
Abstract
Lentils represent the most common pulse crop found in Southwest Asian Neolithic archaeobotanical assemblages. Using carbon isotope determinations and palaeoclimatic modelling this paper investigates variability and change through time in lentil growth habitats at Gusir Höyük, a Pre-Pottery Neolithic habitation site in the Upper Tigris basin of southeast Anatolia dated to the 12th and 11th millennia cal bp. The elevated lentil carbon isotope values from samples dated to ~ 10,500–10,300 cal bp possibly points to greater water availability and the potential onset of lentil cultivation at Gusir Höyük during the Early Pre-Pottery Neolithic B. The paper revisits previous hypotheses about the process and timing of initial lentil domestication in the region through a re-evaluation of materials available from other Neolithic sites in SW Asia. Its findings highlight the important role of wild lentil populations growing in higher-rainfall localities, which may have responded more readily to increasing water availability in the context of early cultivation. It also explores the potential offered by isotopic approaches for detecting early cultivation, highlighting the need for future studies on pulse and cereal crop progenitors in SW Asia and beyond.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Despite decades of research into the evolution of the domestication syndrome in plants worldwide (reviewed in Fuller et al. 2014) regional processes and pathways remain largely elusive. In Southwest Asia, one of the earliest and best researched centres of agricultural origins in the Old World, most cereal and pulse crop progenitor taxa have unique sets of plant functional traits and broad geographic distributions, which suggests that multiple local populations could have developed specific adaptations to different environmental conditions and varying degrees of plasticity in growth habit, seed size, and yield properties (for a comprehensive review, see Jones et al. 2021). This brings to the fore the need to investigate progenitor taxa individually, taking into account both long-term shifts in environmental conditions and the potential signatures (morphological, chemical, etc.) of their early management and cultivation. Recent work is increasingly demonstrating the diversity and complexity of Pre-Pottery Neolithic (PPN) plant use, early cereal and pulse cultivation, and domestication across SW Asia (Arranz-Otaegui et al. 2016; Caracuta et al. 2017; Kabukcu et al. 2021), also extending to the management of a broad range of other wild seed taxa (Jesus et al. 2021; Whitlam et al. 2018). In addition, recent work has questioned classic archaeobotanical indicators of early cultivation, notably grain-size increase, and increasing yields as ‘intentional’ drivers of cultivation and domestication (Preece et al. 2015; Jones et al. 2021). The value of wild/weedy plant assemblages as indicators of predomestication cultivation (sensu Fuller et al. 2011) has also been comprehensively questioned and re-evaluated (Asouti 2017; Weide et al. 2021).
Genomic research indicates that the most likely progenitor of the domesticated lentil (Lens culinaris Medik. ssp. culinaris) is L. culinaris ssp. orientalis, which has a broad geographic distribution across SW Asia (Alo et al. 2011; Liber et al. 2021). A range of other wild lentil taxa also occur in the region, such as L. nigricans and L. ervoides. All wild relatives of the domesticated lentil are presently found in several regions including southern Anatolia, the northern Euphrates and Tigris basins, and the Levant. Existing evidence on habitat preferences indicates wild lentils are found in semi-arid steppe vegetation, on stony ground and hillslopes as small formations (Zohary 1972; Fuller 2007). Due in part to their plant functional traits (see Warham 2021), wild lentils are not often found in dense stands, or as components of dense grassland/wetland habitats. Their growth habits and tendency to form small clusters in vegetation are often cited as a limiting factor in their intensive harvesting prior to agriculture (Ladizinsky 1993; Abbo et al. 2008). The genomic evidence points to a protracted period of fixation for domestication-related mutations in wild populations distributed across this broad region stretching from southern Anatolia to the Levant (Alo et al. 2011; Liber et al. 2021). However, whether the mutations associated with lentil domestication emerged from a single or a limited number of sub-populations remains unresolved, due to the patchy modern distribution of wild lentils and the high likelihood of introgression between populations (Liber et al. 2021).
While there is consensus in the literature on the progenitors of domesticated lentils and their potential geographic distribution, there are contrasting theories on the development and fixation of specific domestication-related mutations. Based on linear modelling, Ladizinsky (1987) hypothesised that loss of dormancy could arise within a decade in wild populations as a result of intensive year-on-year harvesting prior to the onset of early cultivation. Ladizinsky (1993) also argued that it would not be viable to rely on the cultivation of wild pulses due to their low yields and very low rates of germination, and proposed instead that non-dormant lentils must have already been dominant prior to initial cultivation. A similar argument was put forward by Abbo et al. (2009) backed up by field experiments involving wild pea and lentils. The hypothesis for the fixation of loss of dormancy in lentils prior to the start of their cultivation has been criticized by Zohary (1989), Blumler (1991) and Weiss and Zohary (2011) on the basis that it does not take into account intra-annual variability in seed production and germination. These authors proposed that loss of dormancy is more likely to have developed in response to cultivation favouring the proliferation of non-dormant seeds through repeated planting cycles.
A significant shortcoming of both hypotheses is that they cannot be tested through direct archaeobotanical observation due to the absence of clear markers of phenotypic change in charred plant macrofossils (Weiss and Zohary 2011). Wild lentils, like most legumes, are characterised by high rates of dormancy preventing the free germination of seeds upon planting. Physical dormancy, often referred to as hard-seededness, entails the development of a water impermeable seed coat. This frequently manifests as a thicker seed coat but may also involve plant phenolics, suberin, waxes, or other substances contained in seed coat (testa) layers (Janská et al. 2019). Previous work on the microstructure of pea seeds has shown that wild accessions have considerably thicker seed coats (Butler 1990). Recent work demonstrates that pea seed dormancy is also mediated by the presence of waxes and phenolics in outer testa layers (Smýkal et al. 2014; Janská et al. 2019; Zablatzká et al. 2021). However, it has not been possible to detect similar differences in lentil seed coat thickness: both wild-type and domesticated accessions have relatively thin seed coats (Butler 1990).
Other morphological indicators of domestication remain equally elusive. Pod indehiscence (the loss of free seed dispersal upon ripening) is one secure trait of domesticated lentils (Weiss and Zohary 2011). However, it remains undetected due to the continuing absence of charred lentil pod remains from PPN archaeobotanical assemblages. Lentil seed sizes in modern-day cultivars range between 3 and 9 mm in diameter (ssp. microsperma has smaller pods and seeds of 3–6 mm diameter), while the wild specimens are generally smaller than 3 mm in diameter (Zohary et al. 2012). Archaeological evidence to date indicates that seed size increase was slow in lentils with most Neolithic and earlier specimens having diameters smaller than 3 mm (Fuller 2007). As cultivation and initial domestication, particularly in cereals, is largely established around this time, it has been argued seed size increase in lentils likely represents a much later development that postdates by several centuries the establishment of sustained agricultural practices in SW Asia (Weiss and Zohary 2011; Zohary et al. 2012). It has additionally been argued that the link between domestication and seed-size increase, across a range of cereal and pulse crops, may not have resulted from intentional selective pressures (Preece et al. 2015; Kluyver et al. 2017; Jones et al. 2021), and thus may not be robust indicators of initial cultivation.
An alternative means for assessing early pulse management and cultivation is by examining potential shifts in growth conditions through stable isotopes. In lentils, like most other SW Asian crops and progenitor taxa, carbon isotope values (δ13C ratios) reflect rates of stomatal conductance (Wallace et al. 2013; Fiorentino et al. 2015; Styring et al. 2016). Under conditions of optimal water availability during the growth season, stomatal restriction in CO2 assimilation is minimal: plant tissues assimilate less of the heavier 13C isotope resulting in lower (more negative) δ13C values. Previous research involving crop growth trials and carbon isotope analysis on archaeological cereals and pulses has demonstrated that carbon isotope values (δ13C) likely reflect water availability and uptake during the growth period (Flohr et al. 2011; Wallace et al. 2013). They have thus been used to investigate watering regimes and water availability for cereal and pulse crops in Neolithic and later prehistoric sites in SW Asia and Europe (e.g. Ferrio et al. 2005; Fiorentino et al. 2015; Wallace et al. 2015; Styring et al. 2016). Additionally, studies on oak charcoals have been previously applied in archaeology to provide insights into potential fluctuations in rainfall and water availability in the Iberian Peninsula (Aguilera et al. 2009) and Anatolia (Masi et al. 2013). While tree species are likely to show smaller-scale shifts in carbon isotope values in response to short-term and intra-annual water availability due to their deeper rootstocks, present-day and archaeological work increasingly shows that they can provide reliable indicators of broader climatic shifts, thus offering a good control variable in assessing the impacts of large-scale variations in water availability (e.g. Caracuta et al. 2021). Work done on modern almond wood by Caracuta et al. (2021) demonstrated robust correlation between Δ13C values of almond wood and 10-year averages of annual rainfall specific to collection locations. At collection locations ranging from ca. 550 to ca. 750 mm annual rainfall, the Δ13C on almond wood ranged from 19.4 (± 1.3) to 21.3 (± 1.3) (see Fig. 8; Table 2 in Caracuta et al. 2021). Research to date involving crop growth trials with lentils indicates that seed Δ13C values are likely to reflect plant water status during the grain-filling period, corresponding to April-May (Wallace et al. 2013). Almond nutshell formation also occurs during a similar window, between March-May (Martinez-Gomez et al. 2006), with kernel formation and maturation completing later in the year. Due to the demonstrated potential of Δ13C on archaeological almonds to reflect shifts in water availability, and the occurrence of wild almonds in similar habitats (semi-arid, steppe-like, Asouti and Kabukcu 2014) with wild lentils, carbon isotope determinations on almond nutshell would thus offer a robust control in assessing broad shifts in climatic variables contributing to lentil water status.
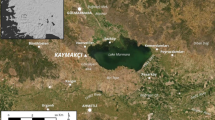
The present paper applies a stable isotope approach to investigating the start of lentil cultivation in PPN SW Asia, using specimens from the site of Gusir Höyük in southeast Anatolia and comparing them to materials available from other Neolithic sites in SW Asia (see Fig. 1 for locations of all sites mentioned in the text). Gusir Höyük (Kabukcu et al. 2021) is one of a handful of excavated and sampled habitation sites in southeast Anatolia that preserve deposits dated from the PPNA through to the Early PPNB horizons (12th and 11th millennia cal bp) a timespan critical for understanding the pace, dynamics and diverse pathways of early cultivation across SW Asia (Asouti and Fuller 2013; Asouti 2017; Kabukcu et al. 2021). The Gusir archaeobotanical assemblage (reported in Kabukcu et al. 2021) is characterised by the predominance of lentils alongside nuts (Pistacia, Amygdalus) throughout the sampled occupation phases. The archaeobotanical and preliminary zooarchaeological results point to a prominent reliance on semi-arid, steppe ecotones (Kabukcu et al. 2021). Sporadic use of wetland ecotones, indicated by the use of riparian woodland vegetation and rushes, as well as occasional hunted fauna found in such ecotones point to the presence of drier and wetter landscape mosaics in the vicinity of the site. As previously reported (Kabukcu et al. 2021), lentil seed diameter measurements indicate the presence of wild-type Lens spp. (all seeds below 3 mm, within a range of 2.79–1.7 mm and average 2.37 mm) in all sampled contexts.The EPPNB marks a critical shift in plant subsistence at Gusir with the appearance of securely dated (~ 10,500–10,300 cal bp) wild-type einkorn and emmer wheat (both absent from PPNA samples) alongside lentils, other pulses and nuts. Wheat seed metrics and chaff morphology indicate that einkorn wheat represents a local population possibly brought into cultivation during the EPPNB, also suggested by the co-occurrence of taxa associated with increasing anthropogenic impacts such as Medicago radiata. Taken together this evidence provides a germane context for exploring whether changes in the carbon isotope values of the Gusir lentils may reflect shifts in soil water availability linked to changing growth conditions (e.g. rainfall, temperature, soil water retention capacity) including anthropogenic impacts (cultivation, water input) from the PPNA to the EPPNB. The isotopic investigation of the Gusir lentil assemblage thus provides a unique opportunity to explore the potential offered by isotopic approaches for elucidating the history of early pulse management and cultivation, to date hindered by the absence of direct archaeobotanical evidence for changes in seed size, testa thickness and/or structure, and dispersal mechanisms.

Map of sites mentioned in the text. Triangles represent sites with Δ13C values reported in the manuscript. Squares represent sites with significant archaeobotanical concentrations of charred lentils considered to represent early lentil cultivation in SW Asia (map prepared using QGIS 3.2, with free vector and raster map data from Natural Earth)
Materials and methods
Carbon isotope measurements were obtained from well-preserved complete lentil seeds (n = 15) retrieved from all sampled PPNA and EPPNB occupation phases at Gusir (Kabukcu et al. 2021). In order to evaluate potential shifts in water availability in the site environs carbon isotope measurements were also obtained from almond (Amygdalus) nutshell (n = 10). Selection of single lentil and nutshell specimens followed previously published research which highlight the importance of the selection of well-preserved specimens to minimise variability due to the loss of seed elements (e.g. testa) (e.g. Fraser et al. 2013; Styring et al. 2013; Wallace et al. 2013). An additional limitation on specimen selection was the lack of closely-controlled charred seed concentrations and/or multiple specimens deriving from stratigraphically directly linked contexts at the site. As a result, it was not justifiable to combine individual specimens from different samples (and different trenches of excavation) to obtain bulk seed samples (sensu Nitsch et al. 2015) for use in carbon isotopic determinations.
The use of individual seeds for carbon isotopic analysis is likely to additionally represent some variability due to naturally occurring within-stand (and amongst the seeds of an individual plant) variability (for further details see Fiorentino et al. 2015). As previously noted, while carbon isotope values in the seeds of modern cultivars can be narrow (e.g. in cereal grains with a standard deviation of ~ 1.3% within a field, Riehl et al. 2014) it has been noted that intrasample variability in archaeological cereal grains could be potentially higher owing to the nature of deposition and charred seed accumulation (Fiorentino et al. 2015). As such, the approach taken in this article, depending on the carbon isotopic values of single specimens, carries with it some interpretative caveats stemming from an expectation of inherently greater variability in measurements.
Lentil seeds from Gusir were examined under a stereo-zoom microscope to assess charring impacts. Specimens exhibiting distortion of surface features were excluded from further analysis. The lentil specimens analysed and reported here represent the only well-preserved, complete specimens (with visibly preserved testa) found at the site. Similarly, criteria for selecting almond nutshell relied on the well-preserved surface of the fragments, avoiding specimens which were split transversally. Additional confirmation of relatively low charring temperatures was achieved by assessing % C preservation indicated by the results of isotopic measurements (50–60% in the specimens reported in this study; see Supplementary Material (ESM Table S1). Due to the low numbers of suitable specimens found in the previously analysed samples (reported in Kabukcu et al. 2021), additional specimens were obtained through sample scanning targeting lentil and almond nutshell remains. An additional limitation was the low numbers of archaeobotanical samples (and excavated deposits) dating to the EPPNB at Gusir Höyük, as a result, the number of analysed lentil specimens from the later period of habitation at the site are limited.
Sample preparation of single seed and nutshell specimens was carried out following experimentally and archaeologically replicated charred plant macro-remain preparation protocols aimed at removing contaminants and homogenising the prepared samples (Vaiglova et al. 2014). Following a gentle acid-only treatment, each specimen was treated with 0.5 M HCl (aqueous solution) at 80 °C for up to 30 min, followed by a minimum of three rinses in Milli-U water. All specimens were freeze-dried and homogenised in an agate pestle and mortar. Carbon isotope ratios were determined using a Thermo Scientific DELTA V Advantage mass spectrometer coupled to a Costech Instruments Elemental Analyser at the Liverpool Isotope Facility for Environmental Research (LIFER) (Department of Earth, Ocean and Ecological Sciences, University of Liverpool). Stable carbon isotopic compositions were calibrated to the VPDB (Vienna PeeDee Belemnite) scale using international calibration standards USGS40 and USGS41 interspersed among archaeological samples in each session. Analytical accuracy (monitoring of systematic errors in measurement) was maintained (< 0.1) and monitored by the LIFER facility via the inclusion of international calibration standards interspersed in each run. Additional monitoring of precision was conducted using an in-house carbonised plant standard prepared from a single archaeological deciduous oak charcoal specimen of at least three replicate aliquots included in each run. Across sessions included in this study, a total of 20 replicate measurements from the plant reference specimen resulted in an average δ13C value of − 24.63 and standard deviation of 0.0735. Single specimen samples were run in duplicates to monitor measurement precision and consistency. Individual carbon isotope measurements, and average and standard deviation values for each specimen, are listed in ESM Table S1.
Following previously published research on carbon isotope ratios used to infer water availability (Ferrio et al. 2005; Fiorentino et al. 2015; Styring et al. 2016) δ13C ratios were converted to account for variations through time in the δ13C value of atmospheric carbon dioxide (δ13Cair) using the equation of Farquhar et al. (1989):
δ13Cair values for each occupation phase at Gusir Höyük (early-late PPNA, EPPNB) and for the lentil δ13C ratios previously reported in the literature (see below) were extracted from the smoothed curve of δ13C of atmospheric CO2 sourced from the CU-INSTAAR/NOAA-CMDL database published by Ferrio et al. (2005).
The Gusir Höyük lentil carbon isotope values were compared to lentil carbon isotope values (from wild-type, genus-level and domesticated lentils) published from SW Asian sites dated to the early Holocene. These include values from ‘Ain Ghazal (Late PPNB; southern Levant) (Wallace et al. 2013), Tell Halula (Middle-LPPNB, PN; northern Levant) (Araus et al. 1999) and Çatalhöyük East (PN; central Anatolia) (Wallace et al. 2013; Vaiglova 2016). ORAU (Oxford Radiocarbon Accelerator Unit) δ13C values reported from radiometrically dated lentil specimens from M’lefaat and Qermez Dere (PPNA; NW Iraq) were also included. The inclusion of ORAU measurements was deemed suitable with the reported uncertainty threshold ~ 0.5–1‰, (VPDB calibrated, see Hedges et al. 1996). While the precision is relatively low, these provide values that can be a robust comparison with lentil specimens reported here and elsewhere in the region. All previously published δ13C values used in this study, their data sources, calibrated age ranges and the variables used in Δ13C calculations are listed in ESM Table S2.
Evaluation of variation in lentil Δ13C values across all sites, reflecting different micro-climatic conditions and time periods, was achieved by extracting four bioclimatic variables using the R (version R 4.2.2, R Core Team 2022), package pastclim 1.2 (Leonardi et al. 2023) which provides access to hind-casted climatic reconstructions based on the HadCM3 model for the last 120k years (Beyer et al. 2020). Extracted bioclimatic variables include annual mean temperature (Bio1), total annual precipitation (Bio12), seasonality of rainfall (Bio15) and seasonality of temperature (Bio4). Bioclimatic variables have been calculated following previously reported bioclimatic indices (Hijmans et al. 2005; O’Donnell and Ignizio 2012) and are used in this paper to assess the potential range of environmental contributors to plant water status at each locality. Extracted bioclimatic variables contained in pastclim 1.2 are bias-corrected and downscaled to 0.5° × 0.5°, thus offering palaeoclimatic reconstructions specific to site localities. For the time periods corresponding to the sites included in this study, bioclimatic reconstructions are available in 1,000-yr time slices.
Results
The Gusir Höyük almond nutshell and lentil seed Δ13C values are presented in Fig. 2a and b. Virtually no change is evidenced in the water levels indicated by almond nutshell isotope determinations through time; Δ13C values remain relatively high throughout the PPNA and EPPNB (ranges: 18.2 to 20.8, average values 19.41 ± 0.73 across all phases; see ESM Table S1 and Table 1) and are higher than the Δ13C almond nutshell values previously reported from Çatalhöyük East (ranges: 17.2 to 18.3; Vaiglova 2016, p. 102). By contrast, the lentil Δ13C values from both PPNA early (average values 17.10 ± 1.52) and later (average values 17.64 ± 0.96) phases display broader variability (see Table 1), including some specimens with much higher values, as well as those with lower values for each of the phases examined. Slightly elevated values in lentil Δ13C are seen during the EPPNB, with average values of 19.32 ± 0.94, again with relatively broad variability (Table 1; Fig. 2b, see also ESM Table S1). Across the analysed assemblage from Gusir Höyük including the PPNA and EPPNB layers, carbon isotope values of lentils are generally higher than those reported as well-watered lentils, from crop growth trials reported in Wallace et al. (2013), with Δ13C values above 17. At the same time, a small number of specimens from the earliest PPNA phase at the site have values around the poorly-watered lentil threshold (~ 15.5) as reported by Wallace et al. (2013) (see ESM Table S1 and Fig. 2b).

a Plot of Δ13C values of almond nutshell (Amyg.) from PPNA and EPPNB samples at Gusir Höyük (n number of specimens; for data sources see ESM Table S1). b Plot of Δ13C values of lentils (Lens) from PPNA and EPPNB samples at Gusir Höyük (n number of specimens; for data sources see ESM Table S1)
A comparison of the Gusir lentil Δ13C values with those available from other PPN and PN sites further underscores the uniquely well-watered status of the Gusir EPPNB lentils (Fig. 3). The Δ13C values from Qermez Dere and M’lefaat (PPNA; NW Iraq) are largely in line with those of the Gusir PPNA lentils. At the same time, the Gusir EPPNB lentil Δ13C values are much higher than those of later Neolithic sites with well-established crop production regimes such as ‘Ain Ghazal (LPPNB), Tell Halula (M-LPPNB, PN) and Çatalhöyük East (PN) (Fig. 3).

Plot of Δ13C values of lentils from Qermez Dere, M’lefaat, Gusir Höyük, ‘Ain Ghazal, Tell Halula and Çatalhöyük East (n number of reported measurements; for data sources see ESM Table S2)
The possibility that the observed variation in lentil Δ13C values could represent local variability in climatic conditions was further explored through an evaluation of the extracted bioclimatic variables (see previous section) for all sites from which carbon isotope data were available. The details of all site locations and the time slices used for extracting bioclimatic variables from pastclim 1.2 are listed in Tables 2 and 3. Total annual precipitation reconstructions for the relevant time slices for sites with reported lentil Δ13C values (Table 2) indicate that, amongst these sites, Gusir likely received the highest amount of annual rainfall across its PPNA and EPPNB occupation phases. Seasonality of rainfall is relatively high at most sites except Çatalhöyük where it is considerably lower compared to all other sites. Çatalhöyük also has the lowest annual mean temperature (11 °C). All sites located in the upper Tigris and Euphrates basins (Gusir Höyük, Qermez Dere, M’lefaat and Tell Halula) show higher seasonality of temperature. However, despite variability in key bioclimatic indicators, lentils from most sites display a similar range of Δ13C values irrespective of their wild or cultivar status (Fig. 3). The high values of EPPNB Gusir represent the sole outlier. The low values of the LPPNB ‘Ain Ghazal lentils may indicate drought-related stress, also suggested by the low total annual precipitation modelled for this locality (see Table 2). However, they are still within the range of variability observed at other sites (except EPPNB Gusir) from which lentil carbon isotope data are available (Fig. 3).
Discussion
The Gusir Höyük lentil Δ13C values show generally greater water availability during both the PPNA to the EPPNB, with potentially a slight increase during the later occupation phase (see Table 1). Both the Gusir almond nutshell Δ13C values (Fig. 2a) and the modelled bioclimatic variables (Table 2) do not point to significant shifts in environmental conditions (annual precipitation, mean temperature, seasonality of rainfall/temperature) that could have affected water availability in the site environs during the PPNA to the EPPNB. Previously published work on irrigation and water deficit growth trials in almonds, with measurements of Δ13C on nutshell (Rahimi-Eichi 2014), indicates that carbon isotope values of almonds are likely to reflect inter-annual variations in water input. As reported by Rahimi-Eichi (2014), at a growth trial site with 214 mm annual rainfall, the irrigation treatments ranging from 476 to 933 mm/year resulted in Δ13C on nutshell ranging from 16.01 to 20.02 (Rahimi-Eichi 2014, p. 27). The author did not however report on within stand variability of measurements, and thus it is unclear if the consistency of almond nutshell values at Gusir indicates buffering of fluctuations in water due to the growth habit of almonds (as a shrub, with deeper roots) or if it indicates the greater likelihood of almonds to be located in more semi-arid/drier locations in the landscape.
Importantly, the broad range of values observed in both PPNA lentils at Gusir, and the slightly elevated values during the EPPNB points to the likelihood that lentils, at least those exploited by the inhabitants of Gusir Höyük, potentially had broader distribution into wetter, alluvial and/or wetland edge habitats, where soils could have exhibited greater water retention. One possible scenario under which this could have happened is through the transplanting and cultivation of lentils in better-watered soils by the site inhabitants. It is also possible that these higher values indicate a shift in wild lentil habitats and harvesting locations, which may have incorporated a greater diversity of wetland edge/riparian ecotones. The available archaeobotanical data from the site (Kabukcu et al. 2021) indicate the increasing presence and/or use of wetland ecotones during the EPPNB (with archaeobotanical finds of Cyperaceae seeds and riparian/wet-woodland taxa charcoals during this period). However, the functional traits and habitat preferences of present-day wild lentils strongly suggest that they tend to occupy generally drier ground, avoiding dense herbaceous ecotones (Zohary 1972; Zohary et al. 2012; Warham 2021), and are thus less likely to be commonly found naturally occurring in wetland habitats in high numbers. Notwithstanding caveats with sample sizes and low specimen numbers (particularly during the EPPNB), these potential sources of shifts and variability observed in PPN lentil carbon isotope values can be tested in future work involving isotope determinations on lentil seeds across a broader range of sites, including several PPN habitations where arguments have already been put forward for potential cultivation and/or domestication based largely on the presence of seed storage contexts (e.g. Garfinkel et al. 1988; Caracuta et al. 2017).
The comparison of the Gusir lentil Δ13C values to those available from other Neolithic sites in SW Asia characterised by different climatic regimes reveals clear similarities between the Gusir PPNA values and those from PPNA Qermez Dere and M’lefaat (Fig. 3) despite the lower annual rainfall modelled for these sites (Table 2). Furthermore, a comparison of regional lentil Δ13C data through time, including populations likely gathered from the wild (PPNA Gusir, Qermez Dere, M’lefaat) and cultivated ones from later PPNB and PN sites (‘Ain Ghazal, Tell Halula, Çatalhöyük East), shows similar carbon isotope value ranges, despite inter-site variations in annual rainfall and (morphologically elusive) differences in loss of dormancy between pre-agricultural and fully dependent on crop production sites.
As demonstrated by the limited range of the regional comparative data, carbon isotopic studies of lentils and other pulses from PPN sites in SW Asia are still in short supply. PPN sites other than Gusir with (non-morphological) evidence pointing to early lentil cultivation include Ahihud, and Yiftah’el (southern Levant), Tell Qaramel, Jerf el Ahmar and Dja’de (Syrian northern Euphrates basin) and Çayönü and Nevali Çori (southeast Anatolia). Sizeable concentrations of charred lentils and other pulses recovered from Ahidud (~ 10,200 cal bp) and Yiftah’el (~ 10,000 cal bp) have been interpreted as indicators of sustained cultivation (Garfinkel et al. 1987, 1988; Caracuta et al. 2017). Neither site has produced evidence of morphological markers for the development of domestication traits, although Garfinkel et al. (1988) argued that lentil cultivation at Yiftah’el likely involved already domesticated seed stock that was accompanied by weeds of cultivation (Galium tricornutum). In a similar vein, Willcox et al. (2009) interpreted the increasing ubiquity and abundance of lentils at PPNA sites in the Syrian northern Euphrates as evidence for their cultivation. Lentil cultivation has also been surmised for the PPNB phases of Çayönü and Nevali Çori in southeast Anatolia based on the abundance and density of charred lentil seed remains found at these sites (van Zeist and de Roller 1994, 2003; Pasternak 1998).
An important commonality between these sites and PPNA/EPPNB Gusir Höyük is revealed by a closer inspection of key bioclimatic variables extracted for all localities (see Table 3) demonstrating the prevalence of annual precipitation > 300 mm. Sites with notably dense lentil remains (Yiftah’el, Ahidud, Çayönü, Nevali Çori) are also characterised by annual precipitation > 450 mm. This pattern suggests that across SW Asia the earliest potential localities of lentil cultivation overlap with higher-rainfall micro-ecologies. Interestingly, recent genomic research has indicated that the domesticated lentil gene pool carries strong adaptive drought-tolerance mutations (Liber et al. 2021), which likely reflects later agronomic selection and/or adaptations to drier environments under a sustained farming regime. On the other hand, the combined isotopic, macrobotanical and palaeoclimatic data presented here suggest that early cultivation probably took hold in higher rainfall areas where local lentil progenitor populations exhibited greater plasticity in their responses to increasing soil water availability, as suggested by the Gusir EPPNB lentil Δ13C values.
Conclusions
This paper presents new evidence on lentil carbon isotope values coupled with a detailed evaluation of palaeoclimatic datasets from the site of Gusir Höyük in southeast Anatolia with an occupation sequence extending from the PPNA through to the EPPNB horizons (12th and 11th millennia cal bp) The results of this study show elevated lentil Δ13C values in the PPNA (~ 11,400–10,900 cal bp) and a slight increase in the EPPNB (~ 10,500–10,300 cal bp) occupation horizons at the site potentially indicating greater water availability. At the same time, almond nutshell Δ13C values remain stable from the PPNA to the EPPNB. Keeping in mind the small sample sizes of lentils available for carbon isotope determinations from the EPPNB, these higher values might suggest a shift in lentil growth conditions that could stem from a shift in the lentil habitats from which gathering was done by the site inhabitants, or as a result of lentil cultivation during the mid-11th millennium cal bp. The majority of lentil carbon isotope values from the PPNA and EPPNB phases at the site show a generally well-watered status, and are in line with modern growth trials involving irrigated lentil crops. In reassessing the potential processes of early lentil cultivation across SW Asia, the Gusir lentil Δ13C values were also compared to previously published lentil isotope data from Neolithic sites in the region alongside modelled palaeo-bioclimatic variables for each locality. The investigation highlights the gaps in knowledge regarding the habitat preferences, growth conditions and likely adaptive responses of wild lentils to early cultivation. The results presented draw attention to the prominence of higher-rainfall habitats, such as the environs of Gusir Höyük, as foci of early lentil cultivation hosting progenitor populations that may have been more responsive to intensive management and cultivation compared to drought-adapted populations in lower rainfall ecotones.
References
Abbo S, Zezak I, Schwartz E, Lev-Yadun S, Gopher A (2008) Experimental harvesting of wild peas in Israel: implications for the origins of near east farming. J Archaeol Sci 35:922–929
Abbo S, Saranga Y, Peleg Z, Kerem Z, Lev-Yadun S, Gopher A (2009) Reconsidering domestication of legumes versus cereals in the ancient near east. Q Rev Biol 84:29–50. https://doi.org/10.1086/596462
Aguilera M, Espinar C, Ferrio JP, Pérez G, Voltas J (2009) A map of autumn precipitation for the third millennium BP in the Eastern Iberian Peninsula from charcoal carbon isotopes. J Geochem Explor 102:157–165. https://doi.org/10.1016/j.gexplo.2008.11.019
Alo F, Furman BJ, Akhunov E, Dvorak J, Gepts P (2011) Leveraging genomic resources of model species for the assessment of diversity and phylogeny in wild and domesticated lentil. J Hered 102:315–329. https://doi.org/10.1093/jhered/esr015
Araus JL, Febrero A, Catala M, Molist M, Voltas J, Romagosa I (1999) Crop water availability in early agriculture: evidence from carbon isotope discrimination of seeds from a tenth millennium BP site on the Euphrates. Glob Chang Biol 5:201–212. https://doi.org/10.1046/j.1365-2486.1999.00213.x
Arranz-Otaegui A, Colledge S, Zapata L, Teira-Mayolini LC, Ibáñez JJ (2016) Regional diversity on the timing for the initial appearance of cereal cultivation and domestication in southwest Asia. Proc Natl Acad Sci USA 113:14001–14006. https://doi.org/10.1073/pnas.1612797113
Asouti E (2017) Human palaeoecology in Southwest Asia during the early Pre-Pottery Neolithic (c. 9700 – 8500 cal BC): the plant story. In: Benz M, Gebel HGK, Watkins T (eds) Neolithic corporate identities. Studies in early near eastern production, subsistence, and environment (SENEPSE) 20. Ex oriente, Berlin, pp 21–53
Asouti E, Fuller DQ (2013) A contextual approach to the emergence of agriculture in Southwest Asia: reconstructing early neolithic plant-food production. Curr Anthropol 54:299–345. https://doi.org/10.1086/670679
Asouti E, Kabukcu C (2014) Holocene semi-arid oak woodlands in the Irano-Anatolian region of Southwest Asia: natural or anthropogenic? Q Sci Rev 90:158–182. https://doi.org/10.1016/j.quascirev.2014.03.001
Beyer RM, Krapp M, Manica A (2020) High-resolution terrestrial climate, bioclimate and vegetation for the last 120,000 years. Sci Data 7:236. https://doi.org/10.1038/s41597-020-0552-1
Blumler MA (1991) Modelling the origins of legume domestication and cultivation. Econ Bot 45:243–250. https://doi.org/10.1007/BF02862051
Butler EA (1990) Legumes in antiquity: a micromorphological investigation of seeds of the Vicieae. University of London, London
Caracuta V, Vardi J, Paz Y, Boaretto E (2017) Farming legumes in the pre-pottery neolithic: new discoveries from the site of Ahihud (Israel). PLoS ONE 12:e0177859. https://doi.org/10.1371/journal.pone.0177859
Caracuta V, Alex B, Regev L et al (2021) The Marine Isotope Stage 3 landscape around Manot Cave (Israel) and the food habits of anatomically modern humans: new insights from the anthracological record and stable carbon isotope analysis of wild almond (Amygdalus sp). J Hum Evol 160:102868. https://doi.org/10.1016/j.jhevol.2020.102868
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annu Rev Plant Physiol Plant Mol Biol 40:503–537. https://doi.org/10.1146/annurev.pp.40.060189.002443
Ferrio JP, Araus JL, Buxó R, Voltas J, Bort J (2005) Water management practices and climate in ancient agriculture: inferences from the stable isotope composition of archaeobotanical remains. Veget Hist Archaeobot 14:510–517. https://doi.org/10.1007/s00334-005-0062-2
Fiorentino G, Ferrio JP, Bogaard A, Araus JL, Riehl S (2015) Stable isotopes in archaeobotanical research. Veget Hist Archaeobot 24:215–227. https://doi.org/10.1007/s00334-014-0492-9
Flohr P, Müldner G, Jenkins E (2011) Carbon stable isotope analysis of cereal remains as a way to reconstruct water availability: preliminary results. Water Hist 3:121–144. https://doi.org/10.1007/s12685-011-0036-5
Fraser RA, Bogaard A, Charles M et al (2013) Assessing natural variation and the effects of charring, burial and pre-treatment on the stable carbon and nitrogen isotope values of archaeobotanical cereals and pulses. J Archaeol Sci 40:4754–4766. https://doi.org/10.1016/j.jas.2013.01.032
Fuller DQ (2007) Contrasting patterns in crop domestication and domestication rates: recent archaeobotanical insights from the Old World. Ann Bot 100:903–924. https://doi.org/10.1093/aob/mcm048
Fuller DQ, Willcox G, Allaby R (2011) Cultivation and domestication had multiple origins: arguments against the core area hypothesis for the origins of agriculture in the near east. World Archaeol 43:628–652. https://doi.org/10.1080/00438243.2011.624747
Fuller DQ, Denham T, Arroyo-Kalin M et al (2014) Convergent evolution and parallelism in plant domestication revealed by an expanding archaeological record. Proc Natl Acad Sci USA 111:6147–6152. https://doi.org/10.1073/pnas.1308937110
Garfinkel Y, Carmi I, Vogel JC (1987) Dating of horsebean and lentil seeds from the Pre-Pottery Neolithic B village of Yiftaḥ’el. Isr Explor J 37:40–42. https://www.jstor.org/stable/27926051
Garfinkel Y, Kislev ME, Zohary D (1988) Lentil in the Pre-Pottery Neolithic B Yiftaḥ’el: additional evidence of its early domestication. Isr J Bot 37:49–51
Hedges REM, Pettitt PB, Bronk Ramsey C, van Klinken GJ (1996) Radiocarbon dates from the Oxford AMS system: archaeometry datelist 22. Archaeometry 38:391–415. https://doi.org/10.1111/j.1475-4754.1996.tb00785.x
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25(1):965–1978. https://doi.org/10.1002/joc.1276
Janská A, Pecková E, Sczepaniak B, Smýkal P, Soukup A (2019) The role of the testa during the establishment of physical dormancy in the pea seed. Ann Bot 123:815–829. https://doi.org/10.1093/aob/mcy213
Jesus A, Bonhomme V, Evin A et al (2021) A morphometric approach to track opium poppy domestication. Sci Rep 11:9778. https://doi.org/10.1038/s41598-021-88964-4
Jones G, Kluyver T, Preece C et al (2021) The origins of agriculture: intentions and consequences. J Archaeol Sci 125:105290. https://doi.org/10.1016/j.jas.2020.105290
Kabukcu C, Asouti E, Pöllath N, Peters J, Karul N (2021) Pathways to plant domestication in Southeast Anatolia based on new data from aceramic neolithic Gusir Höyük. Sci Rep 11:2112. https://doi.org/10.1038/s41598-021-81757-9
Kluyver TA, Jones G, Pujol B et al (2017) Unconscious selection drove seed enlargement in vegetable crops. Evol Lett 1:64–72. https://doi.org/10.1002/evl3.6
Ladizinsky G (1987) Pulse domestication before cultivation. Econ Bot 41:60–65. https://doi.org/10.1007/BF02859349
Ladizinsky G (1993) Lentil domestication: on the quality of evidence and arguments. Econ Bot 47:60–64. https://doi.org/10.1007/BF02862206
Leonardi M, Hallet EY, Beyer R, Krapp M, Manica A (2023) pastclim: an R package to easily access and use palaeoclimatic reconstructions. Ecography 2023:e06481. https://doi.org/10.1111/ecog.06481
Liber M, Duarte I, Maia AT, Oliveira HR (2021) The history of lentil (Lens culinaris subsp. culinaris) domestication and spread as revealed by genotyping-by-sequencing of wild and landrace accessions. Front Plant Sci 12:628439. https://doi.org/10.3389/fpls.2021.628439
Martinez-Gomez P, Sanchez-Pérez R, Dicenta F (2006) Fruit development in almond. Acta Hortic 726:241–246. https://doi.org/10.17660/ActaHortic.2006.726.38
Masi A, Sadori L, Baneschi I, Siani AM, Zanchetta G (2013) Stable isotope analysis of archaeological oak charcoal from eastern Anatolia as a marker of mid-holocene climate change. Plant Biol 15(Suppl1):83–92
Nitsch EK, Charles M, Bogaard A (2015) Calculating a statistically robust d13C and d15 N offset for charred cereal and pulse seeds. STAR. https://doi.org/10.1179/2054892315Y.0000000001
O’Donnell MS, Ignizio DA (2012) Bioclimatic predictors for supporting ecological applications in the conterminous United States. U.S. Geological Survey, Reston
Pasternak R (1998) Investigations of botanical remains from Nevali Çori PPNB, Turkey: a short interim report. In: Damania AB, Valkoun J, Willcox G, Qualset CO (eds) Origins of agriculture and crop domestication. Proceedings of the harlan symposium. ICARDA, Aleppo, pp 170–177
Preece C, Livarda A, Wallace M et al (2015) Were fertile crescent crop progenitors higher yielding than other wild species that were never domesticated? New Phytol 207:905–913. https://doi.org/10.1111/nph.13353
R Core Team (2022) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rahimi-Eichi V (2014) Water use efficiency in almonds (Prunus dulcis (Mill.). University of Adelaide, Adelaide
Riehl S, Pustovoytov KE, Weippert H, Klett S, Hole F (2014) Drought stress variability in ancient Near Eastern agricultural systems evidenced by δ13C in barley grain. Proc Natl Acad Sci USA 111:12348–12353. https://doi.org/10.1073/pnas.1409516111
Smýkal P, Vernoud V, Blair MW, Soukup A, Thompson RD (2014) The role of the testa during development and in establishment of dormancy of the legume seed. Front Plant Sci 5:351. https://doi.org/10.3389/fpls.2014.00351
Styring AK, Manning H, Fraser RA et al (2013) The effect of charring and burial on the biochemical composition of cereal grains: investigating the integrity of archaeological plant material. J Archaeol Sci 40:4:767–4779. https://doi.org/10.1016/j.jas.2013.03.024
Styring AK, Ater M, Hmimsa Y et al (2016) Disentangling the effect of farming practice from aridity on crop stable isotope values: a present-day model from Morocco and its application to early farming sites in the eastern Mediterranean. Anthr Rev 3:2–22. https://doi.org/10.1177/2053019616630762
Vaiglova P (2016) Neolithic agricultural management in the Eastern mediterranean: new insight from a multi-isotope approach. University of Oxford, Oxford
Vaiglova P, Snoeck C, Nitsch E, Bogaard A, Lee-Thorp J (2014) Impact of contamination and pre-treatment on stable carbon and nitrogen isotopic composition of charred plant remains. Rapid Commun Mass Spectrom 28(2):497–2510. https://doi.org/10.1002/rcm.7044
Van Zeist W, de Roller GJ (1994) The plant husbandry of aceramic Çayönü. SE Turk Palaeohistoria 33/34(1991/1992):65–96
Van Zeist W, de Roller GJ (2003) The Çayönü archaeobotanical record. In: van Zeist W (ed) Reports on archaeobotanical studies in the Old World. Groningen Institute of Archaeology, Groningen, pp 143–166
Wallace M, Jones G, Charles M, Fraser R, Halstead P, Heaton THE, Bogaard A (2013) Stable carbon isotope analysis as a direct means of inferring crop water status and water management practices. World Archaeol 45:388–409. https://doi.org/10.1080/00438243.2013.821671
Wallace MP, Jones G, Charles M, Fraser R, Heaton THE, Bogaard A (2015) Stable carbon isotope evidence for neolithic and bronze age crop water management in the Eastern Mediterranean and Southwest Asia. PLoS ONE 10:e0127085. https://doi.org/10.1371/journal.pone.0127085
Warham G (2021) Functional attributes as a tool for understanding the process of cereal and pulse domestication. The University of Sheffield, Sheffield
Weide A, Hodgson JG, Leschner H et al (2021) The association of arable weeds with modern wild cereal habitats: implications for reconstructing the origins of plant cultivation in the Levant. Environ Archaeol online. https://doi.org/10.1080/14614103.2021.1882715
Weiss E, Zohary D (2011) The neolithic southwest asian founder crops: their biology and archaeobotany. Curr Anthropol 52(Suppl4):S237–S254. https://doi.org/10.1086/658367
Whitlam J, Bogaard A, Matthews R, Matthews W, Mohammadifar Y, Ilkhani H, Charles M (2018) Pre-agricultural plant management in the uplands of the central Zagros: the archaeobotanical evidence from Sheikh-e Abad. Veget Hist Archaeobot 27:817–831. https://doi.org/10.1007/s00334-018-0675-x
Willcox G, Buxo R, Herveux L (2009) Late pleistocene and early Holocene climate and the beginnings of cultivation in northern Syria. Holocene 19:151–158. https://doi.org/10.1177/0959683608098961
Zablatzká L, Balarynová J, Klčová B, Kopecký P, Smýkal P (2021) Anatomy and histochemistry of seed coat development of wild (Pisum sativum subsp. elatius (M. Bieb.) Asch. et Graebn. and domesticated pea (Pisum sativum subsp. sativum L.). Int J Mol Sci 22:4602. https://doi.org/10.3390/ijms22094602
Zohary D (1972) The wild progenitor and the place of origin of cultivated lentil—Lens culinaris. Econ Bot 26:326–332. https://doi.org/10.1007/BF02860702
Zohary D (1989) Pulse domestication and cereal domestication: how different are they? Econ Bot 43:31–34. https://doi.org/10.1007/BF02859322
Zohary D, Hopf M, Weiss E (2012) Domestication of plants in the Old World: the origin and spread of domesticated plants in Southwest Asia, Europe, and the Mediterranean Basin, 4th edn. Oxford University Press, Oxford
Acknowledgements
Ceren Kabukcu acknowledges fellowship funding from the Leverhulme Trust (Early Career Fellowship, ECF–284), which supported the data collection reported in this manuscript and currently receives fellowship funding from the Fundação para a Ciência e a Tecnologia (FCT, Portugal), awarded through the University of Algarve (CEECInst-2.ª edição; reference: CEECINST/00052/2021/CP2792/CT0006). The author thanks Jessica Pearson for extending expertise and methodological training in stable isotope analysis during CK’s Leverhulme Early Career Fellowship at the Department of Archaeology, Classics and Egyptology (University of Liverpool). The author also thanks Jade Whitlam and Alexander Weide for their invitation to submit this manuscript to the Special Issue, and two anonymous reviewers, Eleni Asouti, and Hugo Rafael Oliveira for providing insightful feedback and suggestions for improvement on the draft manuscript. Thanks are also due to Necmi Karul, director of the Gusir höyük project, for providing access to the archaeobotanical samples reported in this study.
Funding
Open access funding provided by FCT|FCCN (b-on). Ceren Kabukcu acknowledges fellowship funding from the Leverhulme Trust (Early Career Fellowship, ECF–284) and currently receives fellowship funding from the Fundação para a Ciência e a Tecnologia (FCT, Portugal), awarded through the University of Algarve (CEECInst-2.ª edição) Reference: CEECINST/00052/2021/CP2792/CT0006.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Whitlam.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kabukcu, C. Reassessing the origin of lentil cultivation in the Pre-Pottery Neolithic of Southwest Asia: new evidence from carbon isotope analysis at Gusir Höyük. Veget Hist Archaeobot 32, 533–543 (2023). https://doi.org/10.1007/s00334-023-00935-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-023-00935-z