, Färber-Lorda J (2006) Zooplankton and the oceanography of the eastern tropical Pacific: a review. Prog Oceanogr 69(2–4):318–359
Galbraith GF, Cresswell BJ, McCormick MI, Bridge TC, Jones GP (2022) Contrasting hydrodynamic regimes of submerged pinnacle and emergent coral reefs. PLOS ONE 17(8):e0273092. https://doi.org/10.1371/journal.pone.0273092
Goetze JS, Bond T, McLean DL, Saunders BJ, Langlois TJ, Lindfield S, Harvey ES (2019) A field and video analysis guide for diver operated stereo-video. Methods Ecol Evol 10(7): 1083–1090
Goh NKC, Chou LM (1994) Distribution and biodiversity of Singapore gorgonians (sub-class Octocorallia) - a preliminary survey. Hydrobiologia 285:101–109
Goh NK, Loo MG, Chou LM (1997) An analysis of gorgonian (Anthozoa; Octocorallia) zonation on Singapore reefs with respect to depth. Environ Monit Assess 44:81–89
Goldberg WM (1973) The ecology of the coral-octocoral communities off the southeast Florida coast: geomorphology, species composition, and zonation. Bull Mar Sci 23:465–488
Graham NAJ, Nash KL (2013) The importance of structural complexity in coral reef ecosystems. Coral Reefs 32:315–326
Grinyó J, Gori A, Ambroso S, Purroy A, Calatayud C, Dominguez-Carrió C, Coppari M, Lo Iacono C, López-González PJ, Gili JM (2016) Diversity, distribution and population size structure of deep Mediterranean gorgonian assemblages (Menorca Channel, Western Mediterranean Sea). Prog Oceanogr 145:42–56
Haapkylä J, Seymour AS, Trebilco J, Smith D (2007) Coral disease prevalence and coral health in the Wakatobi Marine Park, south-east Sulawesi, Indonesia. J Mar Biol Assoc United Kingdom 87(2): 403–414
Hartig F (2021) DHARMa: residual diagnostics for hierarchical (multi-level/mixed) regression models. R package version 0.4.4
Hoegh-Guldberg O (2011) Coral reef ecosystems and anthropogenic climate change. Reg Environ Chang 11:215–227
Hughes TP, Barnes ML, Bellwood DR, Cinner JE, Cumming GS, Jackson JBC, Kleypas J, Van De Leemput IA, Lough JM, Morrison TH, Palumbi SR, Van Nes EH, Scheffer M (2017) Coral reefs in the Anthropocene. Nature 546:82–90
Jones CG, Lawton JH, Shachak M (1994) Organisms as ecosystem engineers. OIKOS 69:373–386
Jones CG, Lawron JH, Shachak M (1997) Positive and negative effects of organisms as physical ecosystem engineers. Ecology 78:1946–1957
Kinzie RA III (1973) The zonation of west indian gorgonians. Bull Mar Sci 23:93–155
Kupfner Johnson S, Hallock P (2020) A review of symbiotic gorgonian research in the western Atlantic and Caribbean with recommendations for future work. Coral Reefs 39:239–258
Leversee GJ (1976) Flow and feeding in fan-shaped colonies of the gorgonian coral, Leptogorgia. Biol Bull 151:344–356
Lin MC, Liao CM, Dai CF (2002) Modeling the effects of satiation on the feeding rate of a colonial suspension feeder, Acanthogorgia vegae, in a circulating system under lab conditions. Zool Stud 41:355–365
Maggioni D, Montano S, Voigt O, Seveso D, Galli P (2020) A mesophotic hotel: the octocoral Bebryce cf. grandicalyx as a host. Ecology 101(4):e02950
Maldonado M, Aguilar R, Blanco J, Garcia S, Serrano A, Punzon A (2015) Aggregated clumps of lithistid sponges: a singular, reef-like bathyal habitat with relevant paleontological connections. PLoS ONE 10(5):e0125378
McCauley M, Goulet TL (2019) Caribbean gorgonian octocorals cope with nutrient enrichment. Mar Pollut Bull 141:621–628
McFadden CS, van Ofwegen LP, Quattrini AM (2022) Revisionary systematics of Octocorallia (Cnidaria: Anthozoa) guided by phylogenomics. Bull Soc Syst Biol 1(3)
Mortensen PB, Buhl-Mortensen L (2004) Distribution of deep-water gorgonian corals in relation to benthic habitat features in the Northeast Channel (Atlantic Canada). Mar Biol 144(6):1223–1238
Muzik K, Wainwright S (1977) Morphology and habitat of five Fijian sea fans. Bull Mar Sci 27:308–337
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O'Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2020) vegan: community ecology package. R package version 2.5-7
Paliy O, Shankar V (2016) Application of multivariate statistical techniques in microbial ecology. Mol Ecol 25:1032–1057
Pandolfi JM, Bradbury RH, Sala E, Hughes TP, Bjorndal KA, Cooke RG, Mcardle D, Mcclenachan L, Newman MJH, Paredes G, Warner RR, Jackson JBC (2003) Global trajectories of the long-term decline of coral reef. 301:955–959
Perez-Ruzafa A, Campillo S, Fernández-Palacios JM, Garcia-Lacunza A, Garcia-Oliva M, Ibanez H, Marcos C (2019) Long-term dynamic in nutrients, chlorophyll a, and water quality parameters in a coastal lagoon during a process of eutrophication for decades, a sudden break and a relatively rapid recovery. Front Mar Sci 6:26
Quintanilla E, Madurell T, Wilke T, Sánchez JA (2019) Dynamic interplay of ENSO events and local hydrodynamic parameters drives demography and health status of gorgonian sea fan populations on a remote tropical Eastern Pacific Island. Front Mar Sci 6:1–9
Riedl R, Forstner H (1968) Wasserbewegung im Mikrobereich des Benthos. Sarsia 34(1):163–188
Rodríguez-Lanetty M, Marquez LM, Losada F (2003) Changes in gorgonian morphology along a depth gradient at Isla Alcatraz, San Esteban National Park, Venezuela. Bull Mar Sci 72:1019–1023
Rowley SJ (2018) Environmental gradients structure gorgonian assemblages on coral reefs in SE Sulawesi, Indonesia. Coral Reefs 37:609–630
Rowley SJ, Pochon X, Watling L (2015) Environmental influences on the Indo-Pacific octocoral Isis hippuris Linnaeus 1758 (Alcyonacea: Isididae): Genetic fixation or phenotypic plasticity? PeerJ 2015:1–27
Russell A, Lenth V, Buerkner P, Herve M, Love J, Singmann H, Lenth MR V (2022) Package ‘ emmeans ’ R topics documented. 34:216–221
Sánchez JA (2016) Diversity and evolution of octocoral animal forests at both sides of tropical America. In: Rossi S, Bramanti L, Gori A, Orejas Saco del Valle C (eds) Marine animal forests. Springer, Cham
Sánchez JA, Díaz JM, Zea S (1997) Gorgonian communities in two contrasting environments on oceanic atolls of the southwestern Caribbean. Bull Mar Sci 61:453–465
Slattery M, Lesser MP (2021) Gorgonians are foundation species on sponge-dominated Mesophotic Coral Reefs in the Caribbean. Front Mar Sci 8:1–14
Speciation SD (2016) Speciation, Ecological opportunity, and latitude: (American society of naturalists address)*. Am Nat 187(1):1–18
Sponaugle S (1991) Flow patterns and velocities around a suspension-feeding gorgonian polyp: evidence from physical models. J Exp Mar Bio Ecol 148:135–145
Steiner SCC, Riegl B, Lavorato A, Rodríguez J (2018) Community structure of shallow water Alcyonacea (Anthozoa: Octocorallia) from the southern Tropical Eastern Pacific. Ecol Res 33:457–469
Venables WN, Ripley BD (2002) Modern applied statistics with S, 4th edn. Springer, New York
Verdura J, Linares C, Ballesteros E, Coma R, Uriz MJ, Bensoussan N, Cebrian E (2019) Biodiversity loss in a Mediterranean ecosystem due to an extreme warming event unveils the role of an engineering gorgonian species. Sci Rep 9(1):5911
Wainwright SA, Dillon JR (1969) On the orientation of sea fans (genus Gorgonia). Biol Bull 136(1):130–139
West JM, Harvell CD, Walls AM (1993) Morphological plasticity in a gorgonian coral (Briareum asbestinum) over a depth cline. Mar Ecol Prog Ser 94:61–69
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer-Verlag, New York
Williams GJ, Graham NAJ, Jouffray JB, Norström AV, Nyström M, Gove JM, Heenan A, Wedding LM (2019) Coral reef ecology in the Anthropocene. Funct Ecol 33:1014–1022
Woodhead AJ, Hicks CC, Norström AV, Williams GJ, Graham NAJ (2019) Coral reef ecosystem services in the Anthropocene. Funct Ecol 33:1023–1034
Yoshioka PM, Yoshioka BB (1989) Effects of wave energy, topographic relief and sediment transport on the distribution of shallow-water gorgonians of Puerto Rico. Coral Reefs 8:145–152
Acknowledgements
We acknowledge the Traditional Owners of the Palm Islands, the Manbarra people and thank them for allowing us conduct research in their sea country. We would like to thank Abigal Shaugnessy, Ayla Woolley, Kathy Lisa Connellan, Sina Joelle Strähl, and Tess Jenkins for their assistance with fieldwork, and Saul Gonzalez Murcia for help with statistical analyses. We thank Orpheus Island Research Station, James Cook University, for logistical support. The research was funded by an ARC Discovery Grant (DP19010356) to G.P. Jones and a JCU College of Science and Engineering Competitive Research Training Grant to M. Panero. The research was conducted under GBRMPA permit number G21/45180.1 and JCU Animal Ethics approval number A2725. We thank three anonymous reviewers for their detailed and constructive comments which considerably improved this manuscript.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Panero, M., Galbraith, G.F., Srinivasan, M. et al. Roles of depth, current speed, and benthic cover in shaping gorgonian assemblages at the Palm Islands (Great Barrier Reef). Coral Reefs 42, 1045–1057 (2023). https://doi.org/10.1007/s00338-023-02411-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-023-02411-6