
Abstract
Stenotrophomonas maltophilia is an environmental bacterium that has gained a lot of attention, as a nosocomial pathogen associated with significant mortality rates. Biofilm formation is considered the corner stone for establishing infections in many bacteria including S. maltophilia. The aim of this study was the genotypic characterization of the different virulence-associated genes and the investigation of the effect of ascorbic acid on S. maltophilia biofilm formation. A total of 20 S. maltophilia isolates from different sources were included in this study. Genes encoding different virulence factors were investigated genotypically. These included stmPr1, stmPr2, smlt3773 locus, smf-1, rpfF, rmlA and spgM. Biofilm formation was investigated phenotypically. The effect of ascorbic acid on biofilm formation was investigated using MIC as well as sub-inhibitory concentrations. Many of the isolates harbored both serine proteases genes stmPr-1 and stmPr-2. Fourteen (70%) of the 20 isolates carried stmPr-1 and 15 (75%) had stmPr-2. Most of the isolates (95%) possessed smlt-3773 locus. Genes linked to biofilm formation such as smf-1, rpfF, rmlA and spgM, were found in (90%), (45%), (85%) and (30%) of the isolates, respectively. Phenotypically, all S. maltophilia isolates (100%) were biofilm producers. Fifteen (75%) were strong biofilm producers and 5 (25%) were moderate biofilm producers. In attempts to seek a non-chemotherapeutic alternative that can hinder biofilm formation without provoking antimicrobial resistance, the results, herein, showed that ascorbic acid inhibits biofilm formation in a dose-dependent manner.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Stenotrophomonas maltophilia is a non-fermenting, Gram-negative bacilli. It is an obligate aerobe that is motile with polar flagella [1]. Originally an environmental bacterium, it has gained a lot of attention as a nosocomial pathogen associated with significant mortality rates [2]. However, reports of cases of community acquired infections have also been implicated [1].
Although not abundantly virulent, S. maltophilia possess various virulence-associated factors and it is able to persist on different surfaces due to biofilm formation [2]. These virulence factors include extracellular proteases and esterase [2]. There are two important serine proteases StmPr1 and StmPr2 both of which are substrates for Xps, a type II secretion system (T2SS) [3]. Other virulence factors include fimbriae which are crucial for adherence to surfaces and biofilm formation [4]. Moreover, extracellular polysaccharides (EPS) are essential for biofilm structure. In fact, biofilms are held by these polysaccharides to form multi-layered well-organized structures [5].
Stenotrophomonas maltophilia biofilms are considered a key player in the bacterium ability to cause diseases [6]. Biofilms act as a shield for bacteria within, they guard the bacteria from antibacterial agents and the host immune responses [7]. Moreover, they allow these bacteria to persist and colonize different surfaces including the surfaces of medical equipment, from which they are very challenging to remove [6]. This paves the way for S. maltophilia to establish different infections especially in hospital settings [7].
To overcome this challenge, many agents have been investigated to reduce biofilm formation, to achieve utmost clinical outcomes [2]. Vitamin C (ascorbic acid) has been shown to disrupt biofilm formation in some bacterial species [8, 9]. However, to the best of our knowledge its effect on S. maltophilia biofilms has not been investigated, yet. Hence, the aim of this study was the genotypic characterization of the different virulence-associated genes and the investigation of the effect of ascorbic acid on S. maltophilia biofilm formation.
Materials and Methods
Sample Collection
Stenotrophomonas maltophilia isolates were obtained from microbiology laboratories in different hospitals, in Alexandria, Egypt during a period of 8 months. Identification of the collected isolates was initially done using conventional biochemical methods, then confirmed by Vitek-2 (bioMérieux, France). Susceptibility testing for the isolates was carried out using disc diffusion method according to the CLSI guidelines [10], the antibiotics used were sulfamethoxazole/trimethoprim, minocycline and levofloxacin.
Investigation of Virulence Genes
The bacterial DNA was extracted by boiling method, as described previously [11]. PCR was used to detect different virulence-associated genes. These include genes which encode protease enzymes (stmPr-1, stmPr-2), esterase enzyme (smlt3773 locus) and genes-associated with biofilm formation (smf-1, spgM, rmlA and rpfF). The different primers used in this study are detailed in Table (T-1) in supplementary document (SD1), together with the corresponding amplicon sizes, annealing temperatures and target genes. All primers were purchased from Invitrogen (Thermo Fisher Scientific, California, USA). The PCR master mix used was DreamTaq Green PCR Master Mix (Thermo Fisher Scientific, California, USA). PCR was performed on Biometra T-personal Thermal cycler (Analytik Jena GmbH, Jena, Germany). The amplification was done as follows: activation at 95 °C for 3 min, then 40 cycles of denaturation, annealing and extension, followed by a final extension step at 72 °C for 7 min. The 40 cycles included: denaturation at 95 °C for 30 s, annealing for 30 s, at the temperatures demonstrated in supplementary document (SD1), and extension at 72 °C for 1 min per 1000 bp. For the detection of each gene, we used 12.5 μl of the master mix, 1 μl of each primer (the concentration of each primer was 10 pmol), 2 μl of the previously extracted bacterial DNA, and sterile nuclease-free water was added so that the total volume of the reaction becomes 25 μl. Then, the amplicons were separated using gel electrophoresis on (2%) agarose gel, which contained 0.5 μg/mL ethidium bromide, and the bands were visualized using UV transilluminator (Bio-Rad, California, USA).
Determination of the Antimicrobial Activity of Ascorbic Acid Against Planktonic Culture
The minimum inhibitory concentrations (MICs) of ascorbic acid, against the S. maltophilia clinical isolates and S. maltophilia ATCC 13,637 (Oxoid, London, UK), were determined using broth microdilution method [12]. The range of concentrations of ascorbic acid used was 100 to 0.09765 mg/ml. For each tested isolate, the inoculum was spectrophotometrically adjusted to 1.5 × 108 CFU/ml (OD600 0.12–0.13) and diluted to create a final concentration of 5 × 105 CFU/ml in the microtiter plate [13, 14]. Then, the plates were incubated for 18–20 h at 37 °C. This was performed in triplicates [15].
Determination of Biofilm-Forming Capacity and Antibiofilm Activity of Ascorbic Acid
Each well, of 96-well flat-bottomed microtiter plate, received 100 µl of ascorbic acid solution (1/8 MIC, 1/4 MIC and 1/2 MIC and MIC) and 100 µl of overnight broth subculture of the tested isolate diluted in sterile trypticase soy broth (TSB) (Oxoid, London, UK) corresponding to 1.5 × 108 CFU/ml [13, 14]. This was performed in triplicates [15]. Positive controls containing no ascorbic acid and negative controls containing no bacteria were included [15]. The plates were incubated at 35 °C for 24 h [15]. Then, the medium was discarded and each well was washed three times with phosphate buffer saline (PBS) (pH 7.2) (Sigma-Aldrich, Milan, Italy) and biofilm samples were fixed by incubating the microtiter plates at 60 °C for 1 h [16, 17]. The wells were stained with crystal violet [17]. Then, the dye, which is bound to the biofilm was extracted with ethanol 99.5% [16]. The optical density (OD) of each well was measured at 590 nm and the isolates were categorized as non-biofilm producers, weak, moderate or strong biofilm producers based on the measured OD, as described before by Stepanovic et al [18]. Then, the percentage of the inhibition of the biofilm formation was calculated as described previously by Jadhav et al. [19].
Scanning Electron Microscope (SEM)
One of the isolates (S5) was grown in TSB and TSB supplemented with MIC (3.125 mg/ml) of ascorbic acid in six-well polystyrene plate. The SEM examination and image capturing was carried out using JSM-IT200 (JEOL, Japan), after fixing the adherent cells in 2.5% glutaraldehyde in PBS (PH 7.2), gradual dehydration by ethanol and gold coating the samples as described before by Gad et al [20].
Statistical Analysis
Statistical analysis of the data were performed using IBM SPSS software package version 20.0. (Armonk, NY: IBM Corp) Significance of the results was assessed at the 5% level. The used tests were Chi-square test and Fisher’s Exact. One way ANOVA test was done using Post Hoc Test (Tukey), and the significance of the results was, also, assessed at the 5% level.
Results
A total of 20 S. maltophilia isolates, were collected from different clinical samples, including respiratory tract infections, blood stream infections and wound infections. The different type of samples corresponding to each isolate is shown Table (T-2) in supplementary document (SD2).
The susceptibility of the different strains to sulphamethoxazole/trimethoprim, levofloxacin and minocycline is shown in Table 1.
Genotypic Investigation of Virulence Genes
Genotypic investigation showed that the genes responsible for proteases and esterase activity were present among the isolates. We found that the serine proteases genes stmPr1 (1621 bp), stmPr1(868 bp) and stmPr2 were present in 70%, 15% and 75%, respectively. On the other hand, esterase coding gene smlt-3773 locus was present in 95% of the isolates as shown in Table 2. Moreover, Genes associated with biofilm formation were present. smf-1, rpfF, rmlA and spgM, were found in (90%), (45%), (85%) and (30%) of the isolates, respectively, as demonstrated in Table 2. The figures of the different bands are shown in supplementary document (SD3).
Correlation Between Biofilm-Forming Ability and the Presence of Genes Associated with BIOFILM Formation
The extent of biofilm formation among S. maltophilia isolates was very high. The majority of the isolates were strong biofilm producers 15 (75%) and 5 (25%) were moderate biofilm producers. None of the isolates were either weak biofilm producer nor non- biofilm producer. The biofilm producing ability of each isolate is demonstrated in Table 3.
Statistically, there was no significant correlation between the presence of smf-1, rpfF, rmlA or spgM and the extent of biofilm formation, and this is shown in Table 4.
The Effect of Ascorbic Acid on Planktonic Culture and its Biofilm Inhibition Activity
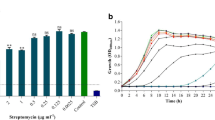
The MIC values of ascorbic acid against the S. maltophilia clinical isolates and S. maltophilia ATCC 13,637 ranged from 0.78 to 50 mg/ml. The modal MIC was 3.125 mg/ml. The MIC50 and MIC90 was 3.125 mg/ml and 6.25 mg/ml, respectively, the MIC value for each isolate is shown in Table 5. The MIC value of ascorbic acid against S. maltophilia ATCC 13,637 was 1.5625 mg/ml, as shown in Table 5.
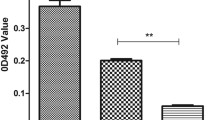
To determine the inhibitory effect of ascorbic acid on biofilm formation, the biofilm-forming ability of S. maltophilia was determined in the presence of variable concentrations of ascorbic acid as well as in the absence of ascorbic acid. The inhibition of biofilm formation was found to be concentration dependent. Similar concentration-dependent manner was shown by S. maltophilia ATCC 13,637, as shown in Fig. 1 and the Table (T-3) in supplementary document SD4. The highest percentage of biofilm inhibition was more evident with MIC, this was shown by the SEM in Fig. 2.

The effect of ascorbic acid on the biofilm-forming capacity of S. maltophilia. Three replica for each group; Data were expressed using Mean ± SD. (SE); SD standard deviation, SE standard error of mean, F F for one way ANOVA test, Pairwise comparison bet. each 2 groups was done using Post Hoc Test (Tukey); p p value for comparing between the studied groups; *: Statistically significant at p ≤ 0.05; a significant with negative control; b significant with positive control; the positive control was the isolate cultured in TSB without ascorbic acid and the negative control was the media without the cultured bacteria

SEM images showing A, B and C which demonstrate isolate (S5) in TSB with different magnifications X2,500, X5000, X10,000, respectively. SEM images showing D, E and F, which demonstrate isolate (S5) in MIC (3.125 mg/ml) of ascorbic acid in TSB with different magnifications X2,500, X5000, X10,000, respectively
Discussion
Stenotrophomonas maltophilia has acquired a lot of interest due to the growing number of nosocomial infections caused by this emerging pathogen [2]. Biofilm formation is considered the corner stone for establishing infections in many bacteria including S. maltophilia [7].
The aim of this study was the genotypic characterization of the different virulence-associated genes and the investigation of the effect of ascorbic acid on S. maltophilia biofilm formation.
Many of the isolates harbored both serine proteases genes; 70% of the isolates harbored stmPr1 (K279a allelic variant of SmtPr1), and 75% harbored stmPr-2. Major (StmPr1) and minor (StmPr2) serine proteases are secreted by Xps, a T2SS in S. maltophilia [3]. Duan et al., showed that stmPr1 and stmPr2 were present in (79.6%) and (95.4%), respectively [21].
Most of the isolates possessed smlt-3773 locus which codes for an esterase enzyme. Nicoletti et al., demonstrated that among all their isolates only three environmental isolates did not harbor smlt3773 locus [4]. Duan et al., reported that smlt3773 locus was present in (52.7%) of their isolates, while in another chinese study, it was found in (91.3%) of the S. maltophilia from pediatric patients [21, 22].
Most of the isolates harbored smf-1, which encodes for type-1 fimbriae, which plays a significant role in adherence to surfaces and the early stages of biofilm formation [2, 23]. Nicoletti et al., reported that smf-1 was present in all their clinically derived S. maltophilia isolates [4]. Azimi et al., showed that smf-1 was present in (99.3%) of their isolates [13].
rpfF plays a critical role in the production of the diffusable signal factor (DSF), which mediates quorum sensing in S. maltophilia. Moreover, disruption to rpfF hampers DSF synthesis and inhibits levels of extracellular proteases [16, 24]. On the other hand, spgM encodes an enzyme with both phosphoglucomutase (PGM) activity as well as a phosphomannomutase activity. spgM has been implemented in biofilm formation [6]. rmlA is aslo associated with attachement and biofilm formation [6]. Among the 20 isolates, rpfF, rmlA and spgM were also investigated, they were found in (45%), (85%) and (30%) of the isolates, respectively. Azimi et al., demonstrated that rpfF, rmlA and spgM were present in (70%), (98%), and (97.3%), respectively [13]. Bostanghadiri et al., reported that rpfF, rmlA and spgM were present in (89.41%), (84.71%) and (100%), respectively [25]. Another study, reported that rpfF, rmlA and spgM were present in (45.2%), (83.7%) and (100%), respectively [22].
The ability to form biofilm was predominant among the 20 S. maltophilia isolates; all the isolates were biofilm producers; 75% were strong biofilm producers and 25% were moderate biofilm producers. Several reports have also witnessed an increased incidence of biofilm-forming ability among their isolates. Azimi et al. and Bostanghadiri et al. reported that (98.7%) and (95.7%) of their isolates, respectively, were biofilm producers with variable capacity of biofilm formation [5, 13]. Zhou et al. showed predominance of biofilm formation (100%) among their isolates, where only 2 isolates were classified as weak biofilm producers [6]. A study based on five European countries by Pompolio et al. revealed that (91.7%) of the isolates tested were biofilm producers categorized into different groups according to the extent of biofilm formation [17].
We found that there was no significant correlation between the presence of smf-1, rpfF, rmlA or spgM and the extent of biofilm formation.
Biofilms allow bacterial cells, to adhere and persist on both biotic and abiotic surfaces, and consequently to establish infections in both community and hospital settings [2]. Different studies have tackled hindering biofilm formation, however many of them used antibacterial agents which may further complicate the antimicrobial resistance challenge [26, 27]. In this context, we decided to evaluate the ability of ascorbic acid to inhibit biofilm formation among the 20 S. maltophilia clinical isolates as well as S. maltophilia ATCC 13,637. Ascorbic acid, also known as vitamin C, is antioxidant and a micronutrient, which is needed to sustain general health and immune system functions [9].
The MIC values of ascorbic acid ranged from 0.78 to 50 mg/ml. The MIC50 and MIC90 was 3.125 mg/ml and 6.25 mg/ml respectively. Kwiecińska-Piróg et al. and Verghese et al. showed that a concentration of 10 mg/ml of ascorbic acid can efficiently inhibit growth among Enterobacteriaceae [28, 29]. Another study conducted by Mumtaz et al., demonstrated that Vitamin C notably hindered the growth of some Gram-positive as well as some Gram-negative bacteria including members of the Enterobacteriaceae family as well as P. aeruginosa [30]. Other reports also investigated the inhibitory effect of vitamin C on several other bacteria including M. tuberculosis and H. pylori, none of which tested the effect of vitamin C on S. maltophilia [31,32,33]. To the best of our knowledge, this is the first study to evaluate the effect of ascorbic acid on S. maltophilia biofilm formation. Ascorbic did not induce biofilm formation, at sublethal doses, in fact it inhibited biofilm formation in S. maltophilia in a concentration-dependent manner. Interestingly, the highest percentage of biofilm inhibition was more evident with MIC. This is consistent with previous reports by Mumtaz et al. [30] and Eydou et al. [9].
Conclusion
Biofilm formation in S. maltophilia represents a therapeutic and a clinical challenge. Biofilm inhibition remains a reasonable solution to meet this challenge. Here, we conclude that Vitamin C can inhibit biofilm formation in S. maltophilia in a concentration-dependent manner. Selecting biofilm inhibitors such as vitamin C and other non-chemotherapeutic agents, spares the use of common antibacterial agents as biofilm inhibitors which paves the way for better clinical outcomes.
Data Availability
Not applicable.
Code Availability
Not applicable.
References
Brooke JS (2012) Stenotrophomonas maltophilia: an emerging global opportunistic pathogen. Clin Microbiol Rev 25(1):2–41. https://doi.org/10.1128/CMR.00019-11
Flores-Trevino S, Bocanegra-Ibarias P, Camacho-Ortiz A, Morfin-Otero R, Salazar-Sesatty HA, Garza-Gonzalez E (2019) Stenotrophomonas maltophilia biofilm: its role in infectious diseases. Expert Rev Anti Infect Ther 17(11):877–893. https://doi.org/10.1080/14787210.2019.1685875
DuMont AL, Karaba SM, Cianciotto NP (2015) Type II secretion-dependent degradative and cytotoxic activities mediated by Stenotrophomonas maltophilia serine proteases StmPr1 and StmPr2. Infect Immun 83(10):3825–3837. https://doi.org/10.1128/IAI.00672-15
Nicoletti M, Iacobino A, Prosseda G, Fiscarelli E, Zarrilli R, De Carolis E, Petrucca A, Nencioni L, Colonna B, Casalino M (2011) Stenotrophomonas maltophilia strains from cystic fibrosis patients: genomic variability and molecular characterization of some virulence determinants. Int J Med Microbiol 301(1):34–43. https://doi.org/10.1016/j.ijmm.2010.07.003
Bostanghadiri N, Ghalavand Z, Fallah F, Yadegar A, Ardebili A, Tarashi S, Pournajaf A, Mardaneh J, Shams S, Hashemi A (2019) Characterization of phenotypic and genotypic diversity of S. maltophilia strains isolated from selected hospitals in Iran. Front Microbiol 10:1191. https://doi.org/10.3389/fmicb.2019.01191
Zhuo C, Zhao QY, Xiao SN (2014) The impact of spgM, rpfF, rmlA gene distribution on biofilm formation in Stenotrophomonas maltophilia. PLoS ONE 9(10):e108409. https://doi.org/10.1371/journal.pone.0108409
Ciofu O, Rojo-Molinero E, Macia MD, Oliver A (2017) Antibiotic treatment of biofilm infections. APMIS 125(4):304–319. https://doi.org/10.1111/apm.12673
Pandit S, Ravikumar V, Abdel-Haleem AM, Derouiche A, Mokkapati V, Sihlbom C, Mineta K, Gojobori T, Gao X, Westerlund F, Mijakovic I (2017) Low concentrations of vitamin C reduce the synthesis of extracellular polymers and destabilize bacterial biofilms. Front Microbiol 8:2599. https://doi.org/10.3389/fmicb.2017.02599
Eydou Z, Jad BN, Elsayed Z, Ismail A, Magaogao M, Hossain A (2020) Investigation on the effect of vitamin C on growth & biofilm-forming potential of Streptococcus mutans isolated from patients with dental caries. BMC Microbiol 20(1):231. https://doi.org/10.1186/s12866-020-01914-4
CLSI (2021) Performance standards for antimicrobial susceptibility testing (31st ed). Clinical and Laboratory Standards Institute, Wayne, PA 36 (3):16–38
Yang JL, Wang MS, Cheng AC, Pan KC, Li CF, Deng SX (2008) A simple and rapid method for extracting bacterial DNA from intestinal microflora for ERIC-PCR detection. World J Gastroenterol 14(18):2872–2876. https://doi.org/10.3748/wjg.14.2872
Institute CaLS (2019) Performance Standards for Antimicrobial Susceptibility Testing, 29th ed. CLSI supplement M100. Clinical and Laboratory Standards Institute, Wayne, PA
Azimi A, Aslanimehr M, Yaseri M, Shadkam M, Douraghi M (2020) Distribution of smf-1, rmlA, spgM and rpfF genes among Stenotrophomonas maltophilia isolates in relation to biofilm-forming capacity. J Glob Antimicrob Resist 23:321–326. https://doi.org/10.1016/j.jgar.2020.10.011
Pollini S, Di Pilato V, Landini G, Di Maggio T, Cannatelli A, Sottotetti S, Cariani L, Aliberti S, Blasi F, Sergio F, Rossolini GM, Pallecchi L (2018) In vitro activity of N-acetylcysteine against Stenotrophomonas maltophilia and Burkholderia cepacia complex grown in planktonic phase and biofilm. PLoS ONE 13(10):e0203941. https://doi.org/10.1371/journal.pone.0203941
Madi H, Lukic J, Vasiljevic Z, Biocanin M, Kojic M, Jovcic B, Lozo J (2016) Genotypic and Phenotypic Characterization of Stenotrophomonas maltophilia Strains from a Pediatric Tertiary Care Hospital in Serbia. PLoS ONE 11(10):e0165660. https://doi.org/10.1371/journal.pone.0165660
Alcaraz E, Garcia C, Papalia M, Vay C, Friedman L, Passerini de Rossi B (2018) Stenotrophomonas maltophilia isolated from patients exposed to invasive devices in a university hospital in Argentina: molecular typing, susceptibility and detection of potential virulence factors. J Med Microbiol 67(7):992–1002. https://doi.org/10.1099/jmm.0.000764
Pompilio A, Ranalli M, Piccirilli A, Perilli M, Vukovic D, Savic B, Krutova M, Drevinek P, Jonas D, Fiscarelli EV, Tuccio Guarna Assanti V, Tavio MM, Artiles F, Di Bonaventura G (2020) Biofilm formation among Stenotrophomonas maltophilia isolates has clinical relevance: the ANSELM prospective multicenter study. Microorganisms. https://doi.org/10.3390/microorganisms9010049
Stepanovic S, Cirkovic I, Ranin L, Svabic-Vlahovic M (2004) Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett Appl Microbiol 38(5):428–432. https://doi.org/10.1111/j.1472-765X.2004.01513.x
Jadhav S, Shah R, Bhave M, Palombo EA (2013) Inhibitory activity of yarrow essential oil on Listeria planktonic cells and biofilms. Food Control 29(1):125–130
Gad GF, El-Feky MA, El-Rehewy MS, Hassan MA, Abolella H, El-Baky RM (2009) Detection of icaA, icaD genes and biofilm production by Staphylococcus aureus and Staphylococcus epidermidis isolated from urinary tract catheterized patients. J Infect Dev Ctries 3(5):342–351. https://doi.org/10.3855/jidc.241
Duan Z, Qin J, Liu Y, Li C, Ying C (2020) Molecular epidemiology and risk factors of Stenotrophomonas maltophilia infections in a Chinese teaching hospital. BMC Microbiol 20(1):294. https://doi.org/10.1186/s12866-020-01985-3
Duan Z, Qin J, Li C, Ying C (2020) Clinical and molecular epidemiology of Stenotrophomonas maltophilia in pediatric patients from a Chinese teaching hospital. Front Cell Infect Microbiol 10:411. https://doi.org/10.3389/fcimb.2020.00411
de Oliveira-Garcia D, Dall’Agnol M, Rosales M, Azzuz AC, Alcantara N, Martinez MB, Giron JA (2003) Fimbriae and adherence of Stenotrophomonas maltophilia to epithelial cells and to abiotic surfaces. Cell Microbiol 5(9):625–636. https://doi.org/10.1046/j.1462-5822.2003.00306.x
Huedo P, Yero D, Martinez-Servat S, Estibariz I, Planell R, Martinez P, Ruyra A, Roher N, Roca I, Vila J, Daura X, Gibert I (2014) Two different rpf clusters distributed among a population of Stenotrophomonas maltophilia clinical strains display differential diffusible signal factor production and virulence regulation. J Bacteriol 196(13):2431–2442. https://doi.org/10.1128/JB.01540-14
Bostanghadiri N, Ardebili A, Ghalavand Z, Teymouri S, Mirzarazi M, Goudarzi M, Ghasemi E, Hashemi A (2021) Antibiotic resistance, biofilm formation, and biofilm-associated genes among Stenotrophomonas maltophilia clinical isolates. BMC Res Notes 14(1):151. https://doi.org/10.1186/s13104-021-05567-y
Sperandio V (2007) Novel approaches to bacterial infection therapy by interfering with bacteria-to-bacteria signaling. Expert Rev Anti Infect Ther 5(2):271–276. https://doi.org/10.1586/14787210.5.2.271
Kim HR, Lee D, Eom YB (2018) Anti-biofilm and anti-virulence efficacy of celastrol against Stenotrophomonas maltophilia. Int J Med Sci 15(6):617–627. https://doi.org/10.7150/ijms.23924
Kwiecinska-Pirog J, Skowron K, Bogiel T, Bialucha A, Przekwas J, Gospodarek-Komkowska E (2019) Vitamin C in the presence of sub-inhibitory concentration of aminoglycosides and fluoroquinolones alters Proteus mirabilis biofilm inhibitory rate. Antibiotics. https://doi.org/10.3390/antibiotics8030116
Verghese RJ, Mathew SK, David A (2017) Antimicrobial activity of Vitamin C demonstrated on uropathogenic Escherichia coli and Klebsiella pneumoniae. J Curr Res Sci Med 3(2):88
Mumtaz S, Mumtaz S, Ali S, Tahir HM, Kazmi SAR, Mughal TA, Younas M (2021) Evaluation of antibacterial activity of vitamin C against human bacterial pathogens. Braz J Biol 83:e247165. https://doi.org/10.1590/1519-6984.247165
Isela N-NR, Sergio N-C, Martínez-Sanmiguel JJ, Hernandez-Delgadillo R, Cabral-Romero C (2013) Ascorbic acid on oral microbial growth and biofilm formation. Pharma Innovat 2((4, Part A)):103
Vilcheze C, Hartman T, Weinrick B, Jacobs WR Jr (2013) Mycobacterium tuberculosis is extraordinarily sensitive to killing by a vitamin C-induced fenton reaction. Nat Commun 4:1881. https://doi.org/10.1038/ncomms2898
Majtan J, Sojka M, Palenikova H, Bucekova M, Majtan V (2020) Vitamin C enhances the antibacterial activity of honey against planktonic and biofilm-embedded bacteria. Molecules. https://doi.org/10.3390/molecules25040992
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
Both authors conceived, designed, and performed the experiments. Both authors wrote and revised the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Ethical Approval
Ethical approval was obtained from the Ethics Committee, Faculty of Pharmacy, Pharos University in Alexandria (PUA). Ethical Approval Number: 01202106203028.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
ElBaradei, A., Yakout, M.A. Stenotrophomonas maltophilia: Genotypic Characterization of Virulence Genes and The Effect of Ascorbic Acid on Biofilm Formation. Curr Microbiol 79, 180 (2022). https://doi.org/10.1007/s00284-022-02869-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00284-022-02869-7