
Abstract
Three sequencing batch reactors (SBRs) were operated in parallel to study the effects of trace erythromycin (ERY) and ERY-H2O on the treatment of a synthetic wastewater. Through monitoring (1) daily effluents and (2) concentrations of nitrogen (N) and phosphorous (P) in certain batch cycles of the three reactors operated from transient to steady states, the removal of carbon, N, and P was affected negligibly by ERY (100 µg/L) or ERY-H2O (50 µg/L) when compared with the control reactor. However, through analyzing microbial communities of the three steady state SBRs on high-density microarrays (PhyloChip), ERY, and ERY-H2O had pronounced effects on the community composition of bacteria related to N and P removal, leading to diversity loss and abundance change. The above observations indicated that resistant bacteria were selected upon exposure to ERY or ERY-H2O. Short-term batch experiments further proved the resistance and demonstrated that ammonium oxidation (56–95%) was inhibited more significantly than nitrite oxidation (18–61%) in the presence of ERY (100, 400, or 800 µg/L). Therefore, the presence of ERY or ERY-H2O (at µg/L levels) shifted the microbial community and selected resistant bacteria, which may account for the negligible influence of the antibiotic ERY or its derivative ERY-H2O (at µg/L levels) on carbon, N, and P removal in the SBRs.
Similar content being viewed by others
Explore related subjects
Find the latest articles, discoveries, and news in related topics.Avoid common mistakes on your manuscript.
Introduction
Macrolide antibiotics are among the most important and common antibacterial agents applied to both human and animal diseases caused by bacteria (Pothuluri et al. 1998; Dubois et al. 2001). Macrolides prevent bacterial growth by binding to the 23S ribosomal RNA (rRNA) of the large 50S ribosomal subunit so as to inhibit protein synthesis in the elongation step (Verdier et al. 2001). Though macrolides can be metabolized by demethylation in the body, the urinary excretion of unchanged parent drugs could reach 5–10% for erythromycin (ERY), 10–20% for clarithromycin, 30% for roxithromycin, 10–20% for spiramycin, and 6–12% for azithromycin (Forth et al. 1992). Unlike other macrolide members, ERY can be inactivated easily by gastric acid to form dehydrated erythromycin (ERY-H2O) by losing one molecule of water (McArdell et al. 2003). Although ERY-H2O has negligible antibacterial activity, it may be involved in inducing bacterial resistance to the parent drug (Majer 1981). The excreted urine, carrying the active antibiotics and their metabolites, will be finally discharged to the wastewater treatment systems.
In contrast to the macrolide antibiotics working principle, mechanisms of bacterial resistance can be the excretion of antibiotic drugs by efflux pumps or the alteration of the target site to avoid binding of these drugs (Amin et al. 2006). To gain resistance, bacteria can also destruct or modify the macrolides directly by the enzymatic hydrolysis (macrolide esterase) and group transfer strategies (phosphate transfer and glycosyltransfer) (Wright 2005). Therefore, the massive use of the macrolide antibiotics has raised questions on the possible long-term consequences of resistance selection because trace levels of macrolides have been detected in aquatic environments (Hirsch et al. 1999; Kolpin et al. 2002; Giger et al. 2003; McArdell et al. 2003; Richardson et al. 2005; Karthikeyan and Meyer 2006; Xu et al. 2007). As one of the macrolide antibiotics, ERY was measured as ERY-H2O in most studies because ERY could be transformed to ERY-H2O in the slightly acidic aquatic environment or during the solid phase extraction at acidic conditions. Dehydrated erythromycin has been detected in most of water or wastewater samples surveyed worldwide, such as America (Kolpin et al. 2002; Karthikeyan and Meyer 2006), Canada (Miao et al. 2004), Germany (Hirsch et al. 1999), Switzerland (Giger et al. 2003; McArdell et al. 2003), Taiwan (Lin and Tsai 2009), Hong Kong (Gulkowska et al. 2008), and China (Richardson et al. 2005; Xu et al. 2007).
It was speculated that the ERY-resistant pathogenic bacteria could be developed even at low ERY concentrations (less than 2 µg/L) in the wastewater treatment plants (WWTPs), which breed a much wider range of environmental bacteria than the natural aquatic environment (Hirsch et al. 1999). Although the potential effects of ERY, at ultra-low concentrations (µg/L), on WWTPs have attracted the attention from the public and scientific communities, few investigations have been made so far. Most studies have been conducted on the detection of ERY-H2O at low concentrations in the WWTPs (Hirsch et al. 1999; Giger et al. 2003; McArdell et al. 2003; Karthikeyan and Meyer 2006). Usually, the resistance or susceptibility of cultures to ERY is judged by inhibitory concentrations. For example, Oleinick (1975) reported that the inhibitory concentrations of ERY to selected pure cultures were 0.01–10 mg/L and 0.1–>200 mg/L to the gram-positive and gram-negative organisms, respectively. Nimenya et al. (1999) concluded the highly significant linear correlation between dose of ERY (10, 20, 40, and 80 mg/L) and its inhibition on ammonia removal (correlation coefficient r = 0.890, probability value p < 0.05) and nitrate production (r = 0.937, p < 0.05), in which the reported inhibition percentage of ammonium oxidation and nitrate production was 0.20% and 0.28% at ERY of 1 mg/L. Additionally, the influence of ERY at relatively high concentrations in the pharmaceutical wastewater was studied in anaerobic reactors. For example, Amin et al. (2006) suggested that the development of antibiotic resistance alleviated the inhibition of ERY (1–200 mg/L) on biogas production and on butyric acid conversion in an anaerobic lab scale reactor with complex microbial communities, accompanying with concentrations of ERY in the effluent ranging from 0.4–150 mg/L. In order to reach the discharge standard of chemical oxygen demand (COD), effluent of anaerobic processes is always treated by aerobic processes, in which ERY might be further removed.
The aim of this study was to assess the influence of ERY or ERY-H2O at low concentrations (µg/L) on the carbon, nitrogen, and phosphorous removal in sequencing batch reactors (SBRs). The inhibitory effects on carbon and nutrients removal were evaluated by a long-term operation of three SBRs, namely R1, R2, and R3 fed with the same synthetic wastewater but spiked with 50 µg/L of ERY-H2O, 100 µg/L of ERY, and no antibiotics, respectively. The microbial communities of the three steady state SBRs were analyzed by employing high-density phylogenic 16S rRNA gene microarrays (PhyloChip) containing 1,440 distinguishable prokaryotic operational taxanomic units (OTUs), and the community shifts in R1 (ERY-H2O) and R2 (ERY) were compared with that in R3 (control). To verify whether the PhyloChip-observed nitrifying bacteria shifts were correlated with their resistance to ERY, short-term running batch experiments were conducted to study higher concentrations (100, 400, and 800 µg/L) of ERY’s inhibition on nitrifying bacteria present in the biomass of the three steady state SBRs.
Materials and methods
Startup and operation of SBRs
Three SBRs (4 L) were started up with the same seeding sludge from a mother reactor. The mother reactor was seeded with sludge collected from an aeration tank of a local domestic WWTP. Before being inoculated to the three SBRs, the sludge in the mother reactor was fed with synthetic wastewater for 8 months to minimize the residue inhibitory and resistant effects of antibiotics possibly existing in the local domestic wastewater, as well as, to achieve optimal nitrogen and phosphorus removal by adjusting operation parameters, such as solids retention time (SRT), hydraulic retention time (HRT), aeration, and batch modes.
The synthetic wastewater consisted of total organic carbon (TOC) of 225 mg/L (equal to theoretical COD of 600 mg/L) from glucose of 563 mg/L, NH +4 –N of 60 mg/L from NH4Cl of 229 mg/L, PO 3-4 –P of 15 mg/L from K2HPO4 of 84 mg/L, alkalinity of 420 mg/L from NaHCO3 of 706 mg/L, and the trace elements in mg/L (EDTA 3; Na2MoO .4 2H2O 1.23; CaSO .4 2H2O 24.6; MgSO4 6; CoCl2 6H2O 4; FeCl3 H2O 15; CuSO4 5H2O 0.002; MnSO4 H2O 0.16; ZnSO4 7H2O 0.002; KI 0.002; and H3BO3 0.002). The mother reactor was fed with synthetic wastewater in half-concentration all the time, while the three SBRs were fed with the synthetic wastewater in half-concentration only during the first 34 days to model the wastewater at relative low concentrations, and then increased to full concentration from day 35 to represent the wastewater at relative high concentrations. During the entire operation, the SBRs were spiked with ERY-H2O of 50 µg/L (R1), ERY of 100 µg/L (R2), and no antibiotics (R3), respectively. From day 56 to 62, the wasted sludge collected from R2 (ERY) was used to set up a fourth reactor R2′. The R2′ (fed with ERY of 100 µg/L) was operated the same as R2 except that it was applied with a shock loading of high TOC up to 6,000 mg/L on day 66, which commonly encountered in pharmaceutical wastewater treatment plants (Amin et al. 2006; Yang et al. 2009), following by various TOC loading until day 130. The TOC and nitrogen contributed by ERY-H2O or ERY were negligible (about 0.6 g TOC and 0.02 g N/g ERY or ERY-H2O). Erythromycin (potency ≥ 850 µg/mg) and all other chemicals were supplied by Sigma–Aldrich (Singapore). Erythromycin stock solution was prepared in 2 mM NaHCO3 (pH = 7.3) because its degradation rate increases logarithmically with a decrease of pH from 7.3 to 3.5 (Kim et al. 2004). Dehydrated erythromycin was prepared according to the previously reported method (McArdell et al. 2003; Abuin et al. 2006).
With an accurate programmable logic controller, the three SBRs were operated simultaneously in the same batch mode of an 8-hour cycle, consisting of inlet (0–14 min), pre-mixing for anoxic/anaerobic condition (0–60 min), aeration for oxic condition (60–300 min), post-mixing for anoxic condition (300–364 min), waste sludge (360–364 min) by decanting ~1/20 volume of 4 L mixed liquid (SRT 6.7 days, which is derived by 20/3 and 3 means three cycles per day), settling (364–404 min), decanting supernatant to 2 L (404–440 min; HRT 16 hr), and idle (440–480 min). The dilution factor was 2 for the three SBRs by discharging 2 L of liquid from the 4 L reactor in each cycle. The pH ranged from 6.6 to 8.2 during one cycle without adjustment. The stable oxygen supply during aeration guaranteed ~5 mg/L of dissolved oxygen (DO) at the end of aeration phase except for R2′ (ERY) during the shock load period. All SBRs were operated under ambient temperature (25–27 °C).
Batch experiments
To test the long-term resistance selection of nitrifying bacteria exposed to ERY-H2O (50 µg/L) in R1 and ERY (100 µg/L) in R2, short-term batch experiments of ammonium oxidation and nitrite oxidation were performed in 250 ml flask bottles with a shaking speed of 150 rpm. The ammonium oxidation batches were conducted in thirty-six flasks, which were filled with 50 ml of diluted ATCC medium 2,265 (Nitrosomonas europaea medium) to a final concentration of 35 mg/L NH +4 –N, except replacing Na2CO3 with NaHCO3 of 490 mg/L. The thirty-six flasks were divided into three groups for inocula from R1 (ERY-H2O), R2 (ERY), and R3 (control), respectively. Then each group of twelve flasks was spiked with ERY at concentrations of 0 (control), 100, 400, and 800 µg/L in triplicates. The nitrite oxidation batches were conducted in the same way as the ammonium oxidation batches in another thirty-six flasks, but filled with 50 ml of diluted ATCC medium 480 (Nitrobacter medium 203) to a final concentration of 21 mg/L NO -2 –N. Similarly to the above experiments on ERY, ammonium oxidation and nitrite oxidation were conducted on inocula from R1, R2, and R3 in a total of seventy-two flasks spiked with ERY-H2O at concentrations of 0 (control), 100, 400, and 800 µg/L. The inocula, 1 ml of biomass taken from the steady state SBRs, were washed three times with the aforementioned media by centrifugation to minimize previously fed antibiotics and organic carbon, and then were added to the liquid medium (2% by volume of inocula). All inocula were freshly applied in only one batch of experiments to avoid artificially selecting for primarily resistant organisms. The flask bottles were placed on a shaker for mixing and aeration at room temperature (25–27 °C).
Statistical analysis of the biomass–source dependent ERY inhibition was carried out by the simple paired Student’s t test to compare the daily product concentrations between any two of the three sources biomass during the 3-day period of the batch experiments. Differences were considered significant if the probability values p ≤ 0.05.
Collection and preparation of samples
During the operation of the three SBRs, equal volumes (5 ml) of effluents from three cycles of each day were collected, and then mixed together as the daily effluent for analysis. Samples in SBRs within one cycle were collected according to online DO and pH signals, which were the indicators for carbon and nitrogen removal (Yang et al. 2007). The frequency of cycle analysis was once for every 3 to 5 days in the transient and the early steady states and decreased to every 7 to 30 days in the later steady states. Samples from SBRs were filtered through a sterile filter membrane (GN-6 Metricel S-Pack, 0.45 µm, 47 mm, mixed cellulose ester, Pall Cop., USA) for TOC, nitrification, and phosphate removal analysis. Therefore, the measured TOC, phosphate, and all kinds of nitrogen were in dissolved forms throughout this study. To avoid sorption on membrane, the samples for antibiotics analysis were centrifuged at 14,000 rpm for 15 min at 25 °C. Samples from batch conical flasks were collected daily and centrifuged as above.
Analytical methods
The concentrations of dissolved TOC and dissolved TN were detected on a TOC analyzer with a TN measuring unit (Shimadzu TOC-Vcsh combined with TNM-1 Unit, Japan). Measurement of the dissolved phosphorus (PO 3-4 –P) and nitrogen in forms of NO -3 –N, NO2 -–N, and NH +4 –N were performed on an ion chromatography (DIONEX DX500 chromatography system, USA). Erythromycin and ERY-H2O were analyzed on a high performance liquid chromatography (Agilent 1100 Series, Agilent Technologies, Germany) with electro-spray tandem mass spectrometry (API 2000TM, Applied Biosystems/MDS Sciex, USA; LC-MS-MS). The previously reported LC method (McArdell et al. 2003) was utilized for analysis of ERY and ERY-H2O on a reverse phase XbridgeTM Phenyl column (50 × 2.1 mm id, 3.5 μm, waters Inc., USA.) protected by a guard column (XbridgeTM Phenyl, 10 × 2.1 mm id, 3.5 μm, waters Inc., USA.). The MS-MS analysis was performed in the positive ion mode (McArdell et al. 2003). One precursor ion and one product ion were chosen for MS-MS determination, that is, m/z 734.5 and 158.2 for ERY and m/z 716.5 and 158.2 for ERY-H2O. The detection limits were 10 µg/L for ERY and 8 µg/L for ERY-H2O.
DNA extraction, polymerase chain reaction, and PhyloChip
Approximately 1 ml of mixed liquid was centrifuged (14,000 g, 5 min) and cell pellets were collected for DNA extraction. The genomic DNA was extracted and purified by using DNeasy Tissue Kit (QIAGEN GmbH, Germany), except with modification of bead beating the cells with glass beads (Diameter 0.1 mm, Biospec, USA) for better cell lysis. The high-density phylogenic 16S rRNA gene microarrays (PhyloChip) employed in this study contain 1,440 distinguishable prokaryotic OTUs identified by 35,000 probes, which were designed according to the approach described previously (DeSantis et al. 2003). Each DNA pool extracted from triplicate sludge samples was tested on triplicate PhyloChips. The 16S rRNA gene amplification, PhyloChip processing, scanning, OTU scoring, and normalization were all performed as previously described (Brodie et al. 2006; Flanagan et al. 2007).
Results
Effects of ERY-H2O on SBR performance
The sequencing batch reactor R1 (fed with 50 µg/L of ERY-H2O) and R3 (control reactor) reached their first steady state (e.g., constant carbon and nutrients removal) on day 21 when added with TOC of 110 mg/L, NH +4 –N of 30 mg/L and PO 34 –P of 7.5 mg/L. After doubling the concentrations of TOC, NH +4 –N, and PO 34 –P on day 35, R1 (ERY-H2O) and R3 (control) once again reached their steady states after day 55. Similar carbon and nitrogen removal profiles were observed in R1 (ERY-H2O) and R3 (control) during the whole running period of 400 days (Fig. 1 showed data of 180 days, and supplementary Fig. S1 showed data of 400 days), which were consistent with the intensive observation on nitrogen removal profile within individual cycle during the steady states (Fig. 2). Concentrations of TOC in effluents of R1 (ERY-H2O) and R3 (control) were below 10 mg/L in most of the days during the operation period, even with doubled concentrations of influent on day 35. The NH +4 –N concentrations were undetected in the daily effluents for most of the days, indicating that ammonium oxidation was completed and not affected by the increased dosage of both TOC and NH +4 –N in the influents on day 35. As the intermediate of nitrification and denitrification, NO -2 –N was not accumulated in the effluents and appeared to be negligible during the aerobic nitrification process. Because the amount of TN and NO -3 –N decreased in the post-mixing period (Fig. 2), endogenous denitrification occurred. Additionally, ERY-H2O concentration in the effluent of R1 (ERY-H2O) was always ~15 µg/L. The missed 70% of ERY-H2O (~35 µg/L) could be absorbed to the sludge or degraded by the biomass.

Daily effluents of R1 (ERY-H2O of 50 µg/L), R2 or R2′ (ERY of 100 µg/L), and R3 (control): the averages of soluble TN (▲), NO -3 –N (■), NO -2 –N (×), NH +4 –N (◆), and TOC (●) or (○) in daily effluents consist of equal volumes of effluents from three cycles of each day

Comparison of nitrogen dynamics within the cycles of a R1 (ERY-H2O) and b R3 (control) during the steady states, c R2′ (ERY) on Day 119, and d R2’ (ERY) on Day 130: Soluble TN (▲), NO -3 –N (■), NO -2 –N (×) and NH +4 –N (◆)
Effects of ERY on SBR performance
Similarly, when comparing the TOC and NH +4 –N removal of R2 (100 µg/L of ERY) with that of R3 (control), ERY was not found to have any effect on the performance of the reactor during the 400 days operation (Fig. 1 and supplementary Fig. S1). To further understand whether the antibiotic ERY at µg/L levels could cause the performance of reactor beyond recovery at accidental conditions (such as the commonly occurred shock loading of high TOC in pharmaceutical wastewater treatment plants), a TOC of 6,000 mg/L in the influent was supplied to R2′ (a duplicate of R2) for 1 week (from day 65 to 72), while R2 (ERY) was operated at normal TOC concentration of 220 mg/L to avoid reactor failure and to minimize possible microbial community changes due to the TOC shock. The results showed that carbon removal in R2′ (ERY) was recovered within 18 days after the shock of high TOC, indicating that ERY did not prevent R2′ from recovery, though the nutrients of nitrogen (N) and phosphorous (P) were insufficient (Fig. 1b′ and supplementary Fig. S1b). The following describes the recovery process. When the concentration of TOC in the influent was reduced to 400 mg/L (from day 73 to 90), the concentration of TOC in the effluent decreased in four levels (3,500 mg/L, 400 mg/L, and 200 mg/L and then lower than 50 mg/L on day 82), indicating that the organic carbon was biodegraded rather than by dilution because of a dilution factor of only 2 for the SBRs. After day 91, with enough aeration and overdosed nutrient, carbon removal in R2’ (ERY) was recovered totally as TOC concentrations in the effluent dropped to 15 mg/L and maintained below 10 mg/L in later days.
Moreover, during the periods of the shock loading and the recovery phases of R2′ (ERY), the results demonstrated typical profiles of nitrification, from nitrite production to complete nitrification (Fig. 1e and supplementary Fig. S1e). The change occurs commonly when SBRs start nitrification even without antibiotics, since ammonium oxidizing microorganisms (AOMs; like Nitrosomonas) grow significantly faster than nitrite oxidizing bacteria (NOB; like Nitrospira) (Watson et al. 1989). When R2′ (ERY) was fed with excess amount of ammonium than assimilation required on day 91, the NO -2 –N and NO -3 –N appeared in the effluent with the decrease of NH +4 –N from day 100, indicating that nitrification resumed. The reduction of TN observed from day 100 to 110 suggests that denitrification resumed from day 100 as well, and reached an equilibrium from day 110 in R2’ (ERY). Interestingly, the intermediate NO -2 –N appeared to be a dominant nitrogen product from day 100 to 106, which was taken over by NO -3 –N as usual after day 106. The transit of nitrification processes was clearly confirmed by the accumulation of NO -2 –N in the aeration period as observed within the cycles on day 119 while not in cycles on day 130 (Fig. 2).
Additionally, during the whole operation period, the concentrations of ERY in the effluent of R2 (ERY) were below the detection limit (10 µg/L), and ERY-H2O which could be produced from ERY was not detected in most of the days either (detection limit of 8 µg/L). Therefore, the missed ERY might be attributed by the sorption to the biomass in R2 (ERY) or degradation to compounds other than ERY-H2O.
Phosphorus removal affected by ERY and ERY-H2O
Since phosphorus removal is another key parameter to evaluate the performance of the reactors, average concentrations of PO 3-4 –P were monitored in the daily effluents of the three SBRs (Fig. 3). The averages of PO 3-4 –P during the 400 days operation were 3.7, 4.6, and 4.8 mg/L for R1 (ERY-H2O), R2 (ERY), and R3 (control), respectively. To capture detailed process dynamics for PO 3-4 –P concentrations, 8-hour cycles were investigated for all three SBRs in the steady states. In all the cycles investigated, the PO 3-4 –P release in R1 (ERY-H2O) during the 60-minute of premixing period was relatively more than that in R2 (ERY) and R3 (control) (Fig. 3). The subtle lower PO 3-4 –P concentrations in effluent of R1 (ERY-H2O) than those in R2 (ERY) and R3 (control) might be due to the higher PO 3-4 –P release in R1 (ERY-H2O) than those in other two reactors.

a The averages of soluble PO 3-4 –P in the daily effluents of R1 (ERY-H2O), R2 (ERY), and R3 (control). b Comparison of phosphorus dynamics within the cycles of R1, R2, and R3 on day 160 during the steady states: soluble PO 3-4 –P of R1 (▲), R2 (□), and R3 (●)
PhyloChip-analyzed changes of microorganisms related to phosphorus and nitrogen removal
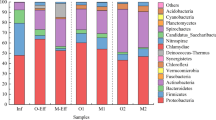
To further understand the slightly improved PO 3-4 –P removal in R1, genomic DNA extracted from steady state R1, R2, and R3 on day 367 was analyzed on the PhyloChips with probes that target the OTUs of Rhodocyclus-related poly-P accumulating organisms (PAOs) under the family of Rhodocyclacea and target the OTUs of glycogen accumulating organisms (GAOs, competing with PAOs for carbon source) under the order of GAO cluster, respectively. With PAOs and GAOs in R3 as the control, results showed that the PAOs fluorescence intensity decreased and the GAOs intensity increased slightly in R1 (ERY-H2O); both intensities of PAOs and GAOs in R2 (ERY) increased (Fig. 4). Also, the diversity of PAOs was lost in both R1 and R2 when comparing with that of R3 (Table 1). Therefore, ERY or ERY-H2O changed phosphorus removal populations, which had potential impact on the phosphorus concentrations in the effluent. Moreover, due to limited OTUs related to PAOs and GAOs on the PhyloChip, the increase of GAOs intensity in R1 and R2 could not indicate the enhancement of GAOs by ERY or ERY-H2O; similarly, the decrease of PAOs diversity and intensity in R1 might not represent the actual resistant PAOs selected by ERY-H2O. The data are not enough to explain the phenomena of the slightly improved phosphorus removal in R1 (than R2 and R3).

PhyloChip analysis of microorganism populations related to nitrification and biological P removal in three steady state SBRs. Bars above the zero line represent bacteria that increased in abundance relative to R3; bars below represent those bacteria that declined in abundance
In addition to phosphorus removal bacteria, nitrifying bacteria is another key group of organisms in wastewater treatment processes. The probes on the PhyloChip cover most of the nitrifying bacteria existing in WWTPs, including the AOMs under the family of Nitrosomonadaceae (including Nitrosomonas, Nitrosospira) and Chromatiaceae (Nitrosococcus oceani), the NOBs under the family of Bradyrhizobiaceae (Nitrobacter) and Nitrospiraceae (Nitrospira), and the anammox bacteria (strictly anaerobic bacteria occupying minor population in SBRs with aerobic conditions) under the family of Anammoxales, Pirellulae, and Planctomycetaceae. All these nitrifying bacteria studied are known as gram-negative. Comparing with those in R3 (control), the fluorescence intensity of AOMs and NOBs decreased significantly in R1 (ERY-H2O) and R2 (ERY), but anammox bacteria increased slightly in R1 and R2 (Fig. 4). The PhyloChip data also demonstrated taxa diversity reduction for nitrifying bacteria in R1 (50 µg/L of ERY-H2O) and R2 (100 µg/L of ERY) comparing with R3 (control; Table 1). The observed taxa diversity loss was the most for Nitrosomonadaceae (the dominant AOMs in WWTPs) in R1 of 80% and in R2 of 70%. The existing OTUs of Nitrosomonadaceae in R1 and R2 had the higher specific intensity than those in R3, suggesting that the less diverse AOMs selected by ERY-H2O or ERY could play the same role as the more diverse community in R3. Indeed, ammonium oxidation was similar in the three SBRs as shown in Figs. 1 and 2. Similarly, nitrifying bacteria (AOMs and NOBs) with the same nitrification performance (Figs. 1 and 2) but the lower diversity and total intensity in R1 and R2 than those in R3 (Table 1 and Fig. 4) may have a higher specific nitrification capacity which suggests a higher antibiotic resistance. For instance, the total intensity of AOMs on the PhyloChip showed a trend of R1 (ERY-H2O) <R2 (ERY) <R3 (control), indicating that the specific ammonium oxidizing capacity and antibiotic resistance of AOMs were in a reverse order of R1 (ERY-H2O) >R2 (ERY) >R3 (control). Similarly, the total intensity of NOBs demonstrated a trend of R2 (ERY) <R1 (ERY-H2O) <R3 (control), indicating that the specific nitrite oxidizing capacity and antibiotic resistance of NOBs could be R2 (ERY) >R1 (ERY-H2O) >R3 (control).
Resistance selection of nitrifying bacteria upon exposure to ERY or ERY-H2O
To verify the resistance selection of nitrifying bacteria indicated by PhyloChip-observed shifts in AOMs and NOBs, biomass from the three steady state SBRs on day 390 were inoculated to the conical flasks with organic carbon free media spiked with different dosages of ERY or ERY-H2O. The impact of biomass growth on the batch experiments can be ignored, since the optical densities of mixing liquid in the flasks showed no difference with time in the 3-day batch experiments. The negligible growth of the biomass can be explained by that fast growing heterotrophic bacteria cannot be supported by these organic carbon free media and the autotrophic nitrifying bacteria (usually occupy 2–3% of total activated sludge biomass) possess relatively long doubling time (such as >8 h at optimal conditions) (Prosser 1989; Koch et al. 2001). In the flasks spiked with ERY-H2O (at concentrations of 100, 400, and 800 µg/L), the biomass from each SBR completely oxidized ammonium or nitrite at similar rates as that of control (without ERY-H2O spiked) within 3 days (data not shown), indicating that ERY-H2O did not inhibit ammonium oxidation or nitrite oxidation in the short-term experiments, although ERY-H2O (50 µg/L in R1) caused microbial community shift in the long-term operation.
Different from ERY-H2O, ERY (at concentrations of 100, 400, and 800 µg/L) inhibited the whole nitrification process, and the inhibition extent depended on the biomass–source and the ERY concentration. For ammonium oxidation, significant difference of ERY inhibition was observed between biomass from R1 (ERY-H2O) and R2 (ERY; p < 0.01), R1 (ERY-H2O) and R3 (control; p < 0.05) based on the statistical analysis of the 3-day product concentrations in the batch experiments. For nitrite oxidation, difference of ERY inhibition was obvious on biomass from R1 (ERY-H2O) and R3 (control; p < 0.01), but less obvious on biomass from R2 (ERY) and R3 (control; p = 0.07). In addition, the difference of ERY inhibition on nitrification processes was studied in detail on the second incubation day by using inhibition percentage (for the same biomass, \( 100\% \times \left( {1 - \frac{{{\text{N}}{{\text{O}}_x} - {\text{N produced with ERY}}}}{{{\text{N}}{{\text{O}}_x} - {\text{N produced without ERY}}}}} \right) \)). The maximum inhibition of ammonium oxidation by ERY (100, 400, and 800 µg/L) occurred on the biomass from R3 (control; 82–93%), followed by R2 (ERY; 71–95%), and R1 (ERY-H2O; 56–91%) after 48 h reaction (Fig. 5), indicating that the resistance of AOMs to ERY was in an order of R1 (ERY-H2O) >R2 (ERY) > R3 (control). Similarly, the maximum inhibition of nitrite oxidation by ERY (100, 400, and 800 µg/L) also occurred on the biomass from R3 (control; 33–61%) after 48 h reaction, but ERY inhibited the nitrite oxidation similarly on the biomass from R1 (ERY-H2O; 21–42%) and R2 (ERY; 18–23%; Fig. 5), indicating that the resistance of NOBs to ERY was in an order of R2 (ERY) ≥R1 (ERY-H2O) >R3 (control). The results also demonstrate that ERY inhibited nitrite oxidation (18–61%) less significantly than ammonium oxidation (56–95%), which is consistent with the less NOBs diversity loss than AOMs as indicated by the PhyloChip readings (Table 1).

In the batch experiments, a ammonium oxidation affected by ERY—(NO -2 –N+NO -3 –N) produced in the batches of R1 (ERY-H2O; ▲), R2 (ERY; ◆), and R3 (control; ■) after 48 h incubation; and b nitrite oxidation affected by ERY—NO -3 –N produced in the batches of R1 (ERY-H2O; ▲), R2 (ERY; ◆), and R3 (control; ■) after 48 h incubation. The values represent means ± standard deviations (n = 3)
Discussion
In this study, a relatively low concentration of ERY (100 µg/L) or dehydrated ERY (ERY-H2O; 50 µg/L) was investigated on their influences to the performance of the SBRs. Both ERY and ERY-H2O showed negligible influence on carbon and nutrients (N and P) removal in the SBRs, which is consistent with the findings of the previous studies on even much higher concentrations of ERY (in mg/L range) (Oleinick 1975; Amin et al. 2006). In contrast, analyzed on high-density microarrays (PhyloChip), bacteria related to N and P removal changed in both diversity and abundance (reflected by fluorescence intensity) due to exposure to ERY-H2O (50 µg/L) or ERY (100 µg/L). The above observations could be explained by the selection of resistant bacteria, which are highly possible to be present in the inoculated activated sludge consisting of high diversity and density of bacteria (Kummerer 2009). This could lead to a shorter adaptation time of bacteria against antibiotics via resistance, such as the ability to biodegrade antibiotics or the efflux of antibiotics to eliminate toxic effects.
The resistant nitrifying bacteria selected upon exposure to low concentrations of the antibiotic ERY or its metabolite ERY-H2O was further confirmed in the short-term batch experiments with higher concentrations of ERY at 100, 400, or 800 µg/L. This is one step further to answer the current open question about the resistance selection due to low concentrations of antibiotics in water by experimental data rather than by speculation (Hirsch et al. 1999). Previous studies reported that ERY-H2O can induce gram-positive Staphylococcus aureus RN 1389 to resist ERY (Majer 1981). The present study demonstrates that ERY-H2O can also induce nitrifying bacteria, which are all known as gram-negative bacteria, to resist ERY. The biomass–source dependent inhibition of ammonium oxidation by ERY (R1 (ERY-H2O) <R2 (ERY) <R3 (control)) suggests that AOMs acclimated to ERY-H2O (50 µg/L in R1) can create stronger resistance to ERY than AOMs acclimated to ERY (100 µg/L in R2). Also, the biomass–source dependent inhibition of nitrite oxidation by ERY (R2 (ERY) ≤R1 (ERY-H2O) <R3 (control)) suggests that NOBs acclimated to ERY-H2O (50 µg/L in R1) can also create resistance to ERY. This shed light on the problem that formation of antibiotic metabolites does not mean elimination of environmental impact of antibiotics. In fact, some metabolites of antibiotics are more toxic to human than their parent drugs, such as acetyl derivative of sulfamethoxazole (Kummerer 2009). Additionally, current batch studies showed higher inhibition of ammonia removal (56–95%) and nitrate production (18–61%) by ERY (100, 400, and 800 µg/L; Fig. 5) than previous inhibition study on ammonium oxidation (0.20%) and nitrate production (0.28%) at ERY of 1,000 µg/L (Nimenya et al. 1999). The inhibition differences could be explained by different experimental conditions, such as Nimenya et al. (1999) used shorter reaction duration, higher concentrations of nitrifying bacteria growing on the biofilms, and ERY solution prepared in deionised water at pH 5.8.
Different from nitrifying bacteria, the PhyloChip-observed information on PAOs and GAOs seems insufficient to explain the phenomena of the slightly improved phosphorus removal in R1 (than R2 and R3), which is mainly due to that PAOs and GAOs cover more complex but still largely uncertain phylotypes than Rhodocyclus-related PAOs and GAO cluster (Seviour et al. 2003). Therefore, more information is needed in the future study.
In addition to the influence of ERY and ERY-H2O on SBRs, the fate of these antibiotics in the reactors is another concern. As researchers suggested, the removal of ERY or ERY-H2O in WWTPs is highly possible due to the sorption by high concentrations of suspended sludge (Giger et al. 2003; McArdell et al. 2003). However, the removal efficiency of ERY-H2O (70%) in R1 of this study was higher than those observed in the WWTPs at about 9–19% (Gulkowska et al. 2008) or ~50% (Yang et al. 2006). Besides possible biodegradation, the difference in sorption may be due to the different characteristics of the activated sludge. The SBRs in this study were fed with synthetic wastewater containing only soluble and easily degradable glucose as organic carbon source, while the real WWTPs contain complex organic carbon source. The different carbon sources could lead to the activated sludge with different extracellular polymeric substances, which would affect the sorption ability (Li and Yang 2007). In addition, the degradation of ERY is another concern during studying its effects on the nutrients removal. Since neither ERY nor ERY-H2O was detected in the effluent of R2 (ERY) and biosorption has never been reported to have such high removal efficiency on ERY or ERY-H2O at even lower concentrations, ERY was suspected to be transformed to products other than ERY-H2O. Our discovery is different from previous studies in which the unchanged ERY was degraded easily to ERY-H2O in the aquatic environment (Hirsch et al. 1999; Kolpin et al. 2002; Giger et al. 2003; McArdell et al. 2003; Richardson et al. 2005; Karthikeyan and Meyer 2006; Xu et al. 2007). The difference may be due to various pH values in the studies or may be due to biodegradation of ERY. Further investigation is needed to identify the degradation mechanism of ERY and the degradation product(s).
In conclusion, ERY-H2O (50 µg/L) and ERY (100 µg/L) exhibit no unfavorable results on the efficiency of SBRs in treating the synthetic wastewater. However, both compounds cause changes of microbial community composition in the reactors, which leads to the selected microorganisms more adaptive and resistant to the inhibitory effects of ERY and ERY-H2O. This study suggests that the influence of ERY and ERY-H2O at the µg/L levels on treatment systems such as the complex wastewater treatment plants, one of the most highly antibiotics-exposed environments, is more likely to induce a pool of ERY resistance genes than to inhibit the treatment.
References
Abuin S, Codony R, Compano R, Granados M, Prat MD (2006) Analysis of macrolide antibiotics in river water by solid-phase extraction and liquid chromatography-mass spectrometry. J. Chromatogr. 1114:73–81
Amin MM, Zilles JL, Greiner J, Charbonneau S, Raskin L, Morgenroth E (2006) Influence of the antibiotic erythromycin on anaerobic treatment of a pharmaceutical wastewater. Environ. Sci. Technol. 40:3971–3977
Brodie EL, DeSantis TZ, Joyner DC, Baek SM, Larsen JT, Andersen GL, Hazen TC, Richardson PM, Herman DJ, Tokunaga TK, Wan JMM, Firestone MK (2006) Application of a high-density oligonucleotide microarray approach to study bacterial population dynamics during uranium reduction and reoxidation. Appl. Environ. Microbiol. 72:6288–6298
DeSantis TZ, Dubosarskiy I, Murray SR, Andersen GL (2003) Comprehensive aligned sequence construction for automated design of effective probes (CASCADE-P) using 16S rDNA. Bioinformatics 19:1461–1468
Dubois M, Fluchard D, Sior E, Delahaut P (2001) Identification and quantification of five macrolide antibiotics in several tissues, eggs, and milk by liquid chromatography-electrospray tandem mass spectrometry. J. Chromatogr. B 753:189–202
Flanagan JL, Brodie EL, Weng L, Lynch SV, Garcia O, Brown R, Hugenholtz P, DeSantis TZ, Andersen GL, Wiener-Kronish JP, Bristow J (2007) Loss of bacterial diversity during antibiotic treatment of intubated patients colonized with Pseudomonas aeruginosa. J. Clin. Microbiol. 45:1954–1962
Forth W, Henschler D, Rummel W, Starke K (1992) Allgemeine und spezielle pharmakologie und toxikologie. BIWissenschaftsverlag, Mannheim, Leipzig, Wien, Zürich
Giger W, Alder AC, Golet EM, Kohler HPE, McArdell CS, Molnar E, Siegrist H, Suter MJF (2003) Occurrence and fate of antibiotics as trace contaminants in wastewaters, sewage sludges, and surface waters. Chimia 57:485–491
Gulkowska A, Leung HW, So MK, Taniyasu S, Yamashita N, Yeunq LWY, Richardson BJ, Lei AP, Giesy JP, Lam PKS (2008) Removal of antibiotics from wastewater by sewage treatment facilities in Hong Kong and Shenzhen, China. Water Res. 42:395–403
Hirsch R, Ternes T, Haberer K, Kratz KL (1999) Occurrence of antibiotics in the aquatic environment. Sci. Total Environ. 225:109–118
Karthikeyan KG, Meyer MT (2006) Occurrence of antibiotics in wastewater treatment facilities in Wisconsin, USA. Sci. Total Environ. 361:196–207
Kim YH, Heinze TM, Beger R, Pothuluri JV, Cerniglia CE (2004) A kinetic study on the degradation of erythromycin A in aqueous solution. Int. J. Pharm. 271:63–76
Koch G, Kuhni M, Siegrist H (2001) Calibration and validation of an ASM3-based steady-state model for activated sludge systems—Part I: prediction of nitrogen removal and sludge production. Water Res 35:2235–2245
Kolpin DW, Furlong ET, Meyer MT, Thurman EM, Zaugg SD, Barber LB, Buxton HT (2002) Pharmaceuticals, hormones, and other organic wastewater contaminants in US streams, 1999–2000: a national reconnaissance. Environ. Sci. Technol. 36:1202–1211
Kummerer K (2009) Antibiotics in the aquatic environment—A review—Part I. Chemosphere 75:417–434
Li XY, Yang SF (2007) Influence of loosely bound extracellular polymeric substances (EPS) on the flocculation, sedimentation and dewaterability of activated sludge. Water Res. 41:1022–1030
Lin AYC, Tsai YT (2009) Occurrence of pharmaceuticals in Taiwan's surface waters: impact of waste streams from hospitals and pharmaceutical production facilities. Sci. Total Environ. 407:3793–3802
Majer J (1981) In vitro introduction of resistance to erythromycin by its metabolite. Antimicrob. Agents Chemother. 19:628–633
McArdell CS, Molnar E, Suter MJF, Giger W (2003) Occurrence and fate of macrolide antibiotics in wastewater treatment plants and in the Glatt Valley Watershed, Switzerland. Environ. Sci. Technol. 37:5479–5486
Miao XS, Bishay F, Chen M, Metcalfe CD (2004) Occurrence of antimicrobials in the final effluents of wastewater treatment plants in Canada. Environ. Sci. Technol. 38:3533–3541
Nimenya H, Delaunois A, La Duong D, Bloden S, Defour J, Nicks B, Ansay M (1999) Short-term toxicity of various pharmacological agents on the in vitro nitrification process in a simple closed aquatic system. ATLA Altern. Lab. Anim. 27:121–135
Oleinick NL (1975) In: Corcoran JW, Hahn FE (eds) Mechanism of action of antimicrobial and antitumor agents. Springer-Verlag, Berlin
Pothuluri JV, Nawaz M, Cerniglia CE (1998) In: Sikdar SK, Irvine RL (eds) Bioremediation: principles and practice. Technomic Publishing Co, Lancaster, Basel, pp 221–248
Prosser JI (1989) Autotrophic nitrification in bacteria. Adv. Microb. Physiol. 30:125–181
Richardson BJ, Larn PKS, Martin M (2005) Emerging chemicals of concern: pharmaceuticals and personal care products (PPCPs) in Asia, with particular reference to Southern China. Mar. Pollut. Bull. 50:913–920
Seviour RJ, Mino T, Onuki M (2003) The microbiology of biological phosphorus removal in activated sludge systems. FEMS Microbiol. Rev. 27:99–127
Verdier L, Gharbi-Benarous J, Bertho G, Mauvais P, Girault JP (2001) A novel mechanism of antibiotic resistance: study of the complex state of peptides with bacterial Staphylococcus aureus ribosomes. Comptes Rendus De L Academie Des Sciences Serie Ii Fascicule C-Chimie 4:745–750
Watson SW, Bock E, Harms H, Koops H-P, Hooper AB (eds) (1989) Nitrifying bacteria. Williams and Wilkins, Baltimore, MD
Wright GD (2005) Bacterial resistance to antibiotics: enzymatic degradation and modification. Adv. Drug Del. Rev. 57:1451–1470
Xu WH, Zhang G, Zou SC, Li XD, Liu YC (2007) Determination of selected antibiotics in the Victoria Harbour and the Pearl River, South China using high-performance liquid chromatography-electrospray ionization tandem mass spectrometry. Environ. Pollut. 145:672–679
Yang Q, Peng YZ, Liu XH, Zeng W, Mino T, Satoh H (2007) Nitrogen removal via nitrite from municipal wastewater at low temperatures using real-time control to optimize nitrifying communities. Environ. Sci. Technol. 41:8159–8164
Yang SW, Cha JM, Carlson K (2006) Trace analysis and occurrence of anhydroerythromycin and tylosin in influent and effluent wastewater by liquid chromatography combined with electrospray tandem mass spectrometry. Anal. Bioanal. Chem. 385:623–636
Yang Y, Wang P, Shi SJ, Liu Y (2009) Microwave enhanced Fenton-like process for the treatment of high concentration pharmaceutical wastewater. J. Hazard. Mater. 168:238–245
Acknowledgements
This research was supported by Academic Research Fund from Singapore Ministry of Education under project number R-288-000-024-112/133 and R-288-000-041-112.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Fig. S1
Daily effluents of R1 (ERY-H2O of 50 µg/l), R2 or R2′ (ERY of 100 µg/l), and R3 (control): the averages of soluble TN (▲), NO3 -–N (■), NO2-–N (×), NH4 +–N (◆), and TOC in daily effluents consist of equal volumes of effluents from three cycles of each day (PPT 148 kb)
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Fan, C., Lee, P.K.H., Ng, W.J. et al. Influence of trace erythromycin and erythromycin-H2O on carbon and nutrients removal and on resistance selection in sequencing batch reactors (SBRs). Appl Microbiol Biotechnol 85, 185–195 (2009). https://doi.org/10.1007/s00253-009-2201-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-009-2201-7