
Abstract
To survive in nutrient-poor waters corals rely on a symbiotic association with intracellular microalgae. However, increased sea temperatures cause algal loss—known as coral bleaching—often followed by coral death. Some of the most compelling evidence in support of the ‘oxidative stress theory of coral bleaching’ comes from studies that exposed corals, cultures of their algal endosymbionts, or the coral model Exaiptasia diaphana to exogenous antioxidants during thermal stress. Here, we replicate these experiments using E. diaphana with the addition of the antioxidants ascorbate + catalase, catechin, or mannitol under ambient and elevated temperatures along with an antioxidant-free control. In the absence of exogenous antioxidants, E. diaphana exposed to elevated temperatures bleached with no change in reactive oxygen species (ROS) levels associated with their microalgal cells. Ascorbate + catalase and mannitol treatments rescued the anemones from bleaching, although microalgal ROS levels increased in these antioxidant treatments under elevated temperature conditions. While bleaching was not associated with changes in net ROS for the intracellular algal symbionts, it is evident from our findings that excess ROS is connected to the bleaching phenotype as exogenous antioxidants were successful in mitigating the effects of thermal stress in cnidarians. This understanding may assist applied research that aims to reduce the impact of climate change on coral reefs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The severe loss of coral cover and subsequent decline of coral reefs over the past several decades are primarily driven by climate change-related increases in sea surface temperature (SST) (Hughes et al. 2017; Stuart-Smith et al. 2018). A spike in SST during unusually hot summers can trigger coral bleaching (Hughes et al. 2018; Hoegh-Guldberg et al. 2019), the dissociation of the coral host and its endosymbiotic, dinoflagellate algae (Symbiodiniaceae). As Symbiodiniaceae provide their coral hosts with most of their organic carbon requirements (Muscatine and Porter 1977; Tremblay et al. 2014), this loss can lead to coral death. Mass bleaching occurred on the Great Barrier Reef (GBR) in 1998, 2002, 2016, 2017, and 2020 and caused extensive coral mortality, with the 2016 and 2020 events having been the most severe and widespread recorded (Hughes and Pratchett 2020).
Several hypotheses describe the mechanisms that drive bleaching (Weis 2008; Cunning and Baker 2012; Wiedenmann et al. 2012; Wooldridge 2013), with a common theme being the transfer of accumulated reactive oxygen species (ROS) from the algal symbiont to the surrounding host cell. ROS are normal products of photosynthesis; the light-triggered splitting of water molecules in the oxygen-evolving complex (OEC) and subsequent transfer of electrons from photosystem II (PSII) to photosystem I (PSI) generate ROS in chloroplasts (Trubitsin et al. 2014; Szabó et al. 2020). Singlet oxygen (1O2) can be formed via energy transfer when ground-state molecular oxygen interacts with chlorophyll triplets (Krieger-Liszkay 2005), a light-dependent process (Rehman et al. 2016). The major site of superoxide (O2•−) production is the thylakoid membrane-bound primary electron acceptor of PSI (as reviewed in Szabó et al. 2020). This O2•− is further converted to hydrogen peroxide (H2O2) within the chloroplast mostly by superoxide dismutase (SOD) (Asada 1987). Hydroxyl radicals (OH) are produced in the Haber–Weiss or Fenton reactions through the interaction of H2O2 and O2•−or directly from H2O2 in the presence of transition metals (Halliwell and Gutteridge 1990).
Symbiodiniaceae have evolved efficient strategies to cope with the accumulation of ROS that are by-products of photosynthesis (Warner and Suggett 2016), but these systems become overwhelmed during periods of thermal stress when ROS production is increased. Elevated temperatures can enhance the activity of photosynthetic enzymes (Iglesias-Prieto et al. 1992), driving increases in photosynthetic rates for in hospite Symbiodiniaceae (Castillo and Helmuth 2005; Hoadley et al. 2016). ROS produced from this amplified photosynthetic activity can trigger the oxidation of essential photosynthetic molecules, such as the D1 protein in PSII (Lesser 1996; Krieger-Liszkay 2005; Wang et al. 2011; Mathur et al. 2014). The damaged PSII is normally repaired, but this process is sensitive to heat (Takahashi et al. 2004, 2009) and excess ROS (Nishiyama et al. 2006), which gives rise to reductions in PSII efficiency, a metric noted for predicting bleaching susceptibility (Takahashi et al. 2008; Dang et al. 2019b; Voolstra et al. 2020). An increase in SST can also affect the stability of thylakoid membranes (Hill et al. 2009; Díaz-Almeyda et al. 2011), preceding the physiological damage from ROS to which cells actually respond (Tchernov et al. 2004; Downs et al. 2013). This damage to membrane integrity can disturb PSI, PSII, and the OEC, while simultaneously inducing the production of ROS (Farooq et al. 2016). The resultant damage forms a positive feedback loop, leading the even further production of ROS (Szabó et al. 2020).
Thermal stress can also inflate cnidarian host ROS production. ROS generation by the mitochondrial electron transport chain (mETC) increases due to thermally induced high respiration or electron transport inhibition, independent of photosynthetic activity (Oakley et al. 2017; Oakley and Davy 2018). Hawkins and Warner (2017) found that activity of the mETC in host and symbiont mitochondria became destabilized in E. diaphana during thermal bleaching, the consequences of which can include heightened O2•− generation (Murphy 2009). Regardless of the source, if not scavenged by the antioxidant network, the damage caused by ROS may trigger signaling cascades ending with a breakdown of host–algae symbiosis (Weis 2008).
Many innovative approaches have been proposed to mitigate bleaching and enhance coral survival during thermal stress (van Oppen et al. 2015; Damjanovic et al. 2017; Peixoto et al. 2017; Dungan et al. 2021, 2022; Maire and van Oppen 2021). Directly targeting ROS has been investigated, with promising but limited evidence that exposure to exogenous antioxidants can reduce ROS levels, mitigate bleaching, or extend survival time in thermally stressed corals (Lesser 1997; Nesa and Hidaka 2008, 2009; Majerová and Drury 2021). In addition, the photosynthetic performance of thermally stressed Symbiodiniaceae has been shown to be improved and net ROS to be decreased within hours of receiving a single antioxidant dose (Lesser 1996; Motone et al. 2020).
In this study, we applied the three exogenous antioxidants (ascorbate + catalase, catechin, and mannitol; Table 1) used in former studies to concurrently assess their effect on bleaching and net ROS in the coral model, the sea anemone Exaiptasia diaphana. We define net ROS as the relative difference between ROS produced and ROS scavenged by enzymatic and non-enzymatic antioxidants and associated with the Symbiodiniaceae cell wall and symbiosome lumen, the space between the host-derived membranes in which the algal symbiont resides (symbiosome) and the Symbiodiniaceae cell wall. This experiment allowed us to address the following questions:
-
(1)
Is thermal bleaching in E. diaphana, as defined by the loss of Symbiodiniaceae cells, accompanied by elevated net ROS?
-
(2)
Can the addition of exogenous antioxidants mitigate thermal bleaching?
-
(3)
Do exogenous antioxidants harm the E. diaphana holobiont?
Materials and methods
Experimental setup
Anemones from Great Barrier Reef (GBR)-sourced E. diaphana genotype AIMS3 were randomly selected from The University of Melbourne culture collection (n = 256) (Dungan et al. 2020). Anemones were equally distributed among 12 × 300-mL lidded glass culture jars and placed in an experimental incubator (Model No. LE-509, Thermoline Scientific, Australia) fitted with white light-emitting diodes (15–20 μmol photons m–2 s–1) on a 12-h/12-h light/dark cycle. During acclimation (11 weeks), anemones were maintained in reconstituted Red Sea Salt™ seawater (RSS, R11065, Red Sea, USA) at ~ 34 parts per thousand (ppt) salinity and fed ad libitum twice weekly with freshly hatched Artemia salina (Salt Creek, Premium GSL, USA). Jars were cleaned 3–5 h after feeding by loosening algal debris with RSS pressure applied through sterile plastic pipettes followed by full RSS changes. Anemones were considered acclimated once their photosynthetic efficiency (Fv/Fm), as measured by pulse-amplitude-modulated (PAM) fluorometry with an imaging-PAM system (IMAG-MAX/L, Walz Heinz, Germany), attained stable, healthy levels as found in the stock culture collection. Seawater temperatures were monitored using submersible data loggers (HOBO UA-001–08) (Fig. S1).
Three exogenous antioxidant treatments were assessed for their effect on bleaching and net ROS, compared to a no-antioxidant control (Table 1). Net ROS was quantified using a fluorescent stain that is activated when oxidized and quantified using flow cytometry. Visual health scores were used to evaluate holobiont health after antioxidant dosing. Stock antioxidants were made with Milli-Q water at 100 × the target concentration and stored at − 20 °C in several aliquots. On the day of dosing, thawed antioxidant aliquots were diluted in 0.22-µm-filtered RSS (fRSS) to the target concentration.
One day before the experiment commenced, anemones were transferred from the lidded glass jars to sterile 12 well plates in fRSS, with eight anemones per plate (one anemone per well) in a random arrangement for a total of 32 plates (4 plates per antioxidant/temperature combination). Given the GBR E. pallida are extremely light sensitive, low light levels were used. When light levels > 20 µmol photons m–2 s–1 were tested, Fv/Fm values did not recover and we therefore conducted the experiment with 15–20 µmol photons m–2 s–1 on a 12-h/12-h light/dark cycle. In the elevated temperature treatment, anemones at 26 °C were ramped up by 1 °C day−1, at 11 h into the light cycle (Fig. 1). By Day 6, elevated temperature treatments reached 32 °C, on Day 10 the temperature was increased to 33 °C, then to 34 °C on Days 11 and 12, respectively. Starting on Day 0, anemones were immersed in respective antioxidant treatments as described in Table 1. fRSS plus antioxidant changes were completed every second day. Anemone plates were randomly rearranged each day inside the incubator to remove plate position as a confounding factor.

Antioxidant dosing and sampling schedule for each antioxidant–temperature combination. Temperature conditions: ambient (26 °C, blue) and elevated (26 °C–34 °C, red). Antioxidant water changes were completed every second day for the duration of the experiment, beginning on Day 0. Stars indicate processing of five anemones from each treatment for Symbiodiniaceae cell density measurements, chl a fluorescence, and ROS quantification. An additional eight anemones were collected on Day 13 for Symbiodiniaceae cell density determination only. Health scoring and imaging PAM were carried out every day at approximately 5 h into the light cycle
Symbiodiniaceae photochemical efficiency measurement
Photochemical efficiency (Fv/Fm) of PSII in Symbiodiniaceae is a commonly used proxy for general holobiont health as lowered Fv/Fm indicates PSII damage (Ralph et al. 2016). Fv/Fm was measured by imaging-PAM fluorometry of the anemone bodies and proximal tentacles at 5 h into the light cycle and after 30 min of dark adaptation. PAM settings were: saturating pulse intensity 8, measuring light intensity 2 (frequency 1), damping 2, and gain 2 to achieve a F0 of 0.1–0.3.
Anemone tissue processing
On Day 0, five anemones were sacrificed for Symbiodiniaceae cell counts, host protein quantification, chl a fluorescence, and ROS quantification to document initial control conditions. On Day 13, five anemones per treatment were killed for chl a fluorescence and ROS quantification, while 13 anemones in each treatment were used to measure Symbiodiniaceae cell counts and host protein concentrations. Anemones were individually homogenized in a sterile glass homogenizer in 1 mL of fRSS from their well. Homogenate for chl a fluorescence and ROS quantification was processed immediately, while aliquots for Symbiodiniaceae cell density and host protein measurement were stored at − 20 °C until quantification.
Symbiodiniaceae cell density and host protein analysis
The − 20 °C preserved homogenate was centrifuged at 5000 × g for 5 min at 4 °C to pellet the Symbiodiniaceae, and 100 µL of the supernatant collected for host protein analysis. Pelleted Symbiodiniaceae were washed with 200 µL fRSS and centrifuged at 5000 × g for 5 min at 4 °C and then resuspended in 200 µL fRSS. Triplicate cell counts (cells mL–1) were calculated with an automated cell counter (Countess II FL, Life Technologies, Australia) from 10 µL of sample. Cell counts were normalized to host protein (mg mL–1) determined by the Bradford assay (Bradford 1976). Each protein sample was assayed in triplicate and measured at 595 nm (EnSpire MLD2300 plate reader, PerkinElmer, Australia) against bovine serum albumin standards (500–0207, Bio-Rad, Australia).
ROS and chlorophyll a fluorescence quantification
Anemone ROS levels were quantified with CellROX® Orange (C10443, Thermo Fisher, Australia). A volume of 4 µL of CellROX® Orange (stock at 500 µM in dimethyl sulfoxide (DMSO)) was added to 400 µL of the sample and 4 µL of DMSO was added to the remaining 400 µL as a control. Both were incubated at sample experimental temperatures for 60 min. Samples were filtered through a 40-µm cell strainer (pluriSelect) and vortexed before processing with a CytoFLEX LX flow cytometer (Beckman Coulter) at a speed of 10 µL min−1. Autofluorescent events strongly excited by the 488 nm laser at emissions 525 ± 20 nm and 690 ± 25 nm were used to gate the Symbiodiniaceae cells and separate them from anemone debris. A forward scatter-height/side scatter-height (FSC-H/SSC-H) plot was then used to select homogenous cells, and singlets were gated on a FSC-H/FSC-Area plot. The CellROX® Orange signal was quantified on singlets gated on previous plots with 561 nm excitation (emission 585 ± 21 nm) (Fig. S2). At least 5000 Symbiodiniaceae singlets per sample were processed, except in samples where Symbiodiniaceae density was low because of advanced bleaching, where between 700 and 2000 singlets per sample were processed. For each sample, the median CellROX® Orange signal of the unstained sample was subtracted from the corresponding stained sample to account for symbiont autofluorescence. Chl a fluorescence was also quantified from the gated singlets using 488 nm excitation (emission 690 ± 25 nm) (Bouchard and Yamasaki 2008).
Prior to ROS quantification and to confirm the flow cytometry results, stained and unstained samples used for flow cytometry were pipetted onto poly-L-lysine (0.1% w v−1 in Milli-Q water)-coated coverslip-bottom well slides (Ibidi). Slides were then visualized with an inverted confocal laser scanning microscope (CLSM, Nikon A1R) by excitation at 561 nm (emission 595 ± 20 nm).
Anemone health
Anemones were assessed daily for overall health via a qualitative health score (Fig. 2), immediately following imaging-PAM fluorometry.

Anemone health was visually scored on a scale of 4 to 0. Anemones with a score of 4 (top) had fully extended bodies and tentacles, actively fed, and responded to water movement; anemones with a score of 3 (left) had curled tentacles and partially withdrawn bodies but actively fed and responded to water movement; anemones with a score of 2 (bottom) had fully retracted bodies with shortened tentacles and showed reduced response to water movement and reduced feeding; anemones with retracted tentacles and/or did not respond to water movement or feeding, but were not dead, were given a health score of 1 (right); dead anemones received a score of 0. No scale bar
Data analysis
All data were analyzed in R v4.0.3 (R Core Team 2018). Statistical tests were considered significant at α = 0.05. Dark-adapted Fv/Fm, Symbiodiniaceae cell density, chl a fluorescence, and ROS measurements were plotted using the R package ggplot2 (Wickham 2020) with data separated by temperature condition. These physiology parameters were analyzed for overall differences between treatments on Day 13 using linear mixed-effects analyses (with lme in the R package nlme; Pinheiro et al. 2019), using temperature, antioxidant, and their interaction as fixed effects and well plate as a random effect. Post hoc pairwise comparisons were made using Tukey’s honestly significant difference test (Tukey 1949) in the R package emmeans (Searle et al. 1980) with Tukey’s adjustment for multiple comparisons to identify where differences occurred. Exploratory graphical analyses of residuals were used to confirm that assumptions of homogeneity of variance, normality, and independence were met for all linear regression modeling. Symbiodiniaceae cell count data were log-transformed to meet these assumptions.
Results
Bleaching, photosynthetic efficiency, and chlorophyll a
On Day 13, Symbiodiniaceae density for E. diaphana was significantly reduced under elevated compared to ambient temperature (F(1,24) = 25.78, p < 0.0001). However, pairwise comparisons showed that this difference was not consistent between antioxidant treatments. There was a significant reduction in Symbiodiniaceae cell density between ambient and elevated temperature for the control (t(24) = 4.27, p = 0.0003) and catechin-treated anemones (t(24) = 2.53, p = 0.0186), with no significant changes for ascorbate + catalase or mannitol-treated anemones (Fig. 3A).

Bleaching metrics of A Symbiodiniaceae cell density normalized to host protein, B photosynthetic efficiency (Fv/Fm), C Chl a fluorescence, and D net ROS D measured on Day 13 from anemones reared at ambient (light colors) or elevated (dark colors) temperature for the no-antioxidant control (purple), ascorbate + catalase (orange), catechin (green), and mannitol (blue) antioxidant treatments. Chl a fluorescence of Symbiodiniaceae cells was measured as median emitted fluorescent signal (488/690 (excitation/emission)), with n = 5 per treatment. Net ROS was measured as signal intensity of CellROX® Orange stained Symbiodiniaceae cells sorted from anemone homogenate, where signal intensity was normalized by subtracting the median signal of unstained cells to control for autofluorescence. n = 5 per treatment for net ROS. Asterisks indicate significant pairwise differences
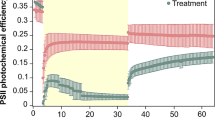
Photosynthetic efficiency was measured each day for the duration of the experiment (Fig. S3). A linear mixed effect model revealed a significant interaction of antioxidant dose and temperature on Fv/Fm on Day 13 (F(3,22) = 3.06, p = 0.0494). Pairwise comparisons showed that anemones in the ambient temperature condition had a significantly higher Fv/Fm compared to those exposed to elevated temperature for each antioxidant treatment. There were no significant differences between antioxidant treatments under ambient temperature conditions (Table S1). Under elevated conditions, however, mannitol-treated anemones had a significantly higher Fv/Fm compared to the no-antioxidant controls (Table S2; Fig. 3B).
There was a significant interaction of antioxidant treatment and temperature on chl a fluorescence (F(3,24) = 4.42, p = 0.013) (Fig. 3C). Only ascorbate + catalase-treated anemones had a significant response to temperature; these anemones at ambient temperature had a significantly higher chl a fluorescence compared to the elevated temperature (t(24) = 4.066, p = 0.0004). In both ambient and elevated conditions, anemones in the ascorbate + catalase treatment had significantly higher chl a fluorescence compared to all other antioxidant treatments and controls (p < 0.0001).
Net ROS in Symbiodiniaceae cell wall and symbiosome lumen
CLSM confirmed that ROS increased the CellROX® Orange signal and that antioxidant addition reduced CellROX® Orange signal in the Symbiodiniaceae cell wall and symbiosome lumen (Fig. S4). On Day 13, net ROS varied significantly with the interaction of antioxidant treatment and temperature (F(3,24) = 8.90, p = 0.0004) (Fig. 3D). At elevated temperature, ascorbate + catalase- and mannitol-treated anemones had higher CellROX® Orange staining compared to the corresponding antioxidant treatment at ambient temperature (tasc+cat(24) = 7.31, p < 0.0001; tman(24) = 3.62, p = 0.0014). There were no significant differences, however, in net ROS between ambient and elevated temperature for the control (no antioxidant) or catechin treatments. At elevated temperature, only ascorbate + catalase-treated anemones had significantly increased CellROX® Orange signal compared to the no-antioxidant controls (t(24) = 7.11, p < 0.0001), with no differences in net ROS at ambient temperature between antioxidant treatments.
Anemone health response to antioxidants
By Day 7, anemones in the ascorbate + catalase treatment in both temperatures showed a drastic decline in health. The health score rating for these two groups continued to decline until the conclusion of the experiment. By Day 11, all elevated temperature anemones displayed reduced health compared to the ambient anemones for each antioxidant treatment. This difference was subtle, however, for all but the ascorbate + catalase treatment anemones. Anemones in the ascorbate + catalase were also visibly darker than all the other treatments, regardless of temperature.
Discussion
Thermal bleaching was not accompanied by elevated ROS in algal symbionts
In the absence of exogenous antioxidants, E. diaphana exposed to elevated temperatures had significantly reduced Symbiodiniaceae cell densities compared to control anemones at ambient temperature, thus confirming that elevated temperature induced bleaching. Moreover, although there was no evidence of chl a pigment bleaching, there was a significant decline in photochemical health (Fv/Fm). However, despite the observed heat-related impacts, we detected no significant change in net ROS for sorted Symbiodiniaceae cells.
The observed bleaching response in the absence of elevated Symbiodiniaceae cell wall and symbiosome lumen-associated ROS corresponds with other studies that show that bleaching can occur without a flux of photosynthetically produced ROS from Symbiodiniaceae to the surrounding host cell (Tolleter et al. 2013) and with discrepancies in enzymatic antioxidant activity between host and symbiont tissue portions (Krueger et al. 2015), raising questions about the importance of symbiont-derived ROS in initiating cnidarian bleaching. Nielsen et al. (2018) found that bleaching independent of ROS can occur with thermal stress, corroborating field observations that coral ROS production is unrelated to bleaching status (Diaz et al. 2016).
Despite the literature reporting variability in net ROS levels from Symbiodiniaceae when exposed to elevated temperatures, there is support for the production of H2O2 in the thylakoid lumen and stroma during photosynthesis (Szabó et al. 2020). While H2O2 can easily pass through chloroplast membranes and accumulate in the Symbiodiniaceae cytosol (Mubarakshina et al. 2010; Borisova et al. 2012), the theory that H2O2 leaks from the Symbiodiniaceae cytosol, through the symbiosome lumen, and into the coral host cell is incomplete (Oakley and Davy 2018). Dye trace measurements have shown that ROS produced by isolated Symbiodiniaceae cells can leak into their surrounding media (Tchernov et al. 2004) and symbiotic Stylophora pistillata produce twice as much O2•− as their aposymbiotic counterparts in the light (Saragosti et al. 2010), but no studies have tracked the production and movement of ROS from Symbiodiniaceae in hospite. With the recent availability of cell cultures (Kawamura et al. 2021), real-time imaging to track the production and movement of ROS in vivo becomes highly feasible.
Exogenous antioxidants prevented bleaching in E. diaphana
Antioxidant treatment resulted in bleaching resistance in the case of mannitol and ascorbate + catalase inoculation, but not for catechin dosed anemones. Treatment with mannitol also led to a significant increase in Fv/Fm under elevated temperature compared to the no-antioxidant controls. Fv/Fm has been described as a proxy for thermal stress response in corals (Ralph et al. 2016; Voolstra et al. 2020) and Symbiodiniaceae cultures (Ragni et al. 2010; Goyen et al. 2017; Dang et al. 2019a) with higher or retained Fv/Fm values, indicating increased bleaching resistance during stress events. The stability in photochemistry was accompanied by stable algal densities.
Ascorbate and mannitol both protect against oxidative damage by·OH, which can be produced in eukaryotic cells via the Haber–Weiss or Fenton reactions through the interaction of H2O2 and O2•− or directly from H2O2 in the presence of transition metals (Halliwell and Gutteridge 1990). H2O2 is ubiquitously formed during cellular respiration and its rate of production in eukaryotic mitochondria has been linked with metabolic state (Boveris et al. 1972), which can be severely impacted in corals experiencing thermal stress (Bahr et al. 2018; Oakley and Davy 2018). Further, O2•− can be generated by the mETC during thermal stress in eukaryotic cells (Murphy 2009). Oxidative damage from excess ROS, such as ·OH or O2•−, may be in the form of enzyme inactivation, lipid peroxidation, or DNA damage (Baird et al. 2009). Host DNA damage followed by programmed cell death is associated with coral bleaching (Weis 2008; Majerová et al. 2020; Majerová and Drury 2021). If the exogenous addition of mannitol or ascorbate leads to the quenching of ·OH or O2•− in cnidarian host tissues, we would expect increased resistance to oxidative stress and reduced DNA damage, potentially explaining why mannitol and ascorbate + catalase addition mitigated bleaching.
In addition to our work here on E. diaphana, a handful of other studies have also observed beneficial effects of exogenous mannitol or ascorbate + catalase addition to corals during periods of thermal stress. Agaricia tenuifolia colonies exposed to elevated temperatures in the presence of mannitol or ascorbate + catalase lost significantly fewer Symbiodiniaceae cells compared to no-antioxidant controls with photosynthesis and respiration rates indistinguishable from their pre-temperature stress rates (Lesser 1997). The addition of mannitol to coral cell cultures extended Pavona divaricata tissue survival at elevated temperature (Nesa and Hidaka 2009), but ROS levels were not recorded. Nesa and Hidaka (2009) also found that an ascorbate + catalase treatment significantly increased survival time for Fungia sp. tissue cultures at 31 °C. Another study revealed that the addition of mannitol was associated with a significant decrease in host P. divaricata DNA damage at elevated temperatures compared to the no-antioxidant controls (Nesa and Hidaka 2008). More recently, mannitol addition to Pocillopora acuta reduced host DNA damage during thermal stress to levels similar to the ambient controls (Majerová and Drury 2021). The cumulative results from these studies and our current work suggest that increased biologically available antioxidants could potentially mitigate bleaching and prolong the life of corals during periods of thermal stress, a promising avenue for assisted evolution approaches.
Ascorbate + catalase treatment was lethal to anemones
Based on the promising results of previous studies (Lesser 1996, 1997; Nesa and Hidaka 2009), we dosed anemones with a combination of ascorbate (125 µM) and catalase (250 U ml−1). Ascorbate is a water-soluble non-enzymatic antioxidant with limited scavenging of ·OH but dominant in vivo scavenging of O2•− and 1O2 (Zhitkovich 2021). Catalase on the other hand is an enzymatic antioxidant efficient at scavenging H2O2. Ascorbate and catalase were used in concert by Lesser (1996) to prevent the accumulation of peroxide as ascorbate was being oxidized (M. Lesser, pers. comm.). Anemones in this treatment had significantly increased chl a fluorescence, regardless of temperature. Exogenous addition of ascorbate to plants (i.e., maize, wheat, barley) has resulted in increased chlorophyll content (Akram et al. 2017), suggesting that ascorbate is driving the change we see here in Symbiodiniaceae chl a fluorescence. An increase in chl a per Symbiodiniaceae cell can be used to cover higher metabolic demands under lower photosynthetic efficiencies (Iglesias-Prieto et al. 1992). This chl a boost in ascorbate + catalase-treated anemones with the accompanying capacity to maintain higher metabolic rates may explain why these anemones did not bleach but had significantly higher net ROS levels under elevated temperatures. However, the health of the ascorbate + catalase dosed anemones declined after Day 7, regardless of temperature. Additionally, several anemones in this treatment group, particularly under elevated temperature, died prior to sampling, driving the low sample size available for Symbiodiniaceae cell density and Fv/Fm measurements. For those anemones that survived, this group had the lowest Fv/Fm values on Day 13.
Mitigation of bleaching by mannitol was accompanied by higher net ROS
Compared to ambient temperature, there was no change in Symbiodiniaceae cell density for mannitol-treated anemones under elevated temperature conditions, suggesting that anemones did not bleach. Anemones receiving this treatment, however, had a significant increase in Symbiodiniaceae-associated net ROS from ambient to elevated temperature. Thus, the mitigation of bleaching by mannitol was not associated with lower Symbiodiniaceae ROS levels. It is possible that the presence of mannitol reduced the natural production of antioxidants by the host and symbionts, thus allowing ROS to accumulate around the Symbiodiniaceae cell wall and in the symbiosome lumen, though this is speculative and warrants further investigation. Higher ROS levels in these anemones that did not bleach under elevated temperature challenge the ‘oxidative stress theory of coral bleaching’ in the context that ROS fluxes from Symbiodiniaceae alone were not enough to cause bleaching.
Role for host-derived ROS in cnidarian bleaching
The oxidative theory of coral bleaching postulates that H2O2 derived from photo-oxidative stress in Symbiodiniaceae might leak into the host tissue when symbiont antioxidant defenses are overwhelmed in heat-stressed corals (Downs et al. 2002). Our results show that anemone bleaching does not necessarily involve a connection to ROS fluxes between host and symbiont as there were no significant increases in ROS associated with Symbiodiniaceae in anemones that bleached. Changes in Symbiodiniaceae net ROS as a result of exposure to elevated temperature are highly variable by species (Lesser 1996, 2019; Suggett et al. 2008; McGinty et al. 2012; Rehman et al. 2016; Wietheger et al. 2018), with observations of bleaching in the absence of photosynthetically produced ROS (Tolleter et al. 2013). It is evident from our findings and other antioxidant dosing studies, however, that excess ROS is connected to the bleaching phenotype as exogenous addition of ascorbate + catalase (this study; Lesser 1997; Nesa and Hidaka 2009) or mannitol (this study; Lesser 1997; Nesa and Hidaka 2008, 2009; Majerová and Drury 2021) has been successful in mitigating the effects of thermal stress in cnidarians. We speculate that host-derived ROS levels may play a larger role in the coral bleaching response than previously thought as they would increase in parallel with respiration rates in thermally stressed corals (Edmunds 2004; Oakley and Davy 2018; Radecker et al. 2021). This hypothesis is supported by increased scavenging of H2O2 in host tissue at high temperatures (33 °C) in both Acropora millepora and Montipora digitata (Krueger et al. 2015), and the consistent upregulation of host genes encoding ROS scavengers in response to elevated temperatures (Louis et al. 2017).
Limitations of the study
While ascorbate + catalase mitigated bleaching, this antioxidant treatment caused extensive mortality. High concentrations of antioxidants can be detrimental in biological systems (Villanueva and Kross 2012). Future experiments should explore their application independently with toxicity tests to determine how each antioxidant might contribute to bleaching rescue and which concentration would be most suited. While CellROX® reagents have been used extensively in cnidarian studies (Levin et al. 2016; Cziesielski et al. 2018; Gegner et al. 2019; Buerger et al. 2020; Radecker et al. 2021; Snyder et al. 2021), the rate constants of these reactants with different ROS have not been published. Future studies should use probes to track specific ROS (Mattila et al. 2015) in vivo for coral cell cultures to elucidate how each ROS individually may be involved in the bleaching process. While this study provides support for the role of non-photosynthetically produced ROS in cnidarian bleaching, the mechanism by which it is produced and leads to bleaching is still one of coral biology’s greatest mysteries.
Studies on the establishment, maintenance, and breakdown of E. diaphana–Symbiodiniaceae symbiosis have proven vital to coral research, but we do recognize that E. diaphana and corals have their clear differences. The symbiosis between E. diaphana and their Symbiodiniaceae endosymbionts is facultative, unlike the obligate symbiosis between scleractinian corals and Symbiodiniaceae. GBR-sourced E. diaphana require very low light levels, such as provided in shaded or deep reef habitats; conversely, shallow unshaded GBR coral habitats can average 1400 µmol photons m–2 s–1 over the course of a day (Bainbridge 2017). These differences in light levels would influence photosynthetic activity and resultant ROS production. Given the ecological differences between E. diaphana and reef building corals, we caution against direct comparisons between the two. However, E. diaphana will remain crucial to the development of tools to document and understand symbiosis as well as in developing and trialling strategies to mitigate the effects of climate change on corals.
Data availability
Detailed protocols, full data sets, and analysis code are available via Dryad, https://doi.org/10.5061/dryad.69p8cz92d
References
Akram NA, Shafiq F, Ashraf M (2017) Ascorbic acid-a potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front Plant Sci 8:613
Asada K (1987) Production and scavenging of active oxygen in photosynthesis. Photoinhibition 227–287
Bahr KD, Rodgers KS, Jokiel PL (2018) Ocean warming drives decline in coral metabolism while acidification highlights species-specific responses. Mar Biol Res 14:924–935
Bainbridge SJ (2017) Temperature and light patterns at four reefs along the Great Barrier Reef during the 2015–2016 austral summer: understanding patterns of observed coral bleaching. Journal of Operational Oceanography 10:16–29
Baird AH, Bhagooli R, Ralph PJ, Takahashi S (2009) Coral bleaching: the role of the host. Trends Ecol Evol 24:16–20
Borisova MM, Kozuleva MA, Rudenko NN, Naydov IA, Klenina IB, Ivanov BN (2012) Photosynthetic electron flow to oxygen and diffusion of hydrogen peroxide through the chloroplast envelope via aquaporins. Biochim Biophys Acta Bioenerg 1817:1314–1321
Bouchard JN, Yamasaki H (2008) Heat stress stimulates nitric oxide production in Symbiodinium microadriaticum: a possible linkage between nitric oxide and the coral bleaching phenomenon. Plant Cell Physiol 49:641–652
Boveris A, Oshino N, Chance B (1972) The cellular production of hydrogen peroxide. Biochem J 128:617–630
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Buerger P, Alvarez-Roa C, Coppin CW, Pearce SL, Chakravarti LJ, Oakeshott JG, Edwards OR, van Oppen MJH (2020) Heat-evolved microalgal symbionts increase coral bleaching tolerance. Sci Adv 6
Castillo K, Helmuth B (2005) Influence of thermal history on the response of Montastraea annularis to short-term temperature exposure. Mar Biol 148:261–270
Cunning R, Baker AC (2012) Excess algal symbionts increase the susceptibility of reef corals to bleaching. Nat Clim Chang 3:259–262
Cziesielski MJ, Liew YJ, Cui G, Schmidt-Roach S, Campana S, Marondedze C, Aranda M (2018) Multi-omics analysis of thermal stress response in a zooxanthellate cnidarian reveals the importance of associating with thermotolerant symbionts. Proc Biol Sci 285
Damjanovic K, Blackall LL, Webster NS, van Oppen MJH (2017) The contribution of microbial biotechnology to mitigating coral reef degradation. Microb Biotechnol 10:1236–1243
Dang KV, Pierangelini M, Roberty S, Cardol P (2019a) Alternative Photosynthetic Electron Transfers and Bleaching Phenotypes Upon Acute Heat Stress in Symbiodinium and Breviolum spp. (Symbiodiniaceae) in Culture. Frontiers in Marine Science 6
Dang KV, Pierangelini M, Roberty S, Cardol P (2019b) Alternative Photosynthetic Electron Transfers and Bleaching Phenotypes Upon Acute Heat Stress in Symbiodinium and Breviolum spp. (Symbiodiniaceae) in Culture. Frontiers in Marine Science 6
Díaz-Almeyda E, Thomé P, El Hafidi M, Iglesias-Prieto R (2011) Differential stability of photosynthetic membranes and fatty acid composition at elevated temperature in Symbiodinium. Coral Reefs 30:217–225
Diaz JM, Hansel CM, Apprill A, Brighi C, Zhang T, Weber L, McNally S, Xun L (2016) Species-specific control of external superoxide levels by the coral holobiont during a natural bleaching event. Nat Commun 7:13801
Downs CA, Fauth JE, Halas JC, Dustan P, Bemiss J, Woodley CM (2002) Oxidative stress and seasonal coral bleaching. Free Radic Biol Med 33:533–543
Downs CA, McDougall KE, Woodley CM, Fauth JE, Richmond RH, Kushmaro A, Gibb SW, Loya Y, Ostrander GK, Kramarsky-Winter E (2013) Heat-stress and light-stress induce different cellular pathologies in the symbiotic dinoflagellate during coral bleaching. PLoS ONE 8:e77173
Dungan AM, Bulach D, Lin H, van Oppen MJH, Blackall LL (2021) Development of a free radical scavenging bacterial consortium to mitigate oxidative stress in cnidarians. Microb Biotech 14(5):2025–2040
Dungan AM, Hartman LM, Blackall LL, van Oppen MJH (2022) Exploring microbiome engineering as a strategy for improved thermal tolerance in Exaiptasia diaphana. J Appl Microbiol 132(4):2940–2956. https://doi.org/10.1111/jam.15465.
Dungan AM, Hartman LM, Tortorelli G, Belderock R, Lamb AM, Pisan L, McFadden G, Blackall LL, van Oppen MJH (2020) Exaiptasia diaphana from the Great Barrier Reef: a valuable resource for coral symbiosis research. Symbiosis 80:195–206
Edmunds PJ (2004) Effect of elevated temperature on aerobic respiration of coral recruits. Mar Biol 146:655–663
Farooq M, Rehman A, Wahid A, Siddique K (2016) Photosynthesis under heat stress Handbook of Photosynthesis. CRC Press, pp697–701
Gegner HM, Radecker N, Ochsenkuhn M, Barreto MM, Ziegler M, Reichert J, Schubert P, Wilke T, Voolstra CR (2019) High levels of floridoside at high salinity link osmoadaptation with bleaching susceptibility in the cnidarian-algal endosymbiosis. Biol Open 8
Goyen S, Pernice M, Szabó M, Warner ME, Ralph PJ, Suggett DJ (2017) A molecular physiology basis for functional diversity of hydrogen peroxide production amongst Symbiodinium spp.(Dinophyceae). Mar Biol 164:46
Halliwell B, Gutteridge J (1990) Role of free radicals and catalytic metal ions in human disease: an overview Methods in enzymology. Elsevier, pp1–85
Hawkins TD, Warner ME (2017) Warm preconditioning protects against acute heat-induced respiratory dysfunction and delays bleaching in a symbiotic sea anemone. J Exp Biol 220:969–983
Hill R, Ulstrup KE, Ralph PJ (2009) Temperature induced changes in thylakoid membrane thermostability of cultured, freshly isolated, and expelled zooxanthellae from scleractinian corals. Bull Mar Sci 85:223–244
Hoadley KD, Pettay DT, Dodge D, Warner ME (2016) Contrasting physiological plasticity in response to environmental stress within different cnidarians and their respective symbionts. Coral Reefs 35:529–542
Hoegh-Guldberg O, Jacob D, Taylor M, Bolaños TG, Bindi M, Brown S, Camilloni IA, Diedhiou A, Djalante R, Ebi KL (2019) The human imperative of stabilizing global climate change at 1.5° C. Science 365:eaaw6974
Hughes TP, Pratchett MS (2020) We just spent two weeks surveying the Great Barrier Reef. What we saw was an utter tragedy The Conversation
Hughes TP, Anderson KD, Connolly SR, Heron SF, Kerry JT, Lough JM, Baird AH, Baum SF, Berumen ML, Bridge T, Claar DC, Eakin CM, Gilmour JP, Graham NAJ, Harrison HB, Hobbs JA, Hoey AS, Hoogenboom M, Lowe RJ, McCulloch MT, Pandolfi JM, Pratchett MS, Schoepf V, Torda G, Wilson SK (2018) Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 359:80–83
Hughes TP, Kerry JT, Alvarez-Noriega M, Alvarez-Romero JG, Anderson KD, Baird AH, Babcock RC, Beger M, Bellwood DR, Berkelmans R, Bridge TC, Butler IR, Byrne M, Cantin NE, Comeau S, Connolly SR, Cumming GS, Dalton SJ, Diaz-Pulido G, Eakin CM, Figueira WF, Gilmour JP, Harrison HB, Heron SF, Hoey AS, Hobbs JA, Hoogenboom MO, Kennedy EV, Kuo CY, Lough JM, Lowe RJ, Liu G, McCulloch MT, Malcolm HA, McWilliam MJ, Pandolfi JM, Pears RJ, Pratchett MS, Schoepf V, Simpson T, Skirving WJ, Sommer B, Torda G, Wachenfeld DR, Willis BL, Wilson SK (2017) Global warming and recurrent mass bleaching of corals. Nature 543:373–377
Iglesias-Prieto R, Matta JL, Robins WA, Trench RK (1992) Photosynthetic response to elevated temperature in the symbiotic dinoflagellate Symbiodinium microadriaticum in culture. Proc Natl Acad Sci U S A 89:10302–10305
Kawamura K, Nishitsuji K, Shoguchi E, Fujiwara S, Satoh N (2021) Establishing Sustainable Cell Lines of a Coral, Acropora tenuis. Mar Biotechnol 23:373–388
Krieger-Liszkay A (2005) Singlet oxygen production in photosynthesis. J Exp Bot 56:337–346
Krueger T, Hawkins TD, Becker S, Pontasch S, Dove S, Hoegh-Guldberg O, Leggat W, Fisher PL, Davy SK (2015) Differential coral bleaching—Contrasting the activity and response of enzymatic antioxidants in symbiotic partners under thermal stress. Comp Biochem Physiol A Mol Integr Physiol 190:15–25
Lesser MP (1996) Elevated temperatures and ultraviolet radiation cause oxidative stress and inhibit photosynthesis in symbiotic dinoflagellates. Limnol Oceanogr 41:271–283
Lesser MP (1997) Oxidative stress causes coral bleaching during exposure to elevated temperatures. Coral Reefs 16:187–192
Lesser MP (2019) Phylogenetic signature of light and thermal stress for the endosymbiotic dinoflagellates of corals (Family Symbiodiniaceae). Limnol Oceanogr 64:1852–1863
Levin RA, Beltran VH, Hill R, Kjelleberg S, McDougald D, Steinberg PD, van Oppen MJH (2016) Sex, Scavengers, and Chaperones: Transcriptome Secrets of Divergent Symbiodinium Thermal Tolerances. Mol Biol Evol 33:2201–2215
Louis YD, Bhagooli R, Kenkel CD, Baker AC, Dyall SD (2017) Gene expression biomarkers of heat stress in scleractinian corals: Promises and limitations. Comp Biochem Physiol C Toxicol Pharmacol 191:63–77
Maire J, van Oppen MJH (2021) A role for bacterial experimental evolution in coral bleaching mitigation? Trends in Microbiology
Majerová E, Drury C (2021) A BI-1 mediated cascade improves redox homeostasis during thermal stress and prevents oxidative damage in a preconditioned reef-building coral. bioRxiv
Majerová E, Carey F, Gates RD (2020) Preconditioning improves bleaching susceptibility in the reef-building coral Pocillopora acuta through modulations in autophagy pathway. Authorea
Marty-Rivera M, Roberson LM, Yudowski GA (2018) Mitigation of coral bleaching by antioxidants. bioRxiv:281014
Mathur S, Agrawal D, Jajoo A (2014) Photosynthesis: Response to high temperature stress. J Photochem Photobiol B 137:116–126
Mattila H, Khorobrykh S, Havurinne V, Tyystjärvi E (2015) Reactive oxygen species: Reactions and detection from photosynthetic tissues. J Photochem Photobiol, B 152:176–214
McGinty ES, Pieczonka J, Mydlarz LD (2012) Variations in reactive oxygen release and antioxidant activity in multiple Symbiodinium types in response to elevated temperature. Microb Ecol 64:1000–1007
Motone K, Takagi T, Aburaya S, Miura N, Aoki W, Ueda M (2020) A Zeaxanthin-Producing Bacterium Isolated from the Algal Phycosphere Protects Coral Endosymbionts from Environmental Stress. Mbio 11:e01019-01019
Mubarakshina MM, Ivanov BN, Naydov IA, Hillier W, Badger MR, Krieger-Liszkay A (2010) Production and diffusion of chloroplastic H2O2 and its implication to signalling. J Exp Bot 61:3577–3587
Murphy MP (2009) How mitochondria produce reactive oxygen species. Biochem J 417:1–13
Muscatine L, Porter JW (1977) Reef corals: mutualistic symbioses adapted to nutrient-poor environments. Bioscience 27:454–460
Nesa B, Hidaka M (2008) Thermal stress increases oxidative DNA damage in coral cell aggregates. 11th International Coral Reef Symposium
Nesa B, Hidaka M (2009) High zooxanthella density shortens the survival time of coral cell aggregates under thermal stress. J Exp Mar Biol Ecol 368:81–87
Nielsen DA, Petrou K, Gates RD (2018) Coral bleaching from a single cell perspective. ISME J 12:1558–1567
Nishiyama Y, Allakhverdiev SI, Murata N (2006) A new paradigm for the action of reactive oxygen species in the photoinhibition of photosystem II. Biochim Biophys Acta Bioenerg 1757:742–749
Oakley CA, Davy SK (2018) Cell Biology of Coral Bleaching. In: van Oppen MJH, Lough JM (eds) Coral Bleaching: Patterns, Processes, Causes and Consequences. Springer International Publishing, Cham, pp 189–211
Oakley CA, Durand E, Wilkinson SP, Peng L, Weis VM, Grossman AR, Davy SK (2017) Thermal shock induces host proteostasis disruption and endoplasmic reticulum stress in the model symbiotic cnidarian Aiptasia. J Proteome Res 16:2121–2134
Peixoto RS, Rosado PM, Leite DCA, Rosado AS, Bourne DG (2017) Beneficial microorganisms for corals (BMC): proposed mechanisms for coral health and resilience. Front Microbiol 8
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Core Team (2019) nlme: linear and nonlinear mixed effects models R Package
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Radecker N, Pogoreutz C, Gegner HM, Cardenas A, Roth F, Bougoure J, Guagliardo P, Wild C, Pernice M, Raina JB, Meibom A, Voolstra CR (2021) Heat stress destabilizes symbiotic nutrient cycling in corals. Proc Natl Acad Sci U S A 118
Ragni M, Airs RL, Hennige SJ, Suggett DJ, Warner ME, Geider RJ (2010) PSII photoinhibition and photorepair in Symbiodinium (Pyrrhophyta) differs between thermally tolerant and sensitive phylotypes. Mar Ecol Prog Ser 406:57–70
Ralph PJ, Hill R, Doblin MA, Davy SK (2016) Theory and application of pulse amplitude modulated chlorophyll fluorometry in coral health assessment. In: Woodley CM, Downs CA, Bruckner AW, Porter JW, Galloway SB (eds) Diseases of coral. Wiley, Hoboken, pp 506–523
Rehman AU, Szabó M, Deák Z, Sass L, Larkum A, Ralph P, Vass I (2016) Symbiodinium sp. cells produce light-induced intra-and extracellular singlet oxygen, which mediates photodamage of the photosynthetic apparatus and has the potential to interact with the animal host in coral symbiosis. New Phytol 212:472–484
Saragosti E, Tchernov D, Katsir A, Shaked Y (2010) Extracellular production and degradation of superoxide in the coral Stylophora pistillata and cultured Symbiodinium. PLoS ONE 5:e12508
Searle SR, Speed FM, Milliken GA (1980) Population marginal means in the linear model: an alternative to least squares means. Am Stat 34:216–221
Snyder GA, Eliachar S, Connelly MT, Talice S, Hadad U, Gershoni-Yahalom O, Browne WE, Palmer CV, Rosental B, Traylor-Knowles N (2021) Functional Characterization of Hexacorallia Phagocytic Cells. Front Immunol 12:662803
Stuart-Smith RD, Brown CJ, Ceccarelli DM, Edgar GJ (2018) Ecosystem restructuring along the Great Barrier Reef following mass coral bleaching. Nature 560:92–96
Suggett DJ, Warner ME, Smith DJ, Davey P, Hennige S, Baker NR (2008) Photosynthesis and production of hydrogen peroxide by Symbiodinium (pyrrhophyta) phylotypes with different thermal tolerances. J Phycol 44:948–956
Szabó M, Larkum AWD, Vass I (2020) A Review: The Role of Reactive Oxygen Species in Mass Coral Bleaching. In: Larkum AWD, Grossmann AR, Raven JA (eds) Photosynthesis in Algae: Biochemical and Physiological Mechanisms. Springer International Publishing, Cham, pp 459–488
Takahashi S, Whitney SM, Badger MR (2009) Different thermal sensitivity of the repair of photodamaged photosynthetic machinery in cultured Symbiodinium species. Proc Natl Acad Sci 106:3237–3242
Takahashi S, Nakamura T, Sakamizu M, van Woesik R, Yamasaki H (2004) Repair machinery of symbiotic photosynthesis as the primary target of heat stress for reef-building corals. Plant Cell Physiol 45:251–255
Takahashi S, Whitney S, Itoh S, Maruyama T, Badger M (2008) Heat stress causes inhibition of the de novo synthesis of antenna proteins and photobleaching in cultured Symbiodinium. Proc Natl Acad Sci U S A 105:4203–4208
Tchernov D, Gorbunov MY, De Vargas C, Yadav SN, Milligan AJ, Häggblom M, Falkowski PG (2004) Membrane lipids of symbiotic algae are diagnostic of sensitivity to thermal bleaching in corals. Proc Natl Acad Sci U S A 101:13531–13535
Tolleter D, Seneca FO, DeNofrio JC, Krediet CJ, Palumbi SR, Pringle JR, Grossman AR (2013) Coral Bleaching Independent of Photosynthetic Activity. Curr Biol 23:1782–1786
Tremblay P, Grover R, Maguer J-F, Hoogenboom M, Ferrier-Pagès C (2014) Carbon translocation from symbiont to host depends on irradiance and food availability in the tropical coral Stylophora pistillata. Coral Reefs 33:1–13
Trubitsin BV, Mamedov MD, Semenov AY, Tikhonov AN (2014) Interaction of ascorbate with photosystem I. Photosynth Res 122:215–231
Tukey JW (1949) Comparing individual means in the analysis of variance. Biometrics:99–114
van Oppen MJH, Oliver JK, Putnam HM, Gates RD (2015) Building coral reef resilience through assisted evolution. Proc Natl Acad Sci USA 112:2307–2313
Villanueva C, Kross RD (2012) Antioxidant-induced stress. Int J Mol Sci 13:2091–2109
Voolstra CR, Buitrago‐López C, Perna G, Cárdenas A, Hume BCC, Rädecker N, Barshis DJ (2020) Standardized short‐term acute heat stress assays resolve historical differences in coral thermotolerance across microhabitat reef sites. Glob Chang Biol 26
Wang J-T, Meng P-J, Sampayo E, Tang S-L, Chen CA (2011) Photosystem II breakdown induced by reactive oxygen species in freshly-isolated Symbiodinium from Montipora (Scleractinia; Acroporidae). Mar Ecol Prog Ser 422:51–62
Warner ME, Suggett DJ (2016) The photobiology of Symbiodinium spp.: linking physiological diversity to the implications of stress and resilience The Cnidaria, past, present and future. Springer, pp489–509
Weis WM (2008) Cellular mechanisms of cnidarian bleaching: stress causes the collapse of symbiosis. J Exp Biol 211:3059–3066
Wickham H (2020) ggplot2: Elegant Graphics for Data Analysis version 3.3. 1. Springer-Verlag, New York
Wiedenmann J, D’Angelo C, Smith EG, Hunt AN, Legiret F, Postle AD, Achterberg EP (2012) Nutrient enrichment can increase the susceptibility of reef corals to bleaching. Nat Clim Chang 3:160
Wietheger A, Starzak DE, Gould KS, Davy SK (2018) Differential ROS generation in response to stress in Symbiodinium spp. Biol Bull 234:11+
Wooldridge SA (2013) Breakdown of the coral-algae symbiosis: towards formalising a linkage between warm-water bleaching thresholds and the growth rate of the intracellular zooxanthellae. Biogeosciences 10:1647–1658
Zhitkovich A (2021) Ascorbate: antioxidant and biochemical activities and their importance for in vitro models. Arch Toxicol
Acknowledgements
The authors thank Leon Hartman, Sarah Jane Tsang Min Ching, and Xavier Smith for their assistance in anemone maintenance, sample collection, and processing. We thank the BioSciences Microscopy Unit (The University of Melbourne) and the Biological Optical Microscopy Platform for the use of their CLSM and Dr. Gabriela Segal for training. Leon Hartman reviewed the manuscript and designed Figure 1. This work was supported by the Australian Research Council [grant numbers DP160101468 and FL180100036].
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Contributions
AMD, LLB, and MJHvO contributed to conceptualization; AMD, JM, and AP-G helped in methodology; AMD performed formal Analysis; AMD and JM contributed to investigation and writing—original draft; AP-G provided resources; AMD, JM, AP-G. LLB, and MJHvO were involved in writing—review and editing; LLB and MJHvO performed funding acquisition and supervision.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Topic Editor Simon Davy
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dungan, A.M., Maire, J., Perez-Gonzalez, A. et al. Lack of evidence for the oxidative stress theory of bleaching in the sea anemone, Exaiptasia diaphana, under elevated temperature. Coral Reefs 41, 1161–1172 (2022). https://doi.org/10.1007/s00338-022-02251-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-022-02251-w