
Abstract
Whale carcasses on the seafloor support unique, ephemeral communities of organisms, and ‘natural’ whale fall sites are infrequently encountered, especially in polar regions. During a manned submersible dive in early 2017, we discovered the skeleton of an Antarctic minke whale (Balaenoptera bonaerensis) at 963 m in Palmer Deep, in the Western Antarctic Peninsula. The site was filmed in HD for approximately two hours, enabling visual identification of representatives from at least eight phyla, although physical sampling was not possible. The remains appeared to be in the late ‘enrichment–opportunistic’ phase (although some mobile scavengers were still present and some sulfonic activity had already commenced), with polychaetes of the order Aciculata, and family Ampharetidae, plus several amphipod species, most abundant. Novel eusirid amphipod and rhodaliid siphonophore taxa were also present. The observed faunal distribution suggests patterns consistent with reports from other Antarctic whale falls (both experimental and natural). This discovery represents the highest-latitude natural whale fall reported to date.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Whale falls represent large organic deposits on the sea floor that may quickly attract a diverse assemblage of opportunistic and specialized organisms. These sites have received increasing attention since the first direct observations were made 30 years ago (Smith et al. 1989), and the diverse reports of whale fall assemblages accumulated in the interim have recently been summarized and reviewed (Smith et al. 2015). Decomposing whale remains progress through a series of four recognizable stages of varying duration and dominance as noted by, e.g., Smith and Baco (2003), Smith et al. (2015), and Silva et al. (2021): 1, ‘mobile-scavenger’ stage (four months to two years post-fall, characterized by necrophagous scavengers removing soft tissue from carcass); 2, ‘enrichment–opportunistic’ stage (months to years in duration, marked by dense colonization of polychaetes and crustaceans utilizing organically enriched sediments and exposed bone); 3, ‘sulfonic’ stage (lasting decades, dominated by an assemblage of sulfur-loving organisms attracted to the sulfide released during anaerobic decomposition of bone and remaining tissue lipids); 4, ‘reef stage’ (nutrient-depleted bones act as a hard substrate for settlement by opportunistic suspension feeders). In whale falls, the availability of food governs ecological succession of the communities such that as one community type is phasing out due to lack of utilizable resources, another community type is transitioning in (Connell and Slatyer 1977). These stages support characteristic communities of organisms (Smith and Baco 2003; Smith et al. 2015), some of which appear endemic to whale falls such as some of the ‘bone-eating’ polychaete worms (genus Osedax; e.g., Rouse et al. 2004, Vrijenhoek et al. 2009), the gastropod genus Rubyspira (Johnson et al. 2010), and numerous polychaetes in the family Dorvilleidae (Smith et al. 2015). Smith et al. (2019) suggested that the first human-driven extinctions in the deep sea may, in fact, have been whale-fall-endemic species, as a result of greatly reduced natural whale fall events throughout the world’s oceans, following 150 years of intensive commercial whaling. Notably, the decline in size of individual whales (and thus the biomass supporting individual whale fall events), through the targeted hunting of larger species, may have played an equal—or greater—role in whale fall specialist species extinctions as the reduction in population size, particularly in the Southern Ocean (Smith et al. 2019).
While the earliest whale fall studies resulted from opportunistic encounters of naturally occurring remains (e.g., Smith et al. 1989; Goffredi et al. 2004), a number of experiments with artificial whale falls have also been undertaken (e.g., Smith and Baco 2003; Dahlgren et al. 2006; Fujiwara et al. 2007; Lundsten et al. 2010). These sites may enable longitudinal studies of whale fall community succession, under more controlled circumstances, including placement of remains, known age of deposition, and the possibility of directly comparing multiple sites (including planned sampling strategies) while reducing potential confounding factors. Such studies have provided considerable insight into whale fall assemblages (in a wide range of geographic locations), including the apparent ubiquity of the genus Osedax, albeit with a high number of possibly geographically restricted species, many undescribed (see Glover et al. 2013). These experiments also suggest that, apart from the whale fall specialists, the majority of animal species in these communities tend to be background deep-sea taxa (Bennett et al. 1994; Smith and Baco 2003). Natural whale falls are, by contrast, generally encountered unexpectedly and without knowledge of the carcass’ origin (circumstances and date of death), and thus may provide only a snapshot glimpse of the assemblage, with data possibly generated solely through physical samples or visual observations. Yet natural whale falls provide intriguing glimpses into these ephemeral oases of productivity and often yield novel insights, such as the possibility of an early skin-sloughing successional stage prior to the mobile-scavenger stage (Smith et al. 2014a).
Modern whale fall observations have been most concentrated in the north Pacific, along the coasts of California and Japan; additional sites are known from (or have been placed) off the coasts of Europe, South America and southern Africa (see Fig. 1 in Smith et al. 2015 for review). High-latitude whale falls comprise a relatively small proportion of these (< 5 observations each above 55°N or below 55°S), and in the Antarctic region, most studies have been made within the Western Antarctic Peninsula (WAP) region. Several of these have been experimental deployments of individual bones (e.g., Glover et al. 2013; Taboada et al. 2013; Amon et al. in prep.; Glover et al. in prep.); a very recently deceased individual was also partially observed by Smith et al. (2014b), and a baleen whale skeleton in advanced stages of decomposition was reported by Stauffer et al. (2022). Observations of implanted whale falls in the WAP reveal the presence here, as elsewhere, of Osedax as a prompt and dense colonizer of whalebone (at the enrichment–opportunist stage and later), alongside diverse polychaetes such as the dorvilleid Ophryotrocha (Glover et al. 2013; Taboada et al. 2013). Reports on natural whale fall assemblages have included, in early decomposition (mobile-scavenger) stages, fishes, crabs, amphipods, echinoderms, nemerteans, and gastropods (Smith et al. 2014a); and in late decomposition stages, the additional presence of a highly diverse invertebrate fauna including sponges, anemones, amphipods, isopods, bivalves, pycnogonids, sabellid and eunicid polychaetes, and sipunculids (Amon et al. 2013). Most WAP sites have comprised the remains of Antarctic minke whales (Balaenoptera bonaerensis), a species whose remains have also been studied elsewhere (e.g., Alfaro-Lucas et al. 2017, 2018).
During a media expedition in January 2017, a nearly complete B. bonaerensis skeleton was discovered while exploring the northeastern slope of Palmer Deep, off Anvers Island. Recognizing the rarity and extreme serendipity of finding natural whale fall sites, extensive video footage was collected; due to permit restrictions, collection of physical specimens was regrettably not possible. These observations enabled subsequent study of the whale fall assemblage, providing unique insight into the decomposition of large organic deposits on the Antarctic seafloor and raising some intriguing questions.
Materials and methods
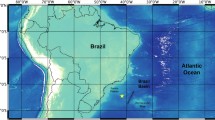
Observations were made opportunistically during a single manned submersible dive from MV Alucia in January 2017. Whale remains were discovered at 963 m depth in a northeastern finger of Palmer Canyon, within Palmer Deep, Western Antarctic Peninsula (64°50.5ʹS 64°09.2ʹW; Fig. 1). The skeleton was identified as that of an Antarctic minke whale (Balaenoptera bonaerensis) based on morphometry of the skull in particular (sharp, abbreviated rostrum, ruling out sei, fin and blue whale, and too short for humpback; N. Pyenson pers. comm.). Observations were made over two hours, with HD footage captured using a Canon EOS C500, Sony α 7 s, and Canon CN7 × 17 17–120 mm; Black Magic Micro Studio 4 K w/ Olympus Zuiko ED 12–50 mm EZ; and multiple white LED lights (6500 K). We subsequently reviewed accumulated footage in detail (KB, MO, CDB) in order to visually identify assembled organisms, using pertinent literature (e.g. Rauschert and Arntz 2015) and authors’ personal databases. Only taxa that could be confidently identified as present and independently confirmed among observers are reported. Due to filming angle, slope aspect, and fine sediment grain sizes (easily disturbed/suspended), the remains were observed from downslope only and attempts to quantify absolute abundance or density of organisms were not made.

Location of a natural whale fall (Antarctic minke whale, Balaenoptera bonaerensis) observed at 963 m depth in Palmer Deep, Western Antarctic Peninsula
Results
The skeleton rested in a slight gully at 963 m in Palmer Deep, bottom temperature 1.4 °C, and was mostly intact (Figs. 2, 3; Online Resource 1), with only small amounts of connective tissue remaining (Online Resource 2). The caudal vertebrae were located a short distance up a steep slope (30–40°) (Fig. 4; Online Resource 3) suggesting that the bulk of the carcass had slid downslope over time. Most thoracic and lumbar vertebrae rested upright on the seafloor, remaining articulated with intervertebral discs still intact. The few disarticulated vertebrae had sunk into the fine, silty sediment surrounding the carcass. One mandible was visible and a length of intact baleen plates was evident (Fig. 2); numerous shorter sections were scattered downslope of the skull. Skull length was estimated at 2 m.

Remains of an Antarctic minke whale (Balaenoptera bonaerensis) and associated biota, observed at 963 m in Palmer Deep, Western Antarctic Peninsula. 2, Entire skeleton, largely intact, with baleen plates downslope (lower left) and caudal vertebrae upslope (upper right). 3, Anterior view. 4, Enlargement of disarticulated caudal vertebrae. a, Anemone (Actinaria indet.); b, Brachioteuthid squid Slosarczykovia circumantarctica; d, Drift algae (Cystosphaera jacquinotii and Himantothallus grandifolius); m, ‘Sea pig’ Protelpidia murrayi; n, Barracudina Notolepis coatesi; r, Rhodaliid siphonophore (novel taxon); s, Salpa thompsoni; z, Deep-sea eelpout (Zoarcidae indet.)
At least 26 distinct taxa from at least eight phyla were filmed on/around the bones and surrounding enriched sediments (Table 1). Of these, ten were directly associated with the skeleton. The most abundant visible taxa were polychaetes: a tube-forming ampharetid polychaete (which also formed dense tube fields extending several meters out from the skeleton across the surrounding sediment), a free-crawling/swimming taxon (Figs. 5–8, Online Resources 2, 4, 5; likely a hesionid or the chrysopetalid Boudemos sp.), and a mostly stationary taxon observed raising its head, likely Ophryotrocha sp. (T. Dahlgren, A. Glover, and H. Wiklund, pers. comms). Mucous deposits within the ampharetid tube fields may have indicated the presence of Osedax, but no individuals could be discerned. Numerous yellow lysianassid or uristid amphipods were closely aggregated in compact patches, primarily on the spinal processes of lumbar vertebrae (Fig. 6); a distinctly larger, yellowish-orange amphipod species with a red head and large red eyes (probably an undescribed eusirid or pontogeneiid) was also present in smaller numbers (Figs. 5–8, Online Resources 2, 4). Additional less-abundant taxa included munnopsid isopods and another species of eusirid or pontogeneiid amphipod (whitish, with small rounded red eyes). Numerous amphipods and several ostracods were also observed swimming around the skeleton. The highest densities of organisms on the bones were observed at the vertebral junctions, in the hollows of the skull, and on the dorsal projections/ spinal processes of the lumbar vertebrae (Figs. 6, 7).

Remains of an Antarctic minke whale (Balaenoptera bonaerensis) and associated biota, observed at 963 m in Palmer Deep, Western Antarctic Peninsula. 5, Posterior skull region and ribs. 6, vertebrae. 7, Anterior view of rostrum. 8, Enlargement intervertebral region with dense assemblage of polychaete worms. d, Drift algae (Himantothallus grandifolius); e, Eusirid or pontogeneiid amphipod (novel taxon); l, Lysianassid or uristid amphipods; p, Hesionid or chrysopetalid polychaete worms; t, Ampharetid polychaete worms; r, Rhodaliid siphonophore (novel taxon); o, Ophyrotrocha sp.; z, Deep-sea eelpout (Zoarcidae indet.)
A further 16 + taxa were present but not directly associated with the skeleton, with the barracudina fish Notolepis coatesi and the ubiquitous Southern Ocean tunicate Salpa thompsoni most abundant. The salps occurred in both chained formations and as free-swimming single individuals, and numerous dead individuals were visible among the ampharetid fields. Zoarcid fishes were also commonly observed (Figs. 2, 4, 5, Online Resources 1–5, along with several pieces of presumptive brown drift algae, greenish in its decayed state (Figs. 3, 6, 8), the elpidiid holothurians Protelpidia murrayi (Figs. 3, 4) and cf. Peniagone vignoni, and several large anemones (Fig. 4, Online Resource 3). Notable singleton taxa included a rhodaliid siphonophore (Fig. 3, Online Resource 5; new species and genus awaiting description; D. Lindsay, pers. comm.), a snailfish (Liparidae: Careproctus sp.) and the Antarctic brachioteuthid squid Slosarczykovia circumantarctica (Online Resource 1).
The remains appeared to be in the late enrichment–opportunist stage, with a considerable annelid and crustacean assemblage; mobile scavengers such as crabs and macrourids, present in the vicinity, were not observed in direct association with the skeleton. Localised sediment darkening suggested some sulfonic activity.
Discussion
These fortuitous observations represent the highest-latitude natural whale fall data reported to date. Permit restrictions prevented us from collecting and examining physical specimens in order to confirm visual IDs (which no doubt would have revealed the presence of many additional taxa); however, close video observations still represent a valuable opportunity to document an in-situ Antarctic whale fall assemblage.
One notable absence in the aggregated footage is any obvious sign of the ‘bone-eating’ polychaete Osedax, a commonly reported member of whale fall assemblages throughout the deep sea (e.g., Braby et al. 2007; Fujiwara et al. 2007; Silva et al. 2021), including nearby locations in the Western Antarctic Peninsula (Glover et al. 2013). When present, these worms are often highly evident on the ribs (J. Alfaro-Lucas, pers. comm.) but were not seen in any location on the present remains. This is somewhat surprising given how rapidly Osedax may colonize whale fall sites (within as little as two weeks to two months; e.g., Onishi et al. 2018; Silva et al. 2021), and the densities of one species, O. antarcticus, observed on B. bonaerensis bones deployed nearby for 14 months (up to 202 individuals/m2; Glover et al. 2013). If present, O. antarcticus should have been readily visible, given their emergent palp length of up to 25 mm and known willingness to colonise vertebrae of this whale species, as observed by Glover et al. (2013). The same study did also identify a much smaller novel species, O. deceptionensis, whose presence would likely be far less obvious. We suspect, but can only indirectly infer (in the absence of direct observations and physical specimens), the unobserved presence of Osedax, possibly corroborated by mucous deposits observed within the surrounding ampharetid tube fields (T. Dahlgren and A. Glover, pers. comm.). Gastropod species established on other whale falls such as the specialist Rubyspira (Johnson et al. 2010) and limpets observed on whale skulls in the Ross Sea (B. Marshall, pers. comm.) were also not observed at the present site; these may similarly have been present but undetected.
Among the visibly present taxa, several novel observations were made, particularly among the amphipods. Eusirid/pontogeneiid amphipods have not been previously reported on whale falls, but in many cases amphipod taxa are not identified precisely, since accurate identifications are often difficult (particularly when based on imagery). Eusirid amphipods are mostly predators (Bousfield and Hendrycks 1995) and, if not attracted by the remains themselves, could well be drawn to the site by abundant prey. The presence of large aggregations of lysianassoid amphipods (comprising typical scavengers) at this late enrichment–opportunist stage has also not been explicitly reported before, but is not unexpected given the presence of some soft tissue. Faunal representatives of the various whale fall stages may overlap as nutrients released during one stage decline and those chararcterizing the next stage become available (e.g., Smith and Baco 2003; Goffredi et al. 2004; Braby et al. 2007;Smith et al. 2015). Given the aforementioned difficulty of identifying amphipods, it is difficult to attribute representative amphipod species to particular decomposition stages and to assess their succession in time. Amphipods of the families Lysianassidae and Uristidae (as well as Alicellidae and Eurytheneidae) are common and abundant mobile scavenger species at shelf and deep-sea depths (see De Broyer et al. 2004, 2007, for the Southern Ocean records). On whale falls, during the scavenger stage, amphipods have in fact often been recorded among the most rapidly attracted scavengers (Jones et al. 1998; Smith and Baco 2003). They are noted to occur in ‘thick swarms’ or in ‘dense populations’ one to four months after experimental deployments, along with hagfish and sleeper sharks (Dahlgren et al. 2006; Lundsten et al. 2010). Amphipod presence has also been recorded during the sulfonic stage by Bennett et al. (1994), Amon et al. (2014) and Alfaro-Lucas et al. (2017). The latter reported the occurrence of the uristid amphipod Stephonyx sp. (a scavenger) on vertebrae preserved from the degradation by Osedax during a ‘highly sulfophilic stage’, with chemosynthetic bacterial mats and numerous epifaunal organisms feeding on them.
Beyond the eusirid/pontogeneiid amphipods, several further potentially novel observations were made, but in the absence of physical samples, remain less concrete than we might wish. Ampharetid worms have been reported in similarly high abundances from whale falls in the northeast Pacific (Smith et al. 2014a), but have not previously been reported from Antarctic whale falls, so there is little regional basis for comparison and not much to be said without the opportunity for detailed morphological examinations. The most abundant polychaete observed crawling over much of the skeleton resembles Ophryotrocha (which has been reported from an experimental whale fall at nearby Deception Island; see Taboada et al. 2013), and a second common species appears to be a hesionid or possibly Boudemos sp. (T. Dahlgren, A. Glover, and H. Wiklund, pers. comms). Neither of these has previously been reported in the region, although they are very common on whale falls in other ocean basins, such as the north Pacific (Smith et al. 2014a, 2015). However, given the substantial number of polychaete species reported on other whale falls where direct specimen observations were possible (e.g., 28 + polychaete species from 13 + families from a single incomplete B. bonaerensis skeleton in the Atlantic; Sumida et al. 2016), it is likely the that diversity of the whale fall fauna at this site is substantially underestimated.
The drift algal pieces represent imported sources of carbon and nitrogen and these observations contribute to the growing evidence of exported nutrient-rich material (blue carbon) to the deep-sea floor (Barnes et al. 2018). Multiple fragments of at least two taxa were present; greenish sheet-like pieces (see Figs. 6, 8) were most likely dead thalli sections of the brown alga Himantothallus grandifolius. The presence of unidentifiable worms on the H. grandifolius decaying fragment (indicated ‘d’ in Fig. 6, 8) suggests that such material is indeed a nutrient source. Other brown algal fragments were clearly Cystosphaera jacquinotii, with characteristic floats (deflated) evident on drift pieces observed nearby (Fig. 4).
Cause and approximate date of death for this Antarctic minke whale remain unknown; the estimated skull length of ~ 2 m suggests a mature individual but no further inferences can confidently be drawn from our observations. The relatively intact state of the bones, high degree of articulation, presence of some soft connective tissue, and enrichment–opportunist assemblage (see Smith and Baco 2003; Smith et al. 2015) suggest a relatively recent death, perhaps as short as one or two years’ residence time on the sea floor (T. Dahlgren and A. Glover, pers. comm.). Based on decomposition and attending fauna, these remains are certainly at an earlier stage than those reported in the same general region by Amon et al. (2013) and Stauffer et al. (2022), perhaps similar in age or slightly younger than those observed by Smith et al. (2014b), particularly given the similarities in attending crustaceans and zoarcids.
The surrounding sediment tended toward the olive-green colouration associated with worm bioturbation observed by Treude et al. (2009), but darkened patches upslope of the carcass likely indicated sulfide presence from anaerobic decomposition and the onset of the sulfonic stage (Smith et al. 2015). We suspect that sediment sampling within the area would have revealed considerable biogeochemical gradients, as well as additional animal taxa and probably substantial numbers of fungi (see Nagano et al. 2020), nitrogen- and sulfur-fixing microbes (e.g., Cavalett et al. 2017), and methanogenic archaea (Goffredi et al. 2008). While some observations on Antarctic whale fall microbial communities report relatively lower diversity compared with lower-latitude sites (Tringe et al. 2005), biotechnological promise has been shown by proteins produced by whale fall bacteria (Smith and Baco 2003); Cardoso de Freitas et al. (2019, p. 26) concluded that the potential “discovery of unique extremophile proteins in deep-sea whale fall metagenomes justifies the allocation of resources for their exploration.”
Conclusion
As with any opportunistically encountered natural whale fall, these observations provide an intriguing snapshot of the successional processes occurring on a food-bonanza site in the deep-sea. Some peculiarities were observed compared with other whale fall reports, such as the possible absence of Osedax and Rubyspira. Due to relative proximity to several existing research stations, we strongly recommend revisiting the site to conduct repeat observations, and augment the present data with physical samples to enable further biological and chemical characterization of this ephemeral polar community.
Data availability
Beyond Online Resources 1–5, the raw footage that supports the findings of this study are available from NHK Enterprises, Ltd, Japan, but restrictions apply. These data were used under license for the current study, and so are not publicly available, but may be provided by the authors (KM, HI) upon reasonable request and with permission of NHK Enterprises, Ltd, Japan.
Change history
09 March 2023
Missing Open Access funding information has been added in the Funding Note.
References
Alfaro-Lucas JM, Shimabukuro M, Ferreira GD, Kitazato H, Fujiwara Y, Sumida PY (2017) Bone-eating Osedax worms (Annelida: Siboglinidae) regulate biodiversity of deep-sea whale-fall communities. Deep Sea Res II 146:4–12. https://doi.org/10.1016/j.dsr2.2017.04.011
Alfaro-Lucas JM, Shimabukuro M, Ogata IV, Fujiwara Y, Sumida PY (2018) Trophic structure and chemosynthesis contributions to heterotrophic fauna inhabiting an abyssal whale carcass. Mar Ecol Prog Ser 596:1–12. https://doi.org/10.3354/meps12617
Amon DJ, Glover AG, Wiklund H, Marsh L, Linse K, Rogers AD, Copley JT (2013) The discovery of a natural whale fall in the Antarctic deep sea. Deep Sea Res II 92:87–96. https://doi.org/10.1016/j.dsr2.2013.01.028
Amon DJ, Wiklund H, Dahlgren TG, Copley JT, Smith CR, Jamieson AJ, Glover AG (2014) Molecular taxonomy of Osedax (Annelida: Siboglinidae) in the Southern Ocean. Zoolog Scr 43:405–417. https://doi.org/10.1111/zsc.12057
Barnes DK, Fleming A, Sands CJ, Quartino ML, Deregibus D (2018) Icebergs, sea ice, blue carbon and Antarctic climate feedbacks. Philos Trans Royal Soc A 376:20170176
Bennett BA, Smith CR, Glaser B, Maybaum HL (1994) Faunal community structure of a chemoautotrophic assemblage on whale bones in the deep northeast Pacific Ocean. Mar Ecol Prog Ser 108:205–205
Bousfield EL, Hendrycks EA (1995) The amphipod superfamily Eusiroidea in the North Pacific region. I. Family Eusiridae: Systematics and distributional ecology. Amphipacifica 1:3–59
Braby CE, Rouse GW, Johnson SB, Jones WJ, Vrijenhoek RC (2007) Bathymetric and temporal variation among Osedax boneworms and associated megafauna on whale-falls in Monterey Bay, California. Deep Sea Res I 54:1773–1791. https://doi.org/10.1016/j.dsr.2007.05.014
Cardoso de Freitas R, Marques HIF, da Silva MAC, Cavalett A, Odisi EJ, da Silva BL, Montemor JE, Toyofuku T, Kato C, Fujikura K, Kitazato H, de Lima AOS (2019) Evidence of selective pressure in whale fall microbiome proteins and its potential application to industry. Mar Genom 45:21–27. https://doi.org/10.1016/j.margen.2018.11.004
Cavalett A, da Silva MAC, Toyofuku T, Mendes R, Taketani RG, Pedrini J, De Freitas RC, Sumida PYG, Yamanaka T, Nagano Y, Pellezari VH, Perez JAA, Kitazato H, de Lima AO, S, (2017) Dominance of Epsilonproteobacteria associated with a whale fall at a 4204 m depth–South Atlantic Ocean. Deep Sea Res II 146:53–58. https://doi.org/10.1016/j.dsr2.2017.10.012
Connell JH, Slatyer RO (1977) Mechanisms of succession in natural communities and their role in community stability and organization. Am Natur 111:1119–1144
Dahlgren TG, Wiklund H, Kallstrom B, Lundalv T, Smith CR, Glover AG (2006) A shallow water whale fall experiment in the north Atlantic. Cah Biol Mar 47(4):385–389
De Broyer C, Nyssen F, Dauby P (2004) The crustacean scavenger guild in Antarctic shelf, bathyal and abyssal communities. Deep Sea Res II 51:1733–1752. https://doi.org/10.1016/j.dsr2.2004.06.032
De Broyer C, Lowry JK, Jazdzewski K, Robert H (2007) Catalogue of the Gammaridean and Corophiidean Amphipoda (Crustacea) of the Southern Ocean with distribution and ecological data. Bull Inst R Sci Nat Belg Biol 77(suppl. 1):1–325
Fujiwara Y, Kawato M, Yamamoto T, Yamanaka T, Sato-Okoshi W, Noda C, Tsuchida S, Komai T, Cubelio SS, Sasaki T, Jacobsen K, Kubokawa K, Fujikura K, Maruyama T, Furushima Y, Okoshi K, Miyake H, Miyazake M, Nogi Y, Yatabe A, Okutani T (2007) Three-year investigations into sperm whale-fall ecosystems in Japan. Mar Ecol 28(1):219–232. https://doi.org/10.1111/j.1439-0485.2007.00150.x
Glover AG, Wiklund H, Taboada S, Avila C, Cristobo J, Smith CR, Kemp KM, Jamieson AJ, Dahlgren TG (2013) Bone-eating worms from the Antarctic: the contrasting fate of whale and wood remains on the Southern Ocean seafloor. Pro Soc Res B 280:20131390. https://doi.org/10.1098/rspb.2013.1390
Goffredi SK, Paull CK, Fulton-Bennett K, Hurtado LA, Vrijenhoek RC (2004) Unusual benthic fauna associated with a whale fall in Monterey Canyon, California. Deep Sea Res I 51:1295–1306. https://doi.org/10.1016/j.dsr.2004.05.009
Goffredi SK, Wilpiszeski R, Lee R, Orphan VJ (2008) Temporal evolution of methane cycling and phylogenetic diversity of archaea in sediments from a deep-sea whale-fall in Monterey Canyon, California. ISME J 2:204–220. https://doi.org/10.1038/ismej.2007.103
Johnson SB, Warén A, Lee RW, Kano Y, Kaim A, Davis A, Strong EE, Vrijenhoek RC (2010) Rubyspira, new genus and two new species of bone-eating deep-sea snails with ancient habits. Biol Bull 219:166–177. https://doi.org/10.1086/BBLv219n2p166
Jones EG, Collins MA, Bagley PM, Addison S, Priede IG (1998) The fate of cetacean carcasses in the deep sea: observations on consumption rates and succession of scavenging species in the abyssal north-east Atlantic Ocean. Proc Roy Soc B 265:1119–1127. https://doi.org/10.1098/rspb.1998.0407
Lundsten L, Schlining KL, Frasier K, Johnson SB, Kuhnz LA, Harvey JB, Clague G, Vrijenhoek RC (2010) Time-series analysis of six whale-fall communities in Monterey Canyon, California, USA. Deep Sea Res I 57:1573–1584. https://doi.org/10.1016/j.dsr.2010.09.003
Nagano Y, Miura T, Tsubouchi T, Lima AO, Kawato M, Fujiwara Y, Fujikura K (2020) Cryptic fungal diversity revealed in deep-sea sediments associated with whale-fall chemosynthetic ecosystems. Mycology 11:263–278. https://doi.org/10.1080/21501203.2020.1799879
Onishi Y, Shimamura S, Yamanaka T, Nakayama R, Ozaki KI, Miyazaki M, Tsunogai U, Fujiwara Y (2018) Variation of geochemical environments associated with whale-fall biomass mineralization processes in the sediment during the mobile scavenger, enrichment opportunist, and sulfophilic stages. Mar Biol 165:141. https://doi.org/10.1007/s00227-018-3398-8
Rauschert M, Arntz WE (2015) Antarctic Macrobenthos. A field guide of the invertebrates living on the Antarctic seafloor. Arntz & Rauschert Selbstverlag, Wurster Nordseeküste, Germany, pp 1–143
Rouse GW, Goffredi SK, Vrijenhoek RC (2004) Osedax: bone-eating marine worms with dwarf males. Science 305:668–671. https://doi.org/10.1126/science.1098650
Silva AP, Colaço A, Ravara A, Jakobsen J, Jakobsen K, Cuvelier D (2021) The first whale fall on the Mid-Atlantic Ridge: Monitoring a year of succession. Deep Sea Res Pt I 178:103662. https://doi.org/10.1016/j.dsr.2021.103662
Smith CR, Baco AR (2003) Ecology of whale falls at the deep-sea floor. Oceanogr Mar Biol 41:311–354
Smith CR, Kukert H, Wheatcroft RA, Jumars PA, Deming JW (1989) Vent fauna on whale remains. Nature 341:27–28
Smith CR, Bernardino AF, Baco A, Hannides A, Altamira I (2014a) The seven-year enrichment: Macrofaunal succession in deep-sea sediments around a 30-ton whale fall in the Northeast Pacific. Mar Ecol Prog Ser 515:133–149
Smith KE, Thatje S, Singh H, Amsler MO, Vos SC, McClintock JB (2014b) Discovery of a recent, natural whale fall on the continental slope off Anvers Island, western Antarctic Peninsula. Deep Sea Res I 90:76–80. https://doi.org/10.1016/j.dsr.2014.04.013
Smith CR, Glover AG, Treude T, Higgs ND, Amon DJ (2015) Whale-fall ecosystems: recent insights into ecology, paleoecology, and evolution. Annu Rev Mar Sci 7:571–596. https://doi.org/10.1146/annurev-marine-010213-135144
Smith CR, Roman J, Nation JP (2019) A metapopulation model for whale-fall specialists: the largest whales are essential to prevent species extinctions. J Mar Res 77(Supp. 2):1–20
Stauffer JB, Purser A, Griffiths HJ, Smith CR, Hoving HJT (2022) Food falls in the deep northwestern Weddell Sea. Front Mar Sci 9:1055318. https://doi.org/10.3389/fmars.2022.1055318
Sumida PY, Alfaro-Lucas JM, Shimabukuro M, Kitazato H, Perez JA, Soares-Gomes A (2016) Deep-sea whale fall fauna from the Atlantic resembles that of the Pacific Ocean. Sci Rep 6:1–9. https://doi.org/10.1038/srep22139
Taboada S, Wiklund H, Glover AG, Dahlgren TG, Cristobo J, Avila C (2013) Two new Antarctic Ophryotrocha (Annelida: Dorvilleidae) described from shallow-water whale bones. Polar Biol 36:1031–1045. https://doi.org/10.1007/s00300-013-1326-4
Treude T, Smith CR, Wenzhöfer F, Carney E, Bernardino AF, Hannides AK, Krüger M, Boetius A (2009) Biogeochemistry of a deep-sea whale fall: sulfate reduction, sulfide efflux and methanogenesis. Mar Ecol Prog Ser 382:1–21. https://doi.org/10.3354/meps07972
Tringe SG, Von Mering C, Kobayashi A, Salamov AA, Chen K, Chang HW, Podar M, Short JM, Mathur EJ, Detter JC, Bork P, Hugenholtz P, Rubin EM (2005) Comparative metagenomics of microbial communities. Science 308:554–557. https://doi.org/10.1126/science.1107851
Vrijenhoek RC, Johnson SB, Rouse GW (2009) A remarkable diversity of bone-eating worms (Osedax; Siboglinidae; Annelida). BMC Biol 7:74. https://doi.org/10.1186/1741-7007-7-7
Acknowledgements
We are forever grateful to NHK Enterprises, Ltd (Japan) for the opportunity to participate in this expedition; in particular, we thank M. Komoda, H. Iwasaki, and G. Hamazaki, as well as cameraman J. Miyake and submersible pilot L. Frey. This unexpected find generated considerable excitement among the science and media crew, and led to many intriguing and fruitful discussions with international colleagues within the deep-sea research community, notably at the 15th Deep-Sea Biology Symposium in Monterey, CA (September 2018), where a preliminary version of these findings was presented as a poster. In particular we thank A. Glover, D. Amon, D. Lindsay, J. Alfaro-Lucas, K. Osborn, and T. Dahlgren. Further identification assistance and valuable discussions were provided by H. Wiklund, N.D. Pyenson, C.R. Smith, B. Marshall, and S. Mills. We also thank C.R. Smith and an anonymous reviewer for their feedback, which has greatly strengthened this manuscript.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. NHK Enterprises,Ltd, Tokyo, Japan.
Author information
Authors and Affiliations
Contributions
MK and HI organised the expedition and procured funding. KB conducted in-situ observations. KB, MA and CDB analysed footage and identified organisms. KB and MA wrote the manuscript. KB prepared the figures and videos. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary file1 (MP4 135291 kb)—Anterior view of Balaenoptera bonaerensis skeleton (963 m depth, Palmer Deep, Western Antarctic Peninsula)
Supplementary file2 (MP4 329904 kb)—Lateral view of Balaenoptera bonaerensis vertebrae and associated biota (963 m depth, Palmer Deep, Western Antarctic Peninsula)
Supplementary file3 (MP4 489067 kb)—Disarticulated caudal vertebrae and associated biota, upslope of main skeleton of Balaenoptera bonaerensis (963 m depth, Palmer Deep, Western Antarctic Peninsula)
Supplementary file4 (MP4 380233 kb)—Anterior view of Balaenoptera bonaerensis skull and associated biota (foreground), 963 m, Palmer Deep, Western Antarctic Peninsula
Supplementary file5 (MP4 254405 kb)—Novel rhodaliid taxon attached to rib of Balaenoptera bonaerensis (963 m depth, Palmer Deep, Western Antarctic Peninsula)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bolstad, K.S.R., Amsler, M.O., Broyer, C.D. et al. In-situ observations of an intact natural whale fall in Palmer deep, Western Antarctic Peninsula. Polar Biol 46, 123–132 (2023). https://doi.org/10.1007/s00300-022-03109-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-022-03109-1