
Abstract
The winter/spring vertical distributions of polar cod, copepods, and ringed seal were monitored at a 230-m station in ice-covered Franklin Bay. In daytime, polar cod of all sizes (7–95 g) formed a dense aggregation in the deep inverse thermocline (160–230 m, −1.0 to 0°C). From December (polar night) to April (18-h daylight), small polar cod <25 g migrated into the isothermal cold intermediate layer (90–150 m, −1.4°C) at night to avoid visual predation by shallow-diving immature seals. By contrast, large polar cod (25–95 g), with large livers, remained below 180 m at all times, presumably to minimize predation by deep-diving mature seals. The diel vertical migration (DVM) of small polar cod was precisely synchronized with the light/dark cycle and its duration tracked the seasonal lengthening of the photoperiod. The DVM stopped in May coincident with the midnight sun and increased schooling and feeding. We propose that foraging interference and a limited prey supply in the deep aggregation drove the upward re-distribution of small polar cod at night. The bioluminescent copepod Metridia longa could have provided the light needed by polar cod to feed on copepods in the deep aphotic layers.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The diel vertical migrations (DVM) of marine zooplankton and fish likely involve the largest displacement of biomass on Earth (e.g., Hays 2003). DVM typically bring migrants from the bathypelagic or mesopelagic zones into the epipelagic zone at night to feed, and back at depth in daytime to avoid visual predators (e.g., Lampert 1989). The amount of carbon taken in the surface layer and carried to depth by these migrations (either as respired, excreted or egested carbon) represents on average from 4 to 34% and up to 70% of the particulate organic carbon flux (see Ducklow et al. 2001 for a review).
While the quest for food and the avoidance of visual predators are the ultimate factors driving the migrations, maximum relative change in light intensity (ΔI/I) at dusk and dawn is believed to be the zeitgeber triggering and synchronizing the DVM (e.g., Ringelberg 1995; Fortier et al. 2001; Pearre 2003). This raises the question of whether high-latitude zooplankton and nekton display DVM during the polar night and the midnight sun when ΔI/I is small (Fortier et al. 2001; Blachowiak-Samolyk et al. 2006; Berge et al. 2009). Polar cod (Boreogadus saida), by far the dominant fish on the extent Arctic shelves, is the central element of the relatively simple herbivory-based food web that dominates the pelagic ecosystem of the Arctic Ocean (e.g., Welch et al. 1992). In summer, the adults are dispersed in habitats ranging from coastal brackish waters to the demersal and pelagic zones of the shallow shelves, including the ice–water interface and the ice pack itself (e.g., Craig et al. 1982; Lønne and Gulliksen 1989; Jarvela and Thorsteinson 1999; Gradinger and Bluhm 2004). In autumn, large polar cod schools are observed in shallow (<10 m) waters (Welch et al. 1993; Hop et al. 1997). The species spawn under sea-ice in winter (Rass 1968; Craig et al. 1982) and spawning and post-spawning polar cod are known to congregate in regions deeper than 200 m (Ponomarenko 1968; Lear 1979; Benoit et al. 2008).
Using continuous multifrequency echosounding validated by trammel nets captures, Benoit et al. (2008) described the progressive build-up of a large, quasi-monospecific aggregation of polar cod during the winter/spring of 2003–2004 at depth range 140–230 m under the ice cover of central Franklin Bay (Canadian Beaufort Sea). In this paper, we document the diel vertical migrations of these polar cod in relation to photoperiod, the thermal structure of the water column, the vertical distribution of its main calanoid copepod prey, and the foraging activity of its predator the ringed seal (Phoca hispida). In particular, we examine how seasonal changes in photoperiod from the polar night in December to the midnight sun in May affect DVM in this key fish species.
Materials and methods
Study area
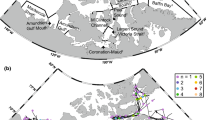
The V-shaped Franklin Bay in SE Beaufort Sea opens on the Amundsen Gulf that connects the Mackenzie Shelf and the Beaufort Sea to the channels of the Canadian Archipelago in the east (Fig. 1). The main water masses in the region comprise the Polar Mixed Layer (0–50 m), the Pacific Halocline (50–200 m), and the deep Atlantic Waters. Typically, landfast ice starts to form in October and covers the whole region by December. Ice reaches its maximum thickness of about 2 m in March. The ice break-up starts with the widening of a large polynya off Cape Bathurst in June, and Franklin Bay is typically ice-free by mid-July.

Bathymetric map of south-eastern Beaufort Sea showing the position of the overwintering station (star) in Franklin Bay
As part of the Canadian Arctic Shelf Exchange Study (CASES), the research icebreaker CCGS Amundsen overwintered at a 230-m-deep station (70.046°N, 126.302°W) in the stable landfast ice of Franklin Bay (Fig. 1). The vertical distribution and movements of fish (essentially polar cod, Benoit et al. 2008) and ringed seals were monitored acoustically from 22 December 2003 to 1 June 2004, while zooplankton and fish were sampled with nets at different times during this period.
Environmental variables
A CTD-rosette system (SBE-911 plus®, Seabird Electronics®) was deployed daily through the 2.4 × 2.3 m wide internal access well (moon pool) of the Amundsen to record temperature, salinity, and light from ~7 m (ship draft) to 225 m. On 13 February, hourly profiles were obtained from 0645 to 1930 hours. Modelled tidal elevation predictions at the overwintering station were calculated with the Webtide 0.65 software. Sunrise and sunset time at Cape Parry (Fig. 1) were obtained from the Weather Underground® meteorological archives (http://www.wunderground.com).
LI-COR LI-192® quantum sensors were used to measure incident photosynthetically available radiation (PAR) and light transmission through snow, ice, and water. PAR at the surface of the snow was measured continuously from 22 January 2004 to 25 May 2004 at a station 1.4 km away from the ship. On 18 April and 20 May, light transmission from the snow surface to the ice–water interface was measured at several locations and averaged over low and high snow covers, while under-ice PAR profiles in the first 30 m of the water column provided the light absorption coefficient of the water (K d). Incident PAR, per cent light transmission through the snow + ice cover, and K d were combined to estimate light intensity at depth for the periods 17–19 April and 19–21 May.
Hydroacoustic observations
Details of the acquisition and processing of the acoustic signal are given by Benoit et al. (2008). In summary, a SIMRAD EK60® three-frequency split-beam echosounder (38, 120, and 200 kHz) calibrated with the standard sphere method (Foote et al. 1987) was operated nearly continuously from 22 December to 29 May to record acoustic backscatter over the entire water column at the 230-m-deep overwintering station. For different technical and logistical reasons, gaps occurred in the record from 7 to 13 January, 21 January to 3 February, 26–27 February, 2–4 and 11–17 March. The unedited acoustic record at 38 kHz available from 22 December to 29 May along with recorded incident PAR at the snow surface are given in Online Resource 1. The 38-kHz signal was used to illustrate the vertical movement of the fish aggregation. For four representative 24-h periods in winter and spring (22 December, 13 February, 6 April, and 29 May), volume backscattering strength (S v in dB re 1 m−1) was calculated for the 38-kHz frequency following Benoit et al. (2008). S v was also calculated for 18–19 March to compare the diel vertical migrations of fish and zooplankton (see below). For each period, the S v hourly profile was computed with vertical echo-integration bins of 3 m, or for the depth interval corresponding to layers sampled with zooplankton nets.
The moon pool opens 3 m forward of the acoustic transducers mounted in the hull. Soon after the icing-in of the ship in Franklin Bay in December, ringed seals readily colonized the moon pool and frequently occupied it, with up to 9 animals in residence at times, until the ship left the area in early June. The length of seals occupying the moon pool was estimated from 24 photographs taken at 5 dates between 20 December and 18 March, using the known length of the moon pool wall as a reference. Following Smith (1987), seals <86 cm were classified as pre-weaning, 86–94 cm as immature, and ≥95 cm as mature. The acoustic tracks of diving seals in the echosounder record were easily distinguished from the much stronger acoustic signature of scientific instruments lowered through the moon pool. When entirely detected by the echosounder, the resulting V- or U-shaped trajectory provided a direct estimate of maximum dive depth. When the diving seal exited the emission cone of the acoustic transducer on the way down or entered the cone on the way up only, the resulting incomplete acoustic track provided no estimate of diving depth but was tallied as a dive.
Sampling and analysis of zooplankton and fish
At irregular intervals from early December to early June, a 0.5-m2 mouth opening multi-layer sampler (Hydrobios®) carrying 200-μm mesh nets was deployed 16 times through the moon pool to sample zooplankton in 4–6 discrete layers between 10 and 220 m. The sampler was lowered cod-end first to 220 m and retrieved at 0.5 m s−1 with nets opening and closing in succession at prescribed depths. Zooplankton samples were preserved in 4% buffered formaldehyde. In the laboratory, samples were sieved into >1 mm and <1 mm size fractions. Species and developmental stages were identified and enumerated under the microscope in a known aliquot of each fraction. Numbers were transformed into carbon biomass following the procedure described byForest et al. (2008). Starting at 0420 hours on 13 March, the multi-layer sampler was deployed 6 times at 4-h intervals, each time targeting the 220–201, 200–171, 170–141, 140–101, 100–61, and 60–10 m intervals. Unfortunately, the EK60 was temporarily out of function from 11 to 17 March, and the S v of the 38-kHz signal recorded on 18–19 March was used to compare the vertical distribution of fish to that of the zooplankton sampled over the daily cycle on 13–14 March.
Through an opening in the ice located 450 m from the ship, a 90-m-long trammel net (outside mesh 10.16 cm, inside mesh 2.54 cm) was deployed vertically over the 140 m to bottom depth interval for 12–24 h on 8 occasions in April and 3 occasions in May. Fresh total length was measured on the 576 polar cod collected, and catch depth (i.e., vertical position in the net) was recorded for the 453 fish that remained entangled in the net upon retrieval. In addition, 106 juvenile and adult fish were collected in the multi-layer Hydrobios zooplankton sampler from March to April and measured. Fish were frozen at −20°C or preserved in 95% ethanol. In the laboratory, 114 fish from the trammel net selected from each depth strata and 102 fish captured in the Hydrobios multi-layer sampler were thawed for stomach content analysis. Total length (cm) and wet weight (mg) were measured, and the stomach and liver were dissected and weighed. Stomachs were opened, and the overall content was weighed. Prey from the dissected stomachs was identified to the lowest taxonomic level possible and counted. A 12.6% carbon-to-wet weight ratio (Crabtree 1995) was used to estimate the carbon content of polar cod.
Results
Thermohaline structure of the water column
Time-depth sections of salinity and temperature illustrate the structure of the water column that prevailed from December to May under the ice of Franklin Bay (Fig. 2). In the 0 to ~40 m layer, the vertical stability provided by the surface halocline (30 to ~32 PSU) maintained an inverted gradient of near-freezing temperatures (−1.7 to −1.4°C). From ~40 to 140 m, an isothermal cold intermediate layer (−1.4°C) spanned the upper part of the Pacific Halocline (32–33 PSU). In the lower part of the Pacific Halocline (140 m to bottom), temperatures (−1.4 to 0.3°C) and salinities (33 to >34.5 PSU) increased with depth.

Time-depth sections of temperature (a) and salinity (b) at the overwintering station in Franklin Bay
Polar cod length, weight, liver weight, and gut content
Polar cod accounted for 97.8% of the 589 fish collected in the trammel net from 9 April to 21 May, and 100% of the fish collected in the multi-layer zooplankton net. Total length ranged from 11.6 to 25.9 cm (mean = 16.4 cm, standard deviation, SD = 2.2 cm) and weight from 4.5 to 95.4 g (mean = 17.8 g, SD = 11.4 g). The ratio of liver weight to overall weight increased with length, indicating that larger fish had disproportionately larger lipid reserves (Fig. 3a). The ratio of gut content weight to body weight changed in no consistent way with length: at any size, most polar cod had relatively low gut content, but a few fish presented much higher values (Fig. 3b). Seasonally, both liver weight and gut content weight standardized to body weight increased with time (regression analysis, P < 0.001) starting in May (Fig. 4).

Ratio of liver weight to fish weight (a) and gut content weight to fish weight (b) in relation to fish length for 264 polar cod captured under the sea-ice cover of Franklin Bay from March to May 2004. The regression in a is liver weight/total weight = 9.33 × 10−5 + 2.15 × 10−3 length (r 2 = 0.15, P < 0.01)

Ratio of liver weight to fish weight (a) and gut content weight to fish weight (b) by sampling date for polar cod captured from March to May 2004 under the sea-ice cover of Franklin Bay
Polar cod preyed primarily on copepods (Table 1). Among the copepods identified to species, the late stages of the calanoids Metridia longa (CVI), Calanus hyperboreus (CIV-CVI), and C. glacialis (CIII-CVI) occurred most frequently. Despite their relatively small size, copepods contributed the bulk of the carbon intake of polar cod because of their high frequency as prey. When preyed upon, the occasional hyperiid amphipod Themisto libellula and the rare fish prey (essentially polar cod) contributed substantially to individual gut content. However, because of low occurrence, their overall importance was negligible. Unidentifiable remains and digested material accounted for a large part of the gut content. Trematode parasites were frequent and nematodes were recorded occasionally (Table 1).
Photoperiod and seasonal changes in the DVM pattern of polar cod
From December to May, the density of the deep scattering layer (SL) increased progressively in the lower part of the Pacific Halocline (Fig. 5), corresponding to the build-up of the large aggregation of polar cod observed and validated by Benoit et al. (2008). During the darkest period of the polar night in late December, the relatively low biomass of scatterers was spread over the entire water column with denser concentrations below 100 m (Fig. 5a). The SL exhibited some faint DVM with migrants moving deeper during a brief period of about 4 h centred around noon. By early January, the biomass was concentrated in the deep half of the water column, and the DVM pattern was sharper (Fig. 5b). The downward and upward migrations corresponded, respectively, with the dawn and sunset civil twilights at Cape Parry.

38-kHz echograms for selected 5-d periods at the overwintering station in Franklin Bay (December 2003 to May 2004). Vertical arrows on top of (a) and (b) indicate time of morning (upward arrow) and evening (downward arrow) theoretical civil twilight at 70°N in December and January. Starting in February, the photosynthetically available radiation (PAR) at the surface of the snow cover is plotted on the echogram (dotted black line). Vertical lines on the echograms are the track of scientific instruments deployed through the moon pool of the ship. Vertical white areas extending from surface to bottom in (a) and (c) correspond to missing record. Grey areas correspond to missing data. S v values below the threshold are in white. Lower threshold for colour scale is −90 dB for 22–26 December (a)
By mid-February and the rising of the sun over the horizon, the biomass of polar cod and the vertical amplitude of the DVM increased (Fig. 5c). By day and night, biomass remained highest in the relatively warm deep layer (−0.4 to 0°C, 190–236 m). Some fish moved into the cold (−1.4°C) intermediate layer at night reaching depths as shallow as 80 m. All fish moved down below 140–160 m in daytime. The rapid upward and downward migrations coincided with sunset and sunrise, respectively (Fig. 5c). By March, the DVM pattern was particularly well defined (Fig. 5d). At night, fish migrating into the cold intermediate layer (70–140 m) and fish remaining in the deep layer (160-bottom) formed two distinct aggregations linked by a vertical vein of higher scattering density at the time of the upward and downward migrations. As nights shortened, the night-time incursions of polar cod into the cold intermediate layer also shortened. By mid-April, the fish occupied the intermediate layer for ca. 7 h only (Fig. 5e), compared to 15 h in mid-March and 17 h in mid-February.
The classical pattern of DVM (shallow at night, deep in daytime) that developed in late December persisted until 10 May. From 11 to 13 May, the DVM blurred, fish tending to remain below 140 m by day and by night (Online Resource 1). By 14 May, the texture of the echogram became highly striated vertically, suggesting unsynchronized migrations of individual fish or fish schools (Fig. 5f). From 14 to 18 May, polar cod were found in the cold intermediate layer (80–140 m) at anytime during the day, with a tendency to return below 140 m around midnight. This behaviour resulted in an inversion of the migration (shallow in daytime, deep at night), which lasted for less than a week before the DVMs ceased altogether.
Based on the fish collected in the trammel net and the stratified zooplankton net, the high-density SL in the 180–225 m depth interval comprised both small and large fish (7–95 g), whereas the polar cod migrating above the 180 m depth and into the cold intermediate layer consisted essentially of small fish <25 g (Fig. 6). The contribution of the liver to overall body weight increased with depth, large fish with large livers being distributed below 180 m (Fig. 7a). The ratio of gut content weight to fish weight varied widely with depth, with a tendency to be high in polar cod collected immediately below the cold layer (140–160 m) and in the deep layer below 180 m (Fig. 7b).

Weight frequency distribution of net-captured polar cod by depth intervals

Vertical distribution of the ratio of liver weight to fish weight (a) and stomach content weight to fish weight (b) for polar cod collected from March to May 2004 at the overwintering station in Franklin Bay. The full circle corresponds to the mean value and the horizontal line to ±1SD
The vertical co-distribution of polar cod and its zooplankton prey
As the fish started to aggregate in Franklin Bay in December, the biomass of polar cod and that of its copepod prey were commensurate (Table 2). By February, polar cod biomass exceeded by over two orders of magnitude than that of copepod prey. The difference widened to 3 and then 4 orders of magnitude in March and April, respectively, as the fish aggregation developed and copepod biomass remained stable or decreased slightly.
In mid-March, the main copepod prey of polar cod was distributed primarily below 100 m (Fig. 8). The three species presented some limited DVM, with an increasing fraction of the biomass moving into the 0–100 m layer during sunset to dawn (2030–0830 hours) and then vacating this layer to concentrate in the 140–200 m layer in the afternoon. With the caveat that the vertical distributions of copepod prey and polar cod were assessed on different dates in mid-March, few polar cod followed their copepod prey in the 0–100 m layer at night (Fig. 8). Below 100 m during twilight and darkness (2030–0830 hours), the vertical distribution of polar cod roughly matched that of its copepod prey, for example at 0030 and 0830 hours. In the afternoon (1230 and 1630 hours), the fish concentrated in the 140–170 m layer, slightly above the depth of maximum concentration of their prey.

Vertical distribution of the biomass of polar cod (g C m−3) and its main copepod prey (mg C m−3) at 4-h intervals under the ice of Franklin Bay in March. Note that the polar cod distribution was measured on 18 March and that of copepods on 13–14 March. The photosynthetically available radiation (PAR in μmole photon m−2 s−1) at the snow surface at the time of sampling is indicated in parentheses. The horizontal stippled reference line at 100 m is provided to facilitate comparisons among panels
Seal size and diving depth
Ringed seals frequenting the moon pool included pre-weaning animals <86 cm long (46%), immatures 86–94 cm long (32%), and mature seals >94–122 cm long (21%). Ringed seals dove from the moon pool to a given depth, returning to the surface after a short period (10–120 s) at that depth. The average descending and ascending rates were equal at 1.17 m s−1 (SD = 0.27). In 517 (14.8%) of the 3,486 dives analysed for the periods 22–24 December, 12–14 February, 5–7 April, and 28–30 May, both the descending and ascending limbs of the trajectory were recorded entirely or partly, and maximum dive depth could be estimated directly or by extrapolating the intersection depth of the two limbs. When pooled over the four selected periods, the frequency distribution of dive depth presented two modes at 110 m and around 190 m, respectively (Fig. 9). Assuming that the shallow and deep modes corresponded to the diving of immature and mature seals, respectively, dives ≤130 and >130 m deep were analysed separately.

Depth frequency distribution of fully detected ringed seal dives (n = 517) at the overwintering station of Franklin Bay. The data for 22–24 December, 12–14 February, 5–7 April and, 28–30 May were pooled
At night, the average shallow dive reached to the −74 dB S v isopleth used here to define the top of the fish aggregation (Fig. 10a–c). Shallow dives were less frequent in daytime than at night and remained short of the deeper daytime aggregation of fish. With the midnight sun in late May, shallow dives no more reached into the fish aggregation (Fig. 10d). The deep dives reached into the core of the fish aggregation (Fig. 10). The average deep dive depth tended to track changes in the depth of maximum S v, with seals diving deeper in daytime when the fish were distributed deeper.

Hourly mean depth (±standard deviation) of fully detected ringed seal dives plotted over the time-depth section of volume backscattering strength (S v in dB re 1 m−1) for four representative periods during the overwintering in Franklin Bay. Full circles represent the mean depth (±SD) of shallow dives (<130 m, presumably by immature seals) and open circles that of deep dives (>130 m, presumably by mature seals). To increase sample size, dive depths for a given hour were averaged over 3 days. The time-depth section of S v is for the central day of the 3-d period
Seal diving frequency varied considerably among the dates selected, and no consistent diel pattern was observed (Fig. 11). In late December, dive frequency remained roughly constant at ca. 10 h−1 over the daily cycle (Fig. 11a). Dive frequency decreased from early morning to the afternoon in mid-February, but increased over the same period in early April (Fig. 11b, c). In late May, dive frequency was maximum at midnight, reached a minimum around noon and increased again in the afternoon (Fig. 11d).

Hourly mean number of ringed seal dives (fully and partially detected) for four representative periods during the overwintering in Franklin Bay. To increase sample size, dive depths for a given hour were averaged over 3 days
Details of polar cod DVM in relation to light, temperature and seal activity
A selected 48-h echogram (38 kHz frequency) illustrates the details of the DVM of polar cod at the time when the fish aggregation reached its maximum density in mid-April (Fig. 12a). Starting at noon on 17 April, all fish were distributed from 140 to 225 m, below the 10−6 μmole photon m−2 s−1 isolume. Highest densities (−40 dB) occurred in a narrow scattering band at 190–200 m. Throughout the 48-h record, this high-density layer tended to spread and re-aggregate within the 180 m to bottom interval (Fig. 12a). Around 2000 hours on 17 April, more polar cod left the high-density layer to enter the 180–140 m layer, coincident with the shallowing of the 10−6 μmole photon m−2 s−1 isolume, but no fish invaded yet the cold layer (−1.4°C) above 140 m. When the rate of decline in incident light intensity (∆I/I) reached its maximum just before midnight, some fish started to move into the intermediate cold layer above 140 m. At 0600 hours, the maximum rate of increase in light intensity triggered the evacuation of the cold layer and the rebuilding of the dense scattering layer in the 180–225 m depth interval. The DVM pattern was similar on the second circadian cycle (18–19 April) except that (1) the pre-migration above 180 m started earlier (1200 hours) than on the previous day (2000 hours); (2) apparently more fish moved into the cold layer during the night; and (3) the morning downward migration was less pronounced (Fig. 12a).

Forty-eight hour echograms centred on 18 April and 20 May (38 kHz). The thick line indicates the calculated depth of the 10−6 μmole photon m−2 s−1 isolume. The dotted line is ∆I/I, the rate of change in light intensity. White circles indicate the depth of ringed seal individual dives. Insert in b is an echogram (from surface to bottom) exemplifying the migrating schools
At the time resolution used in Fig. 12, the trajectory of diving seals are blurred while that of scientific probes remain clear (e.g., the regular CTD casts at 0800 and 2000 hours each day). The fish displayed some avoidance reaction to the instruments, the density of the SL decreasing during instrument profiles (Fig. 12a). The turning depth of shallow (<130 m) seal dives seldom reached the fish aggregation except at night when polar cod migrated upward above 140 m. More often than otherwise, the turning depth of the deep dives coincided with the depth of the highest densities of fish. Deep dives were particularly frequent in the 2 h following a CTD cast, an indication that the diving behaviour of seals was affected by the deployment of instruments through the moon pool they occupied (Fig. 12a).
Under the midnight sun in late May, polar cod showed little sign of synchronized DVM (Fig. 12b). Rather, the vertically striated texture of the echogram (insert on Fig. 12b) suggested intense unsynchronized vertical migrations of schools and/or individual fish between the cold intermediate layer (<140 m) and the deep layer. Within the 60–225 m interval, the vertical distribution of polar cod was affected by seals, the fish tending to avoid the depth interval where seal dives reached (Fig. 12b).
Discussion
From the polar night to the midnight sun: light and polar cod DVM in ice-covered Franklin Bay
In ice-free waters, the DVM of Arctic zooplankton are strong during the succession of day and night in spring and fall, and weak or absent during the polar night and the midnight sun (Buchanan and Haney 1980; Fischer and Visbeck 1993; Falkenhaugh et al. 1997; Blachowiak-Samolyk et al. 2006; Berge et al. 2009). The weakening of ∆I/I, the zeitgeber that synchronizes the migration, explains the weakening of the DVM during the polar night (e.g., Berge et al. 2009). Continuous irradiance and the resulting lack of night-time refuge against visual predators likewise explain the cessation of DVM in ice-free waters during the midnight sun (Hays 1995; Fortier et al. 2001). In ice-covered waters, the snow and sea-ice cover can modify the seasonal pattern in DVM by attenuating underwater light and modifying the intensity of ∆I/I. For example, in Svalbard during the polar night, the dimming of light to almost imperceptible levels stopped the DVM of zooplankton in ice-covered Rijpfjorden, while the migrations continued in ice-free Kongsfjorden (Berge et al. 2009). As well, in Barrow Strait (74.5°N) in summer, sea-ice attenuated irradiance at night below the reported threshold of visual perception of fish predators, and copepods maintained their DVM under the midnight sun (Fortier et al. 2001).
In the present study, seasonal changes in the duration of polar cod DVM closely tracked the lengthening of the photoperiod from the continuous polar night in December until the midnight sun in May. Even at the pinnacle of the polar night on 22 December, the weak upward and downward movements of the scattering layer in the 0–100 m layer were precisely synchronized with the theoretical maximum rate of change in twilight intensity (∆I/I) at 70°N. A first interpretation of this synchrony is that polar cod perceived variations in whatever little downwelling light penetrated the snow and ice cover and reached to 100 m depth. At 70°N in late December, civil twilight at the surface (during which objects can be distinguished by the human eye) prevails for a period of 4 h centred on local noon. In Amundsen Gulf (70°N to 72°N) in 2007–2008, maximum incident PAR during civil twilight raised slowly from 0.5 μmole photon m−2 s−1 on 26 December to 1 μmole photon m−2 s−1 on 5 January and increased exponentially afterwards to reach 7 μmole photon m−2 s−1 on 21 January (T. Papakyriakou, University of Manitoba, unpublished data). The minimum thickness of the snow and ice covers were 0.07 and 0.8 m, respectively, around the overwintering station in December 2003 (Langlois et al. 2006). Using the minimum light attenuation coefficients of snow and sea-ice (4.3 and 1.1 m−1, respectively, e.g., Perovich 1996) and the K d for water measured in April (0.11 m−1), an incident PAR of 0.5 μmole photon m−2 s−1 in late December would have been attenuated to 2.3 × 10−6 μmole photon m−2 s−1 at 100 m depth. The same calculation with the maximum coefficient of attenuation of snow (40 m−1) and ice (1.5 m−1) yields a value of 1.4 × 10−7 μmole photon m−2 s−1 at 100 m depth. These rough estimates of the maximum and minimum light reaching 100 m around noon in late December bracket the 1.9 × 10−7 μmole photon m−2 s−1 threshold to which juvenile walleye pollock (Theragra chalcogramma) respond in the laboratory (Ryer and Olla 1998). Hence, it is conceivable that dark-adapted polar cod possessed the visual acuity to detect the weak solar signal that reached at 100 m in December in Franklin Bay. An alternative explanation is that the precise timing of the DVM of polar cod on maximum ∆I/I in late December was a remanence of an earlier synchronization to the stronger light signal with similar period that prevailed before the setting of the polar night in December.
Based on measured light penetration in April and May, migrating polar cod avoided moving above the 10−6 μmole photon m−2 s−1 isolume (Fig. 12). This threshold is two orders of magnitude lower than the ~1.89 × 10−4 μmole photon m−2 s−1 level at which the feeding of polar cod became significantly impaired in the laboratory (Girsa 1961 cited in Blaxter 1970), which suggests that the feeding of polar cod moving into the cold intermediate layer at night may have been limited by light. However, after several months of adaptation to nearly absolute darkness at depth under the snow and ice cover, polar cod may achieve higher visual acuity than measured under laboratory conditions. Alternatively, when stimulated mechanically or by turbulence, bioluminescent organisms such as Metridia longa (e.g., Lapota et al. 1989) can provide substantial background illumination that can be used by predators (Widder et al. 1992). Within the extremely dense aggregation at depth in Franklin Bay (Benoit et al. 2008), turbulence from swimming fish and/or direct contact between fish and M. longa may have triggered the emission of the light needed by polar cod to detect its prey under the 10−6 μmole photon m−2 s−1 solar isolume. The light emitted by M. longa would make it particularly vulnerable to detection and capture, explaining the relatively high frequency of this copepod in the gut content of polar cod despite its small weight and lipid content and, hence, energy value. Most importantly, bioluminescence by M. longa may also have provided polar cod with the background illumination to detect the large and energy-rich but non-bioluminescent Calanus hyperboreus and C. glacialis.
While the synchronized migration of the scattering layer often stops during the midnight sun, individual migrants may nevertheless perform unsynchronized migrations. For example, in a Svalbard fjord (79°N) in June, the vertical velocity of individual zooplankton (most likely Calanus finmarchicus and C. glacialis) indicated a continuous net downward movement in the surface layer and an upward movement in the deep layer, while the overall scattering layer exhibited no net movement (Cottier et al. 2006). In the present study, the vertical striation of the echogram under the midnight sun in late May suggests as well the rapid vertical migration of individuals or schools with no net synchronized movement of the main aggregation except for the avoidance of seals (Fig. 12b).
Prey, predators and the causes of polar cod DVM under the ice
Among various theories on the causes and trophic advantages of diel vertical migrations in the ocean, the complementary predator-evasion and hunger/satiation hypotheses are most popular (Lampert 1989; Hays 2003; Pearre 2003). Both hypotheses assume that (1) food is limited at depth and available in some shallower layer; (2) in daytime, vulnerability to visual predators is higher in the shallow layer than at depth; and (3) the energy gain of feeding in the shallow layer exceeds the energy expenditure of the migration (Gliwicz and Pijanowska 1988; Lampert 1989). Under these conditions, the maximum in ΔI/I at dusk prompts an upward migration, while the downward migration is triggered by satiation at anytime during the night or by maximum ΔI/I at dawn (Ringelberg 1995; Fortier et al. 2001; Sourisseau et al. 2008). According to the predator-evasion hypothesis, the need to avoid visual predators dictates the downward migration in the morning and the occupation of the deep layer during daytime. Increased risk of predation due to a large size or high visibility limits the amplitude and duration of incursions in the shallow layer at night and increases the depth of daytime residence (Giske et al. 1990; Fortier et al. 2001). According to the hunger/satiation hypothesis, the drive for the night-time upward migration should increase with hunger and only those individuals sufficiently hungry will migrate upward at night (see Pearre 2003 for a review).
Under the ice of Franklin Bay in winter and spring, the greater density of copepod prey in the deep layer (140–225 m) than in the lower part of the cold intermediate layer into which some polar cod migrated at night (100–140 m) seemed to contradict the assumption of food limitation at depth and food availability above. However, the precise assumption is that food limitation prevails at depth for those fish that migrate, not necessarily for all fish. With their heavy livers and heavy gut content relative to body weight, polar cod >25 g showed no sign of food limitation and did not migrate into the intermediate layer (Fig. 6). By contrast, the polar cod that migrated into the intermediate layer were small fish (<25 g) with small livers and gut contents and, perhaps, lesser visual acuity than their larger congeners (Fernald 1990). We suspect that a limitation of food intake due to foraging interference with large polar cod in the extremely dense aggregation at depth drove the upward migration of small polar cod into the cold intermediate layer where the ratio of copepod prey to fish was more favourable. That polar cod feed more successfully outside than inside schools (Hop et al. 1997) supports this interpretation.
Consistent with the hunger/satiation hypothesis, only a small fraction of the polar cod population migrated upward on a given day. Digestion in polar cod is slow, and a total evacuation time of about 400 h (ca. 17 days) has been reported (Hop and Tonn 1998; Saether et al. 1999). Hop and Graham (1995) suggested that polar cod feed infrequently in winter, just enough to minimize weight losses. Hence, small polar cod could have remained at depth for several days between feeding forays into the intermediate cold layer, which would explain the overall low fraction of the population participating in the migration on any given night. If so, the observed migration behaviour of polar cod would represent an extreme case of DVM in which the vertical interchange of individuals actually occurs on a time scale of several days. Under similar conditions of sub-zero temperature and low light availability in the Southern Ocean in winter, Antarctic cod (Notothenia coriiceps) enters a state of dormancy interrupted by awakenings of a few hours every 4–12 days, during which metabolic activity builds back to summer levels (Campbell et al. 2008). The intriguing possibility that small polar cod adopt a similar hibernating strategy, remaining in a state of torpor interrupted by fortnightly feeding incursions into the intermediate layer, could be tested using acoustic tags as in Campbell et al. (2008).
Studies of DVM in the ocean generally assume that the deeper distribution of migrants in daytime is an escape response to visual predators prowling the photic layer. Yet, these visual predators and their predatory impact are seldom identified and quantified. The one and apparently unique predator of polar cod under the ice of Franklin Bay was the ringed seal. Under coastal landfast ice in winter and spring, ringed seals dive beyond 200 m and maximum diving depth often corresponds to the bottom (Lydersen and Hammill 1993; Kelly and Wartzok 1996). In the nearly absolute darkness prevailing at depth under the ice in winter, ringed seals use their highly sensitive vibrissae to detect and capture their prey (Hyvärinen 1989). Dive duration and depth are strongly correlated to body mass (Kelly and Wartzok 1996; Teilmann et al. 1999; Kunnasranta et al. 2002). In the North Water (northern Baffin Bay) in winter and spring, sub-adults equipped with acoustic tags dove only to shallow depth, while an adult performed deep dives under the ice (Born et al. 2004). Born et al. (2004) surmised that immature seals preyed upon small polar cod and the hyperiid amphipod Themisto libellula in the surface layer, whereas adult seals preyed on large polar cod at depth.
Both large mature and small immature ringed seals occupied the moon pool of the ship and, consistent with the scenario of Born et al. (2004), two modes existed in the distribution of dive depths in April: relatively shallow (20–130 m) dives presumably by small seals and deeper dives (130–230 m) by large seals (Fig. 9). In agreement with the observations of Kelly and Wartzok (1996), ringed seals dove by night and by day, and no clear relationship between dive frequency and light was observed. However, the average depth of the deep dives varied with the depth of the fish aggregation (Fig. 10), implying that the energy expended by mature seals to feed on large polar cod varied with the depth of their prey. Hence, on average, residing at depth must have provided polar cod with some protection against predation by ringed seals.
Differences in diving depth and in the prey targeted, respectively, by small and large ringed seals could explain the size-related differences in the migration behaviour of polar cod observed in the present study. In agreement with the prediction of reduced migrations and a deeper distribution of large conspicuous individuals (Giske et al. 1990; Fortier et al. 2001), large polar cod with disproportionately large livers remained at depth, avoiding moving above 180 m (Fig. 6). In the North Atlantic, grey seals (Halichoerus grypus) have been observed to bite off the belly of Atlantic cod (Gadus morhua) including the lipid-rich liver (Fu et al. 2001; Chouinard et al. 2005). In the present study, the livers of large polar cod brought back to the moon pool by large seals were often extruding from the torn belly of the fish (L. Fortier, personnel observation), suggesting that ringed seals similarly sought the lipid-rich liver. For large polar cod targeted by large deep-diving seals and detected by vibrissae rather than visually, moving above 180 m at night afforded no particular foraging advantage or refuge against predation. Accordingly, for those large fish, the best strategy would be to remain as deep as possible at all times so as to increase foraging costs to their predators.
By contrast, the fish migrating at night into the intermediate layer were small polar cod with relatively small livers and, presumably, reduced energy value for large competent seals that can reach the larger fish with larger livers at depth. These small polar cod performed DVM that were precisely synchronized with maxima in ΔI/I at dawn and dusk. Assuming that immature seals are less competent at detecting and capturing their prey in the dark than mature seals, the combination of improved feeding conditions due to reduced interference with large congeners and reduced risk of visual detection by small immature seals at night would drive the DVM of these small polar cod into the intermediate cold layer.
Reverse DVM (shallow in daytime, deep at night) has been documented in zooplankton (see Cohen and Forward 2009 for a review), but the causes and adaptive value of this behaviour are poorly understood (Tester et al. 2004). In the week before the final transition from the day/night cycle to the midnight sun in mid-May, polar cod exhibited a reverse migration (Fig. 5f). During this period of nearly continuous daylight and reduced snow cover, some light may have reached over most of the water column at all times except at depth during the night. Hence, a possible interpretation of the reverse DVM is that, with the disappearance of their refuge from visual predators at depth in daytime, polar cod moved upward to feed in the well-illuminated intermediate cold layer. During the nocturnal twilight, most fish returned below 120 m where dimmer light conditions still afforded some protection against predation by ring seals.
As expected, synchronized DVM stopped under the midnight sun in late May. The end of synchronized DVM coincided with a change in the structure of the fish aggregation, which gradually developed into a dense shoal of schools and ribbons. Along with DVM, schooling is a widespread anti-predation behaviour in fish (e.g., Pitcher et al. 1988; Pitcher and Parrish 1993). After 20 May, the lack of day/night difference in refuge from visual predators apparently triggered schooling as an alternative behaviour to reduce predation (Fig. 12b). Starting in mid-May, the shift from normal DVM to reverse DVM, and then to schooling in the cold intermediate layer at all time of day coincided with the resumption of feeding and the replenishment of liver reserves. Feeding efficiency is another advantage attributed to schooling, which increases detection volume compared to solitary foraging when searching for food concentrations, especially in a zooplankton-poor environment such as often encountered in the Arctic.
Finally, under the midnight sun in May, the vertical distribution and movement of polar cod clearly responded to seal forays in the deep layer (Fig. 12b). In summary, under the ice of Franklin Bay, the lack of DVM by large polar cod and the DVM of small polar cod in winter and spring, as well as reverse DVM and the schooling of polar cod under the midnight sun in May, were all consistent with a need to evade the different predation pressures by mature and immature ringed seals. Our observations suggest that, as in most trophic interactions in Arctic waters (e.g., Falk-Petersen et al. 2009), the quest for lipids dictates the foraging behaviour of ringed seals and, consequently, the observed size-related differences in the DVM of polar cod under the ice in winter and spring.
References
Benoit D, Simard Y, Fortier L (2008) Hydro-acoustic detection of large winter aggregations of Arctic cod (Boreogadus saida) at depth in ice-covered Franklin Bay (Beaufort Sea). J Geophys Res Ocean 113:C06S90. doi:10.1029/2007JC004276
Berge J, Cottier F, Last KS, Varpe Ø, Leu E, Søreide J, Eiane K, Falk-Petersen S, Willis K, Nygård H, Vogedes D, Griffiths C, Johnsen G, Lorentzen D, Brierley AS (2009) Diel vertical migration of Arctic zooplankton during the polar night. Biol Lett 5:69–72. doi:10.1098/rsbl.2008.0484
Blachowiak-Samolyk K, Kwasniewski S, Richardson K, Dmoch K, Hansen E, Hop H, Falk-Petersen S, Mouritsen LT (2006) Arctic zooplankton do not perform diel vertical migration (DVM) during periods of midnight sun. Mar Ecol Prog Ser 308:101–116. doi:10.3354/meps308101
Blaxter JHS (1970) Light; animals; fishes. In: Kinne O (ed) Marine ecology. Vol 1, part 1. Wiley, New York, pp 213–320
Born EW, Teilmann J, Acquarone M, Riget FF (2004) Habitat use of ringed seals (Phoca hispida) in the North Water area (north Baffin Bay). Arctic 57:129–142
Buchanan C, Haney JF (1980) Vertical migrations of zooplankton in the Arctic: a test of the environmental controls. In: Kerfoot WC (ed) Evolution and ecology of zooplankton communities. University Press, Hanover, pp 69–79
Campbell HA, Fraser KPP, Bishop CM, Peck LS, Egginton S (2008) Hibernation in an Antarctic fish: on ice for winter. PLoS ONE 3:e1743. doi:10.1371/journal.pone.0001743
Chouinard GA, Swain DP, Hammill MO, Poirier GA (2005) Covariation between grey seal (Halichoerus grypus) abundance and natural mortality of cod (Gadus morhua) in the southern Gulf of St. Lawrence. Can J Fish Aquat Sci 62:1991–2000. doi:10.1139/F05-107
Cohen JH, Forward RB (2009) Zooplankton diel vertical migration—a review of proximate control. In: Gibson RN, Atkinson RJA, Gordon JDM (eds) Oceanography and marine biology: an annual review, vol 47. CRC press, Boca Raton, pp 77–109
Cottier FR, Tarling GA, Wold A, Falk-Petersen S (2006) Unsynchronised and synchronised vertical migration of zooplankton in a high Arctic fjord. Limnol Oceanogr 51:2586–2599
Crabtree RE (1995) Chemical-composition and energy content of deep-sea demersal fishes from tropical and temperate regions of the western north-Atlantic. Bull Mar Sci 56:434–449
Craig PC, Griffiths WB, Haldorson L, McElderry H (1982) Ecological studies of Arctic cod (Boreogadus saida) in Beaufort Sea coastal waters, Alaska. Can J Fish Aquat Sci 39:395–406
Ducklow HW, Deborah K, Steinberg DK, Buesseler KO (2001) Upper ocean carbon export and the biological pump. Oceanography 14:50–58
Falkenhaugh T, Tande KS, Semenova T (1997) Diel, seasonal and ontogenetic variations in the vertical distributions of four marine copepods. Mar Ecol Prog Ser 149:105–119. doi:10.3354/meps149105
Falk-Petersen S, Mayzaud P, Kattner G, Sargent J (2009) Lipids and life strategy of Arctic Calanus. Mar Biol Res 5:18–39. doi:10.1080/17451000802512267
Fernald RD (1990) Teleost vision—seeing while growing. J Exp Zool 256:167–180. doi:10.1002/jez.1402560521
Fischer J, Visbeck M (1993) Seasonal variation of the daily zooplankton migration in the Greenland Sea. Deep Sea Res Part I 40:1547–1557. doi:10.1016/0967-0637(93)90015-U
Foote KG, Knudsen HP, Vestnes G, MacLennan DN, Simmonds EJ (1987) Calibration of acoustic instruments for fish density estimation: a practical guide. Tech Rep 144, Int Counc Explor Sea, Copenhagen
Forest A, Sampei M, Makabe R, Sasaki H, Barber D, Barber DG, Gratton Y, Wassmann P, Fortier L (2008) The annual cycle of particulate organic carbon export in Franklin Bay (Canadian Arctic): Environmental control and food web implications. J Geophys Res-Oceans 113:C03S05. doi:10.1029/2007JC004262
Fortier M, Fortier L, Hattori H, Saito H, Legendre L (2001) Visual predators and the diel vertical migration of copepods under Arctic sea ice during the midnight sun. J Plankton Res 23:1263–1278
Fu C, Mohn R, Fanning LP (2001) Why the Atlantic cod (Gadus morhua) stock off eastern Nova Scotia has not recovered. Can J Fish Aquat Sci 58:1613–1623
Girsa II (1961) Availability of food animals to some fishes at different conditions of illumination (in Russian). Trudy Soveshch Ikhtiol Kom 13:355–359
Giske J, Aksnes DL, Baliño BM, Kaartvedt S, Lie U, Nordeide JT, Salvanes AGV, Wakili SM, Aadnesen A (1990) Vertical-distribution and trophic interactions of zooplankton and fish in Masfjorden, Norway. Sarsia 75:65–81
Gliwicz MZ, Pijanowska J (1988) Effect of predation and resource depth distribution on vertical migration of zooplankton. Bull Mar Sci 43:695–709
Gradinger RR, Bluhm BA (2004) In situ observations on the distribution and behavior of amphipods and Arctic cod (Boreogadus saida) under the sea ice of the High Arctic Canada Basin. Polar Biol 27:595–603. doi:10.1007/s00300-004-0630-4
Hays GC (1995) Diel vertical migration behaviour of Calanus hyperboreus at temperate latitudes. Mar Ecol Prog Ser 127:301–304
Hays GC (2003) A review of the adaptive significance and ecosystem consequences of zooplankton diel vertical migrations. Hydrobiologia 503:163–170. doi:10.1023/B:HYDR.0000008476.23617.b0
Hop H, Graham M (1995) Respiration of juvenile Arctic cod (Boreogadus saida): effects of acclimation, temperature, and food intake. Pol Biol 15:359–367. doi:10.1007/BF00238487
Hop H, Tonn WM (1998) Gastric evacuation rates and daily rations of Arctic cod (Boreogadus saida) at low temperatures. Polar Biol 19:293–301
Hop H, Welch HE, Crawford RE (1997) Population structure and feeding ecology of Arctic cod (Boreogadus saida) schools in the Canadian High Arctic. In: Reynolds J (ed) Fish Ecology in Arctic North America, Am Fish Soc Symp, vol 19, pp 68–80, Bethesda
Hyvärinen H (1989) Diving in darkness—whiskers as sense-organs of the ringed seal (Phoca-hispida-saimensis). J Zool 218:663–678. doi:10.1111/j.1469-7998.1989.tb05008.x
Jarvela LE, Thorsteinson LK (1999) The epipelagic fish community of Beaufort Sea coastal waters, Alaska. Arctic 52:80–94
Kelly BP, Wartzok D (1996) Ringed seal diving behavior in the breeding season. Can J Zool 74:1547–1555. doi:10.1139/z96-169
Kunnasranta M, Hyvarinen H, Hakkinen J, Koskela JT (2002) Dive types and circadian behaviour patterns of Saimaa ringed seals Phoca hispida saimensis during the open-water season. Acta Theriol 47:63–72
Lampert W (1989) The adaptive significance of diel vertical migrations. Funct Ecol 3:21–27
Langlois A, Mundy CJ, Barber DG (2006) On the winter evolution of snow thermophysical properties over land-fast first-year sea ice. Hydrol Process 21:705–716. doi:10.1002/hyp.6407
Lapota D, Geiger ML, Stiffey AV, Rosenberger DE, Young DK (1989) Correlations of planktonic bioluminescence with other oceanographic parameters from a Norwegian fjord. Mar Ecol Prog Ser 55:217–227
Lear WH (1979) Distribution, size and sexual maturity of Arctic cod (Boreogadus saida) in the Northwest Atlantic during 1959–1978. Can Atl Fish Sci Adv Comm Res Doc 17, Fisheries and Oceans Canada, St. John’s
Lønne OJ, Gulliksen B (1989) Size, age and diet of polar cod, Boreogadus saida (Lepechin 1773), in ice covered waters. Polar Biol 9:187–191
Lydersen C, Hammill MO (1993) Diving in ringed seal (Phoca hispida) pups during the nursing period. Can J Zool 71:991–996. doi:10.1139/z93-131
Pearre S (2003) Eat and run? The hunger/satiation hypothesis in vertical migration: history, evidence and consequences. Biol Rev 78:1–79. doi:10.1017/S146479310200595X
Perovich DK (1996) The optical properties of sea ice. US Army Corps of Engineers. Cold Regions Research and Engineering Laboratory. Monograph 96-1
Pitcher TJ, Parrish JK (1993) Functions of shoaling behaviour in teleosts. In: Pitcher TJ (ed) The behaviour of teleost fishes. Chapman & Hall, London, pp 363–427
Pitcher TJ, Lang SH, Turner JA (1988) A risk-balancing trade off between foraging rewards and predation hazard in a shoaling fish. Behav Ecol Sociobiol 22:225–228
Ponomarenko VP (1968) Some data on the distribution and migrations of polar cod in the seas of the soviet Arctic. Rapp Procès Verbaux Réunions CIEM 158:131–135
Rass TS (1968) Spawning and development of polar cod. Rapp P-V Réun Cons Perm Cons Int Explor Mer 158:135–137
Ringelberg J (1995) Changes in light intensity and diel vertical migration: a comparison of marine and freshwater environments. J Mar Biol Ass UK 75:15–25
Ryer CH, Olla BL (1998) Effect of light on juvenile walleye pollock shoaling and their interaction with predators. Mar Ecol Prog Ser 167:215–226
Saether BS, Christiansen JS, Jobling M (1999) Gastrointestinal evacuation of particulate matter in polar cod Boreogadus saida. Mar Ecol Prog Ser 188:201–205. doi:10.3354/meps188201
Smith TG (1987) The ringed seal, Phoca hispida, of the Canadian Western Arctic. Can Bull Fish Aquat Sci 216, Ottawa
Sourisseau M, Simard Y, Saucier FJ (2008) Krill diel vertical migration fine dynamics, nocturnal overturns, and their roles for aggregation in stratified flows. Can J Fish Aquat Sci 65:574–587. doi:10.1139/F07-179
Teilmann J, Born EW, Acquarone M (1999) Behaviour of ringed seals tagged with satellite transmitters in the North Water Polynya during fast-ice formation. Can J Zool 77:1934–1946. doi:10.1139/cjz-77-12-1934
Tester PA, Cohen JH, Cervetto G (2004) Reverse vertical migration and hydrographic distribution of Anomalocera ornate (Copepoda: Pontellidae) in the US South Atlantic Bight. Mar Ecol Prog Ser 268:195–203
Welch HE, Bergmann MA, Siferd TD, Martin KA, Curtis M, Crawford RE, Conover RJ, Hop H (1992) Energy flow through the marine ecosystem of the Lancaster Sound region, Arctic Canada. Arctic 45:343–357
Welch HE, Crawford RE, Hop H (1993) Occurrence of Arctic cod (Boreogadus saida) schools and their vulnerability to predation in the Canadian high arctic. Arctic 46:331–339
Widder EA, Greene CH, Yougbluth MJ (1992) Bioluminescence of sound-scattering layers in the Gulf of Maine. J Plankton Res 14:1607–1624. doi:10.1093/plankt/14.11.1607
Acknowledgments
This study was conducted within the framework of the Canadian Arctic Shelf Exchange Study (CASES: http://www.cases.quebec-ocean.ulaval.ca/), a Research Network funded by the Natural Sciences and Engineering Research Council of Canada (NSERC) and the Canada Foundation for Innovation, with additional support from Fisheries and Oceans Canada (Canadian Coast Guard), the US National Science Foundation, and the Japanese Ministry of Education, Culture, Sports, Science and Technology. We thank the officers and crew of the CCGS Amundsen for enthusiastic and professional assistance at sea, and the team of technicians, assistants, and colleagues who contributed to the collection of acoustic data, zooplankton, and fish samples, and to laboratory analyses. Zooplankton identification was partially funded by the Arctic Ocean Diversity Census of Marine Life Project. Special thanks to Y. Gratton for the validated physical data and T. Papakyriakou for the light data. Haakon Hop and an anonymous referee provided useful suggestions. This is a contribution to the program of Québec-Océan at Université Laval, the Canada Research Chair on the response of marine arctic ecosystems to climate warming, and the Fisheries and Oceans Canada Research Chair in marine acoustics applied to resources and ecosystem.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Online Resource 1. 38-kHz unedited acoustic record (S v) available from 22 December to 29 May. Black circles represent the incident PAR recorded at the snow surface. Each month of the time-series is provided in a separate file.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Benoit, D., Simard, Y., Gagné, J. et al. From polar night to midnight sun: photoperiod, seal predation, and the diel vertical migrations of polar cod (Boreogadus saida) under landfast ice in the Arctic Ocean. Polar Biol 33, 1505–1520 (2010). https://doi.org/10.1007/s00300-010-0840-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-010-0840-x