
Abstract
Coral reef ecology has advanced in many fields, but disease patterns across depth gradients remain unclear. By comparing the prevalence and extent of bleaching and diseases in 160 colonies of Siderastrea stellata between shallow and mesophotic reefs, we observed that prevalence was high (75%) regardless of depth, but the extent was about two times greater in mesophotic than shallow reefs (14.4% vs. 6.6% of colony area, respectively). Across the shallow reefs, where S. stellata co-occurred with Montastraea cavernosa, M. cavernosa showed lower prevalence (27% of 30 colonies) and extent (1.8% of colony area) compared to S. stellata. Besides bleaching, five coral diseases afflicted S. stellata and two affected M. cavernosa. Because diseases are spread over the entire gradient of depth, any attempt of managing the diseases should consider both shallow and deep reefs to be effective.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Coral reefs are facing unprecedented local and global anthropogenic disturbances (Sully et al. 2019). Threats such as ocean warming, overfishing, pollution, and disease outbreaks are transforming the way that reef ecosystems function (Harborne et al. 2017). Many of these threats are not occurring in isolation, but instead may have additive impacts on coral reef ecosystems (Ferreira et al. 2012). For example, although disease is a natural part of coral lifecycles, interactions with other threats such as ocean warming often exacerbates the negative effects of disease on corals (Randall and Van Woesik 2015; Howells et al. 2020). Indeed, coral disease outbreaks have increased over time, and have now become a major threat to corals in several areas of the world (Maynard et al. 2015).
Although there is an extensive body of literature documenting the impact of disease on corals, most of these studies are based on corals settled in shallow reefs (Montilla et al. 2019; Howells et al. 2020; Bloomberg and Holstein 2021). Our understanding on the dynamics of coral diseases in mesophotic ecosystems (MCEs; between 30 and 150 m depth) is still in its infancy (Weil 2019; Williams et al. 2021; Morais et al. 2022), especially in the Southwestern Atlantic, where in-depth investigations on MCEs are even more recent (Morais et al. 2018; Soares et al. 2018). This lack of scientific research contrasts with the unique MCEs biodiversity and ecological services (Soares et al. 2020b) and calls attention for the understanding of key ecological interactions along depth gradients, including coral diseases (Rogers et al. 2015).
To help bridge this knowledge gap, we sampled the two most abundant coral species of the Southwestern Atlantic, Montastraea cavernosa and Siderastrea stellata (Leão et al. 2016) across a depth gradient from 3 to 61 m. Besides identifying the diseases from the morphological signs, we estimated and compared their prevalence and extent between species and depth categories.
Methods
Study area and data survey
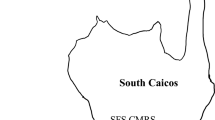
We conducted the study on reefs off the eastern coast of Northeast Brazil, state of Paraiba (Fig. 1). Between January and February of 2017, we surveyed coral colonies with SCUBA in four shallow and four mesophotic reefs (hereafter deep reefs) by employing a video transect method (Safuan et al. 2015). We used two GoPro video cameras to record digital videos over 20 m transects (following Morais and Santos (2018)) in four transects per reef (Table S1). During this study, temperatures aligned with historical averages (about 28–29 °C up to the 50 m depth). The reefs surveyed had on average 3.4% of coral cover, thus density-dependence effects were not expected to play a major role on disease prevalence and extent.

Map showing the sampling points in Northeast Brazil (South Atlantic Ocean). Blue dots represent the shallow reefs (< 30 m), red dots represent the deep reefs (> 30 m). a- Siderastrea stellata. b- Montastraea cavernosa. Photographs by Morais J
Image processing and statistical analyses
During video post-processing, we took screen shots of all frames containing colonies of S. stellata and/or M. cavernosa. We are aware that the taxonomy of the ‘Siderastrea Complex’ is not fully resolved in the South Atlantic due to morphological similarity among S. stellata, S. radians and S. siderea (Menezes et al. 2013). However, we reported the results as S. stellata based on our previous experience (Morais and Santos 2018) and other studies conducted in the study area (e.g., Costa et al. 2008). The pictures were subsequently processed in Adobe Photoshop by selecting the entire colony and obtaining the total area (pixels), corresponding to 100% of colony live tissue area. When signs of bleaching or disease were visible, we selected the corresponding (pixels) and calculated their relative area based on the total area of the colony. Coral diseases were identified using the Coral Disease Handbook (Raymundo et al. 2008) and the Field Guide to Western Atlantic Coral Diseases (Bruckner 2003). It is worth to mentioning that not all tissue damages may be associated with biotic causative agents, just as apparently healthy colonies may be diseased but not displaying gross signs yet. Damages from predators have particular shape and size that allow their differentiation from bleaching and disease signs, thus, they were not considered here.
To compare the prevalence and extent of disease/bleaching between species and in S. stellata between shallow and deep reefs (for M. cavernosa, we were not able to perform the same comparisons given the limited number of colonies below 30 m), we used generalized linear mixed effects models (GLMMs). In the GLMMs, prevalence and extent were the response variables while either species or depth were treated as fixed factors depending on the model in question. Site was treated as a random factor in all models to account for any lack of spatial independence. To examine the differences in the disease prevalence between species and between depths for S. stellata, we used a binomial distribution with logit link function. To evaluate the extent between species and between depths of S. stellata we used a tweedie (log link) and a betabinomial (logit link) distribution, respectively. Model fit and assumptions were assessed using residual plots, all of which were satisfactory. Statistical modelling was performed in the software R (R Core Team, 2020), using the glmmTMB (Brooks et al. 2017) and DHARMa (Hartig 2017) packages.
Results and discussion
We documented a total of 190 coral colonies: 160 colonies of S. stellata (107 in shallow and 53 in deep reefs) and 30 colonies of M. cavernosa (27 in shallow and only 3 in deep reefs). There was a significant difference in the prevalence of disease and bleaching between species (Table S2). Approximately 75% (121) of the S. stellata colonies and 27% (8) of the M. cavernosa colonies showed some sign of disease and bleaching (Fig. 2). The extent of disease and bleaching was also significantly greater in S. stellata, with a median of 8.5% compared to 1.8% in M. cavernosa (Fig. 3a). These results indicate that S. stellata and M. cavernosa are afflicted by disease to different extents. Although these two species have massive growth form and may have similar susceptibility strategy (Loya et al. 2001; Morais et al. 2021), M. cavernosa is usually classified as a stress-tolerant species, while S. stellata is reported as weedy species (Darling et al. 2012). Our finding agrees with this classification and supports previous studies that have shown the genus Siderastrea to have up to four times more prevalence of disease than the genus Montastraea (Calnan et al. 2007).

Proportion of diseased/bleached and intact colonies in shallow (< 30 m depth) and deep reefs (> 30 m depth) of Northeast Brazil. N = 160 colonies of S. stellata and 30 colonies of M. cavernosa

a Percentage of diseased/bleached area in colonies of Montastraea cavernosa and Siderastrea stellata. b Percentage of diseased/bleached area in colonies of S. stellata in shallow and deep reefs. Box-plots show the median (line) and quartiles distributions. Points represent the colonies
There was no significant difference in disease prevalence in S. stellata between shallow and deep reefs (Table S2). Indeed, 73% of the shallow colonies (78 out of 107) and 81% of the deep colonies (43 out of 53) were diseased/bleached (Fig. 2). However, the extent was more than two times greater in deep reefs (median 14.4%) compared to shallow ones (median 6.6%) (Fig. 3b). Given the strong relationship between thermal anomalies and coral disease outbreaks (Miller et al. 2009; Brodnicke et al. 2019), a plausible explanation for this result could be the fact that mesophotic corals live in an environment with more stable temperatures, making them more susceptible to even small temperature variations (Hinderstein et al. 2010; Smith et al. 2016). Consequently, they would be more susceptible to disturbances, including disease infections and reduction in zooxanthellae activity, compared to those living on shallower reefs (Kahng et al. 2014). The long history of exposure to extensive thermal anomalies, high sedimentation, and high nutrient levels (Smith et al. 2008; Camp et al. 2018; Soares et al. 2020a; Chaves-Fonnegra et al. 2021) may have contributed to the greater resistance of shallow corals to diseases compared to deeper corals.
It is important to notice that the shallow reefs of our study region are close to the coast (< 1 km), while the deep reefs are often more than 10 km offshore. This geographic isolation may have precluded gene flow across the shelf and eventually influenced the greater resistance in shallow areas (Weil 2019; Bloomberg and Holstein 2021), but further studies on innate immunity against disease are needed to properly assess this hypothesis. The proximity with the Paraiba coast has also put the shallow colonies in contact with several anthropogenic disturbances over the past five centuries (e.g., pollution, overfishing, mass tourism, sinking of artificial structures; Medeiros et al. 2022), which together changed water quality and sedimentation for corals, their symbionts and pathogens. However, according to our findings, these nearshore abiotic changes were not enough to increase the prevalence and extent of diseases. Similar results have been described for the US Virgin Islands, where many aspects of coral health did not respond to a five-fold increase in the rate of clay and silt sedimentation (Smith et al. 2008). In fact, our findings of S. stellata indicate that the Paraiba mesophotic reefs, although cooler and less exposed to nearshore human stressors, may not act as refuges for this species because their colonies face greater extent of diseases and bleaching [Morais and Santos 2018; see also Bloomberg and Holstein (2021) for a case study with M. cavernosa in the US Virgin Islands]. As demonstrated by Smith et al. (2016), any increase in temperatures above the local mean warmest conditions can lead to coral thermal stress and bleaching, which invalidates the premise that cooler environments—the mesophotic reefs—are protective. The continued monitoring of the diseases and their impact on coral survival and reproduction will be critical to assess this refuge possibility in the South Atlantic.
We recorded five coral diseases and bleaching in our sampling area during the study period (January to February 2017). While only White plague (23%) and White patch (3%) diseases affected M. cavernosa, Dark spot (33.9%), White patch (20.3%), White plague (11.7%), Yellow band (2.4%), Black band (1.2%), and bleaching (7.4%) afflicted S. stellata. Notably, Dark spot was exclusive recorded in shallow habitats, while bleaching was only recorded in deep reefs (Fig. 4). As commonly known, bleaching is periodical and often occur in shallow waters, but did not manifest on the shallow colonies during the study period.

Number of Siderastrea stellata colonies affected by diseases and bleaching in shallow and deep reefs of the Southwestern Atlantic. Photos showing the main diseases of Siderastrea stellata colonies recorded in our study. a Dark spots disease. b White patch disease. c White plague disease. d Yellow band disease. Photographs by Morais J
Identifying the drivers of the five diseases recognized here is urgent, though beyond the scope of this work. We encourage the investigation of the genotype-environment interaction to shed light on the ecoepidemiology underlying the differences between the coral species and between shallow and deep populations of the same species (S. stellata) (Kelley et al. 2021). This is critical to pushing the study of coral diseases in Southwestern Atlantic beyond the observation of bleaching events (Morais et al. 2022).
In summary, our findings revealed remarkable differences in disease prevalence and extent between the two most common reef-building coral species of Southwestern Atlantic. While in general S. stellata is more susceptible to disease than M. cavernosa, this species is also particularly more affected in deep reefs than in shallow reefs. If this difference is enough to put the deep populations at demographic risk is an open question, but it is clear that colonies in shallow and deep reefs show contrasting disease extents. Although we have not measured abiotic conditions, we hypothesize that the more thermally stable environment of deep reefs, with reduced light incidence and increased pressure, potentially make the deep colonies less resistant to temperature fluctuations and disease incidence (Hinderstein et al. 2010). The greater resistance of colonies in shallow reefs may also have a genetic basis promoted by the horizontal isolation between the shallow and deep reefs (Serrano et al. 2014; Bongaerts et al. 2017). Overall, our findings support the notion that MCEs are distinct from shallow reef ecosystems (Soares et al. 2020b). Thus, regulation of economic activities that impact the corals along the depth gradient, such as fishing and tourism, should be implemented accordingly. Because diseases are spread over the entire gradient of depth, any attempt of managing the diseases should consider both shallow and deep reefs to be effective. As recently argued, the ecoepidemiology of coral diseases deserves more space in the research agenda of reef ecosystems, especially in the mesophotic zone (Morais et al. 2022). Survey efforts should describe as many coral and pathogen species as possible, with the specific goals to understand the ecoepidemiology of coral diseases and their interactions with bleaching events (Work and Meteyer 2014; Morais et al. 2022). We also suggest that projections of future distributions of reef-building coral species in the face of climate change (e.g., Principe et al. 2021) should incorporate disease data into the modelling procedures.
References
Bloomberg J, Holstein DM (2021) Mesophotic coral refuges following multiple disturbances. Coral Reefs 40:821–834
Bongaerts P, Riginos C, Brunner R, Englebert N, Smith SR, Hoegh-Guldberg O (2017) Deep reefs are not universal refuges: Reseeding potential varies among coral species. Sci Adv 3:e1602373
Brodnicke OB, Bourne DG, Heron SF, Pears RJ, Stella JS, Smith HA, Willis BL (2019) Unravelling the links between heat stress, bleaching and disease: fate of tabular corals following a combined disease and bleaching event. Coral Reefs 38:591–603
Brooks ME, Kristensen K, Benthem KJV, Magnusson A, Berg CW, Nielsen A, Skaug HJ, Mächler M, Bolker BM (2017) glmmTMB Balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9:378
Bruckner AW (2003) Field guide to Western Atlantic coral diseases and other causes of coral mortality. 1 CD-ROM
Calnan JM, Smith TB, Nemeth R, Kadison E, Blondeau J (2007) Coral disease prevalence and host susceptibility on mid-depth and deep reefs in the United States Virgin Islands. Rev Biol Trop 56:223–234
Camp EF, Schoepf V, Mumby PJ, Hardtke LA, Rodolfo-Metalpa R, Smith DJ, Suggett DJ (2018) The future of coral reefs subject to rapid climate change: lessons from natural extreme environments. Front Mar Sci 5:1–21
Chaves-Fonnegra A, Panassiti B, Smith TB, Brown E, Clemens E, Sevier M, Brandt ME (2021) Environmental and biological drivers of white plague disease on shallow and mesophotic coral reefs. Ecography (cop) 44:1071–1085
Costa CF, Sassi R, Gorlach-Lira K (2008) Zooxanthellae genotypes in the coral Siderastrea stellata from coastal reefs in northeastern Brazil. J Exp Mar Bio Ecol 367:149–152
Darling ES, Alvarez-Filip L, Oliver TA, McClanahan TR, Côté IM (2012) Evaluating life-history strategies of reef corals from species traits. Ecol Lett 15:1378–1386
Ferreira BP, Costa MBSF, Coxey MS, Gaspar a. LB, Veleda D, Araujo M, (2012) The effects of sea surface temperature anomalies on oceanic coral reef systems in the southwestern tropical Atlantic. Coral Reefs 32:441–454
Harborne AR, Rogers A, Bozec Y, Mumby PJ (2017) Multiple stressors and the functioning of coral reefs. Ann Rev Mar Sci 9:445–468
Hartig F (2017) DHARMa: residual diagnostics for hierarchi- cal (multi-level/mixed) regression models
Hinderstein LM, Marr JCA, Martinez FA, Dowgiallo MJ, Puglise KA, Pyle RL, Zawada DG, Appeldoorn R (2010) Theme section on “Mesophotic Coral Ecosystems: Characterization, Ecology, and Management.” Coral Reefs 29:247–251
Howells EJ, Vaughan GO, Work TM, Burt JA, Abrego D (2020) Annual outbreaks of coral disease coincide with extreme seasonal warming. Coral Reefs 39:771–781
Kahng SE, Copus JM, Wagner D (2014) Recent advances in the ecology of mesophotic coral ecosystems (MCEs). Curr Opin Environ Sustain 7:72–81
Kelley ER, Sleith RS, Matz MV, Wright RM (2021) Gene expression associated with disease resistance and long-term growth in a reef-building coral. R Soc Open Sci 8:210113
Leão ZMAN, Kikuchi RKP, Ferreira BP, Neves EG, Sovierzoski HH, Oliveira MDM, Maida M, Correia MD, Johnsson R (2016) Brazilian coral reefs in a period of global change: a synthesis. Brazilian J Oceanogr 64:97–116
Loya Y, Sakai K, Yamazato K, Nakano Y, Sambali H, Van Woesik R (2001) Coral bleaching: the winners and the losers. Ecol Lett 4:122–131
Maynard J, van Hooidonk R, Eakin CM, Puotinen M, Garren M, Williams G, Heron SF, Lamb J, Weil E, Willis B, Harvell CD (2015) Projections of climate conditions that increase coral disease susceptibility and pathogen abundance and virulence. Nat Clim Chang 5:688–694
Medeiros APM, Ferreira BP, Betancur-R R, Cardoso APLR, Matos MRSBC, Santos BA (2022) Centenary shipwrecks reveal the limits of artificial habitats in protecting regional reef fish diversity. J Appl Ecol 59:286–299
Menezes NM, Neves EG, Barros F, de Kikuchi RKP, Johnsson R (2013) Intracolonial variation in Siderastrea de Blainville, 1830 (Anthozoa, Scleractinia): taxonomy under challenging morphological constraints. Biota Neotrop 13:108–116
Miller J, Muller E, Rogers C, Waara R, Atkinson A, Whelan KRT, Patterson M, Witcher B (2009) Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs 28:925–937
Montilla LM, Ascanio A, Verde A, Croquer A (2019) Systematic review and meta-analysis of 50 years of coral disease research visualized through the scope of network theory. PeerJ 7:e7041
Morais J, Cardoso APL, Santos BA (2022) A global synthesis of the current knowledge on the taxonomic and geographic distribution of major coral diseases. Environ Adv 8:100231
Morais J, Medeiros APM, Santos BA (2018) Research gaps of coral ecology in a changing world. Mar Environ Res 140:243–250
Morais J, Morais RA, Tebbett SB, Pratchett MS, Bellwood DR (2021) Dangerous demographics in post-bleach corals reveal boom-bust versus protracted declines. Sci Rep 11:18787
Morais J, Santos BA (2018) Limited potential of deep reefs to serve as refuges for tropical Southwestern Atlantic corals. Ecosphere 9:e02281
Principe SC, Acosta AL, Andrade JE, Lotufo TMC (2021) Predicted shifts in the distributions of atlantic reef-building corals in the face of climate change. Front Mar Sci 8:1–22
Randall CJ, Van Woesik R (2015) Contemporary white-band disease in Caribbean corals driven by climate change. Nat Clim Chang 5:375–379
Raymundo LJ, Couch CS, Bruckner AW, Harvell CD (2008) Coral Disease Handbook Guidelines for Assessment
Rogers AD, Kemp KM, Davies AJ, Lisa Taylor M (2015) The diseases of deep-water corals. Diseases of Coral. John Wiley & Sons Inc, Hoboken, NJ, pp 416–441
Safuan M, Wee HB, Hii Y-S, Liew H-C, Bachok Z (2015) Optimization of coral video transect technique for coral reef survey : comparison with intercept transect technique. Open J Mar Sci 5:379–397
Serrano XM, Baums IB, O’Reilly K, Smith TB, Jones RJ, Shearer TL, Nunes FLD, Baker AC (2014) Geographic differences in vertical connectivity in the Caribbean coral Montastraea cavernosa despite high levels of horizontal connectivity at shallow depths. Mol Ecol 23:4226–4240
Smith TB, Gyory J, Brandt ME, Miller WJ, Jossart J, Nemeth RS (2016) Caribbean mesophotic coral ecosystems are unlikely climate change refugia. Glob Chang Biol 22:2756–2765
Smith TB, Nemeth RS, Blondeau J, Calnan JM, Kadison E, Herzlieb S (2008) Assessing coral reef health across onshore to offshore stress gradients in the US Virgin Islands. Mar Pollut Bull 56:1983–1991
Soares M de O, Tavares TCL, Carneiro PB de M (2018) Mesophotic ecosystems: Distribution, impacts and conservation in the South Atlantic. Divers Distrib 25:ddi.12846
Soares M de O, Cruz ICS, Santos BA, Tavares TCL, Garcia TM, Menezes N, Lopes BD, de Araújo JT, Gurgel ALAR, Rossi S (2020a) Marginal Reefs in the Anthropocene: They Are Not Noah’s Ark. Perspectives on the Marine Animal Forests of the World. pp 87–128, Springer International Publishing, Cham
Soares M de O, Araújo JT de, Ferreira SMC, Santos BA, Boavida JRH, Costantini F, Rossi S (2020b) Why do mesophotic coral ecosystems have to be protected? Sci Total Environ 726:138456
Sully S, Burkepile DE, Donovan MK, Hodgson G, van Woesik R (2019) A global analysis of coral bleaching over the past two decades. Nat Commun 10:1264
Team RC (2020) R: A Language and Environment for Statistical Computing
Weil E (2019) Disease Problems in Mesophotic coral communities. In: Loya Y, Puglise KA, Bridge TCL (eds) Mesophotic Coral Ecosystems. Springer International Publishing, Cham, pp 779–800
Williams SM, García-Sais J, Sabater-Clavell J (2021) Prevalence of Stony Coral Tissue Loss Disease at El Seco, a Mesophotic Reef System off Vieques Island, Puerto Rico. Front Mar Sci 8:1–13
Work T, Meteyer C (2014) To understand coral disease, look at coral cells. EcoHealth 11:610–618
Acknowledgements
We thank Fundação Grupo Boticario de Proteçao à Natureza (Grant number 1044-20152) for funding this study and Mar Aberto Dive Center for logistical support. The Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) provided research grant to BAS (312178/2019-0). We also thank O. A. Silva, M. Russi, I. Just, E. F. Gonçalves, I. Medeiros and S. B. Tebbett.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Topic Editor Steve Vollmer
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Morais, J., Santos, B.A. Prevalence and extent of coral diseases in shallow and mesophotic reefs of the Southwestern Atlantic. Coral Reefs 41, 1317–1322 (2022). https://doi.org/10.1007/s00338-022-02287-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-022-02287-y