
Abstract
Arctic birds migrating southwards face a multitude of challenges such as habitat loss, pollution, overexploitation of food resources and climate change impacts. Long-tailed Duck winter populations in the Baltic Sea have declined in recent decades. However, precise spatial data are lacking, especially from males. Thus, we aimed to identify the wintering grounds, timing of migration and stopover sites of males and females. We studied spatiotemporal distribution patterns of eight male and five female Long-tailed Ducks using implanted ARGOS satellite transmitters. Birds were tagged in the breeding season on Kolguev Island, Russia. After the breeding period, Long-tailed Ducks from Kolguev used three main post-breeding areas: most males undertook long distance eastward post-breeding migration to areas around the Yamal and Gydan peninsulas, while one male and four females moved short to the southeast coast of Kolguev, and one female moved to Mezhdusharsky Island in the Novaya Zemlya Archipelago. Autumn migration included stopover sites in the White Sea, Lake Ladoga, the Gulf of Finland and the Gulf of Riga, and all birds except one spent the winter in the southern part of the Baltic Sea, from the Pomeranian coast and Hoburgs bank to the Gulf of Finland. Only one female stayed in the White Sea for the winter. All but one bird used the White Sea as a stopover site in May, suggesting that this area is of special importance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Animals carry out regular annual migration movements in response to temporal fluctuations in resource availability during the annual cycle (Alerstam et al. 2003; Dingle and Drake 2007). Seasonal environments often provide rich foraging grounds for parts of the year but may become inhospitable at other times. The high Arctic tundra is such a habitat, where the ice-free season is short, and meltwater feeds numerous freshwater lakes (e.g. Smith et al. 2007), in which an intense growth and reproduction of invertebrates takes place. The Arctic thus provides a short but very rich food peak to birds that exploit these habitats. However, during most of the year, the lakes are frozen and thus, the food is not accessible, forcing animals to migrate to wintering sites at lower latitudes. Migration multiplies the threats that animals face as they rely on different sites across the annual cycle (Wilcove and Wikelski 2008; Vickery et al. 2014).
Sea ducks are among the migrant birds that move between breeding areas in the Arctic tundra, and mostly marine wintering areas at lower latitudes. Like many migratory birds, mammals, fish, and invertebrates that experienced population declines in recent decades (Wilcove and Wikelski 2008), this is also the case with many sea duck populations (Austin et al. 2014). In sea ducks, climate change has led to alterations of both the breeding and wintering grounds: As the Arctic warms faster than other latitudes, ecosystem changes affect biological communities, biodiversity, and aquatic food web structure and possibly, the breeding success of sea ducks that depend on the aquatic food web for reproduction (Wrona et al. 2006; Fox et al. 2015). Outside the breeding season, sea ducks move to marine and aquatic habitats, where they spend the winter and energetically demanding periods such as moult (Savard and Petersen 2015). Here, they face a multitude of challenges such as habitat loss, pollution, overexploitation of food resources, and climate change impacts. However, for many populations, key sites in their annual cycle are not known. Better knowledge of annual movements and important sites can help to better understand the drivers behind population declines.
Long-tailed Ducks Clangula hyemalis, have a circumpolar distribution (North America, Greenland, Iceland, Fennoscandia, and Russia) and breed in tundra habitats of the high Arctic and the arctic-alpine zone. The breeding biology in the Russian part of its range has been reviewed recently (Loshchagina et al. 2019; Sokolov et al. 2019). Across the breeding range, Long-tailed Ducks breed from late May, and most males leave the females soon after the start of incubation, in late June or July to gather in moulting flocks. While the males of some populations carry out extensive post-breeding migrations for moulting sites, others moult on waters near the breeding grounds. Females mostly moult in August and September on or close to the breeding grounds (Petersen and Savard 2015; Savard and Petersen 2015). After the post-breeding moult, both males and females migrate south and winter at sea, generally far offshore in waters 10–35 m deep, as well as in brackish estuarine waters and lagoons or more rarely, on inland lakes (del Hoyo et al. 1992).
The diet in the breeding season consists of freshwater plankton and benthos, such as crustaceans, molluscs and insect larvae. In the non-breeding period, the diet consists of various benthic invertebrates and fish usually caught at depths of 5–25 m (del Hoyo et al. 1992; Žydelis and Richman 2015).
In the present study, we followed the migration of Long-tailed Ducks, a species which has undergone declines in Europe (Skov et al. 2011) and North America (Dickson and Gilchrist 2002; Bowman et al. 2015). The Long-tailed Duck winter population in the Baltic Sea, where most of the western Siberian and northern European populations winter, has declined by 65% between 1992 and 1993 (c.4,272,000 individuals) and 2007–2009 (c.1,486,000 individuals; Skov et al. 2011; Heldbjerg et al. 2020). A variety of environmental pressures both in the breeding and non-breeding areas could have contributed to these declines (Hearn et al. 2015). Possible drivers of the decline include unsustainable mortality levels in the non-breeding areas from gillnet fisheries bycatch (Bellebaum et al. 2013; Žydelis et al. 2013), as well as a severe decline in breeding success in recent years (Hearn et al. 2015). The latter is not fully understood, and further research will need to target both changes in Arctic breeding ecosystems and possible carry-over effects from changes in food availability in the wintering area (Hario et al. 2009; Skabeikis et al. 2019).
The Long-tailed Duck population breeding in western Siberia and northern Europe is the most numerous population (1.6 Mio. birds) compared to the populations in Iceland and Greenland (100,000–150,000), eastern Siberia (0.5–1 Mio.) and North America (1 Mio., Wetlands International 2021). Approximately 90% of this Long-tailed Duck population winter in the Baltic Sea (Skov et al. 2011). In a recent tracking study using light-level geolocators, 18 of 19 female Long-tailed Ducks breeding at Kolguev Island in the south-eastern Barents Sea spent their winter in the Baltic Sea and one bird in the White Sea (Karwinkel et al. 2020). Females were selected for that study because they are faithful to the breeding ground and can be recaptured to remove data loggers. This is not the case in males. Long-tailed Ducks usually pair in the wintering grounds and on spring migration. Subsequently, males follow their partner to the breeding area (Glutz and Bauer 1992). Therefore, a different method not requiring recapture had to be employed in the present study focussing on a comparison of male and female Long-tailed Ducks. In Long-tailed Ducks, males usually leave the breeding sites soon after the start of incubation. Therefore, males and females are expected to differ in their annual movements, and this could have important implications in terms of understanding population connectivity and trends.
We thus studied the distribution patterns of Long-tailed Ducks using implanted ARGOS satellite transmitters. The birds were tagged on the breeding grounds of Kolguev Island, Russia, and followed throughout the annual cycle. We aimed to identify the moult and winter distribution, as these two seasons present important periods in the annual cycle that may determine survival and breeding propensity. Specifically, we aimed to analyse the core areas used during moult and winter as well as important sites along the flyway for male Long-tailed Ducks and compare them to females.
Methods
Study site and fieldwork
Kolguev Island (69.16°N, 48.90°E) in the Barents Sea is part of the Nenets Autonomous District. This large island (5020 km2) is located 72 km off the mainland across the Pomor Strait and is dominated by a slightly hummocky moss-lichen tundra with small meltwater lakes in the centre of the island (Kruckenberg et al. 2012). Most of the lakes on the island are shallow thermokarst lakes that freeze to the bottom in winter. There are also deeper glacial lakes with sandy shores in the higher part of the island (Koreisha 2000). The Long-tailed Duck is a common breeding bird here (Glazov et al. 2021). During the breeding season 2019, 14 Long-tailed Ducks (five females and nine males) were equipped with ARGOS satellite transmitters (platform transmitter terminals, PTTs). The birds were captured in the interior of Kolguev Island in June 2019 (Online Resource 1: Table S1) with mist nets erected between small lakes and gillnets located in the tundra lakes with constant monitoring. Individuals were selected for transmitter implantation based on body mass, good health, and the condition that transmitter represents max. ca. 5% of body mass. This transmitter mass is at the upper end of transmitters used for birds, as Phillips et al. (2003) indicated that a transmitter mass equivalent to 3% of body mass is the tipping point for increased trip durations and/or nest desertions. A recent analysis of implanted transmitters (Lamb et al. 2020) has shown an effect of similar implants on the timing of movements in Long-tailed Ducks shortly after the implantation (carried out in winter), affecting the first migration (i.e. spring migration in case of the winter deployed birds), resulting in a later and faster migration. However, the timing and duration of autumn migrations were not affected. In our study, implant surgery was carried out in the breeding season and thus, may have affected the first migration (post-breeding migration), and possibly the autumn migration while effects would be less likely on the subsequent spring migration.
Transmitters were implanted in the abdominal cavity under anaesthesia by an experienced wildlife veterinarian (Korschgen et al. 1996; Spiegel et al. 2017). For some females the transmitters represented up to 5.4% of body weight, as they have smaller body size than males. We used implantable PTTs weighing 26 g, manufactured by Microwave Telemetry, Inc. (Maryland, USA). Transmitters were potted for pressure resistance and programmed using duty cycles of 3–6 transmission hours and 72 h intervals between transmissions.
We documented post-implantation behaviour. As all captured birds were territorial and had their own territories, they would intend to get back to their breeding ponds as soon as they could. Territorial behaviour was determined by observation, when males defended females from other males or females in the pond or pond system. Females were not aggressive, but they did not move to other lakes. After implantation, all birds were released 1–2 km away from capture place to a pond (recovery pond) close to the surgery location with no territorial Long-tailed Ducks. To determine the duration of altered behaviour following instrumentation, we monitored the “recovery pond” 2–3 times a day to record behaviour and presence of birds in the pond. Behaviour of post release birds was variable: after implantation, birds tended to not fly, but rest on shore of the pond, and actively preen. Also passive feeding was observed. We determined the exact location of each instrumented bird according to their different plumage characteristics and antennas of PTT. Furthermore, we considered that a bird was exhibiting “normal” behaviour, when it was observed actively feeding, diving for food and spent most of its time in water or at the shore of the capture pond and behaving territorial and/or aggressive towards conspecifics. Survival was monitored during the 14 days period post surgery (Esler et al. 2000).
Definition of terms and seasons:
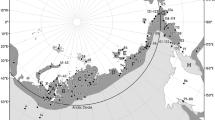
We characterised six periods within the annual cycle of the species, based on longitudinal information (Karwinkel et al. 2020, Fig. 1, Online Resource 2: Fig. S1).

Overview and stages in the annual cycle. Overview map and example of migration pattern of a male Long-tailed Duck from Kolguev Island, Russia. Example for the assignment of the breeding season phases of tracked Long-tailed Ducks, according to longitudinal data
The breeding stage was defined as the time in which the transmitter data from each instrumented bird did not show a change in longitudinal values. The post-breeding migration was the first directional movement away from the breeding site to a post-breeding staging site. This ended when the bird started autumn migration, which was defined by a rapid change in longitudinal values, and a movement past Kolguev into more westerly waters. Autumn stopovers were defined as periods of at least 6 h without longitudinal changes within the migration stage. Autumn migration stopped and the wintering stage began when data on longitudes remained constant again. Subsequently, the onset of spring migration was noted as a further rapid change in longitudinal values and defined the end of the wintering stage. Spring stopovers were defined as periods of at least 6 h without longitudinal changes within the migration stage. Spring migration stopped when longitudinal values showed constancy again. The period following the spring migration was defined as second breeding stage. For all stages, we manually assigned the start and end dates.
Arrival and stopover times may be affected by gaps in the data due to transmission cycles. Since gaps were 72 h long, this method may detect migratory departures or arrivals with a 1–3-day delay, and the duration of short stopovers would be especially affected. If these are only recorded in a single transmission cycle, a range of stopover durations would be possible due to the 72 h intervals between transmissions. We thus provided the observed stopover durations, as well as maximum stopover durations.
Data analyses
Data were filtered to reduce noise produced by location fixes with low or unknown accuracy. Filtering was conducted in two steps. First, the Douglas filter in Movebank was applied (see https://www.movebank.org/cms/movebank-content/argos-data-filters). Secondly, any remaining obvious outliers (n = 2) were removed from the dataset.
After filtering, 8,121 positions were left for further analysis, of which 20% (n = 1652) were categorised in the high-quality location classes 3–1 (Online Resource 1: Table S2). From one transmitter (ES033725) we only received positions from the vicinity of the breeding area for about one month (Online Resource 1: Table S1 and S2). This bird was therefore not included in the analyses. To homogenise the data, mean daily positions were calculated and used in further analyses.
We used ArcGISv.10.2.2. (ESRI 2012) to map migratory patterns and longitude plots to quantify migration movements. Autumn migration was calculated from the first day of departure from the post-breeding site to the first day at the autumn stopover site. For mapping of migratory pathways, we used all positions while birds were on migration, and to characterise the use of stopover sites, we selected the first position after arrival and the last position before departure, respectively, in the corresponding area.
We used a North Pole Azimuthal Equidistant projection to calculate distances. Migration distances are given as minimum estimates assuming straight flight paths between consecutive ARGOS positions. We calculated distances to breeding and wintering sites by summing the length of vectors created from point to point of the PTT transmissions between the first day of departure from the wintering or spring stopover site and the first day at the breeding site.
To estimate home ranges and core areas, we calculated 95% and 50% kernel densities using the adehabitatHR package v0.4.18 (Calenge 2020) in R. We estimated 95% and 50% kernel densities for post-breeding and winter areas with ad hoc adjustments of bandwidth, in order to meet the criteria of a contiguous rather than disjoint polygon that contained no lacuna within the home range (Kie 2013). Additionally, 95% and 50% kernel densities with href bandwidth and MCP are provided (Online Resource 1: Table S3). We used only data from individuals that covered a full season (arrival date until departure date) and only one position per day to avoid overrepresentation of some intervals. We used the 50% kernel density contour to identify core habitat use. To calculate sizes of home ranges and core areas in km2 we projected the shapefile into North Pole Lambert Equal Area projection.
Results
Post-surgery phase in the breeding area
None of the instrumented ducks were lost during the 14 days period post surgery. One male was predated by a Glaucous gull Larus hyperboreus on the 29th day post-surgery. Transmitter masses varied from 4.0% to 5.4% of body mass, with average of 4.7 ± 0.3% and 5.2 ± 0.2% for males and females, respectively. Instrumented birds spent from 2 h to 54 h after surgery in the recovery pond, and this time did not differ between males (on average 32 ± 14 h) and females (34 ± 19 h). The number of days after surgery when birds returned to their breeding ponds and started to behave normally ranged from one day to three days, with an average of 1.8 days in both males and females (Online Resource 1: Table S4).
Data output
We obtained data from 13 Long-tailed Ducks to analyse, including five females and eight males. Of these, we received PTTs transmission for 104–408 days (females: 114–403 days, males 104–408 days). Males and females did not differ in median PTT transmission duration (females: 369 days, males 356 days, Kruskal–Wallis chi-square test: χ2 = 0.086, df = 1, p = 0.770). All 13 birds reached the post-breeding or moult sites, but transmissions stopped for one male in September, at the northern coast of the Yamal Peninsula (ES033717). Autumn migration was thus observed for 12 animals (five females and seven males). The transmissions of three birds stopped in October 2019, shortly after their autumn migration was concluded, in Lake Ladoga (female ES033718) and the Gulf of Finland (female ES030061 and male ES033719). Three females and six males transmitted after reaching the winter sites. Of these, five males and three females transmitted throughout the winter, while one male (ES033715) transmitted in February 2020 for the last time. Of the remaining five males and three females, one PTT failed at the end of the spring migration (male ES033721), and thus, four males and three females reached a second breeding stage in May/June 2020 (Online Resource 1: Table S1).
Post-breeding migration and moult
Male Long-tailed Ducks started their movements away from Kolguev Island in late June/early July, before the beginning of flight feather moult. Most of them performed a long distance eastward migration to their post-breeding and moulting sites (Fig. 2, Table 1). Females left the breeding area significantly later than males and moved a shorter distance to their nearby post-breeding staging areas (Fig. 2, Table 1). All males except one left the breeding area between 30 June and 16 July 2019, while the females stayed in the breeding area either until late July/early August (n = 2), leaving Kolguev just before moulting period, or until early September (n = 3), when wing feather moult is completed (Table 1).

Post-breeding stage. Areas used by Long-tailed Ducks from Kolguev Island, Russia, during the post-breeding stage: a Overview map with positions of males (blue) and females (red), b Post-breeding home ranges of four females and one male on the coast of Kolguev Island, c Post-breeding home ranges of one female on the coast of Mezhdusharsky Island (Novaya Zemlya), and d post-breeding home ranges of seven males close to the Yamal and Gydan peninsulas. (Color figure online)
We observed three main post-breeding areas: All except one male headed east for post-breeding/moulting areas around the Yamal and Gydan peninsulas, which they reached after 8–28 days (Table 1, Fig. 2). One male (ES033722) and most females stayed in freshwater lakes of Kolguev for moult and then moved to the southeast coast of Kolguev, arriving within 1–3 days. Finally, one female moved to Mezhdusharsky Island in the Novaya Zemlya Archipelago (Fig. 2). The distance from the breeding site to the post-breeding sites at the southeast coast of Kolguev was 36–46 km, to Mezhdusharsky Island ca. 260 km and to post-breeding areas around the Yamal and Gydan peninsulas 960–1300 km (Table 1).
Accordingly, the duration of the post-breeding migration was also shorter in females that moved to nearby sites than in males that moved further away (Table 1). The duration of post-breeding staging was 14–77 days (median 41 days) for females, with the two females that moved early from the breeding area reaching the longest stay (64 and 77 days), compared to the females that moved in September (14–41 days). The duration of post-breeding staging amounted to 41–87 days (median 75 days) for males. The shortest staging time in males was observed in male ES033722 that stayed 41 days at the southeast coast of Kolguev. The birds that moved to post-breeding areas around the Yamal and Gydan peninsulas were more mobile and had much larger core areas and home ranges than birds that stayed close to Kolguev (Table 1). This especially applied for two males (ES033713 and ES033727) that spent the first part of the post-breeding period on the coasts of the Gydan Peninsula or nearby islands, but then moved to the western coast of the Yamal Peninsula on 21 and 26 September, respectively (Fig. 2).
Autumn migration and autumn stopovers
Though females Long-tailed Ducks arrived later at their post-breeding sites, the shorter duration compensated for this, and the timing of the onset of autumn migration was similar in males and females, ranging from 24 September to 13 October (Table 2). Male Long-tailed Ducks had longer autumn migrations (i.e. the track from the post-breeding area to the autumn stopover sites) than females but reached similar autumn stopover sites at the same time as females (Table 2). During autumn migration, females travelled 620–1946 km and males 1500–2820 km (Table 2). The travel speeds varied in the range of 148–649 km per day.
Short autumn stopovers were recorded in one female, remaining stationary for one (max. five) days, and two males with stopover durations of three (max. seven) days (Table 2). The remaining females stayed in a stopover area for an observed 9–15 days, and the males for 13–17 days (for maximum times see Table 2), except for ES033722, that spent 54 days in the eastern Gulf of Finland, before moving to the western Gulf of Finland in December.
The autumn stopover sites (Fig. 3) were distributed over the White Sea (2 females), Lake Ladoga (1 female), the Gulf of Finland (1 female and 4 males) and the Gulf of Riga with its offshore islands (1 female and 4 males). Most of the birds were relatively stationary, except for male ES033713 that visited three different areas within 17 days (Fig. 3). Males and females did not differ in mean longitudes during the autumn stopovers (Wilcoxon rank sum test, W = 15, p = 0.594). On her way to Lake Ladoga, one female (ES033718) transmitted intermediate positions from the White Sea on 27/09/2019, but this short stationary period of two hours was not classified as a stopover.

Autumn stopovers. Areas used by Long-tailed Ducks from Kolguev Island, Russia, during autumn stopover: a Overview map with positions of males (blue) and females (red), b Stopover positions at Lake Ladoga and in the Baltic Sea, and c Stopover positions in the White Sea. Males are marked with triangles, and females with filled circles. (Color figure online)
Wintering
Long-tailed Ducks from Kolguev spent the winter in the White Sea (one female), the Gulf of Finland (one female and one male), the Gulf of Riga (one male), off the south-eastern coast of the island of Gotland, Sweden (one female and three males) and the coasts of Eastern Germany and Poland (two males) (Fig. 4, Table 3).

Winter areas. Areas used by Long-tailed Ducks from Kolguev Island, Russia, during winter: a Overview map with home ranges of males (blue lines) and females (red lines), and core area of female ES033724 in the White Sea (red area), b Home ranges (lines) and core areas (filled areas) of Baltic Sea winter distributions. (Color figure online)
The wintering stage lasted for 135 to 206 days (4.5 to 6.8 months, without autumn or spring stopovers), and this duration did not differ between males and females (Table 3). The total wintering time (including autumn and spring stopovers and thus, the period when Long-tailed Ducks are affected by challenges in the wintering habitat) was much less variable. It ranged from 206 to 219 days (6.8 to 7.2 months) and was similar in females (206–212 days) and males (206–219 days, Wilcoxon rank sum test, W = 7, p = 1).
Individual Long-tailed Ducks used relatively restricted wintering sites with core areas of 688 km2 in the White Sea and 1014–2625 km2 in the Baltic Sea (Table 3). The distance between first and last winter positions ranged from 8.8 km to 190 km (Table 3, Online Resource 2: Fig. S2). The largest distance and core area were recorded by male ES033721 that switched repeatedly between offshore banks off Gotland and the coast of Poland (Online Resource 2: Fig. S2).
Spring stopovers, spring migration and breeding sites in the second year
The time of the onset of spring migration varied between 25 March and 20 May (Table 3). Long-tailed Ducks that spent winter in the central and southern parts of the Baltic Sea started their movements earlier (between 25 March and 19 April) than birds overwintered in the Gulf of Riga and the Gulf of Finland, which left their wintering sites only in May (Table 3). Birds that initiated their movements earlier shifted to a different site inside the Baltic Sea, where they stayed for more than one week (median: 18 days), which was considered a stopover. Long-tailed Ducks that started considerable movements in May, quickly left the Baltic Sea for their stopover sites in the White Sea. Therefore, while all Long-tailed Ducks made spring stopovers, we recorded two kinds of them: a long stopover within the Baltic Sea north-easterly from the wintering sites, and a (probably shorter) stopover in the White Sea after mid-May (Table 4, Fig. 5). As ARGOS data transmissions become less frequent towards the end of the battery life, infrequent recording was apparent in the data towards spring. Thus, we could not precisely estimate spring stopover durations, especially in May. We therefore calculated minimum and maximum spring stopover durations as days between the previous recorded position and the first breeding time position (Table 4).

Spring staging. Areas used by Long-tailed Ducks from Kolguev Island, Russia, during spring staging (April–May 2020): a Overview map with positions of males (blue) and females (red), b Positions in the Baltic Sea, c Positions in the White Sea. Males are marked with triangles, and females with filled circles. Long stopovers (> 1 week) are marked with large symbols, and short stopovers (observed duration 1–2 days) with smaller symbols in (b) and (c). (Color figure online)
Long spring stopovers (> 1 week) were observed in one female and four of five males (Table 4). Compared to the winter area, all these individuals had a more easterly longitude and more northerly latitude during their spring stopovers (Table 4). Three males and one female moved in north-easterly direction within the Baltic Sea and reached the Stockholm and Åland archipelagos, or the Gulf of Finland and the coasts of Estonia and Latvia in the South-eastern Baltic Sea (Fig. 5). All three females and four of five males also made stopovers in the White Sea in May; however, the duration of them could not be accurately estimated, but at least one female and one male stayed there more than a week (Table 4, Fig. 5).
All birds left the Baltic Sea between 7 and 23 May, with birds arriving at the breeding sites in late May to mid-June (Table 4). Three females and four males were tracked until arrival to the breeding sites (Table 4). All females returned to Kolguev Island for the second breeding season, as did two males (Table 4, Online Resource 2: Fig. S3). Of these returning males, ES033722 remained in coastal waters until late July, while only two fixes were obtained for ES033727 until 15 June. The remaining two males went to Vaygach Island (70.128°N, 58.781°E) and the Kola Peninsula (68.448°N, 36.555°E), where they stayed inland until the end of June. Another male (ES033721) was last recorded near Vaygach on 21 May (70.151°N, 58.557°E), possibly underway or close to a breeding site. Three males (ES033713, ES033722, ES033727) could be tracked until the post-breeding period again in 2020, when they all showed the same behaviour as in 2019.
Discussion
Our study of Long-tailed Duck movements between the breeding area on Kolguev Island and the wintering areas shows similarities between individual males in their eastward post-breeding migration, and between birds of both sexes in choosing the White Sea as a spring stopover site.
Post-surgery behaviour
For Long-tailed Ducks during breeding season on Kolguev, it took two days to return to normal behaviour following implantation, which was three days shorter than described by Lamb et al. (2020) and 12 days shorter than in the work by Esler et al. (2000). In studies of Esler et al. (2000) and Lamb et al. (2020) most of the sea ducks (various species) were captured and implanted during wintering or moult period at open sea, while in this study we captured birds in breeding area. In this study we had the possibility to make direct observation on the impact of the surgery on sea ducks, while in Lamb et al (2020) the behaviour was calculated form transmitters data not seeing individual birds. Our method can therefore give more precise evaluation of the behaviour of individual birds. An additional factor of such fast return to the breeding ponds can be the need to defend the nesting site. However, none of the females with implants or non implanted birds had been observed incubating or rearing broods. We think that the most important factors of positive shorter time to recovery could be that the birds had enough time to rest and a good source of easily accessible food after surgery. During the fieldwork we observed birds in the first two days after surgery sitting on the bank of the pond preening. Also, after just two days, males would actively chase other males in flight. Considering that birds were instrumented at the breeding site, this could effectively increase survival compared with birds implanted on wintering grounds (Sea Duck Joint Venture 2015) where higher mortality and effect of implantation were registered.
Moulting
When moulting their flight feathers in late summer, sea ducks are flightless for 4–5 weeks and thus depend on rich food resources and protection from predators (Savard and Petersen 2015). While females with chicks usually moult at their breeding site, males and non-breeders of many species tend to aggregate in large numbers in preferred moulting sites. Such aggregations may occur somewhere along the route between breeding and wintering grounds, or far away from these routes (Savard and Petersen 2015).
Moulting site choice in Long-tailed Ducks is highly variable. Unless attending broods, birds of both sexes moult their flight feathers in shallow coastal waters, lagoons, or lakes. Moulting flocks are found throughout the breeding range and may include thousands of birds (Petersen and Savard 2015; Savard and Petersen 2015). Such flocks are also found in many coastal waters along the Russian Arctic coast from the Barents Sea to Eastern Siberia (Loshchagina et al. 2019; Sokolov et al. 2019).
Until mid-August all except one male from Kolguev moved > 900 km to the east to moulting sites at the coasts of the Northern Yamal and Gydan peninsulas. These birds were probably taking advantage of food-rich shallow waters around the Ob and Yenisei estuaries with benthic communities dominated by polychaetes, isopods and bivalves (Kozlovskiy et al. 2011; Vedenin et al. 2015; Galkin and Vedenin 2015). Particularly large moulting concentrations were already known to occur in this region (Sokolov et al. 2019), but the range of origin of the birds was unknown. Post-breeding migrations over even longer distances from breeding areas in Siberia to Alaska have previously been confirmed by recoveries of ringed Long-tailed Ducks, particularly males (King 1973).
Unlike males, all but one females stayed at Kolguev during and after flight feather moult. Three of them moulted on inland lakes, and one female moved to the coastal lagoon in the eastern part of the island for moult. Only one female (ES033714) left Kolguev and moved eastwards for moult, but not as far as the males. From a sample of 49 other females from Kolguev equipped with light-level geolocators in 2017 and 2018, only five birds left Kolguev and the surrounding Barents Sea after wing feather moult. During the subsequent body feather moult until October, these birds probably stayed in an area around Novaya Zemlya Archipelago (Karwinkel et al. 2020 and unpublished data), thus confirming that in the study population, females usually moult on or close to Kolguev and only males undertake post-breeding/moult migrations eastwards as far as the Yamal and Gydan peninsulas. After completion of the flight feather moult, birds that moulted on freshwater lakes inland of Kolguev performed their post-breeding migration to the coastal waters of Kolguev and stayed there until October. Apparently, this post-breeding staging period is important for birds to accumulate resources for subsequent autumn migration (Leafloor et al. 1996).
Tracking male Long-tailed Ducks moulting in coastal lagoons in Alaska with radio transmitters for 2–3 weeks, Flint et al. (2016) found home ranges of only 10–20 km2, like the core areas in coastal waters of Kolguev and Novaya Zemlya. Males moulting off the Northern Yamal and Gydan peninsulas used much larger post-breeding home ranges and core areas, which were approximately 50–67% as large as during winter (Tables 1, 3). This difference matches the average duration of the respective periods (75 vs. 166 days, Tables 1, 3). Flightless time is estimated at 26 days for Long-tailed Ducks (Savard and Petersen 2015). In our study (Table 1), the post-breeding stage of all males lasted much longer than the flightless time. It is possible that these males occupied much smaller home ranges during actual wing feather moult, and afterwards showed a spatial behaviour like in the winter period.
Wintering
Long-tailed Ducks from the East Atlantic Flyway population predominantly winter in the Baltic Sea, where they occupy shallow coastal waters and offshore banks often in high densities (Skov et al. 2011; Nilsson 2016). Five males and one female tracked throughout winter spent the winter season in these high-density areas, with some birds moving between different offshore banks or coastal areas. Of the remaining, two birds wintered at the southern coast of Finland, where only low densities of Long-tailed Ducks were recorded in the early 1990s (Durinck et al. 1993). At least one other female from Kolguev spent the winter 2017–2018 in this area (Karwinkel et al. 2020).
Small proportions of the flyway population winter outside the Baltic in coastal waters off Norway and the Kola Peninsula (Heggøy and Shimmings 2016), and a few birds stay in ice-free parts of the White Sea (Loshchagina et al. 2019; Karwinkel et al. 2020). Our present study is the second to find one female from Kolguev wintering in the White Sea, the previous being Karwinkel et al. (2020). While both studies together still form a comparatively small sample, it is notable that several birds were found wintering north-east of the core wintering range, in the Gulf of Finland and the White Sea. This might be explained by a recent northward shift of winter distribution due to increasing water temperatures, as already observed in other species wintering in the Baltic (Lehikoinen et al. 2013). In Steller’s eiders Polysticta stelleri, the winter distribution has shifted eastwards along the Kola Peninsula and into the White Sea as more areas has become ice-free during winter (Aarvak et al. 2013). Monitoring of off-shore wintering Long-tailed Ducks in the Baltic Sea is also challenging (Fox et al. 2019). Future surveys of wintering sea ducks should therefore pay more attention to ice-free areas outside the main wintering areas. Surveys ignoring these more northern wintering areas off Finland, Norway, and Russia will risk missing out an increasingly important part of the total wintering population as global warming continues.
Spring migration
After wintering in the central and southern Baltic Sea, four birds moved north or north-east inside the Baltic Sea to an extended spring stopover (Table 4). Three birds wintering in Riga Bay, or the Gulf of Finland showed no such movements. All birds occupied shallow coastal waters, broadly overlapping with traditional herring Clupea harengus spring spawning sites in archipelagos and bays (Hede Jørgensen et al. 2005a, b). The onset of the spawning season varies with water temperature, first spawning activities in the Archipelago Sea occur in late April in mild springs (Rajasilta et al. 1993; Polte et al. 2021). Spring stopovers in the Baltic Sea would thus probably match the early phase of Herring spawning in most years. Fish eggs provide a protein rich seasonal food source frequently exploited by staging sea ducks, including Long-tailed Ducks (e.g. Žydelis and Ruskyte 2005; Lok et al. 2012).
All but one bird made stopovers in the White Sea, which was therefore the most predictable stopover site in this study. The longest stopover and also the latest departure was observed in a male (ES033722) which probably was not paired and may have tried to acquire a mate in this area.
Our data suggest that the typically long stopovers at different sites inside the Baltic may serve to accumulate body reserves, utilising seasonal food such as herring spawn. The White Sea may be a crucial location for most birds for a shorter stopover in early May. The extended stay of one probably unmated male suggests that some late matings might occur here, too. However, due to the limited life cycle of the implanted transmitters, other methods such as GLS with saltwater loggers would be better suited to obtain stopover durations more accurately for this part of the season.
Food intake during spring migration may affect the condition of the birds upon arrival at the breeding sites and thus the ability to produce eggs and incubate in time during the short Arctic spring. Warming water temperatures already impact timing and amount of herring reproduction in the Baltic (Polte et al. 2021) and may affect the food resources in the White Sea as well. The possible impacts of such changes on the breeding performance of sea ducks will be important to study in future.
Second summer
Ducks establish pair bonds only for the single breeding season. Pairs are formed during winter or spring migration and males follow the highly site-faithful females to their previous breeding sites (Mallory 2015). As expected, all three females returned to their breeding area in the centre of Kolguev, while at least two males followed their new mates to breeding sites far away from Kolguev. Since Kolguev forms a rather small part of the breeding range of the flyway population, there is a limited probability for a male to mate repeatedly with a female from Kolguev. The locations of the two males returning to Kolguev in 2020 did not indicate breeding activities. Probably these two males did not acquire a mate in 2020 and returned alone to their last year’s breeding area. Like in most ducks (Blums and Mednis 1996), the adult sex ratio of Long-tailed Ducks is highly skewed with twice as many adult males than females (Stempniewicz 1994, own observations), leaving many males unmated every summer. While males are not faithful to their breeding grounds, our observations of three males over two summer seasons suggest fidelity to their moulting sites.
Conclusions
The present data suggested that, in addition to breeding and wintering sites, coastal waters off the Yamal and Gydan peninsulas, as well as the White Sea, play a more important role in the annual cycle of Long-tailed Ducks breeding in the Barents Sea than was previously known. We observed strong sexual differences in the post-breeding movements. Our data further stress the importance of the southern part of the Baltic Sea as the main wintering site, such that declines in the wintering population in these waters cannot be explained by any changes in the choice of the wintering sites by Long-tailed ducks.
Data availability
The datasets supporting the conclusions of this article are deposited in the movebank.org repository under the study name “Long-tailed Ducks ARGOS 2019”.
References
Aarvak T, Øien IJ, Krasnov YV, Gavrilo MV, Shavykin AA (2013) The European wintering population of Steller’s eider Polysticta stelleri reassessed. Bird Conserv Int 23:337–343
Alerstam T, Hedenström A, Åkesson S (2003) Long-distance migration: evolution and determinants. Oikos 103:247–260
Austin J, Slattery S, Clarke RG (2014) Waterfowl populations of conservation concern: learning from diverse challenges, models and conservation strategies. Wildfowl Special Issue 4:470–497
Bellebaum J, Schirmeister B, Sonntag N, Garthe S (2013) Decreasing but still high: bycatch of seabirds in gillnet fisheries along the German Baltic coast. Aquatic Cons 23:219–221
Blums P, Mednis A (1996) Secondary Sex Ratio in Anatinae. Auk 113:505–511
Bowman TD, Silverman ED., Gilliland SG, Leirness JB (2015) Status and trends of North American sea ducks. Pp. 1–28 in Savard J-PL, Derksen DV, Esler D, Eadie JM (editors). Ecology and conservation of North American sea ducks. Studies in Avian Biology (no. 46), CRC Press, Boca Raton, FL.
Calenge C (2020) https://cran.r-project.org/web/packages/adehabitatHR/adehabitatHR.pdf. Accessed 18 Nov 2020
Dickson DL, Gilchrist HG (2002) Status of marine birds of the southeastern Beaufort Sea. Arctic 55(SUPPL. 1):46–58
Dingle H, Drake VA (2007) What is migration? Bioscience 57:113–121
Durinck J, Skov H, Andell P (1993) Seabird distribution and numbers in selected offshore parts of the Baltic Sea, winter 1992. Ornis Svecica 3:11–26
Esler D, Mulcahy DM, Jarvis RL (2000) Testing assumptions for unbiased estimation of survival of radiomarked harlequin ducks. J Wildlife Managem 64:591–598
ESRI - Environmental Systems Research Institute (2012). ArcGIS Release 10.1. Redlands, CA.
Flint PL, Reed JA, Lacroix DL, Lanctot RB (2016) Habitat use and foraging patterns of molting male long-tailed ducks in Lagoons of the Central Beaufort Sea, Alaska. Arctic 69:19–28
Fox AD, Jónsson JE, Aarvak T, Bregnballe T, Christensen TK, Clausen KK, Clausen P, Dalby L, Holm TE, Pavón-Jordan D, Laursen K, Lehikoinen A, Lorentsen S-A, Møller AP, Nordström M, Öst M, Söderquist P, Therkildsen OR (2015) Current and potential threats to Nordic duck populations — a horizon scanning exercise. Ann Zool Fennici 52:193–220
Fox AD, Nielsen RD, Petersen IK (2019) Climate-change not only threatens bird populations but also challenges our ability to monitor them. Ibis 161:467–474
Galkin SV, Vedenin AA (2015) Macrobenthos of Yenisei Bay and the adjacent Kara Sea shelf. Oceanology 55:606–613
Glazov PM, Loshchagina JA, Kondratyev AV, Zaynagutdinova EM, Kruckenberg H, Pokrovsky IG (2021) The long-term monitoring of bird populations on Kolguev Island in the Barents Sea. Arctic 74.
Glutz von Blotzheim UN, Bauer KM (1992) Handbuch der Vögel Mitteleuropas, Anseriformes (1.Teil), 2nd edn. AULA-Verlag, Wiesbaden
Hario M, Rintala J, Nordenswan G (2009) Dynamics of wintering Long-tailed Ducks in the Baltic Sea – the connection with lemming cycles, oil disasters, and hunting. Suom Riista 55:83–96
Hearn RD, Harrison AL, Cranswick PA (2015) International Single Species Action Plan for the Conservation of the Long-tailed Duck Clangula hyemalis, 2016–2025. AEWA Tech Ser 57.
Hede Jørgensen HB, Møller Hansen M, Loeschcke V (2005a) Spring-spawning herring (Clupea harengus L.) in the southwestern Baltic Sea: do they form genetically distinct spawning waves? ICES J Mar Sci 62:1065–1075
Hede Jørgensen HB, Hansen MM, Bekkevold D, Ruzzante DE, Loeschcke V (2005b) Marine landscapes and population genetic structure of herring (Clupea harengus L.) in the Baltic Sea. Mol Ecol 14:3219–3234
Heggøy O, Shimmings P (2016) Long-tailed Duck - common in the north, rare in south. Vår Fuglefauna 39:12–22 (In Norwegian)
Heldbjerg H, Peterse, IK, Ezhov AV (2020) Long-tailed Duck. p. 120 in: Keller V, Herrando S, Voříšek P et al. (eds) European Breeding Bird Atlas 2: Distribution, Abundance and Change. European Bird Census Council & Lynx Edicions, Barcelona.
del Hoyo J, Elliot A, Sargatal J 1992. Handbook of the Birds of the World, Vol. 1: Ostrich to Ducks. Lynx Edicions, Barcelona, Spain.
Karwinkel T, Pollet IL, Vardeh S, Kruckenberg H, Glazov P, Loshchagina J, Kondratyev A, Merkel B, Bellebaum J, Quillfeldt P (2020) Year-round spatiotemporal distribution pattern of a threatened sea duck species breeding on Kolguev Island, south-eastern Barents Sea. BMC Ecol 20:1–13
King JG (1973) A cosmopolitan duck moulting resort; Takslesluk Lake Alaska. Wildfowl 24:103–109
Koreisha MM (2000) Kolguev Island. J Geocryol 1:69–91 (in Russian)
Korschgen CE, Kenow KP, Gendron-Fitzpatrick A, Green WL, Dein FJ (1996) Implanting intra-abdominal radio transmitters with external whip antennas in ducks. J Wildlife Manag 60:132–137
Kozlovskiy VV, Chikina MV, Kucheruk NV, Basin AB (2011) Structure of the macrozoobenthic communities in the southwestern Kara Sea. Oceanology 51:1012–1020
Kruckenberg H, Kondratyev A, Zöckler C, Zaynagutinova E, Mooij JH (2012) Breeding waders on Kolguev Island, Barents Sea, N Russia, 2006–2008. Wader Study Group Bull 119:102–113
Lamb JS, Paton PW, Osenkowski JE, et al. (2020) Implanted satellite transmitters affect sea duck movement patterns at short and long timescales. Condor 122: duaa029.
Leafloor JO, Thompson JE, Ankney CD (1996) Body mass and carcass composition of fall migrant old squaws. Wilson Bull 108:567–572
Lehikoinen A, Jaatinen K, Vähätalo AV, Clausen P, Crowe O, Deceuninck B, Hearn R, Holt CA, Hornman M, Keller C, Nilsson L, Langendoen T, Tománková I, Wahl J, Fox AD (2013) Rapid climate driven shifts in wintering distributions of three common waterbird species. Global Change Biol 19:2071–2081
Lok EK, Esler D, Takekawa JY, De La Cruz SW, Boyd WS, Nysewander DR, Evenson JR, Ward DH (2012) Spatiotemporal associations between Pacific herring spawn and surf scoter spring migration: evaluating a ‘silver wave’ hypothesis. Mar Ecol Prog Ser 457:139–150
Loshchagina J, Vardeh S, Glazov P, Pollet IL, Quillfeldt P (2019) Long-tailed Duck (Clangula hyemalis) ecology: insights from the Russian literature. Part 2: European part of the Russian breeding range. Polar Biol 42:2277–2297
Mallory ML (2015) Site fidelity, breeding habitats, and the reproductive strategies of sea ducks. Pp. 337–364 in J.-P. L. Savard, D. V. Derksen, D. Esler, and J. M. Eadie (editors). Ecology and conservation of North American sea ducks. Studies in Avian Biology (no. 46), CRC Press, Boca Raton, FL.
Nilsson L (2016) Changes in numbers and distribution of wintering Long-tailed Ducks Clangula hyemalis in Swedish waters during the last fifty years. Ornis Svecica 26:162–176
Petersen MR, Savard J-PL (2015) Variation in migration strategies of North American sea ducks. Pp. 267–304 in Savard J-PL, Derksen DV, Esler D, Eadie JM (editors). Ecology and conservation of North American sea ducks. Studies in Avian Biology (no. 46), CRC Press, Boca Raton, FL.
Phillips RA, Xavier JC, Croxall JP (2003) Effects of satellite transmitters on albatrosses and petrels. Auk 120:1082–1090
Polte P, Gröhsler T, Kotterba P,von Nordheim L, Moll D, Santos J,Rodriguez-Tress P, Zablotski Y, Zimmermann C (2021) Reduced reproductive success of Western Baltic Herring (Clupea harengus) as a response to warming winters. Front. Mar. Sci. 8: 589242.
Rajasilta M, Eklund J, Hänninen J, Kurkilahti M, Kääriä J, Rannikko P, Soikkeli M (1993) Spawning of herring (Clupea harengus membras L.) in the Archipelago Sea. ICES J Mar Sci 50:233–246
Savard J-PL, Petersen MR (2015) Remigial molt of sea ducks. Pp. 305–335 in Savard J-PL, Derksen DV, Esler D, Eadie JM (editors). Ecology and conservation of North American sea ducks. Studies in Avian Biology (no. 46), CRC Press, Boca Raton, FL.
Sea Duck Joint Venture (2015) Atlantic and Great Lakes Sea Duck Migration Study: Progress Report June 2015. http//seaduckjv.org/science-resources/atlantic-and-great-lakes-sea-duck-migration-study/.
Skabeikis A, Morkūnė R, Bacevičius E, Lesutienė J, Morkūnas J, Poškienė A, Šiaulys A (2019) Effect of round goby (Neogobius melanostomus) invasion on blue mussel (Mytilus edulis trossulus) population and winter diet of the Long-tailed Duck (Clangula hyemalis). Biol Inv 21:911–923
Skov H, Heinänen S, Žydelis R, Bellebaum J, Bzoma S, Dagys M, Durinck J, Garthe S, Grishanov G, Hario M, Kieckbusch JJ, Kube J, Kuresoo A, Larsson K, Luigujoe L, Meissner W, Nehls HW, Nilsson L, Petersen IK, Roos MM, Pihl S, Sonntag N, Stock A, Stipniece A, Wahl J (2011) Waterbird populations and pressures in the Baltic Sea. Nordic Council of Ministers, Copenhagen
Smith LC, Sheng Y, MacDonald GM (2007) A first pan-Arctic assessment of the influence of glaciation, permafrost, topography and peatlands on northern hemisphere lake distribution. Permafrost Periglacial Proc 18:201–208
Sokolov V, Vardeh S, Quillfeldt P (2019) Long-tailed Duck (Clangula hyemalis) ecology: insights from the Russian literature. Part 1: Asian part of the Russian breeding range. Polar Biol 42:2259–2276
Spiegel CS, Berlin AM, Gilbert AT, Gray CO, Montevecchi WA, Stenhouse IJ, Ford SL, Olsen GH, Fiely JL, Savoy L, Goodale MW, Burke CM (2017) Determining Fine-Scale Use and Movement Patterns of Diving Bird Species in Federal Waters of the Mid-Atlantic United States Using Satellite Telemetry. U. S. Department of the Interior, Bureau of Ocean Energy Management, Office of Renewable Energy Programs, Sterling (USA).
Stempniewicz L (1994) Marine birds drowning in fishing nets in the Gulf of Gdańsk (southern Baltic): numbers, species composition, age and sex structure. Ornis Svecica 4:123–132
Vedenin AA, Galkin S, Kozlovskiy VV (2015) Macrobenthos of the Ob Bay and adjacent Kara Sea shelf. Polar Biol 38:829–844
Vickery JA, Ewing SR, Smith KW, Pain DJ, Bairlein F, Skorpilov J et al (2014) The decline of Afro-Palaearctic migrants and an assessment of potential causes. Ibis 156:1–22
Wetlands International (2021) Waterbird Population Estimates. Retrieved from wpe.wetlands.org on 23 Sept 2021.
Wilcove DS, Wikelski M (2008) Going, going, gone: is animal migration disappearing? PLoS Biol., 6, e188.
Wrona FJ, Prowse TD, Reist JD, Hobbie JE, Lévesque LM, Vincent WF (2006) Climate change effects on aquatic biota, ecosystem structure and function. Ambio 35:359–369
Žydelis R, Ruskyte DR (2005) Winter foraging of Long-tailed Ducks (Clangula hyemalis) exploiting different benthic communities in the Baltic Sea. Wilson J Orn 117:133–142
Žydelis R, Small C, French G (2013) The incidental catch of seabirds in gillnet fisheries: a global review. Biol Cons 162:76–88
Žydelis R, Richman SE (2015) Foraging behavior, ecology, and energetics of sea ducks. Pp. 241–265 in Savard J-PL, Derksen DV, Esler D, Eadie JM (editors). Ecology and conservation of North American sea ducks. Studies in Avian Biology (no. 46).
Acknowledgements
We want to thank all cooperation partners in the Long-tailed Duck project. We thank Sandra Vardeh for logistic support, and Daniela Tritscher, Eric Neuling and Irina Ilina for volunteering in the fieldwork in the Arctic tundra. We thank Rebecca McGuire, Peter Tickle and one anonymous reviewer for their constructive and helpful comments.
Funding
Open Access funding enabled and organized by Projekt DEAL. Funding for the collection and analysis of data was granted by the German Federal Agency for Nature Conservation (Bundesamt für Naturschutz, BfN), award MEERESENTEN (FKZ 3516821500), by the Vogelschutz-Komitee e.V. (VsK), Germany, by the Russian Ministry of Science and Higher Education (Projects No. AAAA-A19-119022190168-8, AAAA-A17-117122790002-8). Morten Ekker at the Norwegian Environmental Agency facilitated funding from the Cooperation on Biodiversity through the Joint Norwegian-Russian Agreement on Environmental Protection for the use of eight PTTs.
Author information
Authors and Affiliations
Contributions
PQ, HK, JB and PG conceived the study and PQ, HK, IJØ and JB obtained the funding. HK, PG, JL, JM and AK conducted the fieldwork on Kolguev Island. PQ analysed the data. PQ, JB and JM wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Ethical approval
All animal work was conducted according to relevant national and international guidelines and pursuant to applicable legislation of Russian Federation. Transmitter implantation was carried out under the permission of the Ethics Committee for Animal Research of St. Petersburg State University (SPbSU) No. 13-03-4 of 20 May 2019. Transmitters were only implanted in individuals determined to be in good health and for which the transmitter weight did not exceed 5.4% of body weight, as per accepted best practices. All implanted birds completely recovered after implantation and behaved normally. All stages of captures (capture, handling, surgery, recovery, and release) were supervised by a qualified veterinarian.
Consent for publication
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Quillfeldt, P., Morkūnas, J., Kruckenberg, H. et al. Year-round movements of Long-tailed Ducks Clangula hyemalis from Kolguev Island, Barents Sea. Polar Biol 45, 71–87 (2022). https://doi.org/10.1007/s00300-021-02973-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-021-02973-7