
Abstract
In the last three decades, several interdisciplinary studies investigated the marine ecosystems off the West Antarctic Peninsula (WAP), one of the most impacted areas of the Southern Ocean by the global warming. Although the extent of near-shore habitats along the WAP is wider than elsewhere in Antarctica, the coastal fish communities have been rarely studied. Complementing these previous studies, we provide new data on the species composition, population structure and relative abundance of the inshore fish community living off the Argentine Islands (Bellingshausen Sea). Fish samples were caught all the year round during four different periods spread over ten years (from 2006 to 2017). The fish fauna consisted of fourteen high-Antarctic and low-Antarctic species of notothenioids, most of them belonging to the Nototheniidae. Notothenia coriiceps was by far the most abundant species, followed in decreasing abundance by Chaenocephalus aceratus, Notothenia rossii, Trematomus newnesi and Trematomus bernacchii. Our findings provide context for future ecological studies as this area represents either a spawning and nursery area for multiple species in this study. More generally, the inshore waters off the Argentine Islands represent the southern limit of distribution for several low-Antarctic species, and our results provide critical baseline data for assessing possible disruptions in population dynamics driven by the ongoing climate change.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The marine ecosystems of the West Antarctic Peninsula (WAP) extends for ~ 1500 km from the Bellingshausen Sea to the northern tip of the peninsula. As one of the most impacted Antarctic region by the recent global warming (Ducklow et al. 2013), it drew the attention of several interdisciplinary studies, such as the Research on Antarctic Coastal Ecosystem Rates (Huntley et al. 1991), the Palmer Antarctic Long-Term Ecological Research (Smith et al. 1995), and the Southern Ocean Global Ocean Ecosystems Dynamics program (Hofmann et al. 2004). As life cycles and population dynamics of organisms in the Antarctic coastal marine ecosystem largely depend on the annual cycle and interannual variations in sea ice cover that, in turn, are affected by the ongoing rapid regional warming (Smith et al. 1998), it would be advisable to acquire long-term data to evaluate and foresee changes in species composition and relative abundance within the ecological communities.
A relatively poorly known area along the WAP is the Argentine Islands, located in the Wilhelm Archipelago between 65°13′–65°16′ S and 64°10′–64°20′ W. According to Daniels and Lipps (1982), the Argentine Islands are of high faunistic interest, showing the highest species richness (18 species) and diversity (Shannon index H′ = 0.84) among twenty-two sites located off the WAP across a wide latitudinal range. The species composition of shallow coastal waters off the Argentine Islands closely resembles those reported elsewhere along the WAP, consisting mainly of notothenioid fish. During five sampling years (2002–2006), a total of 15 species were collected, although the bulk of catches was primarily represented by Notothenia coriiceps and Trematomus bernacchii, followed in decreased order by Trematomus newnesi, Chaenocephalus aceratus and Notothenia rossii (Manilo 2006; Manilo et al. 2009). More recent data on the local fish fauna are sparse and refer only to single sampling seasons (Veselskyy and Khoetskyy 2018; Zinkovskyi et al. 2021).
During their remarkable evolutionary adaptive radiation, notothenioids occupied all habitats available on the continental shelf and slope, colonizing both benthic and pelagic environments. However, as a consequence of the extension of the ice sheet in the late Eocene-early Oligocene, the extent of near-shore and coastal habitats around the Continent is largely reduced, except in the shelf areas along the Scotia Arc and off the Antarctic Peninsula (Clarke and Johnston 1996). In addition, the near-shore environments are permanently covered by anchor ice and rearranged by the iceberg scouring (Barnes 1999), influencing the species composition and population dynamics of the local fish assemblages (e.g. Ruhl et al. 2003; Jurajda et al. 2016). The fish fauna inhabiting the shallow coastal waters has been seldom investigated on both sides of the Antarctic Peninsula, such as off James Ross Island on the eastern side (Jurajda et al. 2016) or at Doumer Island (Moreno et al. 1977) and off the Danco Coast (Casaux et al. 2003) on the western side. In these sites, the benthic fish community consisted of species of both low and high-Antarctic origin, including the genera Notothenia, Gobionotothen and Chaenocephalus and Trematomus, respectively.
The aim of the present study is to provide data on species composition, relative abundance and population structure of the inshore fish community off the Argentine Islands based on multi-year sampling activities. Complementing data reported by previous authors, we attempt to evaluate over a period lasting more than a decade possible changes in the fish community that could be related to the current regional warming.
Materials and methods
Study area
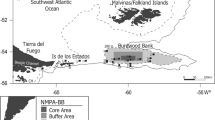
The Argentine Islands are located in the western side of the Antarctic Peninsula, being situated southwest of Petermann Island and separated from the Graham Land by the Penola Strait (Fig. 1). The archipelago includes several small, low and largely snow-covered islands (Irizar, Uruguay, Galindez, Winter, Skua, Corner, Grotto, Barchans and Forge), covering an area of approximately 5 km2. The islands are separated by channels and deep fjords, characterized by inshore shallow waters (< 50 m) with rocky bottoms and relatively weak currents flowing from variable directions, mainly influenced by the tidal activity (Artamonov et al. 2003). Since 1996, the Ukrainian Akademik Vernadsky station is operating at Marina Point, Galindez Island.

Map of the Argentine Islands, showing the main topographical features and the sampling sites. The Vernadsky Station located in Galindez island is indicated by an asterisk
Field sampling
Sampling activities were conducted all the year round during the XI (2006–2007), XII (2007–2008), XIV (2009–2010) and XXI (2016–2017) Ukrainian Antarctic Expeditions. Fish catches were obtained from several sites, most of them located off Galindez and Grotto Islands at depths between 5 and 50 m (Fig. 1). Due to environmental constraints (ice cover, ice floes, strong winds), fish samples were collected almost exclusively by fishing rods deployed from a boat or through holes in the pack ice (in winter) and, in case of bad weather conditions, directly from the shore. Fresh meat, molluscs and fish were used as bait. Fishing time was about 3–4 h a day, and it was considered a single haul. Overall sampling data, including hauls per month, number of fish per month and average number of fish per haul (i.e. CPUE) have been calculated to summarize fishing effort.
Laboratory work and data analyses
After capture, each specimen was identified to the species level according to main guides on Antarctic fishes (Fisher and Hureau 1985; Gon and Heemstra, 1990). Species nomenclature followed that proposed by Eastman and Eakin (2021). Fish were measured as total length (TL) and standard length (SL) with an accuracy of 0.1 cm, weighed as total weight (TW) and gutted weight (GW) with an accuracy of 1 g, and sexed. Gonads were removed from the abdominal cavity and weighed (gw) using a digital balance with an accuracy of 0.1 g.
A series of statistical tests were performed to infer demographic characteristics of sampled populations. Length frequency distributions were computed for each species and tested for sex-related differences applying the Kolmogorov–Smirnov two-samples test. Departure from the expected 1:1 sex ratio was tested using a χ2 goodness-of-fit test. To describe fish body growth, length–weight relationships were calculated for each sex applying an exponential model in the form TW = aSLb, where TW is the total weight, SL is the standard length, a and b are constants. In some cases, the low number of available specimens and/or the narrow size range of fish undermined the statistical power of the analysis. Isometric growth departure (i.e. b ≠ 3) was assessed by a t-test applied to the equation t = (b − 3) SE−1, where SE is the standard error of b. Allometric indices (b) were compared between sexes by applying a F-test (Sokal and Rohlf 1995). Finally, the gonadosomatic index (GSI) was calculated for each specimen as the proportion of gonad to somatic weight (gw/TW × 100). To evaluate the heterogeneity of the local fish community, diversity (H′, Shannon and Weaver 1949), evenness (J′, Pielou 1966) and species richness (SR, Margalef 1958) indices were calculated for each sampling year. All statistical analyses were performed using the STATISTICA 8 software (StatSoft, Inc., Tulsa, OK, USA), and tests were applied with a significance level of 0.05.
Results
During the four Antarctic Expeditions, a total of 227 hauls yielded 1,485 specimens belonging to 14 species of notothenioids (Table 1). The most speciose taxonomic group were the family Nototheniidae (11 species), followed by the Bathydraconidae, Channichthyidae and Harpagiferidae each with one species. The number of species (and the total number of fish) collected per year was proportional to the number of hauls, whereas the fish per haul were comparable among sampling years (Table 2). Except for the 2009–2010, the evenness (J′) was comparable among the other sampling years, whereas both diversity (H′) and species richness (SR) increased markedly in the more recent years (2016–2017), probably due to the risen fishing effort (Fig. 2). Sampling data and biological characteristics of each species, obtained from a representative subsample in case of massive catches, are summarized below.

Annual trend of diversity indices of the fish community living in inshore waters off the Argentine Islands. H′ diversity, J′ evenness, SR species richness
Trematomus peninsulae
This relatively rare species was caught in the Meek Channel during the sampling year 2016–2017, at depth of 30–35 m (Fig. 3). The sample was collected mainly in May and consisted of seven females and three males ranging between 14.0–19.5 cm and 12.5–15.6 cm SL, respectively. Females were all in spawning conditions (with GSI between 8.7 and 15.4%), indicating that this species is an autumn spawner.

Sampling depth (mean ± SD) for each species collected off the Argentine Islands
Gobionotothen gibberifrons
A single male of 21.0 cm SL was caught in August 2016 at 30 m depth (Fig. 3).
Lepidonotothen squamifrons
This species was represented by relatively small specimens collected in May 2016 at depth of 25 m (Fig. 3), consisting of two females and one male of 22.2–22.4 cm and 23.0 cm SL, respectively.
Nototheniops larseni
It was caught exclusively in the Meek Channel in May 2016 at 25–30 m (Fig. 3). The sample was composed of three females and one male measuring 15.7–18.2 cm and 16.2 cm SL, respectively. The larger female was close to spawning (GSI 8.0%), that likely took place in June-July.
Nototheniops nudifrons
A total of seven females ranging between 11.6 and 20.0 cm SL were collected in late autumn-early winter (May–August 2016) at 25 m depth (Fig. 3). The low value of GSI (0.3–2.5%) suggested they were far from the spawning season.
Notothenia coriiceps
It represents overwhelmingly the most abundant and widespread notothenioid fish inhabiting the inshore waters of the Argentine Islands from 1 to 50 m depth (Fig. 3). The CPUEs were one-two orders of magnitude higher than the other species, without any evident annual trend except for a slight decrease in 2009–2010 (Fig. 4). Similarly, the relative abundance ranged approximately between 76 and 91%, except for in 2016–2017 which accounted for 67% (Fig. 5). The selected sample was composed of 233 females and 273 males ranging between 17.0–40.0 cm and 15.2–40.5 cm SL, respectively (Fig. 6), with a sex ratio of 1:1 (χ2 = 3.16, df = 1, p = 0.07). The length frequency distributions differed between sexes, females showing a higher proportion of large specimens than males (D = 0.14, p = 0.016). Males and females showed a similar and isometric body growth (Table 3). The relatively high GSI of two females caught in May (12.2% and 15.6%) indicates they are approaching the reproductive season, that likely takes place in June.

Annual trend of catch per unit of effort (CPUE, i.e. number of fish per haul) calculated for the most abundant species sampled in inshore waters off the Argentine Islands. Note the logarithmic scale used for the CPUE

Annual trend of relative abundance calculated for the most abundant species sampled in inshore waters off the Argentine Islands

Length frequency distributions calculated for the most abundant species sampled in inshore waters off the Argentine Islands
Notothenia rossii
This species represented one of the most abundant species, having been caught in all sampling sites during the three most recent sampling years over a wide depth range (Fig. 3). The bulk of catches were recorded in 2007–2008, then relative abundance and CPUE dropped abruptly in the last two sampling years (Figs. 4 and 5). A total of 31 females and 38 males ranging between 18.2–40.8 cm and 21.2–34.5 cm SL were analysed (Fig. 6), with a sex ratio of 1:1 (χ2 = 0.52, df = 1, p = 0.47). The length frequency distributions did not differ between males and females (D = 0.29, p = 0.053), as well as the isometric body growth found in both sexes (Table 3). A single female of 44.5 cm TL caught in April was approaching the reproductive season (GSI of 13.8%), and likely was ready to migrate offshore with the breeding population.
Trematomus borchgrevinki
This species was sampled only in two occasions in the Meek Channel at 30–35 m (Fig. 3). A single male and female of 21.0 and 15.1 cm SL were caught in October 2006 and in May 2016, respectively.
Trematomus bernacchii
This species was caught over a wide depth range during all sampling years, mainly along the Meek and Stella Creek Channels. Starting from 2006 to 2007, relative abundance and CPUE dropped significantly in the following two sampling years, rising again in 2016–2017 (Figs. 4 and 5). The analysed sample consisted of 18 females and 12 males with size range of 14.4–27.0 cm and 12.5–18.0 cm SL, as well as three unsexed juveniles (Fig. 6). The sex ratio was 1:1 (χ2 = 0.83, df = 1, p = 0.36). The length frequency distributions were different between sexes (D = 0.66, p = 0.02), females being significantly larger than males. The length–weight relationships did not differ between sexes, showing both an isometric body growth (Table 3). Adult females sampled in August had a GSI range of 6.1–8.4%, suggesting they likely will spawn in the next spring, as reported in recent studies (e.g. Novillo et al. 2022).
Trematomus hansoni
A single, small female (18.2 cm SL) was caught in the Meek Channel on May 2016 at 25 m (Fig. 3).
Trematomus newnesi
This species was collected in the three most recent sampling years, mainly in the Meek Channel and off the Barchans Islands over a depth range of 20–30 m (Fig. 3). Relative abundance and CPUE were relatively high in 2007–2008, dropped significantly in 2009–2010, then picked in 2016–2017 (Figs. 4 and 5). The sex ratio of analysed fish sample was 1:1 (χ2 = 0.04, df = 1, p = 0.84), including 12 females and 12 males between 15.6–21.8 and 14.3–19.3 cm SL, respectively (Fig. 6). The length frequency distributions did not differ statistically between sexes (D = 0.33, p = 0.26), although females attained a slightly larger size than males. Males and females showed an isometric body growth, with no significant difference between sexes (Table 3). In this species, the spawning peak likely took place in June, when GSI of females increased at 14.1–20.0%.
Chaenocephalus aceratus
Chaenocephalus aceratus was the only channichthyid and the second most abundant species collected off the Argentine Islands. It was sampled all around the islands throughout all sampling periods over a wide depth range (Fig. 3). Both relative abundance and CPUE slowly decreased between 2006–2007 and 2009–2010, reaching a peak in 2016–2017 (Figs. 4 and 5). The analysed sample consisted of 65 females and 12 males ranging between 43.0–59.0 and 38.0–55.0 cm SL, respectively (Fig. 6). The length frequency distributions did not differ between sexes (D = 0.25, p = 0.28), whereas the sex ratio significantly departed from 1:1 (χ2 = 35.11, df = 1, p = 3.11 × 10–9). The length–weight relationships did not show significant differences in slopes (b) between sexes, although body growth of females was allometric negative (Table 3). Based on the GSI values (10.4–18.9%), four females (49.0–58.0 cm SL) caught in December and February were in spawning conditions, indicating this species as a summer spawner.
Parachaenichthys charcoti
This species was collected over a wide depth range (15–50 m) (Fig. 3) all around the Argentine Islands during the sampling years 2007–2008 and 2016–2017. The sample consisted of nine males and eleven females ranging between 40.0–47.0 cm and 40.5–49.5 cm SL, respectively (Fig. 6). The length frequency distributions were similar between sexes (D = 0.32, p = 0.35), and the sex ratio did not depart from 1:1 (χ2 = 0.05, df = 1, p = 0.82). About half of the specimens caught in October–November were close to spawning condition, with a GSI of 3.8–8.3% in males and 11.3–16.4% in females. Hence, the spawning season would occur from December onwards, in agreement with previous studies (Novillo et al. 2018).
Harpagifer antarcticus
Two females of 7.8–7.9 cm SL were caught in the Stella Creek Channel at 1–2.5 m (Fig. 3) in May 2016. One of them was in spawning condition, with a GSI of 21.8%.
Discussion
The present manuscript provides a general overview of the inshore fish community living off the Argentine Islands, complementing the previous data recorded in different sampling periods in the same area (Manilo et al. 2009; Veselskyy and Khoetskyy 2018). The species inventory of this area includes a total of 16 notothenioid fish, most of them described in the present study, besides Trematomus brachysoma (Pappenheim, 1912) and Trematomus pennellii Regan 1914. Other species previously found throughout a wide latitudinal gradient off the WAP (DeWitt and Hureau 1979; Daniels and Lipps 1982), such as Trematomus scotti (Boulenger, 1907) and Pagetopsis macropterus (Boulenger, 1907), are caught only offshore or are totally absent from the Argentine Islands, respectively. Trematomus peninsulae, an endemic benthopelagic species of the WAP (Daniels 1981), was caught in inshore waters of the Argentine Islands at about 50 m, which represents the upper limit of their depth range (Eastman 2017).
All the species collected in inshore waters off the Argentine Islands are demersal or benthic, with an overall high-Antarctic (T. peninsulae, N. coriiceps, T. borchgrevinki, T. bernacchii, T. brachysoma, T. hansoni, T. newnesi, T. pennellii) or low-Antarctic (G. gibberifrons, N. larseni, N. nudifrons, N. rossii, P. charcoti, C. aceratus, H. antarcticus) geographic distribution (Eastman 2020). Most of them are eurybathic, being distributed from shallow water down to 350–900 m. Conversely, T. borchgrevinki and T. brachysoma are found at a maximum depth of 70 and 90 m, respectively, whereas H. antarcticus lives in shallow waters down to 5 m depth (Eastman 2017). As a consequence, the inshore waters off the Argentine Islands represent the uppermost depth limit for most of the species caught. In addition, this area represents the southern boundary along the WAP for some low-Antarctic species, such as N. rossii, P. charcoti (although it was occasionally sampled off Peter I Island, Matallanas and Olaso 2007), C. aceratus and H. antarcticus. Despite of a possible biased sampling due to the exclusive use of fishing rods, the species composition of the inshore fish community of the Argentine Islands closely resembles those obtained in other neighbouring areas along the WAP using different fishing gears, such as the Danco coast (Casaux et al. 2003) and the Palmer Archipelago (Daniels and Lipps 1982).
Looking at the population structure of the most abundant species sampled off the Argentine Islands, some of them (Notothenia spp. and T. bernacchii) are represented by relatively small specimens (i.e. juveniles and sub-adults), with a few adults. The inshore waters with rocky bottoms and beds of macroalgae constitute a preferential habitat for these species, as reported elsewhere off the South Shetland Islands (e.g. Eastman et al. 2011) and Adelie Land (Hureau 1970). On the other hand, the finding of spawning adults of T. newnesi, P. charcoti and C. aceratus in different periods of the year (respectively in winter and summer) clearly indicates that this area is an important breeding site. The importance of inshore waters as spawning grounds has been previously reported for these and other notothenioids, such as N. nudifrons, C. aceratus and P. charcoti off Potter Cove, South Shetland islands (Novillo et al. 2019, 2021a). As reported here off the Argentine Islands, the spawning of N. coriiceps took place in May–June in Admiralty Bay, King George island (Sapota 1999). The capture of a single gravid female of H. antarcticus in May is consistent with the eggs release and nesting behaviour observed in June off Arthur Harbour (Anvers Island) by scuba divers (Daniels 1978), supporting an extended spawning period across their geographical distribution (Novillo et al. 2021b). Present data on T. peninsulae presumably extend the spawning period of this species from February (Daniels 1981) to May, as well as their maximum size at 22 cm TL (Eastman 2019).
The annual trend of relative abundance revealed a significant catch of Notothenia rossii starting from 2007–2008 (Manilo 2009), whereas catches of C. aceratus considerably increased in the most recent sampling years (present study; Veselskyy and Khoetskyy 2018). The concurrent increase of water temperature observed in recent years along the WAP might have potentially driven the shift of these typically sub-Antarctic species towards more southern areas of the Antarctic Peninsula than before. Therefore, it should be emphasized the importance to continuously monitor the species composition and relative abundance of fish communities in faunistic boundary areas like the Argentine Islands, to be able to assess ongoing latitudinal or depth distribution changes in advance.
Data availability
The datasets analysed during the current study are available from the corresponding author on reasonable request.
References
Artamonov Y, Romanov A, Vnukov Y, Lomakin P, Perov A, Stepura I (2003) Peculiarities of the hydrological and hydrochemical water structure in the region of Argentine Islands in February-March 2002. Ukranian Antarct J 1:17–24 ((in Russian))
Barnes DKA (1999) The influence of ice on polar nearshore benthos. J Mar Biol Assoc UK 79:401–407. https://doi.org/10.1017/S0025315498000514
Casaux R, Barrera-Oro E, Baroni A, Ramon A (2003) Ecology of inshore notothenioid fish from the Danco Coast, Antarctic Peninsula. Polar Biol 26:157–165. https://doi.org/10.1007/s00300-002-0463-y
Clarke A, Johnston IA (1996) Evolution and adaptive radiation of Antarctic fishes. Trends Ecol Evol 11:212–218. https://doi.org/10.1016/0169-5347(96)10029-X
Daniels RA (1978) Nesting behaviour of Harpagifer bispinis in Arthur Harbour, Antarctic Peninsula. J Fish Biol 12:465–474. https://doi.org/10.1111/j.1095-8649.1978.tb04190.x
Daniels RA (1981) Cryothenia peninsulae, a new genus and species of nototheniid fish from the Antarctic Peninsula. Copeia 1981:558–562. https://doi.org/10.2307/1444559
Daniels RA, Lipps JH (1982) Distribution and ecology of fishes of the Antarctic Peninsula. J Biogeogr 9:1–9. https://doi.org/10.2307/2844726
DeWitt HH, Hureau J-C (1979) Fishes collected during “Hero” Cruise 72–2 in the Palmer Archipelago, Antarctica, with the description of two new genera and three new species. Bull Mus Natl Hist Nat Paris 4(1A):775–820
Ducklow HW, Fraser WR, Meredith MP, Stammerjohn SE, Doney SC, Martinson DG, Sailley SF, Schofield OM, Steinberg DK, Venables HJ, Amsler CD (2013) West Antarctic Peninsula: An ice-dependent coastal marine ecosystem in transition. Oceanogr 26:190–2032. https://doi.org/10.5670/oceanog.2013.62
Eastman JT (2017) Bathymetric distributions of notothenioid fishes. Polar Biol 40:2077–2095. https://doi.org/10.1007/s00300-017-2128-x
Eastman JT (2019) An analysis of maximum body size and designation of size categories for notothenioid fishes. Polar Biol 42:1131–1145. https://doi.org/10.1007/s00300-019-02502-7
Eastman JT (2020) The buoyancy-based biotope axis of the evolutionary radiation of Antarctic cryonotothenioid fishes. Polar Biol 43:1217–1231. https://doi.org/10.1007/s00300-020-02702-6
Eastman JT, Eakin RR (2021) Checklist of the species of notothenioid fishes. Antarct Sci 33:273–280. https://doi.org/10.1017/S0954102020000632
Eastman JT, Barrera-Oro E, Moreira E (2011) Adaptive radiation at a low taxonomic level: divergence in buoyancy of the ecologically similar Antarctic fish Notothenia coriiceps and N. rossii. Mar Ecol Progr Ser 438:195–206. https://doi.org/10.3354/meps09287
Fischer W, Hureau JC (1985) FAO species identification sheet for fishery purposes. Southern Ocean (Fishing areas 48, 335 58 and 88) (CCAMLR Convention Area). FAO, Rome
Gon O, Heemstra PC (1990) Fishes of the Southern Ocean. JLB Smith Institute of Ichthyology, Grahamstown
Hofmann EE, Wiebe PH, Costa DP, Torres JJ (2004) An overview of the Southern Ocean Global Ocean Ecosystems Dynamics Program. Deep-Sea Res II 51:1921–1924. https://doi.org/10.1016/j.dsr2.2004.08.007
Huntley ME, Karl DM, Niiler PP, Holm-Hansen O (1991) Research on Antarctic coastal ecosystem rates (RACER): an interdisciplinary field experiment. Deep-Sea Res Part A 38:911–941. https://doi.org/10.1016/0198-0149(91)90090-3
Hureau J-C (1970) Biologie comparée de quelques poissons antarctiques (Nototheniidae). Bull Inst Oceanogr Monaco 68(1391):1–244
Jurajda P, Roche K, Sedláček I, Všetičková L (2016) Assemblage characteristics and diet of fish in the shallow coastal waters of James Ross Island, Antarctica. Polar Biol 39:2299–2309. https://doi.org/10.1007/s00300-016-1896-z
Manilo LG (2006) Ichthyofauna and morphobiological characteristics of mass fish species of coastal waters of Argentine Islands (Antarctica). Zbirnyk Prac Zool Muz (kyiv) 38:5–22 ((in Russian))
Manilo LG, Ignatiev SM, Chesalin MV (2009) The state of the coastal ichthyofauna and features of the biology of mass species of the fishes in the area of Ukrainian Antarctic station Akademik Vernadsky. Fish Ukr 64:7–23 ((in Russian))
Margalef R (1958) Information theory in ecology. Gen Syst 3:36–71
Matallanas J, Olaso I (2007) Fishes of the Bellingshausen sea and Peter I Island. Polar Biol 30:333–341. https://doi.org/10.1007/s00300-006-0189-3
Moreno CA, Zamorano JH, Duarte WE (1977) Distribución y segregación espacial de las poblaciónes de peces en bahía South (Isla Doumer, Antártica). INACH Ser Cient 5:45–58
Novillo M, Moreira E, Macchi G, Barrera-Oro E (2018) Reproductive biology in the Antarctic bathydraconid dragonfish Parachaenichthys charcoti. Polar Biol 41:2239–2248. https://doi.org/10.1007/s00300-018-2359-5
Novillo M, Moreira E, Macchi G, Barrera-Oro E (2019) Reproductive effort in Chaenocephalus aceratus validated by gonadal histology: inshore sites serve as spawning grounds for some notothenioids. Polar Biol 42:1959–1972. https://doi.org/10.1007/s00300-019-02571-8
Novillo M, Elisio M, Moreira E, Macchi G, Barrera-Oro E (2021a) New insights into reproductive physiology in Antarctic fish: a trial in Lepidonotothen nudifrons. Polar Biol 44:1127–1139. https://doi.org/10.1007/s00300-021-02879-4
Novillo M, Moreira E, Macchi G, Barrera-Oro E (2021b) Histological analysis provides further insights into Harpagifer antarcticus reproductive biology at the western Antarctic Peninsula. Polar Biol 44:2165–2175. https://doi.org/10.1007/s00300-021-02953-x
Novillo M, Desvignes T, Moreira E, Barrera-Oro E (2022) Egg predation in Antarctic fish: the ingestion by Notothenia coriiceps of an entire Trematomus bernacchii spawn identified by molecular techniques. Estuar Coast Shelf Sci 266:107742. https://doi.org/10.1016/j.ecss.2022.107742
Pielou EC (1966) Shannon’s formula as a measure of specific diversity: its use and misuse. Am Nat 100:463–465
Ruhl HA, Hastings PA, Zarubick LA, Jensen RM, Zdzitowiecki K (2003) Fish populations of Port Foster, Deception Island, Antarctica and vicinity. Deep-Sea Res II 50:1843–1858. https://doi.org/10.1016/S0967-0645(03)00094-8
Sapota MR (1999) Gonad development and embryogenesis of Notothenia coriiceps from South Shetlands—Antarctica. Polar Biol 22:164–168. https://doi.org/10.1007/s003000050406
Shannon CE, Weaver W (1949) The mathematical theory of communication. University of Illinois Press, Urbana
Smith RC, Baker KS, Fraser WR, Hofmann EE, Karl DM, Klinck JM, Quetin LB, Prézelin BB, Ross RM, Trivelpiece WZ, Vernet M (1995) The Palmer LTER: a long-term ecological research program at Palmer Station, Antarctica. Oceanography 8:77–86
Smith RC, Baker KS, Stammerjohn SE (1998) Exploring sea ice indexes for polar ecosystem studies. Bioscience 48:83–93. https://doi.org/10.2307/1313133
Sokal RR, Rohlf FJ (1995) Biometry: the principle and practice of statistics in biology research. Freeman, New York
Veselskyy MV, Khoetskyy PB (2018) Ichthyofauna of the waters of the archipelago of the Argentine Islands (Ukrainian Antarctic Expedition 2015–2016). Nauk Zap Ternop Nat PedUniv 72:36–43
Zinkovskyi AV, Dykyy IV, Trokhymets VM (2021) Allocation of the diet of the Argentine Islands’ inshore ichthyofauna. Biosyst Divers 29:67–72
Acknowledgements
The authors are grateful to the National Antarctic Scientific Center of Ukraine for financing and organizing the collection and transport of the samples, as well as for the full support to the study. We are much indebted to the Institute of Biology and Medicine of Taras Shevchenko National University of Kyiv and to the Institute of Hydrobiology of the National Academy of Sciences of Ukraine for providing all the laboratory facilities. The corresponding author would like to dedicate the paper to all Ukrainian people.
Author information
Authors and Affiliations
Contributions
Each author provided the following contributions: VT conceived the study, MLM wrote the manuscript, OS, AZ, OG, ID, DL and AB conducted field and lab work. All authors contributed to the manuscript editing and approved the final draft.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare they have no conflicts of interest (both financial or non-financial) relevant to the content of this article, and that the study was conducted in accordance with the ethical standards of their own institutional and national research committee.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Trokhymets, V., Savytskiy, O., Zinkovskyi, A. et al. Species composition, distribution and relative abundance of the inshore fish community off the Argentine Islands, Bellingshausen Sea. Polar Biol 45, 845–855 (2022). https://doi.org/10.1007/s00300-022-03040-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-022-03040-5