
Abstract
We investigated lake sediments from Großer Ursee, Landkreis Ravensburg, Allgäu, Baden-Württemberg, to explore whether human activity was delayed and less intense at higher sites compared to those in the adjacent lower area around Bodensee (Lake Constance). A 11.1 m high resolution pollen profile from the centre of the lake was analysed, which covers the late Würm and complete Holocene sequence, providing a continuous pollen record of 1,092 samples. Human and climate induced vegetation and landscape change is discussed. The history of human impact indicated in this profile at around 700 m in the western Allgäu, with a precipitation above 1,000 mm/year and with a less favourable climate is compared with human impact in adjacent lower landscapes, with a more favourable climate for agriculture such as the Bodensee region. Since the Neolithic period, differences in vegetation development between the regions were mainly caused by a varying intensity of human impact which itself was strongly influenced by climatic differences. In the lowlands with a warm and dry climate, human impact started earlier and was stronger than in less favourable areas. Finally, the regular occurrence of Trapa natans at Großer Ursee, much more frequent there than in the adjacent warmer landscape to the west, is discussed in terms of various ecological factors.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The success of agriculture depends on soil quality and climatic conditions. The present day climatic range of crop cultivation is broader than in the past, a consequence of progress in agricultural technology and plant breeding (Flint 2015). The climatic range of agriculture in the past is estimated to have been within a mean annual ground surface temperature range of 7–20 °C and a total annual precipitation of 400–1,000 mm. The required precipitation depends on the temperature, the higher the temperature, the higher the required precipitation. Agriculture has its optimal conditions when the precipitation is totally consumed by evaporation and plant transpiration, and there is no runoff, because runoff means loss of plant nutrients (Wright 1977). Focusing on temperate Europe, the landscapes best suited for agriculture are the loess-covered plain in the lowlands with mean annual ground surface temperatures between 7 and 10 °C and a total annual precipitation between 450 and 650 mm. These are the landscapes which were settled since the early Neolithic and with strong human impact in all periods (Preuß 2008). The low mountain ranges of central Europe, the lowlands of northern central Europe, as well as the pre-alpine lowlands have a wetter and cooler climate than the lowlands of the loess belt. The question is, if this last area was settled later and also less intensely for most of the time as the archaeological record suggests (for example, Engelhardt 2006; Nadler 2006), what were the climatic limits for agriculture in the past? For this, high resolution pollen evidence can be more meaningful than archaeological evidence, which is restricted by the state of the art. We focus on a site where there is hardly any archaeological evidence older than the medieval period, where its suitability for agriculture is questionable, and which is not cultivated now. Because no prehistoric lake shore settlements were known in the eastern part of the Rhine glacier region with an annual precipitation above 1,000 mm, it was thought that prehistoric farmers were not able to colonize and cultivate these regions of eastern Oberschwaben and Allgäu, which today have no arable farming, but only animal husbandry.
A high-resolution lake pollen profile from Großer Ursee, Landkreis Ravensburg, situated in this wet climatic region, should, in comparison with profiles from the dryer landscapes, answer this question. The aim was to provide a high resolution standard pollen record of the vegetation history and human impact in the western part of the Allgäu where such data have been scarce up to now, compared to adjacent areas to the east and west.
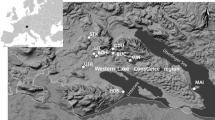
Another aim was to compare this site and its evidence for human impact with the nearby region to the west, which is better suited for agriculture and where archaeological evidence for strong human occupation goes back to the later Neolithic (ca. 4300–3500 bc), the western Bodensee area. To the east part of the Allgäu, in Bayern (Bavaria), close to the Alps is also discussed (Fig. 1).

Map of the study area in southwestern Germany showing location of Großer Ursee and other pollen sites in the Allgäu on the right and pollen records from the western Bodensee (Lake Constance) region on the left (from www.deine-berge.de, modified)
Geographical setting
The Rhine glacier covered an area of about 10,000 km2 outside the Alps at the time of its maximum extension in the Würm period (Keller and Krayss 1993, 2005), which is today represented by the regions Ostschweizer Mittelland in eastern Switzerland and Oberschwaben in southwestern Germany, with Bodensee (Lake Constance) in the centre. The glacier formed its terminal moraines close to the Adelegg hills, which consist of molasse conglomerate, and close to the Iller glacier in the east. This area is also part of the European main watershed between the rivers Rhine and Danube (Scholz 2016) and there is a group of lakes and bogs here, the south-easternmost of which is Großer Ursee (10° 01′32″ E, 47° 45′ 10″ N). It covers 20 ha, with a maximum water depth of 10 m (Fig. 1). Its watershed is at 695 m a.s.l., but in contrast, the sites around Bodensee are at between 395 and 450 m. Seven km to the southeast of Großer Ursee, the Adelegg hills reach an altitude of more than 1,100 m. The temperatures in the Allgäu are not only lower than at Bodensee, but the precipitation is also much higher, as the Allgäuer Alpen and Bregenzer Wald cause increasing precipitation to the east. Because of the unfavourable climatic conditions of the Allgäu, the only form of farming there today is stock grazing. The potential natural vegetation there is considered to be Fagus (beech) woods with Abies (fir) and some Picea (spruce), but the woodlands would mainly consist of Abies in some places (Müller et al. 1974). For soils, eutric cambisols cover most of the Würm glaciated area of the southwestern German pre-alps. Soils with stagnic and gleyic properties as well as histosols occur in depressions (Hornig et al. 1991). There is a precipitation gradient from west to east across Bodensee so that over 100 km, the annual precipitation increases from less than 700 mm in the Hegau (Singen) to more than 1,200 mm in the Allgäu (Meteoblue 2019). This is caused by a reduction in precipitation from the rain shadow of the Schwarzwald (Black Forest) to the west of this area and the windward slope of the Alps increasing it to the east.
Material and methods
A 12 m profile was collected from Ursee, consisting of 2 m segments taken in the centre of the lake, using an UWITEC piston corer. This was analysed continuously from a depth of 1,110 cm to the top at intervals of 1 cm, with a sum of 1,000 arboreal pollen in each sample. The preparation included treatment with hot HCL, HF where necessary, acetolysis and staining in glycerol (Berglund and Ralska-Jasiewiczowa 1986). The resulting data are shown in a pollen diagram on the basis of 1,092 samples, established using Tilia and Tiliagraph (https://www.tiliait.com/). After establishing an outline diagram with a sampling interval of 10 cm (Rösch and Hahn 2016), the core was shared between the two first authors of this paper, PS analysing the late Würm and early Holocene part, and MR the upper part from the first occurrences of Picea, Abies and Fagus upwards. The chronology is based on 17 radiocarbon dates measured on terrestrial plant remains, resulting in a Bayesian time model calculated using Oxcal v.4.3.2 (ESM Table 1; Bronk Ramsey 2009). Reversed or outlying radiocarbon ages were not used for the age model. The chronology of the Late Glacial part of the pollen diagram was modified because there is only one radiocarbon date within the Younger Dryas. We have used ages for climatically caused vegetation developments, such as the strong rise of Juniperus, from other well dated Late Glacial sites to the north of the Alps to create a robust time model, based on the assumption that the late Würm vegetation development was synchronous over rather large areas (Table 1; Eusterhues et. al. 2002; Ammann et al. 2013). Otherwise, the modelled ages based on extrapolation would have been considerably too young. Additionally, we constructed an age-depth diagram which was also calculated using Oxcal v.4.3.2 (Fig. 2; Bronk Ramsey 2008, 2009). Loss-on-ignition analysis was carried out using the same sampling system as for pollen analysis. The pollen data from the Bodensee region used for comparison were obtained with the same methods and standards. The zonation of the pollen diagram from Großer Ursee follows Firbas (1949, 1952) for the Firbas zones (such as Preboreal, Boreal etc.) and Bastin (1979) for the description of the pollen events. The pollen zones are defined by the dominance of one or several pollen types, the subzones by significant changes in subdominant taxa. The zone definition follows Mangerud et al. (1974) and Hedberg (1976).

Age-depth diagram for Großer Ursee
Results and interpretation
The pollen results are presented in Figs. 3 and 4. According to changes of taxon dominance the diagram is divided into nine pollen zones which are further sub-divided into sub-zones (Fig. 3, ESM Table 2).

Pollen diagram of Großer Ursee, late Würm and Holocene part

Pollen diagram of Großer Ursee, Subatlantic
Lithology
The lake deposits reach a thickness of 1,107 cm in the deepest part, underlain by lacustrine basin clay which was deposited during the ice retreat and is probably several metres thick. From 1,107 to 1,062 cm a silty to sandy calcareous mud with organic material and olive to dark grey in colour overlies the light grey basal lake clay. This mud deposit represents the Bølling and Allerød warmer periods. From 1,062 to 976 cm the grey calcareous mud has a decreased organic content and increased clastic material, reflecting the sedimentation conditions during the Younger Dryas. From 976 cm upwards, a dark grey mud consisting of detritus with around 10% calcium carbonate was formed during the Preboreal and Boreal (Fig. 5). At the end of the Boreal, the organic content of the mud increases clearly to over 50% for the first time (Fig. 5) and the colour turns to brownish grey. This sediment was deposited during the Atlantic and the Subboreal periods. From 620 cm onwards, layers with higher amounts of clastic material occur intercalated in the mud (Fig. 5). These layers are connected with erosion caused by agricultural activity and woodland clearing in the lake catchment in the Subatlantic.

Loss-on-ignition graph of Großer Ursee
Pollen zones
Vegetation development starts at the bottom of the profile with dominance of non-arboreal pollen and high Salix values, indicating the Oldest Dryas steppe vegetation with herbs and dwarf shrubs in pollen zone PZ 1. Then a Juniperus shrub/dwarf shrub stage of the earlier part of the Bølling interstadial is visible in PZ 2, with a short Juniperus maximum of 40% and fast declining NAP. The later part of the Bølling interstadial with a Betula maximum of more than 50% in PZ 3 is also a short period. PZ 4 is characterized by predominance of Pinus sylvestris. It can be divided into three subzones, PZ 4a with increasing Pinus to a first maximum of 80% and decreasing Betula, 4b with a NAP increase to 20%, together with an increase of Juniperus to more than 5%, in combination with a slight decrease of Pinus and 4c with a second Pinus maximum. PZ 4a corresponds to the Allerød interstadial, 4b to the Younger Dryas stadial and 4c marks the transition to the Preboreal. PZ 5 is a zone of rapid vegetation change, from Pinus dominance to a short peak of Betula and Ulmus, before Corylus avellana becomes dominant, corresponding to the Preboreal and the transition to the Boreal. PZ 6, corresponding to the Boreal is characterized by dominance of Corylus. Three subzones can be distinguished: PZ 6a has high Ulmus percentages and increasing Corylus. In 6b Corylus has its maximum of more than 50%. It decreases slowly in 6c and Ulmus and especially Fraxinus excelsior increase. The longest lasting zone, more than four millennia, is PZ 7, the Atlantic period, which is characterized by dominance of Corylus and Ulmus, high percentages of Quercus, Tilia and Fraxinus together with increasing Fagus sylvatica and Abies alba. Three subzones can be distinguished: Corylus still dominates during PZ 7a. Among the tall trees only Picea is present. In 7b, Corylus and Ulmus have more or less the same values, Fraxinus reaches its maximum, Tilia is already decreasing and records of Fagus and later also of Abies, are continuous. In 7c, the Atlantic, Fagus increases and also Abies to a lesser degree. PZ 8, the Subboreal, is characterized by predominance of Fagus, while Abies, Corylus and Quercus are subdominant. Picea increases to 10%. Four subzones are visible: in PZ 8a Fagus increases and Ulmus values are still above 5%. In 8b Fagus has its maximum of about 40%. In 8c Fagus is reduced to about 25% and Betula, Corylus, Abies and NAP have their maxima. In 8d Fagus has recovered and the NAP is lower. PZ 8c, the first clear phase of woodland clearance, can be linked to the La Tène and Roman period. PZ 9, the Subatlantic, is shown in Fig. 4 in more detail. It has predominance of NAP and subdominance of Fagus, Picea and Corylus. Three subzones are visible: PZ 9a has high Betula percentages and increasing NAP, with a maximum of 25% at ad 1200. In 9b the NAP is slightly reduced, Pinus increases and Cannabis values are about 10%. In 9c the NAP has a second maximum, from high percentages of Poaceae. Fagus, Abies and Quercus decrease.
Short summary of the woodland development around Großer Ursee
In the Bølling interstadial the spread of Betula occurs, preceded by a shrub phase with Juniperus and Hippophaë. Afterwards Pinus spreads, leading to open pine woods with Betula. The climatic cooling during the Younger Dryas is indicated by decreased signs of woodland as shown mainly by an increase of Juniperus. During the Preboreal a Betula expansion takes place, which is characteristic for the Rhine glacier area (Eusterhues et. al. 2002), while Ulmus and Corylus immigrate and start to spread. Later Acer, Tilia, Quercus and Fraxinus also become established in the western Allgäu, forming the woodlands there during the Boreal and the Atlantic periods. Pinus decreases, because it was outcompeted. Through Atlantic times Fagus and, with a delay, Abies slowly become established. During the Subboreal, dense woods dominated by Fagus were common. With the spread of Fagus and Abies, there was a synchronous decline of Ulmus in the woodland. From ca. 2500 cal bc onwards, human activities are detected, indicated by the regular occurrences of Plantago lanceolata, Rumex, Artemisia, Triticum-type, Hordeum-type and Cerealia undiff. The loss-on ignition results indicate erosion phases in the second half of the Holocene, which are connected to human impact (Fig. 5). So, for example, the first phase of woodland clearance is detected, affecting mainly Fagus. Clear erosion phases are noticeable in the 2nd century bc, between the 2nd and 6th century ad, in the 10th, 13th, 15th and from the late 17th century onwards. They correlate with phases of increased human impact indicated in the pollen record or with climatic deterioration.
Discussion
Comparison of the vegetation development in the western Bodensee region and the eastern Allgäu
A comparison of the pollen results from Großer Ursee with the western Bodensee region shows some differences, which can partly be explained by the differences in climate and altitude (Figs. 1 and 6, Rösch 1990, 1992, 1993, 2013; Lechterbeck 2000; Rösch et al. 2014a, b; Rösch and Lechterbeck 2016; Rösch and Wick 2018, 2019; Lechterbeck and Rösch accepted). The results from Hornstaad were chosen as an example of the vegetation development at Bodensee (Rösch 1992, 1993). The main causes for the differences between this area and the Allgäu are the increasing total annual precipitation sum and the decreasing mean annual temperatures from west to east. During the early Holocene, before the expansion of tall trees, there are no or only slight differences in the vegetation development in other landscapes of central Europe. So, Ulmus is much more abundant in the western Allgäu than in the western Bodensee region, and the Ulmus decline around 4000 bc is very clear there, whereas in the Allgäu it is very slow and lasted until 2000 bc. Fagus arrived in the Allgäu shortly before 6000 bc, but spread very slowly there, starting its full expansion more than 2,000 years later. It spread very rapidly around 5000 bc in the western Bodensee area, 1,000 years after its arrival and it remained dominant there from 3000 bc until ad 500, with only a short interruption in the La Tène and Roman period. There are six phases of Fagus dominance around Bodensee, each lasting for one or more centuries. The first four, date to about 4500, 3500, 2500 and 1500 bc, are interrupted by maxima of Corylus and Betula. The last two date to about 500 bc and ad 500 and they start with maxima of Betula and are interrupted by maxima of Quercus. In contrast, at Großer Ursee, Quercus and Corylus are never dominant after the Fagus increase and show only slight fluctuations. Most of the weak peaks at Großer Ursee can be correlated in time with the strong peaks at sites around Bodensee. Another difference between the two regions is the record of Abies during the last five millennia. At Großer Ursee, Abies is subdominant from 3000 bc onwards, but around western Bodensee it plays only a minor role and seldom exceeds 5%. There is also no phase of Abies dominance preceding the Fagus phase at Großer Ursee, as in the Schwarzwald, Vosges or southwestern Alps (Drescher-Schneider 1978; Schloss 1979; Lang 2005), but Abies is much more frequent than at Bodensee, playing a secondary role after Fagus from 3000 bc onwards. Even bigger is the difference with Picea, which is so rare in the pollen records from the western Bodensee area before the 19th century ad that its actual occurrence there is dubious. In the western Allgäu it established itself as a minor woodland component about 3000 bc and became an important part during the medieval period. In contrast, in the eastern Allgäu Picea had already spread there in the Atlantic period. Also noteworthy is a rather strong representation of Ulmus at Großer Ursee even after the elm decline, with an increase up to 5% in modern times. So, in the relatively undisturbed Allgäu woodlands Fagus could dominate for more than three millennia, while around Bodensee the beech woods were strongly reduced by human impact several times during prehistory and only spread again when human pressure on the landscape lessened, as indicated by pollen and charcoal proxy evidence (Rösch and Lechterbeck 2016). These differences between the two regions may be partly the result of differing amounts of human activity, but they also seem to have been partly driven by differences in climate, insofar as the Allgäu landscape with its less favourable climate was later and less densely settled (Miera et al. 2019). The western Bodensee region has a climate more favourable for agriculture than the Allgäu, where especially the precipitation is higher. As a consequence, the Bodensee region was densely settled from the Late Neolithic onwards, whereas permanent human occupation is not evident in the Allgäu area before the late pre-Roman Iron Age. The fact that people farmed less in the Allgäu is therefore also caused by the climate.

Simplified pollen diagrams of Hornstaad, western Bodensee (Lake Constance) and Großer Ursee, Allgäu, showing the main differences in the vegetation changes since the late Würm
A slight increase of NAP is visible in the second half of the 5th millennium bc at Großer Ursee (Fig. 3). Plantago lanceolata as an indicator of fallow land and pasture (Behre 1981) and a few cereals also occur. This is during the very slow Fagus increase and hints at some kind of human impact with modest agricultural activity and woodland pasturing in the early Neolithic. At the same time, human impact at Bodensee is even weaker to nonexistent (Fig. 6). Only in the Hegau region, where Neolithic Linear Pottery settlements are known, are human impact phases with slight woodland clearance visible. In the late 5th and 4th millennia BC there are at least two very distinct phases of land use at Bodensee, indicated by secondary Corylus maxima, increased charcoal, more or less continuous cereal curves, but a very slight increase of NAP, if at all. In Fig. 6, these impact phases are indicated by declines of Fagus. At the same time, evidence of human impact is very slight or absent. A secondary Corylus maximum is indistinct. In the eastern part of the Allgäu the high resolution pollen records so far show no significant human impact (Küster 1988; Stojakowits 2014). In the 3rd millennium bc, human impact at Großer Ursee is still more or less absent, but the increase of Fagus and Abies continues there. At Bodensee at the same time there is evidence of the first human impact, which is much more clearly visible than previously, because it is indicated by a distinct increase of NAP, especially in the Hegau, which is correlated with the Glockenbecher (Bell Beaker) culture and the beginning of the early Bronze Age during the 2nd millennium BC. Three impact phases can be correlated with the early, middle and late Bronze Age, and three very weak phases of increased NAP during the 2nd millennium bc are also visible at Großer Ursee. However, the first clear phase of woodland clearance occurs here towards the end of the 1st millennium bc, in two sub-phases, which correspond most probably with the (late) pre-Roman Iron Age (La Tène) and the Roman period. At Bodensee there are two to four NAP maxima which can be distinguished during the 1st millennium bc, with values sometimes more than 20%. Großer Ursee has such values only during the short La Tène NAP maximum. During the first half of the 1st millennium ad, NAP is reduced there, increasing again in the second half, but with a short decrease towards the end of this millennium. In the 3rd millennium bc, human influence is still weak in the eastern Allgäu and only at Haslacher See and at Langegger Filz is there a delayed slight increase of NAP and cereals indicating small-scale agriculture (Küster 1988). During the 2nd millennium bc, in the eastern Allgäu weak phases of increased NAP are evident, too. There was even farming in the inner Alpine Illertal during the late Bronze Age (Schmeidl 1962). During the Iron Age, human impact is much reduced in the Allgäu compared to the Bodensee region, although there is one distinct land use phase at Haslacher See with a NAP maximum value around 20%, covering the Hallstatt period. During the La Tène period just the two sites, Haslacher See and Langegger Filz, show slightly increasing land use. After the Roman period a distinct Betula peak indicates regrowth of pioneer woodland on abandoned areas previously cleared. In the early Middle Ages NAP values rise again in the Allgäu. At Bodensee there is also a phase of increased woodland in the first half of the 1st millennium ad, followed by an increase of NAP afterwards, which is earlier and stronger than at Großer Ursee. Towards the end of this millennium, there is no decrease in signs of human activity. In the 2nd millennium ad, signs of clearance of woodland are strongest during the 11th and 12h centuries ad and again in modern times from the 18th century ad onwards. The second NAP rise is mainly of Poaceae. At Großer Ursee the events and chronological patterns of the last millennium are the same as at Bodensee, and this also applies to the eastern Allgäu. Due to afforestation mainly with Picea abies since the 19th century ad, openness decreases slightly.
In general, human impact before the medieval period was never strong enough to change the woodland cover completely by interrupting the dominance of Fagus at Großer Ursee, as was the case in the Bodensee region (Rösch 2018). The importance of the conifers Abies and Picea in woodland composition was greater in the Allgäu than in the lowlands.
Phases of land abandonment shown by short Betula peaks
Reductions of human impact are often more clearly visible in increases of the Betula curve, indicating pioneer woodland colonising abandoned fields, rather than by a decrease of human indicators and NAP values (Müller 1962; Rieckhoff and Rösch 2019). Großer Ursee is one of the pollen diagrams with the most short-term Betula peaks among the south-central European pollen results (Fig. 7).

Land abandonment phases indicated by short Betula peaks
Obviously, abandonment of previously cleared and farmed land was rather common, particularly from the Bronze Age onwards. In the Hegau region with its fertile soils and warm dry climate, which was settled early, there were only a few interruptions in land use in the Neolithic and Bronze Age. The profiles from the western Bodensee region show between 10 and 17 such interruptions. The Großer Ursee results show the most, 26, in spite of the fact that Neolithic abandonment phases are rare here. Even during the high medieval period, when land use around Bodensee was continuous, there are Betula peaks at Großer Ursee. Of particular interest are the chronological patterns. The most interruptions occurred in the late La Tène period. Considering the inaccuracy of the chronology we can suppose that there were interruptions at all sites, indicating not local but regional or supra-regional emigration as interpreted from the archaeological evidence (Rieckhoff 1995; Rieckhoff and Rösch 2019). Land abandonment was also common in the 14th and 17th centuries ad, evidently for climatic and historical reasons. The presence of Betula peaks like those of the 21st century bc or in Neolithic phases does not necessarily suggest abandonment of land, but rather a specific kind of land use like coppicing for a period of time. In a natural beech wood no regeneration from open land takes place and so there is no Betula peak. In the pre-alpine lowlands of southwest Germany, the climate gradient from north to south has an increasing total annual precipitation and decreasing mean annual temperatures as well. This creates the same pattern of land use as shown for regions around Bodensee and also Großer Ursee, which is also obvious in the pollen diagrams from around Ammersee, Landsberg, Bayern (Küster 1995) and from the eastern Allgäu (Küster 1988) closer to the Alps, so that the more unfavourable the climate is, the more interruptions of land use occur.
Occurrence of Trapa natans
Another striking difference between the Großer Ursee and Bodensee results is in the presence of Trapa natans (water chestnut, caltrop). The northernmost border of its distribution during most parts of the Holocene before the Subatlantic period was 850 km further to the north, reaching almost 64°N, as indicated by many fruit and pollen finds in southern Scandinavia (Gams 1926; Lang 1994; Vanhanen and Pesonen 2016). The occurrence of this annual floating leaf plant has been thought to be mainly affected by the water temperature, as a minimum of 20 °C is necessary during summer for flowering (Gams 1926). At Großer Ursee, water chestnut is indicated by the pollen record throughout almost the entire Holocene except the Preboreal, whereas in nine pollen profiles from the western Bodensee region with more than 5,000 pollen samples and more than 5 million pollen grains identified, only one single grain of T. natans has been found, from Buchensee, dating to the 1st century AD, and another grain from Hartsee, dating to the 16th century ad. T. natans has not been detected in lakes in southern Bavaria, because it is neither shown in the pollen diagrams nor mentioned in the published texts. Further east, in southern Bohemia, Czech Republic, numerous macrofossil remains were found dating to the Mesolithic period (Pokorny et al. 2010). Pollen analytical evidence of T. natans in the Czech Republic comes from Komořanské Jezero, together with macrofossils (Jankovská 1988; Houfková et al. 2017), dating to the Atlantic and Subboreal period, and one single grain from Knížecí pláně, dating to about 8,200 cal bp (Svobodová et al. 2001).
So, T. natans was very rare in the rather warm climate of the Bodensee region, but in the cooler climate of the western Allgäu it was common at Großer Ursee. It was also common at Federsee with a rather cool climate too (Karg 2006; Rösch 2006; Maier and Harwath 2011) and in southwestern Bavaria at Elbsee in the eastern Allgäu (Fig. 1; Stojakowits et al. unpublished data). In accordance with the air temperatures, Bodensee and the small lakes in its vicinity reach water temperatures of up to 25 °C during the summer, whereas the water of the higher Allgäu lakes is several degrees cooler. Therefore, the summer temperature cannot be the only reason for the presence or absence of T. natans, at least in this situation. Its presence at Großer Ursee until the 19th century is proof that it even survived the Little Ice Age, when mean July temperatures were even lower than the average temperature of 17 °C of today, which casts doubt on the assumption that a water temperature of at least 20 °C is needed for it to be able to flower (Gams 1926). Other possible reasons are the distribution of this useful plant by humans for its arrival, or changes to the nutrient status of lakes for its disappearance (Lang 1994). To gain better insight, we must consider the biology and ecology of this species (Gams 1926; Sebald et al. 1992; Oberdorfer 2001). As an annual floating leaf plant, its survival depends on the germination of its fruits. In autumn, the ripe nuts float on the water surface, later sinking to the bottom of the lake. According to new investigations, T. natans seeds germinate in a wide temperature range, with an alternating temperature of 14/6 °C, but freshly matured seeds need a cold stratification lasting at least nine weeks to overcome their dormancy (Phartyal et al. 2018). After germination in spring, a thin, 1–2 m long stem develops that connects the weak roots of the plant in the lake sediment with the leaf rosette floating on the water surface. Therefore Trapa is restricted to water 1–2 (2.5) m deep, although its maximum known growing depth reaches 3.6 m (Hummel and Kiviat 2004). Trapa usually grows in eutrophic, humus rich mud on the lake bed. Federsee is a large, eutrophic shallow water lake with an organic mud lake bed and an average water depth of 1 m, with a maximum of 3 m today, therefore ideally suited for T. natans. The smaller glacial lakes around Bodensee have, or had, only narrow littoral zones with shallow water, at least in prehistoric times. The Untersee, part of Bodensee, now has extensive littoral zones with shallow water, but as pollen analyses show, these developed mainly by increased silting up in the medieval period and later. The same is the case with south-eastern Buchensee with the only Trapa grain from the Bodensee region. Today it is a shallow lake with only 2 m water depth, but before the medieval period, it had a depth of at least 7 m. Furthermore, the littoral lake sediments in this region are very rich in lime and rather poor in organic matter. Bodensee and the small glacial lakes nearby were mostly oligotrophic before the medieval period, so perhaps the nutrient content of the water was too low for Trapa. The large lake itself probably has too strong waves and too great a seasonal change of the water table for floating water plants to succeed. These two reasons for the absence of T. natans in Bodensee have already been mentioned (Tolar et al. 2011). Altogether, there are enough ecological reasons why T. natans was, against the climatic trend, very rare in the Bodensee region, but common at Federsee and in the Allgäu. The consequences for other regions should be rethought, for perhaps the late Holocene extinction of T. natans in northern Europe was not caused by climatic change, but by the acidification of lakes and their transition from a mesotrophic/eutrophic to a dystrophic stage.
A general implication should be, that first all other possible reasons for frequency or distribution changes of plants should be checked before they are interpreted as evidence for climatic change. Schofield and Bunting (2005) explain the mid Holocene occurrence of T. natans in a mere (lake) in Holderness (East Yorkshire, UK) through a combination of a favourable climate and the availability of a suitable habitat. Cultivation of water chestnuts is known during the Neolithic period (for example, Vuorela and Alto 1982). However, there is no solid evidence that T. natans was cultivated in Meso- and Neolithic times at Großer Ursee. Nowadays T. natans does not occur at Großer Ursee any more, and its closest occurrence is near Ravensburg, the only one in southwest Germany outside of the northern upper Rhine area (Naturkundemuseum 2019). In the eastern Allgäu there is actually no water chestnut at all (Bayern Flora 2019).
Conclusions
The late Würm, early and middle Holocene vegetation development until the expansion of Fagus at Großer Ursee is very similar to the situation around Bodensee and in southern central Europe in general. It consists of a succession of different woodland types caused by the climatic change at the transition between the late Würm and the Holocene and controlled by the development of soils, the different migration speeds of the various taxa and the competition between them, in stages which are well known (Firbas 1949, 1952). Rather special from the point of view of lowland landscapes is the high importance of Ulmus at Großer Ursee. Fagus, after its rather late establishment there, has a vegetation history characteristic of this region, with clear differences to elsewhere and it remained the dominant woodland tree for more than three millennia, from 3000 bc until ad 500. The dominance of Fagus was not interrupted by increases of Corylus, Betula and Quercus as in the Bodensee region or other low-lying land, most probably as a consequence of weaker human impact. There is also no phase of Abies dominance preceding the Fagus phase as in the Schwarzwald, Vosges or southwestern Alps, but Abies is much more frequent than around Bodensee, playing a secondary role after Fagus from 3000 bc onwards. Picea, gains importance and overtakes Abies in the medieval period, but it is absent further to the west, especially at low elevations, before its planting as a forest tree in modern times.
Strong human impact is not evident at Grosser Ursee before the La Tène culture in the late pre-Roman Iron Age. Earlier human impact, indicated by a Corylus or Quercus increase, charcoal increase, NAP increase, or human indicators such as Cerealia or Plantago lanceolata, is present, but very weak. Of particular interest is the evidence of human impact in the early Neolithic, which is in contrast to the Bodensee region, where distinct human impact is not visible before the late Neolithic. After the expansion of Fagus, Abies and Picea from around 3000 bc until 200 bc, traces of human impact at Großer Ursee are so weak that local occupation around the lake is unlikely. This changes during the late pre-Roman Iron Age, when the landscape was obviously settled by farmers. In the eastern Allgäu, the first unequivocal phases of woodland clearance occurred earlier than at Großer Ursee, but clear human impact also started much later there than around western Bodensee.
Short-term (less than 100 years) interruptions of human impact, indicated by distinct peaks in the Betula curves, occur in all profiles from the Neolithic to modern times. They are most frequent at Großer Ursee and least frequent in the Hegau, obviously depending on the climate. It seems that marginal sites were often given up, while climatically better areas for farming show fewer interruptions in land use. Phases with very common interruptions occur at the time when Celtic tribes migrated before the Roman occupation, during the migration period of Germanic tribes in the 4–5th centuries ad, in the 14th century with the plague and the end of the medieval warm period and in the 17th century ad at the beginning of the Little Ice Age, with their well-known climatic and demographic crises.
It is worth mentioning that the regular occurrence of Trapa natans in the Allgäu is more frequent than in adjacent warmer landscapes and therefore against the climatic trend, and this appears to have been caused by factors other than the climate alone, an implication worth checking in other regions, too. In some cases, water chestnuts were cultivated according to archaeological evidence from lake sites as at Federsee. Another study, from Finland, has shown that habitat availability for T. natans and local conditions should also be taken into account (Korhola and Tikkanen 1997). In general, the cool and wet climate of the Allgäu, compared with the Bodensee region, seems, at least in the second half of the Holocene, to have determined the vegetation not directly, but by influencing human impact, which started earlier but was weaker under the less favourable climatic conditions there.
References
Ammann B, van Leeuwen JFN, van der Knaap WO, Lischke H, Heiri O, Tinner W (2013) Vegetation responses to rapid warming and to minor climatic fluctuations during the Late-Glacial Interstadial (GI-1) at Gerzensee (Switzerland). Palaeogeogr Palaeoclimatol Palaeoecol 391 Part B 1:40–59
Bastin B (1979) Essai de définition d’une terminologie précise applicable au commentaire des diagrammes polliniques se rapportant au quaternaire. Bull Soc R Bot Belg 112:7–12
Bayern Flora (2019) https://daten.bayernflora.de/de/info_pflanzen.php. Accessed 20 Feb 2018
Behre K-E (1981) The interpretation of anthropogenic indicators in pollen diagrams. Pollen Spores 23:225–245
Berglund BE, Ralska-Jasiewiczowa M (1986) Pollen analysis and pollen diagrams. In: Berglund BE (ed) Handbook of Holocene palaeoecology and palaeohydrology. Wiley, Chichester, pp 455–484
Bronk Ramsey C (2008) Deposition models for chronological records. Q Sci Rev 27:42–60
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51:337–360
Drescher-Schneider R (1978) Pollenanalytische Untersuchungen zur Kenntnis der spät- und postglazialen Vegetationsgeschichte am Südrand der Alpen zwischen Turin und Varese (Italien). Bot Jahrb Syst Pflanzengesch Pflanzengeogr 100:26–109
Engelhardt B (2006) Wie die Bayern Bauern wurden: Das Neolithikum. In: Gesellschaft für Archäologie in Bayern EV (ed) Archäologie in Bayern—Fenster zur Vergangenheit. Pustet, Regensburg, pp 54–75
Eusterhues K, Lechterbeck J, Schneider J, Wolf-Brozio U (2002) Late- and post-glacial evolution of Lake Steisslingen (I): sedimentary history, palynological record and inorganic chemical indicators. Palaeogeogr Palaeoclimatol Palaeoecol 187:341–371
Firbas F (1949) Spät- und nacheiszeitliche Waldgeschichte Mitteleuropas nördlich der Alpen, vol 1. Allgemeine Waldgeschichte, Fischer, Jena
Firbas F (1952) Spät-und nacheiszeitliche Waldgeschichte Mitteleuropas nördlich der Alpen, vol 2. Waldgeschichte der einzelnen Landschaften, Fischer
Flint CL (2015) Progress in agriculture. Arkose Press. https://openlibrary.org/publishers/Arkose_Press
Gams H (1926) Hydrocaryáceae. In: Hegi G (ed) Illustrierte Flora von Mitteleuropa, vol V/2. Hanser, München, pp 882–894
Hedberg HD (1976) International stratigraphic guide. Wiley, New York
Hornig W, Mickley W, Stahr K (1991) Genese, Eigenschaften und Verbreitung der Böden im Westallgäuer Hügelland. Jahrb Geol Landesamt Baden-Württemberg 33:199–217
Houfková P, Bešta T, Bernardová A, Vondrák D, Pokorný P, Novák J (2017) Holocene climatic events linked to environmental changes at Lake Komořany Basin, Czech Republic. Holocene 27:1,132–1,145
Hummel M, Kiviat E (2004) Review of world literature on water chestnut with implications for management in North America. J Aquat Plant Manag 42:17–28
Jankovská V (1988) Palynologische Erforschung archäologischer Proben aus dem Komořanské jezero-See bei Most (NW-Böhmen). Folia Geobot Phytotaxon 23:45–77
Karg S (2006) The water chestnut (Trapa natans L.) as a food resource during the 4th to 1st millennia BC at Lake Federsee, Bad Buchau (southern Germany). Environ Archaeol 11:125–130
Keller O, Krayss E (1993) The Rhine-Linth glacier in the upper Würm: a model of the last alpine glaciation. Q Int 18:15–27
Keller O, Krayss E (2005) Der Rhein-Linth-Gletscher im letzten Hochglazial. Vierteljahrsschr Nat forsch Ges Zür 150:19–32
Korhola AA, Tikkanen MJ (1997) Evidence for a more recent occurrence of water chestnut (Trapa natans L.) in Finland and its palaeoenvironmental implications. Holocene 7:39–44
Küster H (1988) Vom Werden einer Kulturlandschaft. Vegetationsgeschichtliche Studien am Auerberg (Südbayern). (Quellen und Forschungen zur prähistorischen und provinzialrömischen Archäologie 3) VCH, Acta Humaniora, Weinheim
Küster H (1995) Postglaziale Vegetationsgeschichte Südbayerns. Geobotanische Studien zur prähistorischen Landschaftskunde. Akademie Verlag, Berlin
Lang G (1994) Quartäre Vegetationsgeschichte Europas. Fischer, Jena
Lang G (2005) Seen und Moore des Schwarzwaldes als Zeugen spätglazialen und holozänen Vegetationswandels. (Andrias 16) Staatliches Museum für Naturkunde, Karlsruhe
Lechterbeck J (2000) „Human Impact“ oder „Climatic Change“?: Zur Vegetationsgeschichte des Spätglazials und Holozäns in hochauflösenden Pollenanalysen laminierter Sedimente des Steißlinger Sees (Südwestdeutschland). (Tübinger mikropaläontologische Mitteilungen 25) Institut und Museum für Geologie und Paläontologie, Tübingen
Lechterbeck J, Rösch M (accepted) Contributions to the European pollen database, Western Lake Constance (Germany): Böhringer See. Grana
Maier U, Harwath A (2011) Detecting intra-site patterns with systematic sampling strategies. Archaeobotanical grid sampling of the lakeshore settlement Bad Buchau-Torwiesen II, southwest Germany. Veget Hist Archaeobot 20:349–365
Mangerud J, Andersen ST, Berglund BE, Donner JJ (1974) Quaternary stratigraphy of Norden, a proposal for terminology and classification. Boreas 3:109–126
Meteoblue (2019) https://www.meteoblue.com/de/wetter/vorhersage/modelclimate/isny-im-allg%C3%A4u_deutschland_2895638. Accessed 11 Nov 2019
Miera JJ, Henkner J, Schmidt K, Fuchs M, Scholten T, Kühn P, Knopf T (2019) Neolithic settlement dynamics derived from archaeological data and colluvial deposits between the Baar region and the adjacent low mountain ranges, southwest Germany. E&G Q Sci J 68:75–93
Müller H (1962) Pollenanalytische Untersuchung eines Quartärprofils durch die spät- und nacheiszeitlichen Ablagerungen des Schleinsees (Südwestdeutschland). Geol Jahrb 79:493–526
Müller T, Oberdorfer E, Philippi G (1974) Potentielle natürliche Vegetation von Baden-Württemberg. (Veröffentlichungen der Landesstelle für Naturschutz und Landschaftspflege in Baden-Württemberg/Beiheft 6) Landesstelle für Naturschutz und Landschaftspflege, Ludwigsburg
Nadler M (2006) Der lange Weg in eine neue Gesellschaft: Die Kupferzeit. In: Gesellschaft für Archäologie in Bayern EV (ed) Archäologie in Bayern—Fenster zur Vergangenheit. Pustet, Regensburg, pp 76–99
Naturkundenmuseum (2019) https://www.flora.naturkundemuseum-bw.de/start2.htm. Accessed 11 Nov 2019
Oberdorfer E (2001) Pflanzensoziologische Exkursionsflora für Süddeutschland, 8th edn. Ulmer, Stuttgart
Phartyal SS, Rosbakh S, Poschlod P (2018) Seed germination ecology in Trapa natans L., a widely distributed freshwater macrophyte. Aquat Bot 147:18–23
Pokorný P, Šida P, Chvojka O, Žáčková P, Kuneš P, Světlík I, Veselý J (2010) Palaeoenvironmental research of the Schwarzenberg Lake, southern Bohemia, and exploratory excavations of this key Mesolithic archaeological area. Památky Archeologické 101:5–38
Preuß J (2008) Das Neolithikum in Mitteleuropa: Kulturen—Wirtschaft—Umwelt vom 6. bis 3. Jahrtausend v.u.Z.—Übersichten zum Stand der Forschung, 2nd edn. Beier & Beran, Weißbach
Rieckhoff S (1995) Süddeutschland im Spannungsfeld von Kelten, Germanen und Römern. Studien zur Chronologie der Spätlatènezeit im südlichen Mitteleuropa. (Trierer Zeitschrift für Geschichte und Kunst des Trierer Landes und seiner Nachbargebiete, Beiheft 19). Rheinisches Landesmuseum, Trier
Rieckhoff S, Rösch M (2019) Ein keltischer Exodus? Archäologisch-Botanische Überlegungen zum Übergang Eisenzeit Römische Kaiserzeit in Südwestdeutschland. In: Leskovar J, Karl R (eds) Interpretierte Eisenzeiten—Fallstudien, Methoden, Theorie 8. Linzer Gespräche zur interpretativen Eisenzeitarchäologie. Studien zur Kulturgeschichte von Oberösterreich, pp 57–87
Rösch M (1990) Vegetationsgeschichtliche Untersuchungen im Durchenbergried. In: Landesdenkmalamt Baden-Württemberg (ed) Siedlungsarchäologie im Alpenvorland 2. (Forschungen und Berichte zur Vor- und Frühgeschichte in Baden-Württemberg 37) Theiss, Stuttgart, pp 9–64
Rösch M (1992) Human impact as registered in the pollen record: some results from the western Lake Constance region, Southern Germany. Veget Hist Archaeobot 1:101–109
Rösch M (1993) Prehistoric land use as recorded in a lake-shore core at Lake Constance. Veget Hist Archaeobot 2:213–232
Rösch M (2006) Wassernuß. In: Beck H, Geuenich D, Steuer H (eds) Reallexikon der Germanischen Altertumskunde, Wagen und Gewichte—Wielandlied, vol 33, 2nd edn. De Gruyter, Berlin, pp 293–295
Rösch M (2013) Change of land use during the last two millennia as indicated in the pollen record of a profundal core from Mindelsee, Lake Constance region, southwest Germany. In: von Carnap-Bornheim C, Dörfler W, Kirleis W, Müller J, Müller U (eds) Von Sylt bis Castanas, Festschrift für Helmut Johannes Kroll. (Offa 69/70, 2012/13) Wachholtz, Neumünster, pp 355–370
Rösch M (2018) Evidence for rare crop weeds of the Caucalidion group in Southwestern Germany since the Bronze Age: palaeoecological implications. Veget Hist Archaeobot 27:75–84
Rösch M, Hahn S (2016) Besiedlung und Landnutzung im Allgäu von der Jungsteinzeit bis zur Neuzeit—ein interdisziplinäres Forschungsprojekt: Isny, Kreis Ravensburg. Archäologische Ausgrabungen in Baden-Württemberg 2015:45–50
Rösch M, Lechterbeck J (2016) Seven Millennia of human impact as reflected in a high resolution pollen profile from the profundal sediments of Litzelsee, Lake Constance region, Germany. Veget Hist Archaeobot 25:339–358
Rösch M, Wick L (2018) Contributions to the European pollen database 41. Western Lake Constance (Germany): Überlinger See. Mainau Grana 58:78–80
Rösch M, Wick L (2019) Contributions to the European pollen database 43. Western Lake Constance (Germany): Buchensee. Grana 58:308–310
Rösch M, Kleinmann A, Lechterbeck J, Wick L (2014a) Botanical off-site and on-site data as indicators of different land use systems: a discussion with examples from Southwest Germany. In: Bittmann F, Gerlach R, Rösch M, Schier W (eds) Farming in the forest—ecology and economy of fire in prehistoric agriculture. (Vegetation History and Archaeobotany 23, Suppl 1). Springer, Berlin, pp 121–133
Rösch M, Kleinmann A, Lechterbeck J, Wick L (2014b) Erratum to: botanical off-site and on-site data as indicators of different land use systems: a discussion with examples from Southwest Germany. Veget Hist Archaeobot 23:647–648
Schloss S (1979) Pollenanalytische und stratigraphische Untersuchungen im Sewensee. Ein Beitrag zur spät- und postglazialen Vegetationsgeschichte der Südvogesen. (Dissertationes Botanicae 52) Cramer, Vaduz
Schmeidl H (1962) Der bronzezeitliche Prügelweg im Agathazeller Moor. Bayerische Vorgeschichtsblätter 27:131–142
Schofield JE, Bunting MJ (2005) Mid-Holocene presence of water chestnut (Trapa natans L.) in the meres of Holderness, East Yorkshire. UK Holocene 15:687–697
Scholz H (2016) Bau und Werden der Allgäuer Landschaft. Schweizerbart, Stuttgart
Sebald O, Seybold S, Philippi G (1992) Die Farn- und Blütenpflanzen Baden-Württembergs, vol 4. Ulmer, Stuttgart
Svobodová H, Reille M, Goeury C (2001) Past vegetation dynamics of Vlatavský luh, upper Vltata river valley in the Šumava mountains, Czech Republic. Veget Hist Archaeobot 10:185–199
Stojakowits P (2014) Pollenanalytische Untersuchungen zur Rekonstruktion der Vegetationsgeschichte im südlichen Iller-Wertach-Jungmoränengebiet seit dem Spätglazial. Dissertation, Universität Augsburg
Tolar T, Jacomet S, Velušček A, Čufar K (2011) Plant economy at a late Neolithic lake dwelling site in Slovenia at the time of the Alpine Iceman. Veget Hist Archaeobot 20:207–222
Vanhanen S, Pesonen P (2016) Wild plant gathering in Stone Age Finland. Q Int Part A 404:43–55
Vuorela I, Aalto M (1982) Palaeobotanical investigations at a Neolithic dwelling site in southern Finland, with special reference to Trapa natans. Ann Bot Fenn 19:81–92
Wright HE Jr (1977) Environmental change and the origin of agriculture in the old and new worlds. In: Reed CA (ed) Origins of agriculture. Mouton, The Hague, pp 281–318
Acknowledgements
Open Access funding provided by Projekt DEAL. Oliver Korch, Stefan Hahn, Daniel Quentin, Natalia Ryabogina and Katrin Unger helped with the field work. SH provided eight pollen samples from his unpublished doctoral thesis. The owner of the lakes, Fürst von Waldburg-Zeil, gave us permission to take cores and to use his fishing hut for assembling our equipment and setting down the raft on the water.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T. Giesecke.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rösch, M., Stojakowits, P. & Friedmann, A. Does site elevation determine the start and intensity of human impact? Pollen evidence from southern Germany. Veget Hist Archaeobot 30, 255–268 (2021). https://doi.org/10.1007/s00334-020-00780-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-020-00780-4