
Abstract
This paper reports on new research into the timing and nature of post-glacial environmental change at Lake Flixton (North Yorkshire, UK). Previous investigations indicate a succession of wetland environments during the early Holocene, ultimately infilling the basin by ca 7,000 cal bp. The expansion of wetland environments, along with early Holocene woodland development, has been linked to changes in the human occupation of this landscape during the Mesolithic (ca 11,300–6,000 cal bp). However, our understanding of the timing and nature of environmental change within the palaeolake is poor, making it difficult to correlate to known patterns of Mesolithic activity. This paper provides a new record for both the chronology and character of environmental change within Lake Flixton, and discusses the implications for the Mesolithic occupation of the surrounding landscape.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Since the late 1940s evidence for intensive phases of early prehistoric activity has been recorded around Lake Flixton in the eastern Vale of Pickering (North Yorkshire, UK) (Fig. 1a, b). The lake formed at the start of the Late-glacial interstadial (ca 14,600 ka cal bp) and gradually in-filled with calcareous and organic sediments that preserved a rich record of both the human and environmental history of the landscape (e.g. Clark 1954; Walker and Godwin 1954; Mellars and Dark 1998; Conneller and Schadla-Hall 2003; Taylor 2011; Milner et al. 2018). The area is best known for the early Mesolithic site of Star Carr where a large assemblage of bone and antler artefacts, faunal remains and worked flint were recovered from sediments at the former lake edge (Clark 1954). However, a further 20 sites, dating to the final Palaeolithic, terminal Palaeolithic and the early and late Mesolithic have also been recorded (Fig. 2) (Conneller and Schadla-Hall 2003). Early Mesolithic activity (ca 11,300–9,500 cal bp) was particularly intensive. Recent work at Star Carr has shown that the site was occupied for ca 800 years, with evidence for the construction of large timber platforms, post-built structures, and the deliberate management of lake-edge vegetation (Dark 1998a; Conneller et al. 2012; Milner et al. 2018). Other sites also show evidence for multiple phases of occupation, in some cases spanning centuries (Conneller and Schadla-Hall 2003; Taylor 2018), as well as evidence for post-built structures (Taylor and Gray Jones 2009) and controlled burning of the wetland vegetation (Cummins 2003). Together this suggests a long-term commitment to particular places in this landscape that stands in contrast to established views of the period (Conneller et al. 2012; Milner et al. 2018). Late Mesolithic occupation (ca 9,500–6,000 cal bp) is also known from multiple locations, in some cases continuing from the earlier part of the period. Whilst these sites have been less extensively studied there is evidence for comparably intensive forms of activity including the use of structures (Taylor 2012) and management of both wetland and terrestrial vegetation through burning (Cummins 2003).

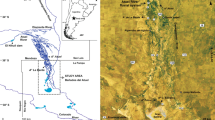
Maps showing the location of the study area; a the study area in relation to northwest Europe, b Lake Flixton in relation to the present-day North Sea coastline and the coastal towns of Scarborough and Filey

Map of Lake Flixton showing the approximate extent of the lake at the start of the Mesolithic, based on an estimated lake level of 24 m a.s.l. (see Taylor and Alison 2018), and taking into account the local topography. a Survey area at Flixton School House; b position of Dark’s (1998b) deep lake profile. Known areas of Mesolithic activity are marked as follows: 1: Flixton School House Farm. 2: Flixton School Field. 3: Barry’s Island. 4: Lingholme Farm Site A. 5: Killerby Carr. 6: Lingholme Farm Site C. 7–9: Cayton Carr. 10: Manham Hill. 11: Seamer C. 12: Seamer B and Rabbit Hill. 13: Seamer Carr K. 14: Seamer D. 15: Seamer Carr L and N. 16: Seamer Carr F. 17: Ling Lane. 18: Star Carr. 19: Flixton 9. 20: VP site D. 21: VP site E. 22: Woodhouse Farm. 23: Flixton Island site (1) 24: Flixton Island site (2) 25: No Name Hill
It has been argued that the scale of early Mesolithic activity was due to the rich and productive wetland environments forming within the lake, and that the subsequent in-filling of the basin, coupled with changes to the terrestrial woodland, led to the abandonment of sites and an overall decline in activity (Mellars 1998, pp 229–231). However, our ability to determine the significance of environmental change is hampered by a poor understanding of the timing and nature of wetland succession. The principal issue is chronology. There are only two reliably dated records for the spatial and temporal development of the wetlands, both of which relate to the lake-edge environments in the early Mesolithic. The most detailed comes from Star Carr where Dark (1998a) documented the development of the marginal vegetation by recording pollen and plant macrofossil profiles at intervals from the shore. However this record does not continue later than ca 10,000 cal bp and only extends ca 10 m into the lake. A more spatially extensive record was established by the author at Flixton School House Farm (henceforth Flixton SHF), where plant macrofossil analysis was carried out on samples from two cores, one at the former lake edge and the other 50 m into the basin (Taylor 2011). Again, however, this provides no information on the timing and character of wetland succession in the deeper parts of the lake during the late Mesolithic. Of the remaining dated records, most consist of individual pollen or plant-macrofossil profiles from sediments in the relatively shallow lake margins close to the shore (Cummins 2003; Taylor 2011), with only two profiles documenting the later expansion of wetlands into the deepest parts of the basin (Day 1996; Dark 1998b; Cummins 2003). As a result, there is little indication of the rate at which wetland succession occurred or the varying extents of the different environments other than at the very edge of the lake in the early Mesolithic.
The sequence of wetland succession and the effects of changes in the lake’s hydrology are also debated. Cloutman (1988b, p 31) argued that the hydrosere was reversed during the late Mesolithic by a rise in lake level that caused areas of Carr to be replaced by reedswamp. He also suggested that rising lake levels caused organic deposits to form over areas of previously dry ground above the lakeshore (Cloutman 1988a, p 14). However, Cummins (2003, pp 320–321) has argued that, whilst there were short-lived fluctuations in lake level, the hydrosere developed in a broadly linear manner with no evidence for a sustained rise in lake level.
These issues make it difficult to establish the potential effects that environmental change had upon the character of Mesolithic settlement. To resolve this a new programme of plant macrofossil analysis and radiocarbon dating was undertaken to establish a more detailed account of the spatial and temporal development of the lake environments. By bringing the results of this study together with previous palaeo-environmental and archaeological research in the area, it is now possible to describe the evolution of the wetland and terrestrial environments during the Mesolithic, and to discuss the implications for the human occupation of this landscape.
Materials and methods
Fieldwork was undertaken between 2008 and 2012 at Flixton SHF (Figs. 2, 3a, b) in conjunction with archaeological investigations of an early and late Mesolithic settlement (Taylor and Gray Jones 2009). Three trenches (F, H and I) were excavated to sample the deposits at the former lake edge (trench F) and the adjacent higher ground (trenches H and I), and an auger transect was recorded between them to produce a complete stratigraphic profile. Three cores (A–C) were taken to sample the deposits within the lake basin (Fig. 3a, b). The early Mesolithic sequences at trench F and core A have previously been reported (Taylor 2011).

a Coring and trench locations at Flixton School House Farm in relation to basal topography, and b detailed map showing location of the trenches. The location of the survey area in relation to the lake is shown in Fig. 2
Plant macrofossils
A contiguous sequence of samples, 35–50 mm thick, was cut from the east-facing section of trench F, bulk samples (1 l) were taken from stratigraphic units in trenches H and I, and cores A–C were taken using a Russian auger, carried off-site, and divided into 35–50 mm thick samples. These were sub-sampled (sample volumes: 50 cm3 from the cores, 100 cm3 from trench F, and 1,000 cm3 from H and I), disaggregated in 8% sodium hydroxide, and washed through nested sieves (2 mm–125 µm mesh). Each sieve fraction down to 125 µm was analysed using a Zeiss Stemi DV4 stereo microscope and, where necessary, a Zeiss light microscope (400 × and 1,000 × magnification). All macrofossils that could be identified to a specific taxon were counted, with the exception of fragments of Nymphaea alba seed and fern sporangia which were recorded in terms of abundance. The results were plotted quantitatively as the number of macrofossils of each taxon/species per sample using program C2 version 1.6.6 (Juggins 2010), with the exception of trenches H and I where the material was too sparse. Identifications were carried out using Cappers et al. (2006), Grosse-Brauckmann (1972, 1974, 1992), and a modern reference collection. Higher plant nomenclature follows Stace (2010).
Wood taxonomic identifications
Fifty-one samples were taken from nine layers of wood in the lower part of trench F (23.47–24.18 m a.s.l.). A further 10 samples were taken from a homogenous layer of wood toward the top of the trench (25.52–25.66 m a.s.l.) but were too poorly preserved for taxonomic identification. Wood from the intervening deposits consisted entirely of intrusive root and was not analysed. Samples were taken during the excavation, and the levels recorded using a Leica T300 total station. Thin sections of wood samples were removed from the radial, transverse and transversal planes, mounted on slides, examined under 10 × to 30 × magnification and identified using Schweingruber (1990).
Radiocarbon dating
Nineteen radiocarbon dates were obtained on samples from trench F and cores A–C to establish a chronology for the environmental profiles (Table 1 and ESM Table 1). Samples were taken from monolith tins/cores, and from bulk macrofossil samples. Dating was undertaken by the Oxford Radiocarbon Accelerator Facility with the exception of sample FSH-A-1, which was dated by Beta Analytic. Radiocarbon dates were calibrated using OxCal 4.2 (Bronk Ramsey 2009a, b) and the IntCal13 calibration curve (Reimer et al. 2013). Bayesian analysis has been undertaken using the functions in the OxCal software; deposition models (P-Sequence) were produced for trench F and core A, sequence models were produced for cores B and C, and the Difference function was used to estimate the durations of key stratigraphic units and hiatuses in peat formation. Dates are expressed at two standard deviations (95.4%) unless otherwise stated, and rounded to the nearest 10 years.
Results
Stratigraphy
Full sediment logs are available in supplementary files (ESM Table 2a, b). At the lake edge (trench F), the mineral substrate was overlain by a sequence of fine detrital mud, coarse reed peat and wood peat, sealed by a poorly sorted silty sand (ESM Table 2a). Above this was a coarse herbaceous detritus with two bands of much darker, denser humified peat. The herbaceous detritus and the uppermost layer of humified peat continued upslope into trench H, with the humified deposit extending into trench I. Overlying these was a deposit of wood peat, extending across all three trenches, which in trench F graded into a mixed herbaceous/woody detritus. Sealing the sequence in all three trenches was a highly humified organic sediment grading into topsoil. The basin deposits were characterised by marl overlain by fine detrital mud (ESM Table 2b). In core A, this was succeeded by a sequence of reed peat, herbaceous detritus and wood peat, whereas in cores B and C a coarse, fibrous detritus overlay the fine detrital mud.
Chronology
In trench F organic sedimentation began at 11,190–10,730 cal bp (Table 1). The deposition model shows a rapid and even rate of deposition up to 24.48 m a.s.l., becoming slightly slower and irregular above this (Fig. 4a). The duration over which the sand layer formed was modelled at 0–260 years. Hiatuses in peat formation were identified at the interface between each of the humified peat layers and the overlying sediment; the first (dated by OxA-22177 and OxA-22178) of between 70 and 350 years, the second (dated by OxA-22063 and OxA-22206) of between 10 and 230 years. The modelled age for the start of plant macrofossil zone F4b is poorly constrained but assuming an even rate of deposition in this part of the sequence it is likely that this transition occurred in the centuries around 8,000 cal bp.
In core A the onset of marl formation has been modelled at 11,690–10,460 cal bp (Table 1). The deposition model shows rates of sedimentation increasing from 22.86 m, at the transition from fine to coarse detrital mud (Fig. 4b). Due to the lack of dates above 23.55 m no attempt has been made to model the top of the sedimentary sequence or the upper zones in plant macrofossil profile A.

a Deposition model for trench F in relation to the stratigraphic sequence and the plant macrofossil zones, produced using OxCal v4.3.2 (Bronk Ramsey 2017) and the intCal13 calibration curve (Reimer et al. 2013). Probability distributions are shown at 95.4%. b Deposition model for core A in relation to the stratigraphic sequence and the plant macrofossil zones produced using OxCal v4.3.2 (Bronk Ramsey 2017) and the intCal13 calibration curve (Reimer et al. 2013). Probability distributions are shown at 95.4%
There were insufficient dates to create reliable deposition models for cores B and C. Dates from core B established the age for the start of plant macrofossil zones B3 (OxA-33254) and B4a (OxA-33255), whilst those from core C dated the start of zone C3 (OxA-33256) and C4 (OxA-33257).
Wood analysis
Taxonomically identifiable material was recorded from the lower deposits of trench F (23.47–24.18 m) (ESM Table 3). Most of the wood consisted of stems or side branches of Salicaceae (Salix or Populus spp.) or Betula spp., with Viburnum spp. occurring at 24.07 m, and Corylus avellana present from 24.07 m to 24.18 m. A single stump of Salix/Populus was recorded at 24.18 m. None of the material showed evidence of human modification, but sample 28F had been worked by beaver (ESM Table 3).
Plant macrofossil analysis
The results of the plant macrofossil analyses are presented in Figs. 5, 6 and 7 and Tables 2, 3, 4 and 5, with the exception of trenches H and I where the material was too sparse to be quantified. In Core C only the part of the sequence incorporating the decline and disappearance of aquatic material was investigated.

Plant macrofossil profile and chronology, trench F (principal taxa). Total numbers of macrofossils of each taxon/species are presented, with the exception of Nymphaea alba seed fragments and fern sporangia which are expressed on an abundance scale. The profile has been divided into six zones based on changes in the plant macrofossil assemblages. The results are displayed stratigraphically, and biological taxa are grouped by broad habitat. The chronology is derived from the deposition model (see Fig. 4a) and is expressed as median ages. For rare taxa see Fig. S1

Plant macrofossil profile and chronology, core A (all taxa). Total numbers of macrofossils of each taxon/species are presented, with the exception of Nymphaea alba seed fragments and fern sporangia which are expressed on a scale of abundance. The profile has been divided into six zones based on changes in the plant macrofossil assemblages. The results are displayed stratigraphically, and biological taxa are grouped by broad habitat. The chronology is derived from the deposition model (see Fig. 4b) and is expressed as median ages

a, b Plant macrofossil profiles, core B (top) and C (bottom). Total numbers of macrofossils of each taxon/species are presented, with the exception of N. alba seed fragments and fern sporangia which are expressed on a scale of abundance. Profile B has been divided into five zones, and profile C into four zones based on changes in the plant macrofossil assemblages. The results are displayed stratigraphically, and biological taxa are grouped by broad habitat. Dates are expressed as modelled ages (cal bp) derived from the sequence model (see Table 1)
In trenches H and I, fragments of Eupatorium achenes occurred throughout the deposits, but in very low quantities, with Thalictrum achenes, and Betula fruits occurring occasionally. Alnus fruits were present in the upper deposits coinciding with the transition to wood peat.
Discussion
Interpretation of the macrofossil and sedimentary sequences
Trench F. Zone F1: 11,190–10,730 to 10,690–10,340 cal bp. Sedimentation began in standing water, with Phragmites, probably P. australis (Haslam 1972), growing at the site. A range of emergent and aquatic taxa were present in the wider area (notably Nymphaea, Potamogeton spp., Carex (including C. paniculata and C. rostrata), Menyanthes, and Typha spp.), though given the tendency of plant material to accumulate at lake edges these were not necessarily growing locally (e.g. Birks 1973; Zhao et al. 2006; Koff and Vandel 2008). The local growth of trees (B. pubescens, B. pendula, P. tremula, and possibly Salix spp.) at the shore is demonstrated by large Betula and Salix/Populus stems and branches within the sediments.
Zone F2: 10,690–10,340 to 10,530–10,270 cal bp. The decrease in the quantity and range of macrofossils reflects a reduction in the volume of water reaching the sediments (see also Dark 1998a, p 131), which is tentatively interpreted as a shift to a seasonally flooded environment. Cladium and Typha spp. began to grow locally, probably in response to shallower conditions. Corylus, Salix/Populus, and Viburnum spp. were present on the adjacent dry ground.
Zone F3: 10,530–10,270 to 10,110–9,770 cal bp. The absence of aquatic material and the decline in other taxa mark the point where the deposits formed beyond the reach of the lake. The presence of a tree stump indicates the in-situ growth of Salix/Populus. Emergent vegetation (Cladium, Carex spp. and Typha spp.) was present nearby. Peat formation was interrupted at 10,170–9,890 cal bp by the deposition of a layer of silty sand.
Zone F4a: 10,110–9,770 to 8,620–7,450 cal bp. From the composition of the sediments and the macrofossil assemblage, herbaceous plants (including fen taxa Thalictrum and Eupatorium) had colonised the area as organic sedimentation resumed. Quasi-terrestrial conditions were present locally but it was sufficiently wet to support Menyanthes, at least for part of the time. The first band of humified peat reflects a drop in the local water table, causing the existing deposit to dry out and humify. This started at 9,590–9,480 cal bp with the deposition model indicating a hiatus in peat formation of 70–350 years, before organic sedimentation resumed at 9,440–9,150 cal bp.
Zone F4b: 8,620–7,450 to 7,420–7,090 cal bp. The increasing quantity of macrofossils suggests that the surface of the peat became slightly wetter (improving preservation) though the local vegetation remained unchanged. A subsequent fall in the water-table is marked by the second band of humified peat, which again caused the existing deposits to dry out. This began at 7,560–7,420 cal bp, with a hiatus in peat formation of 10–230 years, before organic sedimentation resumed at 7,430–7,250 cal bp.
F5: 7,420–7,090 to 6,970–6,440 cal bp. Alnus, Urtica, Carex spp. and possibly Corylus colonised the local area. The presence of Alnus cones and wood in the sediment indicating in-situ growth of alder.
Zone A1: 11,690–10,460 to 11,120–10,360 cal bp. Open water was present at the sampling point with beds of Characeae present locally.
Zone A2: 11,120–10,360 to 10,160–9,690 cal bp. The shift to organic sedimentation reflects a decrease in water depth. Macrofossil records indicate the local presence of aquatic (Potamogeton spp. and Nymphaea) and emergent (S. lacustris, Typha, and Carex) taxa.
Zone A3: 10,160–9,690 to 9,400–9,000 cal bp. The absence of aquatic macrofossils in the basal samples reflects a temporary fall in lake level leaving the deposits above lake-water level. Based on the deposition model this event had a duration of up to 370 years. Cladium and Carex spp. colonised the local area before a rise in lake level (modelled at 10,100–9,500 cal bp) submerged the sampling point, and aquatic and deep-water emergent taxa (Potamogeton spp. and S. lacustris) become re-established. The water-depth gradually shallowed in the upper half of the zone and a Cladium swamp formed at the sampling point.
Zone A4a: start: 9,400–9,000 cal bp; end: uncertain age. The consistent absence of aquatic material indicates that the deposits began to form beyond the extent of the lake. Herbaceous plants, including Hydrocotyle vulgaris, and also ferns were growing locally.
Zone A4b: Undated. Conditions became wetter leading to increased preservation of plant macrofossils, though the deposits remained terrestrial in character. Eupatorium, Urtica and Mentha grew locally.
A5: Undated. Alnus, Betula spp. and Carex spp. colonised the area. As with trench F the presence of Alnus cones and wood in the deposits indicate in-situ growth of alder.
B1: Undated. Open water was present at the sampling point with Characeae and Myriophyllum spicatum present in the surrounding area.
B2: start: uncertain age; end: 8,980–8,640 cal bp. The change to organic sediments marks a shift to shallower water. Aquatic and emergent plants (notably Nymphaea, Typha spp., S. lacustris and Carex spp.) grew locally.
B3: 8,980–8,640 to 8,520–8,340 cal bp. The absence of aquatic macrofossils marks the point where the deposits began to form beyond the reach of the lake. Cladium and Juncus spp. were growing locally.
B4a: start: 8,520–8,340 cal bp; end: age uncertain. The deposits remained terrestrialised. Eupatorium, Juncus spp. and ferns were growing locally.
B4b: Undated. Eupatorium, Alnus and Betula spp. grew locally or in the surrounding area. Failure to record Alnus cones or wood suggests that alder was not present at the sampling point.
C1: Undated. Open water was present at the sampling point with Characeae and Myriophyllum growing locally.
C2: start: uncertain age; end: 8,040–7,880 cal bp. The switch to detrital muds indicates a shallowing of the water, emergent and aquatic plants (notably Nymphaea, Potamogeton spp., S. lacustris, and Carex spp.) colonise the local area. Eupatorium seeds reflect the presence of fen nearby.
C3: 8,040–7,880 to 7,830–7,660 cal bp. The decline in the quantity of aquatic macrofossils suggests that the water depth at the sampling point continued to fall. Carex spp., Juncus spp. and Eupatorium grew locally.
C4: start: 7,830–7,660 cal bp; end: age uncertain. The absence of aquatic material marks the point where the deposits formed beyond the extent of the lake. Herbaceous plants, possibly including Cladium and Juncus spp., were present locally.
Trenches H and I
Based on the character of the basal organic deposits and the lack of aquatic plant macrofossils, deposition began in a terrestrial fen environment with Eupatorium and Thalictrum growing locally and Betula spp. present in the surrounding area. The deposits subsequently dried out creating a band of dark humified peat. This corresponds stratigraphically with the second band of humified peat recorded in trench F (see ESM Table 2). The start of this event (in trench F) dates to 7,560–7,420 cal bp. Fen environments returned before shrubs or trees (including Alnus, and Betula spp.) colonised the area resulting in the layer of wood peat. This transition to Carr was dated in trench F to 7,420–7,090 cal bp, and is likely to be broadly contemporary in trenches H and I.
Correlating the records
A radiocarbon chronology for the sequence of environmental change recorded in the plant macrofossil and sedimentary profiles is shown in Fig. 8. In addition, the increase in fen taxa in the upper part of trench F and core A, which reflects a shift to a slightly wetter depositional environment (zones F4b and A4b, respectively), is assumed to be broadly contemporary.

Evidence for hydrological changes
Two forms of hydrological change have been documented in this study; the gradual shallowing of the lake due to ongoing sedimentation in the basin, and more rapid fluctuations in lake-level/water-table. The chronological relationship between these changes is shown in Figs. 8, 9.

Schematic representation of the simplified sequence of environmental succession and the key hydrological events in each of the profiles. The vertical lines represent the modelled range of the dates for environmental succession (on the left of each column) and the hydrological changes (on the right of each column). Where no error range is shown the date of environmental change is unknown
At a general level, the data show the linear development of the lake environments. At each of the sampling points the plant macrofossil assemblage reflects an overall trend from wetter to drier conditions, with the macroremains of aquatic and emergent taxa giving way to macroremains from plants characteristic of wet or boggy ground, and fen or carr environments. With the exception of the earliest fluctuation in lake level recorded in core A (see below), once conditions at each sampling point were such as to be beyond the reach of the lake they remained terrestrialised. Similarly, the chronological relationship between the profiles shows that open water and aquatic environments were succeeded by increasingly shallow and ultimately terrestrial conditions within the basin over time (see Fig. 9).
This broad pattern was interrupted by changes in lake or local water-table levels. The most significant was the fluctuation in lake level recorded in core A. This began at 10,160–9,690 cal bp with a fall in lake level that left the deposits around the sampling point beyond the reach of the lake for up to 370 years. This is contemporary with the deposition of the sand layer in trench F (Figs. 8, 9). Assuming a causal as well as chronological relationship, the sand may have derived from erosion of sediments on the higher ground above the lake edge during this drier phase. However, there is no evidence for a shallowing of the water in cores B or C at this time (Fig. 8) indicating that the fall in lake level was not large enough to alter the environment in the deeper parts of the basin.
The subsequent rise in lake level in core A at 10,100–9,500 cal bp occurs within the same modelled age range as the transition from carr to fen in trench F (10,110–9,770 cal bp, zone F3–F4a) (Fig. 8). The correlation of these two events suggests that the rising level of the lake caused the deposits at the lake edge to become too wet to support the local growth of trees and thus resulted in fen development. This development is also seen elsewhere within the basin. In trench F the transition from carr to fen coincided with a switch from wood peat to a coarse herbaceous detritus (ESM Table 2a), and a similar stratigraphic horizon was recorded at Seamer Carr (Cloutman 1988b) and other parts of the basin (Paul Lane, personal communication).
The remaining events had a lesser impact on the wider environment. A transition to a slightly wetter environment was inferred from an increase in the quantity of macrofossils recorded in trench F and core A (zones F4b and A4b). The macrofossils show no evidence for a significant change in the local vegetation, and the effects were probably limited to a localised shift to wetter ground conditions that increased macrofossil survival. Two fluctuations in the local water-table were also identified from the sedimentary and chronological record in trench F. In both cases these were represented by a layer of heavily humified peat and a hiatus in organic sedimentation at the interface between the humified layer and the overlying deposit. These represent periods when the surface of the peat dried out causing the extant deposits to humify and peat formation to cease, before a subsequent shift to wetter conditions caused organic sedimentation to resume. There is no evidence for a comparable event in cores A–C, suggesting that the effects were relatively minor in character and thus limited to the edges of the basin.
Finally, there is no evidence that a rising lake level caused peat-forming environments to expand over previously dry ground (contra Cloutman 1988a, p 14); aquatic material was not recorded in the deposits in trenches H and I, and the sedimentary sequences and plant macrofossils are indicative of a terrestrial fen. Comparable environments were also recorded in Cloutman’s pollen analysis at Seamer Carr (Profile K5), which described a herb and fern-rich fen forming above the former lake shore (Cloutman 1988b, pp 28–30). Rather, the expansion of peat-forming environments over areas of dry ground was probably caused by the accumulation of peat at the lake edge, which would have inhibited drainage from the adjacent mineral sediments above the shore causing them to become waterlogged (a form of edaphic paludification) (Rydin and Jeglum 2006, pp 125–126).
The timing and nature of wetland succession in the Lake Flixton basin
Organic sediments began to form at the Flixton SHF lake edge in the centuries around 11,000 cal bp with marl accumulating in the deeper water at the site of core A (and potentially B and C). Species diversity was high within the lake. Phragmites was growing at trench F, and other emergent species, notably Menyanthes, Typha spp. and Carex (including C. paniculata and C. rostrata), were growing in the surrounding area. A similar range of taxa was recorded from contemporary sediments at Star Carr (Dark 1998a; Taylor and Alison 2018) and No Name Hill (Taylor 2011), indicating the rich and diverse nature of the marginal vegetation. Aquatic plants Nymphaea, Myriophyllum and Potamogeton spp. were also present, probably in deeper water further from the shore, along with beds of the aquatic algae Characeae. Trees (Populus tremula, Betula pubescens and B. pendula, and Salix spp.) were growing at the water’s edge. Beyond the lake, pollen analysis has recorded a landscape of grassland and scrub with Juniperus, Salix and Betula that was replaced by Betula woodland from ca 11,100 cal bp (Fig. 4 in Blockley et al. 2018).
The lake-edge environments changed rapidly over the following centuries. Emergent and aquatic species had colonised core A by ca 10,750 cal bp. At trench F the deposits were only seasonally submerged by ca 10,500 cal bp, encouraging the local growth of Cladium, with the deposits forming beyond the reach of the lake by ca 10,400 cal bp. This allowed trees (Salix/Populus) to spread onto the peat. The same sequence has been recorded at Star Carr where Cladium increases following the shift to a shallower environment (Taylor and Alison 2018, p 130), before the lake edge deposits became terrestrialised and resulted in fen and carr forming by 10,450–10,165 cal bp (Supp Info 20–21 in Blockley et al. 2018,). Assuming a similar rate of sedimentation in other parts of the basin the embayments at Seamer Carr, Lingholme, and Cayton (Fig. 2) were probably also becoming terrestrialised at this time (see Taylor et al. 2018b, p 49). However, within the main body of the lake the extent of these environments was still limited to the area close to the shore with vegetation characteristic of deeper water persisting at core A, and open water also persisting in the vicinity of cores B and C. On the shore, Viburnum spp. and Corylus were growing at Flixton SHF from ca 10,500 cal bp, the presence of the latter in the wider landscape also being reflected in the low but consistent presence of its pollen (e.g. Cloutman 1988b; Dark 1988a, b; Day 1996). The dating undertaken by Cloutman (1988a) shows that peat-forming environments were already established on areas of dry ground at Seamer Carr Site K (Fig. 2).
The period from ca 10,000 to ca 9,000 cal bp was characterised by changes to both the wetland and terrestrial environments. At ca 9,900 cal bp the lake level fell causing the area around core A to become terrestrialised. Emergent and aquatic plants probably expanded further into the basin creating an extensive area of wetland vegetation (though not as far as core B where open water conditions persisted). A subsequent rise in the level of the lake, up to 370 years later, resulted in flooding in the area around core A and reversion to an aquatic swamp. Deposits at the lake edge were now too wet to support growth of trees. Fen communities consisting of herbaceous plants (including Thalictrum and Eupatorium) replaced carr at trench F and other locations around the former shore. At ca 9,500 cal bp the local water-table fell causing the deposits around the edge of the lake to dry out and humify, before wetter conditions returned up to 350 years later. Peat was starting to encroach over previously dry ground at VP Site E and parts of the Star Carr peninsula (see Fig. 2) by this time (see Cloutman 1988a), and was probably also developing at Flixton SHF. On drier ground Corylus became the principal component of the woodland, an event dated at Star Carr to 10,250–9,730 cal bp (8,940 ± 90 bp. OxA-4377) (Dark 1988a, p 133), and Ulmus, and to a lesser extent Quercus, began to expand into the area, shading out much of the understory vegetation (Day 1996, pp 16–17; Dark 1998b, p 170).
Despite these hydrological changes the lake environments continued to develop in the manner of a classic hydrosere. By ca 9,200 cal bp the deposits at core A were forming beyond the reach of the lake water. Emergent species were probably already present around core B, and by ca 8,800 cal bp had been replaced by Cladium fen, creating a zone of terrestrialised wetland that extended lakewards at least 150 m from the former shore. Aquatic and deep water emergent communities may also have been present around core C from this time, and were colonising the deeper parts of the basin by 8,600–8,300 cal bp (94.6% probability) (7,640 ± 83 bp. OxA-4042) (Day 1996, p 17; Dark 1998b, p 170). On dry ground, the pollen evidence indicates that Quercus became more common, and, along with Ulmus and Corylus, formed a significant component of the woodland whilst Alnus spread to the area (Day 1996, p 17; Dark 1998b, p 170), and probably colonised drier areas within the wetland.
From ca 8,000 cal bp the hydrosere entered its final phase as the water around core C became increasingly shallow and then terrestrialised at ca 7,750 cal bp. Standing water may have persisted in some places, and may have resulted in small pools within a wider mosaic of swamp, fen and carr. The deposits around trench F dried out for a second time at ca 7,500 cal bp, though the effects appear to have been limited to the edge of the basin. Shortly after organic sedimentation resumed Alnus colonised the site (ca 7,250 cal bp) and probably the area around core A. Fen environments persisted around cores B and C, though the occurrence of Alnus and Betula macrofossils suggests at least some tree cover locally.
Implications for Mesolithic activity
The first Mesolithic groups arrived in the area at ca 11,300 cal bp (Milner et al. 2018), with evidence for activity at multiple locations around the lake within the following centuries (Conneller et al. 2016; Taylor 2018). This phase of settlement is strongly associated with the lake environments and the exploitation of wetland resources. Activity areas were located on or close to the shore and have yielded evidence for fishing, fowling, hunting of aquatic mammals, and the collection and processing of wetland plants (Clark 1954; Taylor 2011, 2012, 2018; Taylor et al. 2018a; Robson et al. 2018).
Initial changes to the lake environments had little effect on this pattern of activity with sites continuing to be occupied as the lake edge became shallower and ultimately terrestrialised, and emergent vegetation extended further from the shore (see Taylor 2011, 2012, 2018; Milner et al. 2018). Indeed, the early stages of wetland succession were probably beneficial to early Mesolithic groups as the expansion of swamp and carr within the shallower embayments created a mosaic of habitats suitable for fish, waterfowl, and some of the larger mammals.
This pattern of settlement began to change from the latter stages of the early Mesolithic as sites on low-lying ground close to the shore were abandoned. The recent excavations at Star Carr, which focused on the area between 24 m and 25 m a.s.l., show occupation ending ca 10,500 cal bp (Milner et al. 2018; Blockley et al. 2018), whilst activity at Seamer Carr site C (which lay at a similar elevation) had ceased by ca 10,000 cal bp (Conneller et al. 2016) (site locations in Fig. 2). In addition, there is no evidence for late Mesolithic settlement on early Mesolithic occupation sites at VP-D, VP-E, Seamer Carr site D, and Flixton Island (Table 1 in Conneller and Schadla-Hall 2003), all of which lie on or below 25 m a.s.l. (Fig. 2), and only minimal activity at Seamer Carr site K, which lies at a similar elevation. In contrast sites on higher ground, such as Flixton SHF, Flixton School Field, Barry’s Island and No Name Hill continued to be occupied into the late Mesolithic along with new sites (such as Rabbit Hill) on similarly elevated positions (Fig. 2).
The changes described above were probably a response to the expansion of peat-forming environments over areas of dry ground, a process that would have rendered low-lying areas adjacent to the shore unsuitable for settlement. In turn, the shift to higher ground changed the environmental context of the later sites, which now lay either at the junction between fen and the terrestrial woodland or, in the case of the former islands or peninsulas, on small wooded hills surrounded by wetlands. Indeed, from ca 9,000 cal bp it is more appropriate to describe these sites as occupying a wetland edge, rather than a lake-edge environment, as areas of open water receded into the basin.
As well as the shift in settlement, patterns of economic activity would have changed as both terrestrial woodland and wetland succession altered the availability of plant resources and the habitats of prey animals. However, these changes need not have been as unfavourable to the human population as has been suggested (contra Mellars 1998, p 230). Though the lake was infilling with fen, this would have been bordered by an extensive woodland-edge environment made up of younger saplings and shade intolerant species such as Salix and Populus tremula. This would have resulted in habitats suitable for browsers such as red and roe deer. Given that the development of closed canopy woodland is generally thought to have reduced the size of large mammal populations (e.g. Mellars 1976), the persistence of large areas of fen and woodland-edge resulted in the Lake Flixton landscape remaining an important area for both settlement and hunting throughout the much of the Mesolithic.
Conclusions
Plant macrofossil analysis and radiocarbon dating have established the timing and nature of wetland succession in Lake Flixton. Fluctuations in lake-level had a significant effect on the local environment, although there is no evidence for a sustained rise in lake-level. The early development of the wetlands had little effect upon the pattern of human activity and may even have been beneficial for the early Mesolithic populations by creating a diverse range of highly productive environments. From the later part of the early Mesolithic (ca 10,500 cal bp), environmental change had affected patterns of settlement and economic activity, though the area remained inhabited. This project demonstrates the importance of plant macrofossil studies for tracking changes in lakes and wetlands associated with human activity and settlement, and the need for well-dated palaeo-environmental records at a local scale when interpreting the effects of early Holocene environmental change on patterns of human activity.
References
Birks HH (1973) Modern macrofossil assemblages in lake sediments in Minnesota. In: Birks HJB, West RG (eds) Quaternary plant ecology. Blackwell, Oxford, pp 173–189
Blockley S, Candy I, Matthews I et al (2018) The resilience of postglacial hunter-gatherers to abrupt climate change. Nat Ecol Evol 2:810–818
Bronk Ramsey C (2009a) Bayesian analysis of radiocarbon dates. Radiocarbon 51:337–360
Bronk Ramsey C (2009b) Dealing with outliers and offsets in radiocarbon dating. Radiocarbon 5:1,023–1,045
Bronk Ramsey C (2017) Methods for summarizing radiocarbon datasets. Radiocarbon 59:1,809–1,833
Cappers RTJ, Bekker RM, Jans JEA (2006) Digitale zadenatlas van Nederland (Digital seed atlas of the Netherlands). Groningen Archaeological Studies 4. Barkhuis, Eelde
Clark JGD (1954) Excavations at Star Carr. Cambridge University Press, Cambridge
Cloutman E (1988a) Palaeoenvironments in the Vale of Pickering, part 1: stratigraphy and palaeogeography of Seamer Carr, Star Carr and Flixton Carr. Proc Prehist Soc 54:52–69
Cloutman E (1988b) Palaeoenvironments in the Vale of Pickering, part 2: environmental history at Seamer Carr. Proc Prehist Soc 54:21–36
Conneller C, Schadla-Hall RT (2003) Beyond Star Carr: the Vale of Pickering in the tenth millennium bp. Proc Prehist Soc 69:85–105
Conneller C, Milner N, Taylor B, Taylor M (2012) Substantial settlement in the European early Mesolithic: new research at Star Carr. Antiquity 86:1,004–1,020
Conneller C, Bayliss A, Milner N, Taylor B (2016) The resettlement of the British landscape: towards a chronology of Early Mesolithic lithic assemblage types. Internet Archaeol. https://doi.org/10.11141/ia.42.12
Cummins G (2003) Impacts of hunter-gatherers on the vegetation history of the Eastern Vale of Pickering, Yorkshire. PhD thesis, Durham University
Dark P (1998a) Lake edge sequences: results. In: Mellars P, Dark P (eds) Star Carr in context: new archaeological and palaeoecological investigations at the early Mesolithic site of Star Carr, North Yorkshire. McDonald Institute for Archaeological Research, Cambridge, pp 125–146
Dark P (1998b) The lake centre sequence: results. In: Mellars P, Dark P (eds) Star Carr in context: new archaeological and palaeoecological investigations at the early Mesolithic site of Star Carr, North Yorkshire. McDonald Institute for Archaeological Research, Cambridge, pp 163–178
Day P (1996) Devensian late-glacial and early Flandrian environmental history of the Vale of Pickering, Yorkshire, England. J Quat Sci 11:9–24
Grosse-Brauckmann G (1972) Über pflanzliche Makrofossilien mitteleuropäischer Torfe. I: Gewebereste krautiger Pflanzen und ihre Merkmale. Telma 2:19–55
Grosse-Brauckmann G (1974) Über pflanzliche Makrofossilien mitteleuropäischer Torfe. II: Weitere Reste (Früchte und Samen, Moose u.a.) und ihre Bestimmungsmöglichkeiten. Telma 4:51–117
Grosse-Brauckmann G (1992) Über pflanzliche Makrofossilien mitteleuropäischer Torfe. III: Früchte, Samen und einige Gewebe (Fotos von fossilen Pflanzenresten). Telma 22:53–102
Haslam SM (1972) Biological flora of the British Isles, Phragmites communis. Trin J Ecol 60:585–610
Juggins S (2010) C2 1.66 Software for ecological and palaeoecological data analysis and visualization. https://www.staff.ncl.ac.uk/stephen.juggins/software/C2Home.htm. Accessed 12 Feb 2016
Koff T, Vandel E (2008) Spatial distribution of macrofossil assemblages in surface sediments of two small lakes in Estonia. Estonian J Ecol 57:5–20
Mellars P (1976) Fire ecology, animal populations and man: a study of some ecological relationships in prehistory. Proc Prehist Soc 42:15–45
Mellars P (1998) Postscript: major issues in the interpretation of Star Carr. In: Mellars P, Dark P (eds) Star Carr in context: new archaeological and palaeoecological investigations at the early Mesolithic site of Star Carr, North Yorkshire. McDonald Institute for Archaeological Research, Cambridge, pp 215–242
Mellars P, Dark P (1998) Star Carr in context: new archaeological and palaeoecological investigations at the early Mesolithic site of Star Carr, North Yorkshire. McDonald Institute for Archaeological Research, Cambridge
Milner N, Taylor B, Conneller C, Bayliss A (2018) Interpretative narrative of the history of occupation. In: Milner N, Conneller C, Taylor B (eds) Star Carr, vol 1: a persistent place in a changing world. White Rose University Press, York, pp 225–244
Reimer PJ, Bard E, Bayliss A et al (2013) IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal bp. Radiocarbon 55:1,869–1,887
Robson HK, Little A, Jones AKG et al (2018) Scales of analysis: evidence of fish and fish processing at Star Carr. J Archaeol Sci Rep 17:895–903
Rydin H, Jeglum J (2006) The biology of peatlands. Oxford University Press, Oxford
Schweingruber FH (1990) Microscopic wood anatomy. Structural variability of stems and twigs in recent and subfossil woods from Central Europe, 3rd edn. Swiss Federal Institute for Forest, Snow and Landscape Research, Birmensdorf
Stace C (2010) New flora of the British Isles, 3rd edn. Cambridge University Press, Cambridge
Taylor B (2011) Early Mesolithic activity in the wetlands of the Lake Flixton Basin. J Wetl Archaeol 11:63–84
Taylor B (2012) The occupation of wetland landscapes during the British Mesolithic: case studies from the Vale of Pickering. Ph.D. thesis, University of Manchester
Taylor B (2018) Subsistence, environment and Mesolithic landscape archaeology. Camb Archaeol J 28:493–510
Taylor B, Alison E (2018) Palaeoenvironmental investigations. In: Milner N, Conneller C, Taylor B (eds) Star Carr, vol 2: studies in technology, subsistence and environment. White Rose University Press, York, pp 123–149
Taylor B, Gray Jones A (2009) Definitely a pit, possibly a house? Recent excavations at Flixton School House Farm in the Vale of Pickering. Mesolith Misc 20:21–26
Taylor B, Conneller C, Milner N, Elliott B, Little A, Knight B, Bamforth M (2018a) Human lifeways. In: Milner N, Conneller C, Taylor B (eds) Star Carr, vol 1: a persistent place in a changing world. White Rose University Press, York, pp 245–272
Taylor B, Blockley S, Candy I et al (2018b) Climate, environment and Lake Flixton. In: Milner N, Conneller C, Taylor B (eds) Star Carr, vol 1: a persistent place in a changing world. White Rose University Press, York, pp 41–54
Walker D, Godwin H (1954) Lake stratigraphy, pollen-analysis and vegetational history. In: Clark JGD (ed) Excavations at Star Carr: an early Mesolithic site at Seamer near Scarborough, Yorkshire. Cambridge University Press, Cambridge, pp 25–69
Zhao Y, Sayer CD, Birks HH, Hughes M, Peglar SM (2006) Spatial representation of aquatic vegetation by macrofossils and pollen in a small and shallow lake. J Palaeolimnol 35:335–350
Acknowledgements
The plant macrofossil analysis for trenches F, H and I, and core A were carried out as part of the author’s AHRC-funded doctoral research (AHRC Doctoral Award 2007/135399). The radiocarbon chronologies for those profiles were supported by a Grant from NERC/ORAU (NF/2009/1/20), and the fieldwork at Flixton was supported by funding from by the Vale of Pickering Research Trust. My thanks to Amy Gray Jones who co-directed the excavations, Nick Overton and Pete Ryan for assistance with the coring, and Jeff Blackford, Mel Giles and Jamie Woodward for advice and support throughout my doctoral thesis. I also thank the two anonymous referees and Michael O’Connell for their comments on the earlier drafts of this paper. All errors remain my own.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M. O’Connell.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Taylor, B. Early Holocene wetland succession at Lake Flixton (UK) and its implications for Mesolithic settlement. Veget Hist Archaeobot 28, 559–573 (2019). https://doi.org/10.1007/s00334-019-00714-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-019-00714-9