
Abstract
Purposes
Strontium ranelate (SrRN) is a novel and effective anti-osteoporotic drug used to promote bone formation and restrain bone resorption. However, studies of the effects and mechanism of SrRN on adipocytic differentiation from bone marrow mesenchymal stem cells (BMMSCs) remain limited.
Methods
The cytoactivity of BMMSCs was analyzed by CCK-8 and annexin V-FITC/PI kit. Then the differentiation efficiency was analyzed by Oil Red O staining and western blotting. The level of autophagy was evaluated by immunofluorescence and western blotting. The autophagy inhibitor chloroquine and the selective Akt antagonist API-2 were used to study the relationship between autophagy regulation and adipocytic differentiation. Finally variations in β-catenin protein levels were also analyzed by western blotting, as were proteins related with autophagy and differentiation signaling pathways.
Results
SrRN had no influence on the vatility of BMMSCs (bone marrow mesenchymal stem cells) and exhibited anti-adipocytic effects in dose-dependent manner. At the early stage of adipocytic differentiation, autophagy level significantly increased and SrRN inhibited the variation tendency. Then we detected the PPAR-γ and β-catenin variation when applied with autophagy inhibitor CQ (chloroquine) and the same way, after Akt selective inhibitor applied in the test, the effect of SrRN was compromised. At last, we detected the increased phosphorylation of Akt-related pathway at 1.00 mM concentration of SrRN in comparison with the adipocytic induction.
Conclusions
SrRN could disturb the process of adipocytic differentiation of BMMSCs and interfere with the variation tendency of autophagy level after adipocytic induction in Akt-dependent manners.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Osteoporosis is usually characterized by increased bone resorption and/or decreased bone formation, and has been a growing epidemiological problem in our rapidly aging society. Osteoporosis-related fractures are a cause of chronic disability and are a cause of increased risk of death. Recent research has revealed that most patients suffering from osteoporosis, including osteoporosis accompanied by fracture, do not receive adequate pharmacotherapy.
Strontium ranelate (SrRN) is both an anti-resorptive and an anabolic drug that has been indicated for oral administration to treat osteoporosis. Osteoblasts have been found to play key roles in the mechanisms of action of SrRN, by upregulating the mRNA and protein expression of osteoprotegerin and downregulating those of receptor activator of NF-κB ligand. Osteoblasts are derived from bone marrow mesenchymal stem cells (BMMSCs) present in the bone marrow stroma, which are able to differentiate into a number of lineages, including osteoblasts, chondroblasts, and adipocytes. Recent studies on bone marrow adipocytes have shown that bone marrow fat (BMF) is only a simple filling tissue, but is also an indication of bone fragility. Bone marrow adipocytes share the same precursors as osteoblasts, indicating a theoretical inverse relationship between adipocytic and osteogenic lineage commitment and differentiation [1, 2]. The differentiation of BMMSCs towards the osteoblast phenotype usually occurs at the expense of the adipocyte phenotype. Recently, it has been proposed that the balance between osteogenic and adipogenic MSC differentiation is disrupted in osteoporosis [3, 4]. It is known that high BMF may be associated with weak bone mass in osteoporosis, especially during aging and post-menopause; we therefore designed a study to research the effects of SrRN on adipogenic differentiation of BMMSCs.
A large number of molecular events are involved in adipocytic differentiation of BMMSCs, including a series of complicated physiological processes. However, we know little about the cell-intrinsic mechanisms of this cell lineage commitment to differentiate. Autophagy is also a complicated process which degrades intracellular components by formation of autophagosomes, which then fuse with lysosomes to form autolysosomes [5]. Growing evidence suggests that autophagy is implicated in lipid accumulation and adipocytic differentiation [6,7,8]. The Wnt/β-catenin pathway, and especially β-catenin, has been associated with the degradation effects of autophagy [9, 10]. The present study was designed to investigate the relationship between the effects of SrRN on adipocytic differentiation from BMMSCs and autophagy regulation of β-catenin at the protein level.
Materials and methods
Cell culture
Rat BMMSCs were purchased from Cygen Biosciences Biotech (Guangzhou, China). Cells were cultured in basal medium (DMEM/F12 1:1) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin. Cells were plated into 25-cm2 culture flasks and incubated at 37 °C in a 95% humidified atmosphere with 5% CO2. Culture medium was replaced every two days. When adherent cells reached 80% confluence, they were detached by mild treatment with trypsin-EDTA (0.25%/0.02%) and replaced at 1/3 density for continued culture. Cells passaged eight times were used for subsequent experiments. Rat BMMSCs were treated with SrRN (Sigma-Aldrich, St Louis, MO, USA) at different concentrations (0.00, 0.10, 0.50, or 1.00 mM) in adipocytic medium. Adipocytic differentiation medium was used to induce differentiation of BMMSCs.
CCK-8 cell proliferation assay
BMMSCs were plated in 96-well plates at a density of 1 × 103 cells/well. After 4 h of cell culture, the basal medium was replaced with fresh medium with or without SrRN (0.00, 0.05, 0.10, 0.50, 1.00, or 2.00 mM). The medium was replaced every two days, and the CCK-8 assay was implemented at different time-points (1, 2, 3, 4, 5, 6, and 7 days). Briefly, to perform the assay the medium was replaced with 100 μL fresh medium, and the 10 μL CCK-8 reagent (Dojindo, Kumamoto, Japan) was added into each well. Cell culture was continued for one hour, then the optical density (OD) was quantified at a wavelength of 420 nm. Samples were assayed in triplicate and the assay was performed in three cultures for each group.
Annexin V-FITC/PI apoptosis detection
To further analyze the effect of SrRN on the viability of BMMSCs, the apoptosis of BMMSCs was detected in cells treated with or without SrRN (0.00, 0.05, 0.10, 0.50, 1.00, or 2.00 mM) for three days using an annexin V-FITC/PI kit (Dojindo). BMMSCs were detached by trypsinization without EDTA and washed twice with cool PBS. The cells were centrifuged at 500×g for five minutes and resuspended in 500 μL binding buffer, then 5 μL annexin V-FITC and 5 μL PI were added and the cells were incubated in the dark at room temperature for 15 minutes. The samples were analyzed immediately using a FACSCalibur flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA) and the data were analyzed using CellQuest software.
Adipocytic differentiation of BMMSCs: Oil Red O staining and triglyceride accumulation
To investigate the anti-adipocytic effect of SrRN, BMMSCs were plated into 60 mm petri dishes at a density of 5 × 105 cells/3 mL. After 48 hours of continuous culture, the medium was replaced by adipocytic medium containing different concentrations of SrRN. The adipocytic medium consisted of serum-free DMEM (glucose 4.5 g/L) supplemented with 10 μg/mL insulin, 0.5 mM 3-isobutyl-1-methylxanthine, 200 μM indomethacin, and 1 μM dexamethasone [11]. The medium was changed every two days, and after two weeks of treatment adipogenesis was evaluated by Oil red O staining and triglyceride quantification.
Oil Red O staining and quantitative analysis of Triglyceride
After 14 days of induction and treatment with or without SrRN, cells were washed with PBS three times and fixed in 4% paraformaldehyde for ten minutes at room temperature, washed with distilled water and 60% isopropanol, stained with newly prepared oil red O for ten minutes, and finally restained with Mayer’s haematoxylin. Samples were observed under an inverted phase contrast microscope. Cells were then destained in 1 mL 100% isopropanol for 15 minutes, and the optical density of the destained solution was measured at 540 nm as previously described [12].
Triglyceride quantification test: a triglyceride content determination kit (Solarbio Science & Technology, Beijing, China) was utilized to assay the total triglyceride content of the cell samples after 14 days of induction and treatment with or without SrRN, according to the manufacturer’s protocol. Briefly, 3 × 105 cells were collected and reacted with the reagents provided. The OD was measured spectrophotometrically at a wavelength of 420 nm.
LC-3 and PPAR-gamma immunocytochemistry
BMMSCs were seeded into 24-well plates; after three days of induction and treatment with SrRN (1.00 mM), cells were fixed in 4% paraformaldehyde, permeabilized in 0.5% Triton X-100 (Solarbio Science & Technology) and washed with PBS three times. The samples were blocked in 5% bovine serum albumin (BSA) for one hour at 37 °C, then incubated overnight with rabbit monoclonal anti-LC3B antibody (Abcam, Cambridge, UK). Next, the samples were incubated with a goat anti-rabbit IgG peroxidase secondary antibody and were then counterstained with DAPI. Similarly, BMMSCs induced and treated with or without SrRN were treated with chloroquine (CQ, 20 μM) or API-2 (10 nM) in each group. They were then stained with a rabbit monoclonal anti-PPAR-γ antibody (Abcam, Cambridge, UK) following the above procedure. Finally, the samples were viewed under a fluorescence microscope (Olympus, Japan).
Western blotting
Cell samples were collected, washed and lysed with RIPA lysis buffer supplemented with PMSF (Beyotime, Jiangsu, China) to extract total protein. The protein concentration was quantified using a BCA assay kit (CWBio, Beijing, China). Aliquots of protein (30 μg) were denatured and separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis, then transferred to a PVDF membrane. The membranes were blocked with 5% BSA in TBST for two hours, then incubated with the primary antibodies overnight. The next day, samples were incubated with the secondary antibody (ZSBG-BIO, Beijing, China) at 1:10,000 for two hours then visualized with enhanced chemiluminescence (ECL, Beyotime) and analyzed using Image J software (NIH, Bethesda, MD, USA).
Statistical analysis
Data are presented as mean ± SD, and comparisons of quantitative data were analyzed by one-way ANOVA. P < 0.05 was considered significant, and all experiments were performed at least three times.
Results
SrRN has no effect on viability of BMMSCs
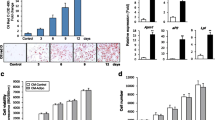
The viability of BMMSCs at different concentrations of SrRN was analyzed by CCK-8 and annexin V assay, and results are shown respectively as a growth curve in Fig. 1a and a bar graph in Fig. 1b. During the 7 days of culture with or without SrRN, the cell growth curve showed no significant differences between groups. Similar results were obtained in the apoptosis assay. After 72 h of cell culture with different concentrations of SrRN, the cell samples were stained and analyzed by flow cytometry. No differences in either early or late apoptosis were found between groups.

SrRN has no effect on viability of BMMSCs and SrRN has anti-adipocytic effect. a Cell growth in BMMSCs at 1–7 days with different concentrations of SrRN, measured by CCK-8, with triplicates of each group; data expressed as mean ± SD. b Flow cytometric analysis of apoptosis in BMMSCs at different drug concentrations, FITC/PI double staining; Q2 and Q4 represent early apoptosis and late apoptosis in each experimental group. c Oil Red O staining of BMMSCs after 14 days in culture with different concentrations of SrRN (b × 200). d Quantitation of the extracted oil red O at 520 nm. e Analysis of triglyceride content in the different groups at 14 days using a triglyceride quantification kit
SrRN has an anti-adipocytic effect
To investigate the anti-adipocytic effect of SrRN on the differentiation of BMMSCs, induction efficiency was compared by Oil Red O staining after 14 days of adipocytic differentiation with or without SrRN treatment (0.00, 0.10, 0.50, or 1.00 mM). Figure 1c shows that SrRN downregulated lipid accumulation. Quantitative results are presented in Fig. 1d, e, confirming this trend. When compared with the control, the results reflected both the anti-adipocytic effect and dose-dependence in the SrRN treatment group.
SrRN downregulates essential adipocytic differentiation factors
After 5 days of induction and treatment with or without SrRN, protein levels of PPAR-γ, an essential protein for adipocytic differentiation of BMMSCs, were detected by fluorescence staining (Fig. 2a). The expression of PPAR-γ in the SrRN treatment group showed dramatic differences, compared with the control group. To confirm this effect, the intracellular proteins PPAR-γ and C/EBP-α, critical proteins in adipocytic induction, were analyzed by western blotting. After 14 days of induction and treatment with or without SrRN (0.00, 0.10, 0.50, or 1.00 mM), the relative expression of PPAR-γ and C/EBP-α proteins was downregulated in all the treatment groups, with the most obvious effect in the 0.50 and 1.00 mM treatment groups (P < 0.01 for both; Fig. 2b).

SrRN downregulates essential adipocytic differentiation factors. a Fluorescence intensity of PPAR-γ. b SrRN significantly reduced the protein expression of PPAR-γ and C/EBP-α in cells after 14 days of adipocytic differentiation with and without SrRN. c Immunofluorescent intensity of the autophagy-related protein, LC3, (× 200) induced by adipocytic differentiation in the presence or absence of SrRN elate
SrRN negatively regulates autophagy in adipocytic differentiation of BMMSCs
In previous studies, autophagy was found during adipocytic differentiation and had a complicated effect on adipogenesis. To investigate the regulatory effect of autophagy and SrRN on the process of adipocytic differentiation, the intracellular expression of LC3 protein was visualized using immunofluorescence technology. After adipocytic induction of BMMSCs with or without SrRN for three days, autophagy levels were reflected by fluorescence intensity (Fig. 2a). SrRN significantly inhibited the expression of LC3 protein, which suggested that SrRN might suppress autophagy in the early stages of adipocytic differentiation. In order to detect dynamic changes in autophagy during early adipocytic differentiation, cell samples were collected at four different time-points (days 1, 3, 5, and 7). Western blotting was used to detect the expression of LC3-II/I and P62 proteins at different time-points (Fig. 3a). Quantitative analysis showed that the level of LC3-II/I was significantly increased in the early stage of adipocytic differentiation, while the expression of was P62 significantly decreased in the induction group (Fig. 3b, c). However, in the treatment group (1.00 mM SrRN) the expression of the autophagy-related proteins, LC3-II/I and P62, exhibited no significant changes over the same period (Fig. 3b, c). These results indicated that the autophagy levels increased in the early stage of adipocytic differentiation and that treatment with SrRN might significantly downregulate autophagy over the same period.

SrRN negatively regulates autophagy in adipocytic differentiation of BMMSCs and positively protect β-catenin by downregulating the autophgy. a Western blotting was used to detect the expression of the autophagy-related proteins, LC3-I/II and P62. b, c SrRN effectively reduces the early increase in autophagy levels during adipocytic differentiation of BMSCs at 1–7 days. d The autophagy inhibitor CQ and the Akt selective antagonist API-2 were respectively added to the induction group and the treatment group. Five days later, the fluorescence intensity of PPAR-γ was visualized. e, f Western blotting was used to detect dynamic changes of β-catenin in the early stages of adipocytic differentiation
Inhibition of autophagy negatively regulates adipocytic differentiation and the effect of SrRN is Akt-dependent
During differentiation from BMMSCs into adipocytes, the autophagy inhibitor CQ and the highly-selective Akt inhibitor API-2 were added to the induction group and treatment group, respectively. As the most critical factor in adipocytic differentiation, PPAR-γ was visualized (fluorescence intensity) five days later (Fig. 3d). The results suggested that CQ might suppress the process of adipocytic differentiation and that API-2 might disturb the anti-adipocytic effect of SrRN during differentiation.
Autophagy-dependent variation of β-catenin protein during adipocytic differentiation of BMMSCs
The Wnt/β-catenin signaling pathway plays an essential role during the osteoblastic and adipocytic differentiation of BMMSCs. Previous studies revealed complex effects of autophagy on adipocytic differentiation. On the one hand, the inhibition of autophagy might interfere with adipocytic differentiation [6, 7], while on the other hand, autophagy might reduce lipid accumulation [13, 14]. By adding an autophagy inhibitor and Akt selective antagonist to the induction and treatment group, respectively, we investigated the connection between autophagy and adipocytic differentiation, as well as the likely mechanism of SrRN associated with these processes. Expression of β-catenin protein was detected and quantified by western blotting (Fig. 3c, e).
In the induction and API-2 groups, β-catenin protein showed a significant and time-dependent decrease. However, in the SrRN and CQ groups, β-catenin levels were stabilized at a relatively high level over the same period (Fig. 3e, f). From 1 to 7 days, the β-catenin protein levels in the treatment and CQ groups also decreased, but the differences were not statistically significant.
The intracellular regulatory mechanism of SrRN
According to previous studies, the mechanism of SrRN is likely linked to the calcium-sensing receptor (CaSR) [15, 16], which then further activates the PI3K/Akt signaling pathway. This suggested that the autophagy-regulatory effect of SrRN might rely on the PI3K/Akt/mTOR signaling pathway. The protein levels of p-Akt, Akt, p-mTOR, mTOR, p-GSK3-β, and GSK3-β were detected by western blotting (Fig. 4a–d). While Akt, mTOR and GSK3-β showed no significant changes within 12–36 h, p-Akt, p-mTOR, and p-GSK3-β exhibited different changes over the same period. During the early stage of adipocytic differentiation, from 12 to 36 hours, the phosphorylation levels of Akt, mTOR, and GSK3-β were obviously higher in the treatment group than in the induction group. In addition, lower phosphorylation levels were observed in the API-2 group compared with the induction group, which suggested that the effect of SrRN might be Akt-dependent.

The intracellular regulatory mechanism of SrRN a Akt and p-Akt, mTOR and P-mTOR, and GSK3-β and p-GSK3-β protein levels in cell samples were detected by western blotting. b, c, d Quantitative analysis of GSK3-β and p-GSK3-β, mTOR and P-mTOR, and Akt and p-Akt
Discussion
We analyzed the proliferative effects of different concentration of SrRN (0.00 to 2.00 mM) on rat BMMSCs and found no obvious difference in proliferation. However, in previous studies, SrRN or strontium chloride exhibited different proliferative effects on BMMSCs at different concentrations [11, 17]. Although, our finding was not consistent with past reports, the results still verified the safety of SrRN below 2 mM.
To investigate the anti-adipocytic effects of SrRN, rat BMMSCs were induced in adipocytic medium with various concentrations of SrRN (0.00, 0.10, 0.50, or 1.00 mM Sr2+). After 14-day induction and culture, lipid, and triglyceride accumulation were detected by Oil Red O staining and quantitative measurement, and critical protein expression was detected by western blotting and immunofluorescence. The results revealed that the anti-adipocytic effects of SrRN are due to intrinsic changes such as downregulation of differentiation. At the same time, the efficiency of anti-adipogenic activity was dose-dependent in the treatment group. Although, studies about the anti-adipogenic effects of SrRN are still limited, similar results were observed in previous studies [11, 18, 19]. A study by Liet al. showed that SrRN decreased the mRNA levels of PPAR-γ2, adipocyte lipid-binding protein 2, and lipoprotein lipase by RT-PCR, as well as the protein expression of PPAR-γ in western-blot analysis after one or two weeks of treatment in adipogenic medium [11]. Fournier et al. not only revealed the anti-adipocytic effects of SrRN but also explained the effects anterior to osteoblastic effects. In their study, SrRN reduced PPAR-γ2 and C/EBP-α mRNA in a dose-dependent manner without affecting RUNX2 gene expression whereas it repressed ALP mRNA. Later (day 5), PPAR-γ2 and CEBP-α mRNA remained inhibited by SrRN, preventing adipocyte lipid accumulation, while Runx2 and ALP mRNA were increased under adipogenic conditions [18]. Saidak et al. explained that the age-related switch of osteoblastic to adipocytic differentiation could be antagonized by SrRN treatment via mechanisms involving NFATc/Maf and Wnt signaling [19]. All these results indicated that Sr inhibited adipocytic differentiation of BMMSCs.
In recent studies, autophagy had been identified as a regulator of cellular differentiation, suggesting that autophagy might regulate the process of adipocytic differentiation [7, 8, 20]. Through genetic inhibition of critical autophagy-related gene 7 (Atg7), the function of autophagy in regulating adipocytic development was examined by Singh et al. and similar results were obtained by Zhang et al. [7, 8]. In their studies, mouse models with a targeted deletion of Atg7 exhibited decreased white adipose mass and enhanced insulin sensitivity, while lipid accumulation and protein levels of adipocyte differentiation factors were inhibited. All these results suggested that adipocytic differentiation was attenuated after inhibition of autophagy [7, 8], and revealed the phenomenon of the accumulation of autophagic vesicles and intracellular lipid-like vesicles [21]. The results of LC3 and P62 expression in our study (Fig. 3a–c) suggested that autophagic levels increased in the early stage of adipocytic differentiation (1–7 days) and obviously higher than the treatment group over the same time period; therefore, SrRN might attenuate changes in autophagy. The adipogenic core factor PPAR-γ was obviously altered with the application of CQ and API-2 (Fig. 3d) and the results explained the crossover effect between adipogenic differentiation and autophagy. These results also revealed the probable regulatory mechanism of SrRN for further study.
The Wnt/β-catenin signaling pathway and autophagy activity interact by mutual feedback regulation. On the one hand, β-catenin suppressed autophagosome formation and directly repressed p62/SQSTM1 (encoding the autophagy adaptor p62) while on the other hand, β-catenin might be selectively degraded via autophagy [9]. Similar results have also been proposed by other researchers [22, 23]. To further investigate anti-adipocytic mechanisms of SrRN, β-catenin expression was detected by western blotting. In SrRN and CQ groups, the β-catenin protein level was obviously higher than in the induction group, which suggested that β-catenin itself might be targeted by autophagic clearance upon adipogenic induction. Since there was an internal crosstalk between β-catenin and PPAR-γ, which suggested that β-catenin might inhibit the expression of PPAR-γ in the differentiated lineage [4, 24], the anti-adipocytic effect of SrRN might be dependent on stabilization of the protein β-catenin.
The CaSR was involved in the anti-osteoporotic effects of SrRN [15, 16]. To further study the intracellular mechanisms, the Akt/mTOR pathway and Akt/GSK3-β pathway were further examined in our study. The Akt/mTOR pathway is essential for the regulation of autophagy. The different phosphorylation levels and trends shown in Fig. 4c and d, suggested that autophagy, at the early stage of adipocytic differentiation, might be regulated by the Akt/mTOR signaling pathway. In the SrRN treatment group, the Akt/mTOR pathway was significantly activated with an increased phosphorylation level, compared with the induction group. When API-2, the selective inhibitor of intracellular protein Akt, was added to the treatment group, the activation by SrRN was suppressed. Guo et al. presented similar results in their recent study about the internal mechanism of SrRN. The PI3K/Akt/mTOR signaling pathway was activated by SrRN to trigger angiogenesis in human umbilical vein endothelial cells [17].
β-catenin is the main functional executor of the Wnt/β-catenin signaling pathway. In our study, the intracellular protein levels of β-catenin in the treatment group were obviously higher than in the induction group, which might be related to the stabilization of β-catenin. The results presented in Fig. 4b suggested that the phosphorylation tendency of GSK3-β decreased upon adipocytic induction, whether with or without SrRN treatment, but phosphorylation levels were significantly higher in the SrRN treatment group. When API-2 was added to the treatment group, the results were significantly altered. The level and tendency were similar to those in the induction group. Rybchyn et al. suggested a similar conclusion, that the anti-osteoporotic effect of SrRN might be Akt-dependent, increasing the nuclear translocation of β-catenin [25].
References
Devlin MJ, Rosen CJ (2015) The bone–fat interface: basic and clinical implications of marrow adiposity. Lancet Diabetes Endocrinol 3(2):141–147
Patsch JM et al (2013) Bone marrow fat composition as a novel imaging biomarker in postmenopausal women with prevalent fragility fractures. J Bone Miner Res Off J Am Soc Bone Miner Res 28(8):1721
Ullah I, Subbarao RB, Rho GJ (2015) Human mesenchymal stem cells—current trends and future prospective. Biosci Rep 35(2)
Yuan, Z., et al.,(2016) PPARγ and Wnt signaling in adipogenic and osteogenic differentiation of mesenchymal stem cells. Current Stem Cell Research & Therapy. 11(3): p. -
Klionsky DJ et al (2016) Guidelines for the use and interpretation of assays for monitoring autophagy, 3rd edn. Autophagy 12(1):1–222
Song BQ et al (2015) Inhibition of notch signaling promotes the adipogenic differentiation of mesenchymal stem cells through autophagy activation and PTEN-PI3K/AKT/mTOR pathway. Cell Physiol Biochem 36(5):1991–2002
Singh R et al (2009) Autophagy regulates adipose mass and differentiation in mice. J Clin Investig 119(11):3329
Zhang Y et al (2009) Adipose-specific deletion of autophagy-related gene 7 (atg7) in mice reveals a role in adipogenesis. Proc Natl Acad Sci U S A 106(47):19860–19865
Petherick, KJ., et al.,(2013) Autolysosomal β-catenin degradation regulates Wnt-autophagy-p62 crosstalk. Embo J. 32(13): p. 1903–1916
Shi ZY et al (2017) Protective effect of autophagy in neural ischemia and hypoxia: negative regulation of the Wnt/beta-catenin pathway. Int J Mol Med 40(6):1699–1708
Li Y et al (2012) Effects of strontium on proliferation and differentiation of rat bone marrow mesenchymal stem cells. Biochem Biophys Res Commun 418(4):725–730
Sen A et al (2015) Adipogenic potential of human adipose derived stromal cells from multiple donors is heterogeneous. J Cell Biochem 81(2):312–319
Singh R, Cuervo AM (2012) Lipophagy: connecting autophagy and lipid metabolism. Int J Cell Biol 2012:282041
Singh R et al (2009) Autophagy regulates lipid metabolism. Nature 458(7242):1131–1135
Caverzasio J (2008) Strontium ranelate promotes osteoblastic cell replication through at least two different mechanisms. Bone 42(6):1131
Coulombe J et al (2004) In vitro effects of strontium ranelate on the extracellular calcium-sensing receptor. Biochem Biophys Res Commun 323(4):1184–1190
Guo X et al (2016) Dose-dependent effects of strontium ranelate on ovariectomy rat bone marrow mesenchymal stem cells and human umbilical vein endothelial cells. Int J Biol Sci 12(12):1511–1522
Fournier C et al (2012) Reduction by strontium of the bone marrow adiposity in mice and repression of the adipogenic commitment of multipotent C3H10T1/2 cells. Bone 50(2):499
Saidak, Z. et al.,(2012) Strontium ranelate rebalances bone marrow adipogenesis and osteoblastogenesis in senescent osteopenic mice through NFATc/Maf and Wnt signaling. Aging Cell. 11(3): p. 467–474
Skop V et al (2014) Autophagy inhibition in early but not in later stages prevents 3T3-L1 differentiation: effect on mitochondrial remodeling. Differ; Res Biol Divers 87(5):220–229
Gruia AT et al (2016) Mesenchymal stromal cells differentiating to adipocytes accumulate Autophagic vesicles instead of functional lipid droplets. J Cell Physiol 231(4):863–875
Gao C et al (2010) Autophagy negatively regulates Wnt signalling by promoting Dishevelled degradation. Nat Cell Biol 12(8):781–790
Wang T et al (2017) Regulation of autophagy inhibition and inflammatory response in glioma by Wnt signaling pathway. Oncol Lett 14(6):7197–7200
Li Y et al (2017) PPAR-γ and Wnt regulate the differentiation of MSCs into adipocytes and osteoblasts respectively. Curr Stem Cell Res Ther 12
Rybchyn MS et al (2011) An Akt-dependent increase in canonical Wnt signaling and a decrease in Sclerostin protein levels are involved in strontium Ranelate-induced osteogenic effects in human osteoblasts. J Biol Chem 286(27):23771–23779
Funding
This work was supported by the National Natural Science Foundation of China.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests
The authors declare that they have no conflict of interest.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Xu, Z., Bai, X. Strontium ranelate-induced anti-adipocytic effects are involved in negative regulation of autophagy in rat bone marrow mesenchymal stem cells. International Orthopaedics (SICOT) 42, 2483–2490 (2018). https://doi.org/10.1007/s00264-018-3960-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00264-018-3960-5