
Abstract
In recent years, functional changes in Southern Ocean are becoming more noticeable, due to climate change and increasing human impacts, including a growing fishery that is concentrating in the Antarctic Peninsula (AP) region. Antarctic krill Euphausia superba is often the primary prey species for animals such as Pygoscelis penguins, a sentinel species for ecosystem monitoring and management. During the last two decades in the AP gentoo penguin numbers (Pygoscelis papua) have increased and their range has shifted southward, in contrast to the decline in numbers of Adélie (P. adeliae) and chinstrap (P. antarcticus) penguins. Given divergent population trends, the goal of this study was to examine differences in their diet, and size structure of Antarctic krill recovered from penguin diet samples. The study is based on diet samples collected during the austral summers on King George Island (South Shetland Islands) where P. adeliae, P. antarcticus, and P. papua breed in mixed colonies. Results indicate that the penguins consumed krill of similar sizes during the breeding period. In contrast to prior diet studies, we found higher proportions of krill in the gentoo diet and changes in the percentage of krill in the diet relative obtained during 1970s. The similarity in diets among all three species suggests that the availability prey items (e. g., fishes) may be changing and driving higher dietary overlap. Moreover, we also check differences in krill length among penguin individuals and we did not find any statistically significant differences. We also found plastic debris in penguin stomachs during both summers.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The marine ecosystem in the Antarctic Peninsula region is strongly dependent on the availability of a small, swarming crustacean, Euphausia superba, (hereafter krill), which is the food base for numerous whale, seal and seabird species (Hill et al. 2006). For some of the predators, such as Pygoscelid penguins, krill can constitute up to 99% of their diet (Tierney et al. 2008; Juáres et al. 2018; Panasiuk et al. 2020). Krill biomass is estimated at around 380 million tons (Atkinson et al. 2009) and its harvesting is regulated by the Commission for the Conservation of Antarctic Marine Living Resources (CCAMLR). Krill are predicted to be affected by harvesting and the effects of climate change, including warming sea-surface temperatures, melting of sea ice, and changes in phytoplankton composition and abundance (Hill et al. 2006). In particular, the effects of climate warming on krill growth may have significant implications for krill biomass which can cause negative consequence for dependent predators (Klein et al. 2018).
Among the important krill-dependent predators are penguins, which constitute nearly 90% of Antarctic avian biomass (Croxall et al. 2002). As the main component of Antarctic avifauna, Pygoscelid penguins exhibit extensive overlap in their distributions and nesting sites (Volkman et al. 1980; Lishman 1985). They exhibit different foraging locations and foraging-depth ranges during the breeding season, but also during winter period (Hinke et al. 2019; Korczak-Abshire et al. 2021), which helps maintain their co-occurrence throughout the AP region. Despite such differences in niche, the similarity of diet among the Pygoscelid penguins suggests a shared constraint on their performance (Hinke et al. 2007). Their diet is heavily dependent on krill (70 to 100% of the diet) (Volkman et al. 1980; Hinke et al. 2007; Panasiuk et al. 2020), thus their foraging distribution and abundance trends are usually linked with krill biomass variability (Trivelpiece et al. 2011a, b; Stryker et al. 2020).
Relatively few research sites afford the opportunity to examine the diet of the three Pygoscelid penguin species simultaneously. Long-term research on breeding populations of Adélie (Pygoscelis adeliae), gentoo (P. papua), and chinstrap (P. antarcticus) penguins in Admiralty Bay, King George Island included study of their trophic niches. Volkman et al. (1980) and Trivelpiece et al. (1987) demonstrated that gentoo diets contained more fish than Adélie or chinstrap diets. More recently, Miller et al. (2010) highlighted local differences in diets, reporting that from 1997–1998 to 2003–2004 fish constituted 28.8% of a diet of gentoo and 0.6% of chinstrap at Cape Shirreff (Livingston Island), while in Admiralty Bay (King George Island) fish represented 1.6% of a diet of gentoo and 0.1% of chinstrap. Within the last three decades in Admiralty Bay, the breeding populations of Adélie and chinstrap penguins have decreased, while the population of gentoo penguins has increased (Watters et al. 2020), consistent with regional trends (Lynch et al. 2012). Such large-scale changes in the abundance and distribution of each species may affect their competitive relationships for food resources.
The main goal of our study was to analyze the diet composition and krill sizes consumed by sympatric Adélie, gentoo and chinstrap, and to compare our results with historical diet data. As a null hypothesis, we expected similar diet compositions of Adélie and chinstraps, but a higher proportion of fish in gentoo diets. We also expected similar distributions of the sizes, sex, and maturity stages of krill in the diets of three sympatric penguin species that initiate foraging trips from Admiralty Bay, leading to more similar diet characteristics that previously observed. We used two consecutive years of diet data to address our hypotheses related to dietary composition of each species.
Materials and methods
Sampling collection and laboratory studies
Diet samples were collected in the Austral summers (December–February) of 2011/2012 and 2012/2013. Fieldwork was conducted at King George Island, South Shetland Islands (62° 10’ S, 58° 30’W), within the Antarctic Specially Protected Area (ASPA) No. 128 where Adélie, chinstrap, and gentoo penguins breed in five colonies (Fig. 1). Following procedures of the Conservation of Antarctic Marine Living Resources (CCAMLR) Ecosystem Monitoring Program (CEMP), the population size, expressed in the total number of occupied nests, and breeding success, expressed as the mean number of chicks alive during the crèche stage per nest were calculated. Diet samples were collected from adult penguins after their foraging trips using the water-offloading technique (Wilson 1984) with CEMP standard methods modifications (see more in Panasiuk et al. 2020). Diet samples were sorted to identify the frequency of occurrence by number and weight of krill, fish, and other, unidentifiable diet items. All krill individuals collected in the diet samples were counted (based on the pairs of eyes for digested animals). Whole, undamaged individuals were sexed, and measured for total length, according to CCAMLR standard protocol (https://www.ccamlr.org/): from the anterior side of the eyeball to the tip of the telson (Makarov and Denys 1980). Diet samples from 2011/12 were used to estimate the difference between krill sizes consumed by penguins and to compare with krill sizes presented by Panasiuk et al. (2020). Diet samples from both seasons were used for comparing krill sizes and maturity stages. We only used diet samples that contained at least 30 whole individuals for comparing krill sizes and maturity stages. Sexed animals were grouped into the following clusters: F—females, Fs—sexually active females defined by presence of spermatophores, and M—males. Additional groups—Es and J—were used for individuals without clear sex indication and individuals with damaged bodies (Es), and juvenile individuals.

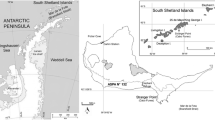
Breeding colony distribution of Adélie penguins (P. adeliae), chinstrap penguins (Pygoscelis antarcticus) and gentoo penguins (Pygoscelis papua) in 2012/13, within the Antarctic Specially Protected Area No. 128 (ASPA 128) at the western shore of Admiralty Bay at King George Island, South Shetland Islands, (contour of the Antarctic continent coast source: http://www.marineregions.org/gazetteer.php?p=details&id=1926, contour of the King George Island coast source: Scientific Committee on Antarctic Research Antarctic Digital Database, version 6.0, 1993–2015)
Statistical analyses
Statistical analyses were carried out using Rstudio software package, Rversion 4.0.3. We used a one-way ANOVA with a Tukey’s HSD post hoc test to compare differences in krill length and maturity stages in the penguin diet samples. Additionally, we used a nested ANOVA to compare differences inn krill lengths in which krill length was a response variable and years and penguin species were factors with year nested within species. Differences in krill sizes between years were statistically significant (p < 0.0001), however, because our main goal was to check differences between penguin species and inter-individual differences within species we pooled samples to examine sex and maturity stages of the krill population eaten by penguins. We also checked interindividual differences in krill lengths using the Kruskal–Wallis test.
Results
Krill, by mass and number, constituted more than 99% of the diet in all samples (Table 1), with only small contributions of other prey, including amphipods, and other euphausiids (Thysanoessa macrura, and Euphausia frigida) and fish. In case of frequency of occurrence—tiny fish remains or otoliths were reported in 70% of gentoo penguin stomachs in season 2012/2013 and 20% in 2011/2012. For Adélie and chinstrap, however, fish prey was present in first researched season in only 10% of individuals.
Most krill individuals in the diet samples were classified as adults with a median length of 40 mm (mean 39.76 ± 6.93). Juvenile krill were rare (Fig. 2, Table 2) and were most abundant in the Adélie diet. On average, the largest krill were observed in the diet samples of gentoo penguins, while diet samples from Adélie and chinstrap penguins contained smaller krill (Fig. 2, Table 1). The mean sizes of krill consumed by Adélie penguins was 38.40 ± 7.28 mm and chinstrap penguins was 40.68 ± 5.55 mm, while in gentoo diets was 41.7 ± 6.22 mm (Fig. 3, Table 1). The differences between the length of a krill in the diets were biggest between Adélie and chinstrap penguins and Adélie and gentoo penguins (Tukey HSD, p < 0.0001). Smaller difference was noted between gentoo and chinstrap penguins (Tukey HSD, p = 0.015). We also checked interindividual differences, and results of the Kruskal–Wallis test showed no statistically significant differences among Adélies (p = 0.4616), gentoos (p = 0.4478), and chinstraps (p = 0.4289).

Histograms of pooled krill E. superba lengths in stomach content of all three penguin species in 2011/12 and 2012/13

Differences in krill E. superba length in a diet of the three investigated penguin species between seasons 2011/2012 and 2012/2013
Female krill formed the majority of the diet for all penguin species (Table 2). The largest average size of krill in each sex and maturity stage was found in the gentoo diet (Table 2). The biggest krill was a 73 mm female with spermatophore (Fs), and this group represented, on average, the largest animals in all samples with mean lengths that varied from 44.43 to 46.45 mm (Table 2). Male krill individuals were the second largest group in terms of size (both by median and mean value) (Table 2).
We found algae, stones, feathers and other debris in the Pygoscelis penguin stomachs (Fig. 4). Some net fragments were identified. They were accompanied by plastic film elements of uncertain origin, although some of them looked like fragments of plastic bags. Most of the plastic debris could be classified as microplastics (1–5 mm; 10 plastic parts), mesoplastics (< 5–20 mm; 23 plastic parts) (Barnes et al. 2009) and classified as user plastic (only non-industrial remains of plastic objects) (Van Franeker and Meijboom 2006). In case of color of found debris—there was 1 occurrence of red-pink plastic parts (4 debris in one dietary sample in Adélie stomach) and 7 green strings (5 in Adélie penguin stomach content; 2 in gentoo), while majority (12 debris) were off/white-clear in color. Plastic debris were found in 16 stomach contents (out of 78) which constituted 20% of all samples.

Example of ingested plastic debris found in penguin diets in the summer season 2011–2012
Discussion
Krill dominance and size in Pygoscelis diet
Krill constituted almost 100% of the penguin diets, in weight and by number, during the 2011/2012 summer season, similar to other reports (Miller et al. 2010; Panasiuk et al. 2020). Fishes, in all cases, constituted < 0.1% of mass in the diet (Table 1). The similarities in diet composition among sympatric Pygoscelid penguins observed during our study differ from those observed roughly for decades ago. For example, Trivelpiece et al. (1987) noted that krill constituted 86.5% of in the gentoo penguin diet during the period 1976–1985. Similar proportions were also noted earlier by Volkman et al. (1980), with the krill comprising 84.5% of the gentoo diet and 99.6% for the two other penguin species (Table 3). Jabłoński (1985) recorded even 48.6% of fish in the gentoo diet (Table 3). We explore possible reasons for the increased similarity in diet below.
Adélie and chinstrap penguins in the broader Antarctic Peninsula region typically exhibit a summer diet-based almost exclusively of krill (Lishman 1985). Such high amounts of krill in the diet of all three species suggest sufficient local availability of krill in the environment during the summer breeding period (Hinke et al. 2007; Nardelli et al. 2021). The apparent increase in the occurrence of krill in the diet of gentoo penguins since the early 1980s may imply a decline in availability of secondary prey items, particularly fish. While we noted that 70% of gentoos consumed fish in 2012/2013 season which would corresponds with Miller et al. (2010) findings in Cape Shirreff (Livingston Island), most of noted presence in our study was based on otoliths presence in stomach content. Gentoo penguins were known to consume Pleurogramma antarctica in King George Island vicinity (Volkman et al. 1980). A study of the distribution of Pleurogramma on the Western Antarctic Peninsula shelf exposed a gap in its occurrence which could indicate a local collapse of this fish (no individuals smaller than 150 mm were recorded, and after 1990s smaller contribution in predators diet has been noted) (Parker et al. 2015). Evidence of a decade’s long decline in the occurrence of fish in Antarctic fur seal diets (Klemmendson et al. 2020) provides supporting evidence for potential changes in the preyscape available to penguins in this region. The relative dominance of krill in the diet is also supported by recent results from a molecular study of penguin feces (Zeng et al. 2022) that suggests a diet dominated by Euphausiid species. Therefore, further study is necessary to explain the switch to a diet with higher proportions of krill, especially with increasing population of gentoo penguins in the region, and the concurrent decrease in Adélie and chinstrap penguin populations numbers (Lynch et al. 2012). We note that the stomach contents can be related to sample timing relative to the period of a day or night foraging. Jansen et al. (1998) and Miller and Trivelpiece (2008) observed during their studies that the diet of chinstrap penguins consists of more fish during nights, and samples for this study were collected only in the afternoons.
In our study, the mean length of krill in all penguin stomach contents was 38–42 mm and adult individuals were the majority (Figs. 2, 3). These sizes are comparable to the findings of Volkman et al. (1980) and Coria et al. (1995), but smaller than the animals noted by Trivelpiece et al. (2011a) from the same area of research. Differences in the length of krill consumed by penguins may be caused by differences in the availability of certain age classes, different foraging areas, or year-to-year differences in prey availability, recruitment, and abundance (Coria et al. 1995; Hewitt et al. 2003; Kokobun et al. 2015; Reiss 2016). It is worth noting that our study area is recognized as a region of occurrence of juvenile krill (Fevolden and George 1984) and cycles of krill recruitment are evident throughout the WAP (Miller and Trivelpiece 2007; Saba et al. 2014; Reiss 2016; Conroy et al. 2020). However, few krill less than 30 mm in length were observed in the diet samples, which suggests that presumably krill recruitment in the area was not high in the preceding year.
Our two-year study confirmed that the largest krill were consumed by gentoo penguins, while the smallest animals were typically found in Adélie diets (Figs. 2, 3; Table 1). In general however, the krill sizes in the diets of the three penguin species were similar. It is evident that strong dietary overlap exists between Pygoscelis penguins in the study area, and that dietary overlap may be much more pronounced than reported in the past (Volkman et al. 1980; Jabłoński 1985; Lishman 1985; Trivelpiece et al. 1987).
Krill sex and stages in Pygoscelis diet
During this study, a preference for female krill was evident across penguin species (Table 2). In over 3800 measured krill individuals, only 6.5% were recognized as males. Similar results were observed by Volkman et al. (1980), Reid et al. (1996), and Lynnes et al. (2004). In our study female krill with attached spermatophores constituted more than 20% of all measured females. The disproportionate occurrence of female krill in the diet of predators also was surprising for Reid et al. (1996) around the South Georgia, who surmised that predators opted to catch the largest available krill—gravid females. Sexually active females were the most numerous group in predator diet and net samples in researched done by Reid et al. (1996). Hill et al. (1996) suggested that for macaroni penguins, the predominance of female krill in the diet could be a result of capture avoidance by faster swimming male krill. The higher proportion of female krill in penguin diet may also point to selectivity for the largest individuals available. In the Bransfield Strait, juvenile krill generally form the majority of the population, while adult females represent a smaller fraction of population (Siegel 1983; Fevolden and George 1984). Therefore, the dominance of female krill in penguin stomachs affirms selectivity for the largest krill individuals available. The female krill carrying spermatophores were also the largest krill in the available diet samples with a mean length of 45 mm for all species (Table 2). The preponderance of females in the diet may also affected by digestion. Irvine (2002) suggested that smaller males are usually much more affected by the sampling methods and preservation than females, and tend to break. Those factors may lead to the underestimations of smaller males in samples and could explain the relatively small number of males < 40 mm present in the diet samples.
Krill availability in Pygoscelis diet and their breeding success
Many reports suggest a direct linkage between reproductive success of predators and fluctuations in krill abundance and structure (Lynnes et al. 2004; Trivelpiece et al. 2011b). In the 2011/2012 breeding season, the abundance of Adélie and chinstrap penguins was larger than during the 2012/13 breeding season (Table 1). The situation was in opposite for gentoo penguins. The breeding success of all three penguin species, however, was higher in 2011/2012 than in 2012/2013 (Table 1), similar to other breeding locations like the Lions Rump (Korczak-Abshire personal observations) (ASPA 151), and the Stranger Point (Juáres et al. 2020). This suggests that the 2011/12 season may have been better for the penguins. During our study we did not find any considerable differences in diet composition, krill size, or numerical of krill between both seasons. However, in a krill dominated system, we may not expect the diet composition of penguins to change considerably, but the performance of birds can be negatively affected if foraging trip durations are long (e.g., Hinke et al. 2007). Episodic phenomena, including heavy snowfall events (Spring 2012 and January 2013—personal observation), can also negatively affect reproductive performance (see Hinke et al. 2012). For example, the strong wind, rainfalls, and thick snow events affect the increase of chicks’ energy input and lower thermal insulation factors. Thus, the observed difference in breeding success between years may reflect meteorological anomalies during incubation and guard stages.
What is the reason for dietary overlap?
Historical data shows a distinction between chinstrap and gentoo trophic niche based on diet differences (Volkman et al. 1980) and foraging ranges (Trivelpiece et al. 1987). However, climate change in Antarctic Peninsula region (Meredith and King 2005) and shifts in penguin population trends may affect the trophic niche of the Pygoscelid penguins. Both Adélie and chinstrap breeding populations have decreased in the northern Antarctic Peninsula region (Korczak-Abshire et al. 2012; Juáres et al. 2015). For example, in Turret Point Oasis (King George Island) from 1980 to 2008 the population of Adélie decreased by 84.06% and chinstrap by 50.09% (Korczak-Abshire et al. 2019). In case of gentoo penguins at Stranger Point/Cabo Funes, South Shetland Islands Juáres et al. (2020) noted that their population between 2000/2001 and 2018/2019 increased by more than 60%. The first observations of this trend were made in 1990s by Ciaputa and Sierakowski (1999) and they suggested that penguin population trends and krill abundance were linked. Population changes among gentoo and chinstrap penguins are related to temperature and its anomalies (Petry et al. 2016), while the decline in Adélie penguin numbers is correlated with reduction of sea ice during winter season (Juáres et al. 2015). Taking into consideration increases in air temperature from 1981 to 2011 during summer, autumn and winter seasons, along with increases in sea-surface temperatures and increased variability in sea ice extent (Kejna et al. 2013), we should expect a response in birds population numbers. Yet, between the 2011/2012 and 2012/2013 summer seasons, we didn’t detect substantial differences in krill size composition in Pygoscelis stomachs. Similarity in diet composition across the penguin species can be connected with a lower availability of krill or other alternative prey items with an increasing competition between them. If the numbers of Adélie and chinstrap penguins are in decline, while gentoo populations rise, it could indicate that gentoo penguins are expanding their trophic niche to occupy the vacancy created by decreasing populations of its congeners, and this possibility was also discussed by Miller et al. (2010).
Plastic debris in penguin stomachs
One of the first observations of ingested plastic by marine vertebrates south of the Polar Front was in a snow petrel (Pagodroma nivea) in the 1980s (Van Franeker and Bell 1988). Similar findings for other Antarctic predators remain rare (Waller et al. 2017). We found plastic debris in the Pygoscelis penguins stomachs in two consecutive seasons. Among Antarctic penguins, evidence for plastic ingestion was noted from penguin scats (Bessa et al. 2019) in the regions of Bird Island (South Georgia) and Signy Island (South Orkney Islands), and in regurgitate (Panasiuk et al. 2020) collected from birds at King George Island (South Shetland Islands). Although the Southern Ocean has smaller concentration of micro- and macroplastic than other parts of the World Oceans (Suaria et al. 2020), observations of plastic debris in the penguin stomachs during two breeding seasons (now one decade ago) suggest that its occurrence in diets is not a spurious finding. We found that Pygoscelid penguins are also able to ingest larger pieces of plastic (Fig. 4) and this could indicate additional potential mechanisms to impact penguins.
While there are grounds for supposition, answer the question: are the diets of sympatric Pygoscelid penguins more similar than previously thought? definitely needs more data. It is possible that widely studied belief about the decline of E. superba (e.g., Atkinson et al. 2009) could be now facing its turning point with confirmed big aggregations of returning whales, as fin whales noted in 2018 by Herr et al. (2022). Such situation could suggest that if there is enough of krill to support dietary needs of some predators groups this could also be a reason for higher percentage of dietary overlap of pygoscelid penguins. However, to regard this as a trend—such observation as in presented study need to be continued, as well as studies on what drives these changes.
Data availability
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
References
Atkinson A, Siegel V, Pakhomov EA, Jessopp MJ, Loeb V (2009) A re-appraisal of the total biomass and annual production of Antarctic krill. Deep Sea Res Part I 56(5):727–740
Barnes DK, Galgani F, Thompson RC, Barlaz M (2009) Accumulation and fragmentation of plastic debris in global environments. Philos Trans R Soc B 364(1526):1985–1998
Bessa F, Ratcliffe N, Otero V, Sobral P, Marques JC, Waluda CM, Trathan PN, Xavier JC (2019) Microplastics in gentoo penguins from the Antarctic region. Sci Rep 9(1):1–7
Ciaputa P, Sierakowski K (1999) Long-term population changes of Adélie, chinstrap, and gentoo penguins in the regions of SSSI No. 8 and SSSI No. 34, King George Island, Antarctica. Pol Polar Res 20(4):355–365
Conroy JA, Reiss CS, Gleiber MR, Steinberg DK (2020) Linking Antarctic krill larval supply and recruitment along the Antarctic Peninsula. Integr Comp Biol 60(6):1386–1400
Coria NR, Spairani H, Vivequin S, Fontana R (1995) Diet of Adélie penguins Pygoscelis adeliae during the post-hatching period at Esperanza Bay, Antarctica, 1987/88. Polar Biol 15:415–418
Croxall JP, Trathan PN, Murphy EJ (2002) Environmental change and Antarctic seabird populations. Sci 297(5586):1510–1514
Fevolden SE, George RY (1984) Size frequency pattern of Euphausia superba in the Antarctic Peninsula waters in the austral summer of 1983. J Crustac Biol 4(5):107–122
Herr H, Viquerat S, Devas F, Lees A, Wells L, Gregory B, Giffords T, Beecham D, Meyer B (2022) Return of large fin whale feeding aggregations to historical whaling grounds in the Southern Ocean. Sci Rep 12(1):1–15
Hewitt RP, Demer DA, Emery JH (2003) An 8-year cycle in krill biomass density inferred from acoustic surveys conducted in the vicinity of the South Shetland Islands during the austral summers of 1991–1992 through 2001–2002. Aquat Living Resour 16(3):205–213
Hill HJ, Trathan PN, Croxall JP, Watkins JL (1996) A comparison of Antarctic krill Euphausia superba caught in nets and taken by macaroni penguins Eudyptes chrysolophus evidence for selection. Mar Ecol Prog Ser 140:1–11
Hill SL, Murphy EJ, Reid K, Trathan PN, Constable AJ (2006) Modelling Southern Ocean ecosystems: krill, the food-web, and the impacts of harvesting. Biol Rev Camb Philos Soc 81(4):581–608
Hinke JT, Salwicka K, Trivelpiece SG, Watters GM, Trivelpiece WZ (2007) Divergent responses of Pygoscelis penguins reveal a common environmental driver. Oecologia 153:845–855
Hinke JT, Polito MJ, Reiss CS, Trivelpiece SG, Trivelpiece WZ (2012) Flexible reproductive timing can buffer reproductive success of Pygoscelis spp. penguins in the Antarctic Peninsula region. Mar Ecol Prog Ser 454:91–104
Hinke JT, Santos MM, Korczak-Abshire M, Milinevsky G, Watters GM (2019) Individual variation in migratory movements of chinstrap penguins leads to widespread occupancy of ice-free winter habitats over the continental shelf and deep ocean basins of the Southern Ocean. PLoS ONE 14:1–19
Irvine LG (2002) Sex differences in Antarctic krill (Euphausia superba) retrieved from Adélie penguin stomachs: implications for diet analysis. Polar Biol 25:717–720
Jabłoński B (1985) The diet of penguins of King George Island, South Shetland Islands. Acta Zool Cracov 29:117–186
Jansen JK, Boveng PL, Bengtson JL (1998) Foraging modes of chinstrap penguins: contrasts between day and night. Mar Ecol Prog Ser 65:161–172
Juáres MA, Santos M, Negrete J, Mennucci JA, Perchivale PJ, Casaux R, Coria NR (2015) Adélie penguin population changes at stranger point: 19 years of monitoring. Antarct Sci 27(5):455–461
Juáres A, Casaux R, Corbala A, Blanco G, Pereira GA, Perchivale PJ, Coria NR, Santos MM (2018) Diet of Adélie penguins (Pygoscelis adeliae) at stranger point (25 de Mayo/King George Island, Antarctica) over a 13-year period (2003–2015). Polar Biol 41:303–311
Juáres MA, Casaux R, Negrete J, Rios A, Castillo M, Coria NR, Santos MM (2020) Update of the population size and breeding performance of gentoo penguins (Pygoscelis papua) at stranger Point/Cabo Funes, South Shetland Islands. Polar Biol 43(2):123–129
Kejna M, Araźny A, Sobota I (2013) Climatic change on King George Island in the years 1948–2011. Pol Polar Res 34(2):213–235
Klein ES, Hill SL, Hinke JT, Phillips T, Watters GM (2018) Impacts of rising sea temperature on krill increase risks for predators in the Scotia Sea. PLoS ONE. https://doi.org/10.1371/journal.pone.0191011
Klemmedson AD, Reiss CS, Goebel ME, Kaufmann RS, Dorval E, Linkowski TB, Borras-Chavez R (2020) Variability in age of a Southern Ocean myctophid (Gymnoscopelus nicholsi) derived from scat-recovered otoliths. Mar Ecol Prog Ser 633:55–69
Kokubun N, Choy E-J, Kim J-H, Takahashi A (2015) Isotopic values of Antarctic Krill in relation to foraging habitat of penguins. Ornithol Sci 14(1):13–20
Korczak-Abshire M, Chwedorzewska KJ, Wąsowicz P, Bednarek PT (2012) Genetic structure of declining chinstrap penguin (Pygoscelis antarcticus) populations from South Shetland Islands (Antarctica). Polar Biol 35(11):1681–1689
Korczak-Abshire M, Zmarz A, Rodzewicz M, Kycko M, Karsznia I, Chwedorzewska KJ (2019) Study of fauna population changes on Penguin Island and Turret Point Oasis (King George Island, Antarctica) using an unmanned aerial vehicle. Polar Biol 42:217–224
Korczak-Abshire M, Hinke JT, Milinevsky G, Juáres MA, Watters GM (2021) Coastal regions of the northern Antarctic Peninsula are key for gentoo populations. Biol Lett. https://doi.org/10.1098/rsbl.2020.0708
Lishman GS (1985) The food and feeding ecology of Adélie penguins (Pygoscelis adeliae) and Chinstrap penguins (P. antarctica) at Signy Island, South Orkney Islands. J Zool 205(2):245–263
Lynch HJ, Naveen R, Trathan PN, Fagan WF (2012) Spatially integrated assessment reveals widespread changes in penguin populations on the Antarctic Peninsula. Ecology 93(6):1367–1377
Lynnes AS, Reid K, Croxall JP (2004) Diet and reproductive success of Adélie and chinstrap penguins: linking response of predators to prey population dynamics. Polar Biol 27(9):544–554
Makarov RR, Denys CJ (1980) Stages of sexual maturity of Euphausia superba Dana. BIOMASS Handbook 11:1–13
Meredith MP, King JC (2005) Rapid climate change in the ocean west of the Antarctic Peninsula during the second half of the 20th century. Geophys Res Lett. https://doi.org/10.1029/2005GL024042
Miller AK, Trivelpiece WZ (2007) Cycles of Euphausia superba recruitment evident in the diet of Pygoscelid penguins and net trawls in the South Shetland Islands. Antarctica Polar Biol 30(12):1615–1623
Miller AK, Trivelpiece WZ (2008) Chinstrap penguins alter foraging and diving behavior in response to the size of their principle prey, Antarctic krill. Mar Biol 154:201–208
Miller AK, Kappes MA, Trivelpiece SG, Trivelpiece WZ (2010) Foraging-niche separation of breeding gentoo and chinstrap penguins, South Shetland Islands. Antarctica Condor 112(4):683–695
Nardelli SC, Cimino MA, Conroy JA, Fraser WR, Steinberg DK, Schofield O (2021) Krill availability in adjacent Adélie and gentoo penguin foraging regions near Palmer Station. Antarctica Limnol Oceanogr 66(6):2234–2250
Panasiuk A, Wawrzynek-Borejko J, Musiał A, Korczak-Abshire M (2020) Pygoscelis penguin diets on King George Island, South Shetland Islands, with a special focus on the krill Euphausia superba. Antarct Sci 32(1):21–28
Parker ML, Fraser WR, Ashford J, Patarnello T, Zane L, Torres JJ (2015) Assemblages of micronektonic fishes and invertebrates in a gradient of regional warming along the Western Antarctic Peninsula. J Mar Syst 152:18–41
Petry MV, Valls FCL, de Souza PE, Krüger L, da Cruz PR, dos Santos CR (2016) Breeding sites and population of seabirds on Admiralty Bay, King George Island. Antarctica Polar Biol 39(7):1343–1349
Reid K, Trathan PN, Croxall JP, Hill HJ (1996) Krill caught by predators and nets: differences between species and techniques. Mar Ecol Prog Ser 140(1/3):13–20
Reiss CS (2016) Age, growth, mortality, and recruitment of Antarctic Krill, Euphausia superba. In: Siegel V (ed) Biology and ecology of Antarctic Krill. Springer, Cham, pp 101–144
Saba GK, Fraser WR, Saba VS, Iannuzzi RA, Coleman KE, Doney SC, Schofield OM (2014) Winter and spring controls on the summer food web of the coastal West Antarctic Peninsula. Nat Commun 5(1):1–8
Siegel V (1983) Population structure of Euphausia superba in the eastern part of the Bransfield Strait. In Schnack SB (ed) On the biology of krill Euphausia superba. Proceedings of the Seminar and Report of the Krill Ecology Group, Bremerhaven, 12–16 May 1983, Berichte zur Polarforschung, Sonderheft 4, Alfred-Wegener-Institut Polarforschung, Bremerhaven, pp. 227–238
Strycker N, Wethington M, Borowicz A, Forrest S, Witharana C, Hart T, Lynch HJ (2020) A global population assessment of the Chinstrap penguin (Pygoscelis antarctica). Sci Rep. https://doi.org/10.1038/s41598-020-76479-3
Suaria G, Perold V, Lee JR, Lebouard F, Aliani S, Ryan PG (2020) Floating macro-and microplastics around the Southern Ocean: results from the Antarctic circumnavigation expedition. Environ Int. https://doi.org/10.1016/j.envint.2020.105494
Tierney M, Southwell C, Emmerson LM, Hindell MA (2008) Evaluating and using stable-isotope analysis to infer diet composition and foraging ecology of Adélie penguins Pygoscelis adeliae. Mar Ecol Prog Ser 355:297–307
Trivelpiece WZ, Trivelpiece SG, Volkman NJ (1987) Ecological segregation of Adélie, Gentoo, and Chinstrap Penguins at King George Island. Antarctica Ecol 68(2):351–361
Trivelpiece SG, Will A, Boysen K, Chilton P, Trivelpiece WZ (2011a) Seabird Research at Admiralty Bay, King George Island, Antarctica. In Walsh JG (ed) AMLR 2010–2011a FIELD SEASON REPORT. National Marine Fisheries Service Southwest Fisheries Science Center, La Jolla, USA, pp 24–27
Trivelpiece WZ, Hinke JT, Miller AK, Reiss CS, Trivelpiece SG, Watters GM (2011b) Variability in krill biomass links harvesting and climate warming to penguin population changes in Antarctica. PNAS 108(18):7625–7628
Van Franeker JA, Bell PJ (1988) Plastic ingestion by petrels breeding in Antarctica. Mar Pollut Bull 19(12):672–674
Van Franeker JA, Meijboom A (2006) Fulmar Litter EcoQO Monitoring in the Netherlands 1982–2004 in relation to EU Directive 2000/59/EC on Port Reception Facilities Report for the Ministry of Transport, Public works and Water Management (V&W), contract nr DGTL/ZH/2.53. 2.5012. Alterra, Texel Sampling of Fulmars in the Netherlands is conducted by the Dutch Seabird Group (Nederlandse Zeevogelgroep-NZG)
Volkman NJ, Presler P, Trivelpiece W (1980) Diets of Pygoscelid penguins at King George Island. Antarctica Condor 82(4):373–378
Waller CL, Griffiths HJ, Waluda CM, Thorpe SE, Loaiza I, Moreno B, Pacherres CO, Hughes KA (2017) Microplastics in the Antarctic marine system: an emerging area of research. Sci Total Environ 598:220–227
Watters GM, Hinke JT, Reiss CS (2020) Long-term observations from Antarctica demonstrate that mismatched scales of fisheries management and predator-prey interaction lead to erroneous conclusions about precaution. Sci Rep 10:2314. https://doi.org/10.1038/s41598-020-59223-9
Wilson RP (1984) An improved stomach pump for penguins and other seabirds. J Field Ornithol 55(1):109–112
Zeng YX, Li HR, Han W, Luo W (2022) Molecular dietary analysis of Adélie (Pygoscelis adeliae) and Gentoo (Pygoscelis papua) penguins breeding sympatrically on Antarctic Ardley Island using fecal DNA. Polar Biol 45:999–1011
Acknowledgements
The authors thank the WZ and SG Trivelpiece for providing Pygoscelis penguin diet samples and research data on the Llano Point breeding site. The data on penguin populations used in the paper were collected at the Henryk Arctowski Polish Antarctic Station. We would like to express our gratitude to the reviewers who improved significantly this manuscript, one anonymous reviewer and Mr. Scott Jennings.
Funding
This work was supported by the Polish National Science Centre, Project Miniatura 1 no. 2017/01/X/NZ8/01181, received by Anna Panasiuk.
Author information
Authors and Affiliations
Contributions
JW-B was responsible for the laboratory analysis, dataset preparation, results interpretation, conception, preparation and management of the manuscript. AP responsibility was the coordination of sample collection, statistical analysis with interpretation, and in manuscript preparation. MK-A provided data and results interpretation of penguin population. All authors provided critical feedback and helped shape the research, analysis and manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Ethical approval
All applicable institutional guidelines for the care and use of animals were followed.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wawrzynek-Borejko, J., Panasiuk, A., Hinke, J.T. et al. Are the diets of sympatric Pygoscelid penguins more similar than previously thought?. Polar Biol 45, 1559–1569 (2022). https://doi.org/10.1007/s00300-022-03090-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-022-03090-9