
Abstract
The main target of this paper is to improve the knowledge of the species composition of sea stars in Patagonian Argentine deep sea reaching depths of 2062 m. In addition, these results offer us the opportunity to analyze the possible connections between Argentinian marine fauna and adjacent Antarctic areas that have become a topic of interest in the past few years. This work is based on Atlantic Projects’ surveys carried out on an atypical and especially vulnerable marine ecosystems (canyons created from craters collapse by gas leaks). These are profusely impacted by frequent fishing activities, being one of the most important and international fishing grounds, where 887 records (1878 specimens) of 41 species of asteroids were collected in 217 stations ranging from 219 to 2062 m in depth. Seven of those species are proposed as new records: (Diplasterias octoradiata (Studer 1885), Plutonaster bifrons (Wyville Thomson, 1873), Radiaster elegans Perrier, 1881, Anseropoda antarctica Fisher, 1940, Pillsburiaster calvus Mah, 2011, Paralophaster lorioli (Koehler, 1907), Pteraster flabellifer Mortensen 1933). After refining the database built from literature and open-access databases such as OBIS and AntBIF, the new Argentinian asteroids deep-water checklist contains 2198 records from 64 asteroids species including the 7 new records proposed. Most of these 64 species (89.06%) are present in Antarctic-adjacent waters, and after the study of their occurrences at traditional biogeographic entities, our results support the hypothesis that Argentinian waters (in the case of the class Asteroidea) should be considered part of the sub-Antarctic entity.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The distribution of Argentinian marine fauna and their connection with adjacent areas has become a topic of interest in the last few years. Regarding the north, a good study of these connections can be found in Alvarado and Solís-Marín (2013) and, within this field, the study undertaken by Brogger et al. (2013a) in Argentinian waters. Regarding the south, some authors have tried to explain how species distribution areas can help to analyze the existence of biogeographical entities (regions or provinces) in the Southern Ocean, mainly based on their occurrence (Hedgpeth 1969; Rodriguez et al. 2007; Griffiths et al. 2009; De Broyer et al. 2014; Figuerola et al. 2017; Weir and Stanworth 2019, among others). In the case of echinoderms, the most recent studies have been carried out by Martin-Ledo and Lopez Gonzalez (2014), Danis et al. (2014), Eleaume et al. (2014), Saucède et al. (2014), and Moreau et al. (2017, 2018, 2019, 2021), among others.
The study area is located in the Southwest Atlantic. It extends over 200 miles from the exclusive economic zone of Argentinian waters and reaches a depth of 2062 m. This zone is known as the Patagonian fishing area, owing to the fishing activities undertaken here by many countries, among them Spain (del Rio et al. 2012). These fishing activities are related to the area’s high biodiversity, which is mainly due to the nutrient-rich upwelling during spring and summer as a consequence of the confluence of two ocean currents: the warm, subtropical Brazilian current (BC), which flows from the north, and the cold, deeper current from the Malvinas/Falkland Islands (M/FI) from the south. The latter could be considered a branch of the Antarctic Circumpolar Current (ACC) (Piola and Rivas 1997; Piola and Matano 2001; Rivas et al. 2006; Campagna et al. 2007). In addition, the bottom morphology shows a canyon system with very special features since it was created from the collapse of a series of pockmarks (craters produced by gas leaks).
In this context, the Spanish General Secretariat of Fisheries charged the Spanish Institute of Oceanography (Instituto Español de Oceanografía, IEO), with the undertaking of a series of multidisciplinary research surveys (focused on slope and canyon systems: Atlantis Project; del Rio et al. 2012) aimed at studying the potential existence of vulnerable marine ecosystems (VMEs), where echinoderms, among others, represent one of the most abundant and frequent taxa in the area.
Most studies on echinoderms from this area are restricted to the Argentinian shelf or near the slope, such as Bernasconi (1934, 1935, 1937, 1941a, 1941b, 1943, 1962a, 1962b, 1963, 1964a, 1964b, 1965, 1966a, 1972, 1973a, 1980), Tablado (1982), and Tablado and Maytia (1988), and more recent studies, such as Brogger et al. (2010, 2013b), Martinez and Brogger (2012), Martinez et al. (2015), Souto et al. (2014), Brogger and O’Hara (2015), Di Giorgio et al. (2015), Arribas et al. (2016), Epherra et al. (2015, 2017), Hunter et al. (2016); Martinez (2016), Martinez and Penchaszadeh, (2017), Martinez et al. (2018; 2020), Wilkie and Brogger (2018), Carames et al. (2019), Flores et al. (2019), Martins and Tavares (2019), and Gil et al. (2020). With regard to the class Asteroidea, few papers have been published recently: Romanelli and Tablado (2011), Pérez et al. (2017), Cossi et al. (2017), Arribas et al. (2017), Berecoechea et al. (2017), Rivadeneira et al. (2017), Fraysse et al. (2018), Rivadeneira et al. (2020) and Fraysse et al. (2020), and Moreau et al. (2018, 2021). However, some asteroids were found in deeper areas that were recorded on research cruises, such as the Challenger, Vema, and Walter Herwig, with results published by Sladen (1882, 1883) and Bernasconi (1965, 1966b, 1973b) respectively.
The objectives of the present work are to review the asteroid diversity in the study area considering: (1) the wide depth range covered by the Atlantis project compared with previous studies, (2) the potential record of new species occurrences, with better descriptions of species depth ranges, and (3) quantifying the number of common species shared with Antarctic waters describing potential connections between these two areas.
Material and methods
Field sampling
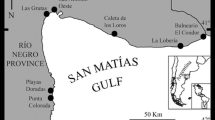
Sampling was done during Atlantis Projects’ surveys (13 multidisciplinary research expeditions) on board the Miguel Oliver vessel from 2007 to 2010 (Fig. 1). Different equipment was used: rock dredge (0.8 m wide and 0.3 m high; mesh size 10 mm), mega box corer dredge, and LOFOTEN trawl (31.20 m × 17.70 m; mesh size 35 mm).

Area of study. a General view with the sampled area labeled as b. b Sampled area; stations are represented by black dots
Fauna was collected at 480 stations; however, only 217 stations were taken into account (219 m to 2062 m depth) because the present study is restricted to the slope and canyons system (Table 1).
Identification and taxonomy
Asteroids were sorted and fixed in 70% ethanol. Identification was based on morphological characters according to: Clark and Downey (1992), Clark (1962), Bernasconi (1934, 1935, 1937, 1941a, 1943, 1957, 1961, 1962a, 1962b, 1963, 1964a, 1964b, 1965, 1966a, 1996b, 1972, 1973a, 1980), Tablado (1982), Tablado and Maytia (1988), McKnight (2006), Mah (2011, 2018), and the original descriptions. Asteroids’ classifications were checked in World Register of Marine Species (WoRMS: Mah 2022), and new record species AphiaID (urn:lsid:marinespecies.org:taxname) were included for consulting and referring to synonymies. Morphological notations follow Clark and Downey (1992).
Datasets
After species identification, the results were georeferenced and included in a dataset.
Several publicly available datasets were used for species distribution analysis: Ocean Biogeographic Information System (OBIS), Antarctic Biodiversity Information Facility (AntBIF), Museo Argentino de Ciencias Naturales (MACN), and online USNM Invertebrate Zoology Collection database. On the other hand, other records were include coming from literature such as Mah (2011), Mah and Blake (2012), Souto et al. (2014), Moreau et al. (2018), Guillaumot et al. (2020), and Moreau et al. (2021). These records were assessed to determine their reliability on the basis of two criteria: (1) records published in scientific journals and (2) records or human observations from international surveys or institutions and identified by experts of recognized standing. The reliable records were included together with Atlantic surveys ones that were curated following a polygon built (ArcGIS 10.7) delimiting the area of study to the appropriated countries borders and depth range (nodes: 37.69°S, 69.37°W; 37.69°S, 48.97°W; 55.78°S, 68.60°W; 55.67°S, 63.48°W; 49.81°S, 63.54°W; 49.63°S, 48.97°W).
Biogeographical approach
Five biogeographical entities were defined to analyze the Antarctic and near waters influence: Argentinian deep waters (ADW), Malvinas/Falkland Islands (M/FI), Scotia Arc (SA), sub-Antarctic Islands, including Bouvet Island, Prince Edward Islands, Crozet Islands and Kerguelen Islands (SAI), and Antarctica (A) following Griffiths et al. (2009), based on Moore et al. (1999).
Results
Studied area checklist and new records
A total of 1878 specimens, belonging to 41 asteroid species (Table 2), were found where Ctenodiscus australis were the most frequent and abundant (22.68%; 50.23%), followed by Cheiraster planeta (9.31%; 28.57%) and Henricia studeri (12.83%; 28.57%).
Twenty-four species expanded their bathymetric range (Table 3), and seven species are proposed as new records for Argentinian waters, which are summarized below (Fig. 2, 3; Online Resource 1).

Images to support diagnosis of: Diplasterias octoradiata: a abactinal view, bar: 2 cm; b detail of marginal species, bar: 0.01 cm; c adambulacral spines, bar: 0.01 cm. Plutonaster bifrons: d abactinal general view, bar: 1 cm; e actinal general view, bar: 0.02 cm; f marginal plates, bar: 0.02 cm; g adambulacral plates and furrow spines, bar: 0.20 cm. Radiaster elegans: h abactinal (right side) versus actinal (left side) general view, bar: 1 cm; i detail of actinal plate arrangement, bar: 0.20 cm; j adambulacral plates and furrow spines, bar: 0.20 cm; k detail of abactinal paxillar plates, bar: 0.20 cm; l superomarginal plates and spines, bar: 0.20 cm. Radiaster tizardi: m abactinal general view, bar: 1 cm; n marginal fringe, bar: 0.50 cm; o actinal general view (plate arrangement), bar: 0.50 cm

Images to support diagnosis of: Anseropoda antarctica: a abactinal general view, bar: 2 cm; b detail of abactinal plates, bar: 0.02 cm; c details of actinal plates, bar: 0.02 cm; d adambulacral plates and furrow spines, bar: 0.02 cm. Pillsburiaster calvus: e abactinal (left side) versus actinal (right side) general view, bar: 1 cm; f detail of abactinal plates, bar: 0.20 cm; g adambulacral plates and spines of the furrow, bar: 0.20 cm; h superomarginal plates, bar: 0.20 cm. Paralophaster lorioli: i abactinal general view, bar: 0.50 cm; j detail of abactinal plates, bar: 0.01 cm; k detail of superomarginal plates, bar: 0.01 cm; l detail of actinal plates, bar: 0.01 cm; m detail of oral plates, bar: 0.02 cm. Pteraster flabellifer: n abactinal general view, bar: 1 cm; o detail of oral plates, bar: 0.20 cm; p marginal plates, bar: 0.20 cm; q adambulacral plates, bar: 0.20 cm
Phylum Echinodermata Klein 1778
Class Asteroidea Blainville 1830
Order Forcipulatida Perrier 1884
Family Asteriidae Gray 1840
Genus Diplasterias Perrier 1891
Diplasterias octoradiata (Studer 1885), new record (Fig. 2a–c)
AphiaID: 172655
Diagnosis Arms 7–9, rarely 5, 6, 10. Abactinal surface rigid, skin pustular, skeleton continuous and delicate with spines numerous with a crossed pedicellariae wreath (Fig. 2a). Superomarginal spines with a wreath of crossed pedicellariae (Fig. 2b). Adambulacral plates without crossed pedicellariae, straight and predominantly monacanthid (Fig. 2c).
Distribution Malvinas/Falkland Islands, The Scotia Arc, Ross Sea, Wilkes Land. Present study: Argentine. New record: 2 specimens (Online Resource 1).
Bathymetric range 7–866 m (Ahearn 1995; USNM1087183 from Invertebrate Collection Database, NMNH). Present study: 849–855 m.
Remarks Diplasterias octoradiata and Diplasterias radiata (Koehler 1923) (7–10 arms) could be confused because of their arms number overlap; however, the number of adambulacral spines are different: D. radiata are diplacanthid, and D. octoradiata are monacanthid.
Order Paxillosida Perrier 1884
Family Astropectinidae Gray 1840
Genus Plutonaster Sladen 1889
Plutonaster bifrons (Wyville Thomson 1873), new record (Fig. 2d–g)
AphiaID: 123904
Diagnosis Quite long arms R/r = 3.4/1–5.0/1, with narrow tips, almost pointed. Abactinal surface with plates with rather thin paxillae, in transverse series (Fig. 2d). Abactinal paxillae with columns oval in cross-section, crowned with 15–25 short spines. Marginal plates, both superomarginal and inferomarginal with only one prominent conical spine, and otherwise covered with very minute spines (Fig. 2f). Actinal plates covered with minute spines or granules and often with a single conical spine (Fig. 2e). Adambulacral plates with 5–10 furrow spines of equal size, outside of which is a single large conical spine (Fig. 2g).
Distribution Mediterranean Sea, Northeast Atlantic Ocean (from Faroe Islands to Gulf of Guinea, including Canary Islands), Northwest Atlantic Ocean (from New Jersey (USA) to Venezuela). Present study: Argentine. New record: 9 specimens (Online Resource 1).
Bathymetric range 100–3587 m (Mortensen 1927; USNM E-31521 from Invertebrate Collection Database, NMNH). Present study 1037–1820 m.
Remarks Related to Plutonaster agassizi (Verrill 1880), P. bifrons shows a narrow and pointed arms with a terminal plate as long as wide, since P. agassizi ones are rounded, their tips are blunt, and their superomarginal plates curve inward. In contrast, the superomarginal armament of P. bifrons has spines on all plates and relatively long and pointed, P. agassizi does not present spines, at least from the distal side, and, if present, they are short, rigid, or conical.
Family Radiasteridae Fisher 1916
Genus Radiaster Perrier 1881
Radiaster elegans Perrier 1881, new record (Fig. 2h–l).
AphiaID: 152510
Diagnosis Arms long R/r = 3.5/1, with small madreporite (Fig. 2h). Marginal plates with double series of spines larger than abactinal and actinal plates (Fig. 2k, 2l). Transverse series of actinal plates (Fig. 2i) different from the adambulacral ones (Fig. 2j). Oral plates markedly enlarged so that each jaw has a double keel of numerous suboral spines.
Distribution Caribe Sea, Venezuela, Present study: Argentine. New record: 1 specimen (Online Resource 1).
Bathymetric range 604–1446 m (USNM E19305, Oregon II expedition 1970, USNM E 31524 from Invertebrate Collection Database, NMNH). Present study 1077 m.
Remarks There are five Radiaster species (including R. elegans): Radiaster elegans Perrier 1881 [Gulf of Mexico (USNM E 31,524) from Leeward I. to north of Guayana (Clark and Downey, 1992)]. It presents long arms and small madreporite; Radiaster gracilis (H.L. Clark, 1916) (New Zealand and Australia, Tasmania: Mah et al. 2009) presents more than 70 adambulacral plates broader than length; Radiaster notabilis (Fisher 1913) (Batjan Islands Molucca Islands: Fisher 1913) presents scarce paxillae with 75 spines, and 13–14 adambulacral spines; Radiaster rowei H.E.S Clark & D. G. McKnight 2000 (New Zealand: Clark and McKnight, 2000; Mah et al. 2009); Radiaster tizardi (Sladen 1882) (North Atlantic Ocean: from Ireland to off Sahara, Grand Bank: Terranova (Clark and Downey, 1992; Murillo et al. 2016) presents moderate arm length (R/r = 2.2/1–3.0/1) and a large madreporite (Fig. 2m–o), while R. elegans presents long arms and a small madreporite.
Order Valvatida Perrier 1884
Family Asterinidae Gray 1840
Genus Anseropoda Nardo 1834
Anseropoda antarctica Fisher 1940, new record (Fig. 3a–d).
AphiaID: 172707
Diagnosis Arms 5. The abactinal plates scalar, thin and imbricate, covered with granuliform spines and inconspicuous papular pores (Fig. 3a, b). Actinal and abactinal plates in decreasing size toward the marginal ones. Abactinal plates form a carinal series. Small superomarginal plates covered by granules. Small inferomarginal plates protrude to form the edge and are also covered by granules. Actinolateral plates in regular oblique series (Fig. 3c). Adambulacral plates with 6 spines on the furrow and joined by a membrane (Fig. 3d).
Distribution Tierra de Fuego, The Scotia Arc, South Shetland Islands. Present study: Argentine. New record: 29 specimens (Online Resource 1).
Bathymetric range 123–3510 m (USNM 1,122,403; USNM 1122101 from Invertebrate Collection Database, NMNH). Present study 111–1543 m.
Remarks Four species (A. antarctica included) could be compared taking into account their distribution area: Anseropoda rosacea (Lamarck 1816) presents 16 arms: Anseropoda macropora Fisher 1913 (shallow waters) presents 5 arms, 5 spines on the furrow, and Anseropoda aotearoa McKnight 1973 (New Zealand) presents 5 arms, 5–7 furrow spines, and only one series of subadambulacral spines, rarely two.
Family Goniasteridae Forbes 1841
Genus Pillsburiaster Halpern 1970
Pillsburiaster calvus Mah 2011, new record (Fig. 3e–h).
AphiaID: 559190
Diagnosis Abactinal plates covered by granules (Fig. 3e). Papulae present at radial region (Fig. 3f). Superomarginal plates convex and nude between 50% and 90% of surface (Fig. 3h). Inferomarginal plates convex with a low percentage of nude surface. Two to three spines in the adambulacral plate and 2 separated by a space on subadambulacral plates. Four to five oral spines (Fig. 3g).
Distribution The Scotia Arc. Present study: Argentine. New record: 84 specimens (Online Resource 1).
Bathymetric range 339–357 m (Mah 2011). Present study 139–1900 m.
Remarks This genus was cited as Pillsburiaster sp. by Brogger et al. (2013c; report submarine canyons survey, II/III B/O Puerto Deseado at Rio de la Plata, Argentine), but the specimens were not identified as P. calvus; therefore, the present record will be first recorded in Argentinian waters. Genus Pillsburiaster includes 4 species that should be taken in account related to P. calvus, 1 species recorded in the South Atlantic Ocean, Pillsburiaster geographicus Halpern, 1970 (Mexico and Gulf of Guinea), and 3 in New Zealand, Pillsburiaster aoteanus McKnight 1973, Pillsburiaster maini McKnight 1973 and Pillsburiaster indutilis McKnight 2006. These species are distinguished on the basis of the arrangement of their papular areas.
Family Solasteridae Viguier 1878
Genus Paralophaster Fisher 1940
Paralophaster lorioli (Koehler 1907), new record (Fig. 3i–m).
AphiaID: 234937
Diagnosis Abactinal surface with isolated paxillae, each with a short peduncle bearing 6–8 elongated spines (Fig. 3i, j). Marginal plates no more than 12 in each series. Large marginal spines in cluster arrangement (Fig. 3k, l). The actinal surface presents a blunt tuber-shaped prominence, with 4–5 spines with denticulations at their ends. Adambulacral plates with 3 spines in the furrow. They have 6 spines on the oral plate at their free edge (Fig. 3m).
Distribution The Scotia Arc, Antarctic Peninsula, Kerguelen, Ross Sea. Present study: Argentine. New record: 1 specimen (Online Resource 1).
Bathymetric range 104–294 m (USNM E 13,476; USNM E 13,474 from Invertebrate Collection Database, NMNH). Present study 1551 m.
Remarks Paralophaster lorioli could be confused with Paralophaster antarcticus (Koehler 1912). Paralophaster lorioli has few marginal plates, no more than 12 in each series with arrangement of few marginal spines in cluster, while P. antarcticus has more than 12 marginal plates that carry many spines much larger than the adjacent paxillae.
Order Velatida Perrier 1884
Family Pterasteridae Perrier 1875
Genus Pteraster Müller and Troschel 1842
Pteraster flabellifer Mortensen 1933, new record (Fig. 3n–q).
AphiaID: 178571
Diagnosis R/r = 1.9/1, 6 arms, abactinal membrane thin, 5–6 slender paxillar spines, small osculum surrounded by a serial of spines forming a ridge that are not embedded in a web (Fig. 3n). The marginal fringe is evident thanks to the stout marginal spines (Fig. 3p), 6–7 adambulacral spines in unequal series like a fan webbed (Fig. 3q), 7 oral spines half of their length webbed, one hyaline, multitipped suboral spine (Fig. 3o).
Distribution Only one record in Cape Town (South Africa) and Durban. Present study. Argentine. New record: 1 specimen (Online Resource 1).
Bathymetric range 272–366 m (Clark and Downey 1992). Present study 723 m.
Remarks: Pteraster flabellifer could be confused with Pteraster obscurus (Perrier 1891), Pteraster stellifer Sladen 1882, Pteraster affinis Smith 1876, or Pteraster militaris (OF Müller 1776). Pteraster obscurus present the 6–8 arms, but paxillar columns bear numerous spines (more than 6), and a thick web where adambulacral and oral spines are webbed for totality of their length; its area of distribution restricted to the north Atlantic (boreal belt). Related to the other remarkably similar species distributed around Atlantic Ocean, P. stellifer and P. affinis, both with 5 arms, are species from the South Hemisphere; however, P. stellifer has a parchment-like supradorsal membrane, large actinolateral spines, and unwebbed oral spines. Although P. militaris evenly could have 6 arms, its distribution is restricted to the North Hemisphere. In any case, Clark discusses about the possibility that these three species (P. affinis, P. flabellifer, and P. militaris) “may eventually prove to favor only a subspecific relationship” (Clark and Downey 1992), based on results reported in Clark (1962).
Argentinian deep-sea asteroids new checklist
The new Argentinian deep-sea checklist consists of 64 asteroids species that emerge from 2198 records: 887 records from Atlantis surveys and 1311 records from publicly available datasets and literature (Table 4, Fig. 4). These results include the seven new records proposed in the present work.

Results from records reliability study. Analysis of 64 valid species after reliability study—new checklist for Argentinian waters. See species list in Table 4
The geographical distribution of Argentinian Deep-sea asteroids
Of the 64 species included in the new Argentinian Asteroid checklist (Table 4), 7 are exclusive for ADW (Asterina stellifera, Hymenaster pergamentaceus, Lethasterias australis, Perissasterias polyacantha, Plutonaster bifrons, Pteraster flabellifer and Radiaster elegans), and the rest are shared with nearby areas: 23 with SA, SAI, and M/FI, and 2 (Henricia diffidens and Odontaster roseus) only with Antarctica (SA were not included) (Fig. 5a).

Argentinian deep-sea asteroid distribution areas (ADW) and their Antarctic (A) connections. Preliminary biogeographical hypothesis following Griffiths et al. (2009). Based on: a Malvinas/Falkland Islands MFI), Scotia Arc (SA), and sub-Antarctic Islands (SI) are independent entities; b ADW and M/FI Patagonian and SA as Antarctic province; c ADW and M/FI sub-Antarctic, and SA as Antarctic province; d ADW, M/FI, and SA sub-Antarctic; e ADW and M/FI Patagonian and SA sub-Antarctic. In red, new records. In blue, species from ADW shared only with A
Discussion
Reliability of the databases and Argentinian new asteroid checklist
Concerning the species composition in the study area, 64 species should be considered valid records in this area (Bernasconi 1937, 1962a, 1963, 1964a, 1966b, 1973a, 1973b, 1979; Gutt and Starmans 1998; Tablado and Maytia 1988; Orovitz and Tablado 1990; Bastida et al. 1992; Clark and Downey 1992; Gutt et al. 1999; Bremec et al. 2000; Rios et al. 2003; Zaixso 2004; Bertness et al. 2006; Mutschke and Rios 2006; Mah 2011; Mah and Blake, 2012; Brogger et al. 2013a; Souto et al. 2014; Arribas et al. 2016; Fraysse et al. 2018; Moreau et al. 2018, 2021; Guillaumot et al. 2020; National Museum of Natural History (NMNH: Research and Collections Information System, Smithsonian Institution), but only 41 were captured. Consequently, 23 of these species were not reported (Table 4; Fig. 4). These absences could be due to various reasons, including: (1) samplers’ limitations or (2) more likely their low frequency, which makes them difficult to be capture. The latter reason could be directly related to the special features of canyons, morphology, granulometry, or even other biotic factors such as species composition (del Rio et al. 2012). A deeper ecological study will be published in the near future providing more useful information that allows us to explain these absences.
The distribution areas of Argentinian asteroids and their Antarctic connections
The first study approach focuses on the number of shared species in ADW and the remainder of the four entities (provinces/regions) defined as M/FI, SA, SAI, and A (discussed in Griffiths et al. 2009, based on Moore et al. 1999) (Fig. 5).
Under the geographical/countries limits and depth range that frame the present work, ADW shares 89.06% of species with these areas, and only ten species should be considered “Patagonian species” (ADW + M/FI) (Fig. 5a, e framed species list), and they are not present outside of this traditionally named area. Three of them correspond to our proposed new records (discussed above), and the remaining seven are: Cryptasterias brachiata and Lethasterias australis, endemic in Patagonian waters; Hymenaster pergamentaceus and Asterina stellifera, frequent only in the Southwest Atlantic Ocean; Anasterias spirabilis and Asterina fimbriata, species living on the limit of the sub-Antarctic area; and Perisasterias polyacantha, which is frequent in Australia, rare in southern Africa, and very scarce in Argentine (Tablado and Maytia 1988; Brogger et al. 2013a).
Despite the number of ADW species present in the nearby waters, M/FI, SA, and SI altogether share only 35.93% of ADW species (Fig. 5a). However, when they were analyzed separately, we realized that ADW and SA share 81.25% of species, whereas M/FI shares a lower percentage (56.25%) despite their proximity (Fig. 5a). These results could support the idea that M/FI would be an independent entity, as supported by Figuerola et al. (2017) and Griffiths et al. (2009) in the case of gastropods (Fig. 5a). However, this argument could be based on a false premise since, in our study, only deep-sea asteroids were considered and many recorded species at M/FI usually live in shallower waters. In any case, authors who defend other hypotheses did not consider ADW and M/FI as independent entities.
Nevertheless, it is remarkable that 100% of the species from ADW recorded at SAI are included in the SA area (Fig. 5a), which endorses our idea of joining these two areas together as a sub-Antarctic area (Fig. 5d, e), and (under this bathyal context), as has been reported by Briggs and Bowen (2012) and Griffiths et al. (2009) in the case of Bryozoa. On the contrary, other authors defend the hypothesis that SA belongs to the Antarctic area [Longhurst 2007; Martin-Ledo and López-González 2014; Griffiths et al. 2009 (Pycnogonida; Bryozoa); Moreau et al 2017, among others] (Fig. 5b, 5c).
On the basis of the discussion presented above, only one hypothesis could adjust to our preliminary biogeographic results: ADW, M/FI, and SA should be considered sub-Antarctic, depending on whether SA could be a separated entity of Antarctica, based on the fact that ADW and SA share a greater number of species than any other (Fig. 5a) when the rest of the Antarctica region is included (Fig. 5d). Therefore, to confirm or refute this hypothesis (Rodríguez et al. 2007; Martin-Ledo and López-González 2014; Weir and Stanworth 2019; Moreau et al. 2017, among others), some considerations should be taken in account: (1) we should include more records from known surveys that did not incorporate public datasets (such as the BENTART projects, and others); (2) we need to analyze the benefit of including shallower species, or follow Watling et al. (2013), who opted for conducting their studies at the bathyal and abyssal provinces levels, depending on the focus; (3) and, finally, a deeper analysis should be carried out using different biogeographical methods that would be associated with environmental variables (our next target).
Data availability
All data included in this paper will be sent to Antarctic Biodiversity Information Facility (AntBIF) after data are published. The material will be shared with different museums.
Code availability
Not applicable.
References
Ahearn CG (1995) Catalog of the type specimens of seastars (Echinodermata: Asteroidea) in the National Museum of Natural History, Smithsonian Institution. Smithson Contrib Zool 572:1–59. https://doi.org/10.5479/si.00810282.572
Alvarado JJ, Solís-Marín FA (2013) Echinoderm research and diversity in Latin America. Springer, Berlin
Arribas LP, Martinez MI, Brogger MI (2016) Echinoderms in San Matías Gulf, Southwestern Atlantic Ocean. Thalassas 32:11–18
Arribas LP, Bagur M, Palomo MG, Bigatti G (2017) Population biology of the sea star Anasterias minuta (Forcipulatida: Asteriidae) threatened by anthropogenic activities in rocky intertidal shores of San Matías Gulf, Patagonia, Argentina. Rev Biol Trop 65:73–84
Bastida R, Roux A, Martinez DE (1992) Benthic communities of the Argentine continental shelf. Oceanol Acta 15:687–698
Berecoechea JJ, Brogger MI, Penchaszadeh PE (2017) New evidence of brooding in the deep-sea brittle star Astroroma agassizii Lyman, 1876 from a South Western Atlantic Canyon. Deep Sea Res Part I 127:105–110. https://doi.org/10.1016/j.dsr.2017.08.007
Bernasconi I (1934) Los equinodermos de los mares argentinos. Observaciones sobre algunos “erizos de mar”, “estrellas de mar” y otras formas comunes en nuestro litoral. Bol Cent Naval 53:1–21
Bernasconi I (1935) Dos nuevas especies de Pteraster en la Argentina. Physis 11:482–483
Bernasconi I (1937) Asteroideos argentinos I: Familia Pterasteridae. An Mus Argentino de Cien Nat “Bernardino Rivadavia” 39:167–187
Bernasconi I (1941a) Dos nuevas especies argentinas de Luidia. Physis 19:117–118
Bernasconi I (1941b) Los equinodermos de la expedición del Buque Oceanográfico ‘“Comodoro Rivadavia”’ ARA. Physis 19:37–49
Bernasconi I (1943) Los asteroideos sudamericanos de la familia Luidiidae. An Mus Argentino de Cien Nat “Bernardino Rivadavia” 61:1–20
Bernasconi I (1957) Equinoideos y asteroideos de la colección del Instituto Oceanográfico de la Universidad de San Pablo. Bol Inst Oceanogr 7:119–148
Bernasconi I (1961) Una Nueva Especie De Asteroideo. Neotrop 7:1–2
Bernasconi I (1962a) Asteroideos argentinos. III. Familia Odontasteridae. Rev Mus Argentino Bernardino Rivadavia 8:27–58
Bernasconi I (1962b) Perknaster densus patagonicus nueva sub especie de la Argentina. Physis 23:257–258
Bernasconi I (1963) Asteroideos argentinos IV. Familia Goniasteridae. Rev Mus Argentino De Cien Nat Bernardino Rivadavia Cienc Zool 9:1–26
Bernasconi I (1964a) Asteroideos argentinos V. Familia Ganeriidae. Rev Mus Argentino De Cien Nat Bernardino Rivadavia Cienc Zool 9:59–89
Bernasconi I (1964b) Asteroideos argentinos. Claves para los órdenes, familias, subfamilias y géneros. Physis 24:241–277
Bernasconi I (1965) Nuevo género y nueva especie abisal de Goniasteridae (Equinodermata, Asteroidea). Physis 25:333–335
Bernasconi I (1966a) Descripción de una nueva especie de Calyptraster. (Asteroidea, Pterasteridae). Physis 26:95–99
Bernasconi I (1966b) Los equinoideos y asteroideos colectados por el buque oceanográfico R/V “Vema”, frente a las costas argentinas, uruguayas y sur de Chile. Rev Mus Argentino De Cien Nat Bernardino Rivadavia Zool 9:147–175
Bernasconi I (1972) Nuevas especies de Asteroidea: Bathydiaster herwigi sp. nov. (Astropectinidae) y Calyptraster vitreus sp. nov. (Pterasteridae). Physis 31:9–14
Bernasconi I (1973a) Asteroideos argentinos VI. Familia Asterinidae. Rev Mus Argentino De Cien Nat Bernardino Rivadavia Hidrobiol 3:333–346
Bernasconi I (1973b) Los equinodermos colectados por el “Walter Herwig” en el Atlántico Sudoeste. Hidrobiol 3:287–334
Bernasconi I (1979) Asteriidae, coscinasteriinae de la argentina y antartida. Rev Mus Argentino De Cien Nat Bernardino Rivadavia Hidrobiol 5:241–249
Bernasconi I (1980) Asteroideos argentinos VII. Familia Echinasteridae Rev Mus Argentino De Cien Nat Bernardino Rivadavia Hidrobiol 4:247–258
Bertness MD, Crain CM, Siliman BR, Bazterrica MC, Reyna MV, Hidalgo F, Farina JK (2006) The community structure of western Atlantic Patagonian rocky shores. Ecol Monogr 76:439–460
Bremec C, Brey T, Lasta M, Valero J, Lucifora L (2000) Zygochlamys patagonica beds on the Argentinian shelf. Part I: energy flow through the scallop bed community. Arch Fish Mar Res 48:295–303
Briggs JC, Bowen BW (2012) A realignment of marine biogeographic provinces with particular reference to fish distributions. J Biogeogr 39:12–30
Brogger MI, O’Hara TD, (2015) Revision of some ophiuroid records (Echinodermata: Ophiuroidea) from Argentina. Zootaxa 3972:432–440
Brogger MI, Martinez MI, Penchaszadeh PE (2010) Reproduction of the sea urchin Arbacia dufresnii (Echinoidea: Arbaciidae) from Golfo Nuevo, Argentina. J Mar Biol Assoc UK 90:1405–1409. https://doi.org/10.1017/S0025315410000445
Brogger MI, Gil DG, Rubilar T, Martinez MI, Díaz de Vivar E, Escolar M, Epherra L, Perez AF, Tablado A (2013a) Echinoderms from Argentina: biodiversity, distribution and current state of knowledge. In: Alvarado JJ, Solís-Marín FA (eds) Echinoderms research and diversity in Latin America. Springer, Berlin, pp 359–402
Brogger MI, Martinez MI, Zabala S, Penchaszadeh PE (2013b) Reproduction of Ophioplocus januarii (Echinodermata: Ophiuroidea): a continuous breeder in northern Patagonia, Argentina. Aquat Biol 19:275–285. https://doi.org/10.3354/ab00537
Brogger MI, Berecoechea JJ, Flores J, Rivadeneira P, Martinez M (2013c) Diversidad de equinodermos de las campañas Talud Continental 2 y 3. In: Penchaszadeh P, Informe de Campaña Cañon Submarino II/III B/O “Puerto deseado”. CONICET, pp 43–50
De Broyer C, Koubbi P, Griffiths HJ, Raymond B, d’Udekem d’Acoz C, Van de Putte AP, Danis B, David B, Grant S, Gutt J, Held C, Hosie G, Huettmann F, Post A, Ropert-Coudert Y (2014) Biogeographic Atlas of the Southern Ocean. SCAR, Cambridge
Campagna C, Piola AR, Marin MR, Lewis M, Zajaczkovski U, Fernández T (2007) Deep divers in shallow seas: Southern elephant seals on the Patagonian shelf. Deep Sea Res Part I 54:1792–1814
Carames A, Martinez M, Concheyro A, Remirez M, Adamonis S (2019) Holothuroidea: new records from the Lower Cretaceous of the Agrio Formation, Neuquén Basin, Argentina. An integrated study with foraminifers and calcareous nannofossils. Alcheringa 43:580–596. https://doi.org/10.1080/03115518.2019.1641617
Clark AM (1962) Asteroidea. B.A.N.Z. Antarctic Research Expedition 1929–1931. Rep Ser B (zool Bot) 9:1–104
Clark AM, Downey ME (1992) Starfishes of the Atlantic. Chapman and Hall, London
Clark HES, McKnight DG (2000) The marine Fauna of New Zealand: Echinodermata: Asteroidea (Sea-stars). Order Paxillosida & Order Notomyotida. NIWA Biodivers Memoirs 116:1–196
Cossi PF, Boy CC, Perez AF (2017) Pattern of energy allocation during the gametogenesis of the asteroid Cosmasterias lurida (Forcipulatida: Stichasteridae) from the Beagle Channel, Ushuaia, Argentina. Rev Biol Trop 65:197–206
Danis B, Griffiths HJ, Jangoux M (2014) Asteroidea. In: De Broyer C, Koubbi P, Griffiths HJ, Raymond B, d’Udekem d’Acoz C, Van de Putte AP, Danis B, David B, Grant S, Gutt J, Held C, Hosie G, Huettmann F, Post A, Ropert-Coudert Y (eds) Biogeographic atlas of the Southern Ocean. SCAR, Cambridge, pp 200–208
Del Río JL, Acosta J, Cristobo J, Martínez J, Parra S, Tel E, Viñas L, Elvira E, Patrocinio T, Rios P, Almón B, Blanco R, Murillo J, Polonia V, Fernández J, Cabanas JM, Gago JM, Gonzalez G, Cabrero A, Besada MV, Schultze F, Franco A, Bargiela J, Garcia X (2012) Estudios de los Ecosistemas Marinos Vulnerables en aguas internacionales del Atlántico sudoccidental. Proyecto Atlantis. Programa Pesquerías Lejanas. Temas de Oceanografía 6. IEO, Madrid
Eleaume M, Hemery LG, Roux M, Améziane N (2014) Southern Ocean Crinoids. In: De Broyer C, Koubbi P, Griffiths HJ, Raymond B, d’Udekem d’Acoz C, Van de Putte AP, Danis B, David B, Grant S, Gutt J, Held C, Hosie G, Huettmann F, Post A, Ropert-Coudert Y (eds) Biogeographic atlas of the Southern Ocean. SCAR, Cambrigde, pp 208–213
Epherra L, Crespi-Abril A, Meretta PE, Cledón M, Morsan EM, Rubilar T (2015) Morphological plasticity in the Aristotle’s lantern of Arbacia dufresnii (Phymosomatoida: Arbaciidae) off the Patagonian Coast. Rev Biol Trop 63:339–351
Epherra L, Martelli A, Morsan EM, Rubilar T (2017) Parámetros poblacionales del erizo de mar Arbacia dufresnii (Arbacioida, Arbaciidae) en golfos norpatagónicos invadidos por el alga Undaria pinnatífida (Laminariales, Alariaceae). Rev Biol Trop 65:101–112
Figuerola B, Barnes DKA, Brickle P, Brewin PE (2017) Bryozoan diversity around the Falkland and South Georgia Islands: overcoming Antarctic barriers. Mar Environ Res 126:81–94
Fisher WK (1913) Four new genera and fifty-eight new species of starfishes from the Philippine Islands, Celebes, and the Moluccas. Scientific results of the Philippine cruise of the Fisheries Steamer “Albatross” 1907–1910. Proc US Natl Mus 43:599–648
Flores JN, Brogger MI, Penchaszadeh PE (2019) Reproduction and development of the brooding sea urchin Austrocidaris canaliculata from deep-sea off Argentina. Deep Sea Res Part I 143:35–42. https://doi.org/10.1016/j.dsr.2018.11.012
Fraysse CP, Calcagno JA, Perez AF (2018) Asteroidea of the southern tip of South America, including Namuncurá Marine Protected Area at Burdwood Bank and Tierra del Fuego Province, Argentina. Polar Biol 41:2423–2433
Fraysse CP, Boy CC, Becker YA, Calcagno JA, Perez AF (2020) Brooding in the Southern Ocean: the case of the Pterasterid Sea Star Diplopteraster verrucosus (Sladen, 1882). Biol Bull 239:1–12. https://doi.org/10.1086/709664
Gil DG, Lopretto EC, Zaixso HE (2020) Reproductive timing and synchronized reproduction of the sea urchin Pseudechinus magellanicus (Echinoidea: Temnopleuridae) in central Patagonia, Argentina. Mar Biol Res 16:311–326
Di Giorgio G, Rubilar T, Brogger MI (2015) Histological analysis after arm tip amputation in the brittle star Ophiopoclus januarii (Echinodermata: Ophiuroidea). Rev Biol Trop 63:297–308. https://doi.org/10.15517/rbt.v63i2.23164
Griffiths HJ, Barnes DKA, Linse K (2009) Towards a generalized biogeography of the Southern Ocean benthos. J Biogeogr 36:162–177
Guillaumot C, Moreau C, Danis B, Saucède T (2020) Extrapolation in species distribution modelling. Application to Southern Ocean marine species. Prog Oceanogr 188:102438. https://doi.org/10.1016/j.pocean.2020.102438
Gutt J, Helsen E, Arntz WE, Buschmann A (1999) Biodiversity and community structure of the mega-epibenthos in the Magellan region (South America). Sci Mar 63:155–170
Gutt J, Starmans A (1998) Structure and biodiversity of megabenthos in the Weddell and Lazarev Seas (Anarctica): ecological role of physical parameters and biological interactions. Polar Biol 20:229–247. https://doi.org/10.1007/s003000050300
Hedgpeth JW (1969) Introduction to Antarctic zoogeography: Distribution of selected groups of marine invertebrates in waters south of 35°S latitude. In: Bushnell VC, Hedgpeth JW (eds) Distribution of selected groups of marine invertebrates in waters south of 35° latitude. Antarctic Map Folio series. American Geographical Society, New York, pp 1–9
Hunter AW, Rushton AWA, Stone P (2016) Comments on the ophiuroid family Protasteridae and description of a new genus from the Lower Devonian of the Fox Bay Formation, Falkland Islands. Alcheringa 40:429–442. https://doi.org/10.1080/03115518.2016.1218246
Longhurst AR (2007) Ecological geography of the sea, 2nd edn. Academic Press, San Diego
Mah CL (2011) Taxonomy of high-latitude Goniasteridae (Subantartic & Antartic): one new genus, and three new species with an overview and key to taxa. Zootaxa 2759:1–48
Mah CL (2018) New genera, species and occurrence records of Goniasteridae (Asteroidea; Echinodermata) from the Indian Ocean. Zootaxa 4539:1–116
Mah CL, Blake DB (2012) Global diversity and phylogeny of the Asteroidea (Echinodermata). PLoS ONE 7:e35644
Mah CL, McKnight DG, Eagle MK, Pawson DL, Ameziane N, Vance DJ, Baker AN, Clark HES, Davey N (2009) Phylum Echinodermata: sea stars, brittle stars, sea urchins, sea cucumbers, sea lilies. In: Gordon DP (ed) New Zealand inventory of biodiversity, vol 1. Kingdom Animalia. Radiata, Lophotrochozoa, Deuterostomia. Canterbury University Press, Christchurch, pp 371–400
Mah CL (2022) World Asteroidea Database. Asteroidea de Blainville, 1830. World Register of Marine Species. https://www.marinespecies.org/aphia.php?p=taxdetails&id=123080
Martinez MI (2016) The sea cucumber Psolus patagonicus (Echinodermata: Holothuroidea) from the southwestern Atlantic: redescription of the holotype and a new synonym. Rev Mus Argentino De Cien Nat 18:237–241
Martinez MI, Brogger MI (2012) Thandarum hernandezi, a new genus and new species of sea cucumber family Sclerodactylidae (Echinodermata: Holothuroidea: Dendrochirotida) from the Southwestern Atlantic Ocean. Zootaxa 3304:63–68
Martinez MI, Penchaszadeh PE (2017) A new species of brooding Psolidae (Echinodermata: Holothuroidea) from deep-sea off Argentina, Southwestern Atlantic Ocean. Deep Sea Res Part II 146:13–17
Martinez MI, Arribas LP, Berecoechea JJ, Brogger MI, Penchaszadeh PE (2015) Echinoderm diversity in the Southwestern Atlantic. Rev Biol Trop 63:115–120
Martinez MI, Alba-Posse EJ, Lauretta D, Penchaszadeh PE (2018) Developmental stages in the brooding sea cucumber Cladodactyla crocea (Lesson, 1830) in the Southwestern Atlantic Ocean. Polar Biol 41:1237–1244. https://doi.org/10.1007/s00300-018-2280-y
Martinez MI, Alba-Posse EJ, Lauretta D, Penchaszadeh PE (2020) Reproductive features in the sea cucumber Pentactella perrieri (Ekman, 1927) (Holothuroidea: Cucumariidae): a brooding hermaphrodite species from the southwestern Atlantic Ocean. Polar Biol 43:1383–1389
Martin-Ledo R, López-González PJ (2014) Brittle stars from Southern Ocean (Echinodermata: Ophiuroidea). Polar Biol 37:73–88
Martins L, Tavares M (2019) Two new species of Psolus Oken from Brazil (Holothuroidea: Psolidae), with neotype designation and redescription of Psolus vitoriae Tommasi, 1971, and a key to the southwestern Atlantic and Magellanic species. Zootaxa 4563:531–546
McKnight DG (2006) The Marine Fauna of New Zealand: Echinodermata: Asteroidea (Sea-stars). 3. Orders Velatida, Spinulosida, Forcipulatida, Brisingida with addenda to Paxillosida, Valvatida. NIWA Biodivers Memoirs 120:1–187
Moore JK, Abbott MR, Richman JG (1999) Location and dynamics of the Antarctic Polar Front from satellite sea surface temperature data. J Geophys Res 104:3059–3073
Moreau C, Saucède T, Jossart Q, Agüera A, Brayard A, Danis B (2017) Reproductive strategy as a piece of the biogeographic puzzle: a case study using Antarctic sea stars (Echinodermata: Asteroidea). J Biogeogr 44:848–860
Moreau C, Mah C, Agüera A, Améziane N, Barnes D, Crokaert G, Eléaume M, Griffiths H, Guillaumot C, Hemery L, Jazdzewska A, Jossart Q, Laptikhovsky V, Linse K, Neill K, Sands C, Saucède T, Schiaparelli S, Sicinski J, Vasset N, Danis B (2018) Antarctic Sub-Antarctic Asteroidea Database Zookeys 747:141–156
Moreau C, Danis B, Jossart Q, Eléaume M, Sands C, Achaz G, Agüera A, Saucède T (2019) Is reproductive strategy a key factor in understanding the evolutionary history of Southern Ocean Asteroidea (Echinodermata)? Ecol Evol 9:8465–8478
Moreau C, Jossart Q, Danis B, Eléaume M, Christiansen H, Guillaumot C, Downey R, Saucède T (2021) The high diversity of Southern Ocean sea stars (Asteroidea) reveals original evolutionary pathways. Prog Oceanogr 190:102472
Mortensen T (1927) Handbook of the echinoderms of the British Isles. Humphrey Milford Oxford University Press, Copenhagen
Murillo FJ, Serrano A, Kenchington E, Mora J (2016) Epibenthic assemblages of the Tail of the Grand Bank and Flemish Cap (northwest Atlantic) in relation to environmental parameters and trawling intensity. Deep Sea Res Part I 109:99–122
Mutschke E, Rios C (2006) Distribución especial y abundancia relativa de equinodermos en el estrecho de Magallanes, Chile. Ciencia y Tecnología del Mar 29:91–102
Orovitz B, Tablado A (1990) Asterodon singularis (Müller y Troschel 1843), única especie del género Asterodon Perrier 1891 (Asteroidea: Odontasteridae). Gayana Zool 54:21–32
Pérez AF, Fraysse C, Boy CC, Epherra L, Calcagno J (2017) Reproductive biology and energetics of the brooding sea star Anasterias antarctica (Echinodermata: Asteroidea) in the Beagle Channel, Tierra del Fuego, Argentina. Rev Biol Trop 65:221–232
Piola AR, Rivas A (1997) Corrientes en la Plataforma Continental. In: Boschi, EE (ed) El Mar Argentino y sus Recursos Pesqueros. INIDEP, Mar del Plata, pp 119–132
Piola AR, Matano RP (2001) Brazil and Falklands (Malvinas) Currents. In: Steele JH, Thorpe SA, Turekian KK (eds) Encyclopedia of ocean sciences. Academic, San Diego, pp 340–349
Ríos C, Mutschke E, Morrison E (2003) Biodiversidad bentónica sublitoral en el estrecho de Magallanes, Chile. Rev Biol Mar Oceanogr 38:1–12
Rivadeneira PR, Brogger MI, Penchaszadeh PE (2017) Aboral brooding in the deep water sea star Ctenodiscus australis Lütken, 1871 (Asteroidea) from the Southwestern Atlantic. Deep Sea Res Part I 123:105–109
Rivadeneira PR, Martinez MI, Penchaszadeh PE, Brogger MI (2020) Reproduction and description of a new genus and species of deep-sea asteriid sea star (Echinodermata; Asteroidea) from the southwestern Atlantic. Deep Sea Res Part I 163:103348
Rivas AL, Dogliotti AI, Gagliardini DA (2006) Seasonal variability in satellite-measured surface chlorophyll in the Patagonian Shelf. Cont Shelf Res 26:703–720
Rodríguez E, López-González PJ, Gili JM (2007) Biogeography of Antarctic sea anemones (Anthozoa, Actinaria): what do they tell us about the origin of the Antarctic benthic fauna? Deep Sea Res Part II 54:1876–1904
Romanelli MV, Tablado A (2011) Leptychaster kerguelenensis (Echinodermata: Asteroidea) from Southwest Atlantic: Redescription and geographic distribution. Zootaxa 2971:59–68
Saucède T, Pierrat B, David B (2014) Chapter 5.26 Echinoids. In: De Broyer C, Koubbi P, Griffiths HJ, Raymond B, d’Udekem d’Acoz C, Van de Putte AP, Danis B, David B, Grant S, Gutt J, Held C, Hosie G, Huettmann F, Post A, Ropert-Coudert Y (eds) Biogeographic atlas of the Southern Ocean. SCAR, Cambridge, pp 213–220
Sladen WP (1882) The Asteroidea of HMS Challenger Expedition Part I. Pterasteridae. J Linn Soc Lond 16:189–246
Sladen WP (1883) The Asteroidea of HMS Challenger expedition Part II. Astropectinidae. J Linn Soc Lond 17:214–269
Souto V, Escolar M, Genzano G, Bremec C (2014) Species richness and distribution patterns of echinoderms in the southwestern Atlantic Ocean (34–56°S). Sci Mar 78:269–280
Tablado A (1982) Asteroideos argentinos. Familia Poraniidae. Com Mus Argentino Cien Nat Hidrobiol 2:87–106
Tablado A, Maytia S (1988) Presencia Perissasterias polyacantha H.L. Clark 1923 (Echinodermata, Asteroidea) en el Atlántico sudoccidental. Com Zool Mus Hist Nat Montevideo 12:1–11
Watling L, Guinotte J, Clark MR, Smith CR (2013) A proposed biogeography of the deep ocean floor. Prog Oceanogr 111:91–112
Weir CR, Stanworth A (2019) The Falkland Island (Malvinas) as sub-Antarctic foraging, migratory and wintering habitat for southern right whales. J Mar Biol Assoc UK 100:1–11
Wilkie IC, Brogger MI (2018) The peristomial plates of ophiuroids (Echinodermata: Ophiuroidea) highlight an incongruence between morphology and proposed phylogenies. PLoS ONE 13:e0202046
Zaixso HE (2004) Bancos de cholga Aulacomya atra atra (Molina) (Bivalvia: Mytilidae) del golfo San José (Chubut, Argentina): diversidad y relaciones con facies afines. Rev Biol Mar Oceanogr 39:61–78
Acknowledgements
We want to acknowledge ATLANTIC Team (Oceanographic Instituto Español de Oceanografía: IEO), especially Dr. José Luis del Río (IEO, Vigo) Principal Investigator (PI) of ATLANTIS Projects, and Dr. Javier Cristobo (IEO, Gijon) Epifauna Responsible for their support and their confidence in our achievements. We also acknowledge Dr. Araceli Muñoz, Geologist from TRAGSA-IEO for improving our bathymetry database, Dr. Pilar Ríos for her cooperation and support, and, finally, SELUMA Team for their patience and support in finishing this work. We also thank Christopher Mah and Narissa Bax for their comments and recommendations that helped us to improve the manuscript, and Sandra Meneaud from FGUMA Translation and Interpreting Service for the English review.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. Funding for open access charge: Universidad de Málaga / CBUA. This work has been funded by ATLANTIS Projects [IEO, Secretary General of Maritime Fishing (SGP) and Economy and Competivity Ministry. Spanish Government (2007–2010)], Puente Projects (Mod.B5) funded by UMA, and Own Found from the Junta de Andalucía, research groups SELUMA.
Author information
Authors and Affiliations
Contributions
J.H.G. and M.E.M.C. contributed equally in conceiving, designing, and writing the manuscript and in research. M.E.M.C. is the senior author.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest regarding this manuscript.
Ethical approval
None.
Consent to participate
We obtained consent to participate.
Consent for publication
We obtained consent for publication.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
Cite this article
Hurtado-García, J., Manjón-Cabeza, M.E. Species composition of sea stars (Echinodermata: Asteroidea) in the Patagonian Argentinian deep sea, including seven new records: connectivity with sub-Antarctic and Antarctic fauna. Polar Biol 45, 1211–1228 (2022). https://doi.org/10.1007/s00300-022-03056-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-022-03056-x