
Abstract
Sea turtles spend most of their life cycle in foraging grounds. Research in developmental habitats is crucial to understanding individual dynamics and to support conservation strategies. One approach to gather information in foraging grounds is the use of cost-effective and non-invasive techniques that allow public participation. The present study aimed to use photographic-identification (photo-ID) to investigate the spatio-temporal distribution of Chelonia mydas and Eretmochelys imbricata. Furthermore, we describe fibropapillomatosis occurrence. This work was carried out at subtropical rocky reefs of the Brazilian coast in Arraial do Cabo (22°57ʹS, 42°01ʹW), within a sustainable conservation unit. A total of 641 images were obtained through social media screening (n = 447), citizen science (n = 168), or intentional capture (n = 26) dated between 2006 and 2021. Additionally, 19 diving forms (between 2019 and 2021) were received from citizen scientists. All diving forms presented at least one turtle. Photo-ID identified 174 individuals of C. mydas, with 45 being resighted, while E. imbricata had 32 individuals, with 7 individuals resighted. The median interval between the first and last individual sighting was 1.7 years for C. mydas and 2.4 years for E. imbricata. Fibropapillomatosis was only observed in C. mydas, with a prevalence of 13.99% (20 of 143 individuals) and regression in 2 individuals (10.00%). Our results indicated that Arraial do Cabo is an important development area with individuals residing for at least 6 years. This study demonstrated that social media, along with photo-ID, can be useful to provide sea turtle estimates in a foraging ground using a non-invasive, low-cost method.
Similar content being viewed by others


Introduction
Research on animal developmental habitats is crucial to providing a deeper understanding of species dynamics and their interactions, which can promote effective conservation and management strategies. The most commonly used monitoring methods include direct observation, tissue analysis, gut content analysis, genetics, and capture-mark-recapture (CMR), all of which provide valuable data on movement, distribution, diet, health assessment, morphometric values, survival, and behavior (Bourlat et al. 2013; Crossin et al. 2014; Katsanevakis et al. 2012; Nielsen et al. 2017). CMR methods are widely used to monitor individuals and populations over time, and to gather information on growth, demographic parameters and population trends (Casale et al 2009; Crossin et al. 2014; Castelblanco-Martínez et al. 2019; Kanive et al. 2019; Tourani 2021). Although these techniques are known for their accuracy in data collection, the difficulties associated with the most applied CMR techniques (telemetry and tags) are the need for capture, handling, complex logistics for implementation tag loss, transmitter failure, and high-cost equipment (Hebblewhite and Haydon 2010; Casale et al. 2017).
The sea turtle species Chelonia mydas and Eretmochelys imbricata have a complex life cycle that includes an early oceanic juvenile stage, followed by recruitment to neritic developmental habitats, where juveniles and adults can be found (Bolten 2003). Monitoring efforts in foraging grounds are particularly important as individuals spend most of their lives in these habitats; juveniles growing to constitute future spawning stocks, and adults occupying these areas during the remigration interval (Wildermann et al. 2018). Over the past few decades, several studies in foraging grounds have highlighted the influence of habitat quality, prey availability, presence of predators, temperature and substrate type on individual development, disease susceptibility, survival, habitat use and fidelity (Metz et al. 2020; Gillis 2020).
Photographic-identification (photo-ID) is a cost-effective and non-invasive CMR technique that relies on comparing natural marks on individuals using photographs (Schofield et al. 2008). For sea turtles, photo-ID compares unique patterns of facial scales on the sides of the head (Carter et al. 2014; Dunbar et al. 2014; Araujo et al. 2016; Calmanovici et al. 2018), dorsal scales of the head (Dunbar et al. 2014), flippers scales (Gatto et al. 2018) or carapace images (Tabuki et al. 2021). These marks remain stable over the years, making them a great long-term identification tool (Carpentier et al. 2016). However, marks may differ between the two sides of the same individual (Su et al. 2015), implying the need to use photographs of both sides or to standardize the use of only one side to determine the number of individuals. As well as the traditional CMR methods, photo-ID has been used to established patterns of movement, behavior, and habitat use of individuals (Baumbach et al. 2019; Papafitsoros et al. 2020; Petit et al. 2020; Schofield et al. 2020; Hanna et al. 2021).
Several algorithms and software have been developed for photo-ID analysis, allowing conservation programs to incorporate underwater images and reducing the need for turtle manipulation (Dunbar et al. 2014, 2021; Williams et al. 2017; Baumbach et al. 2019). Some of these facilitate the participation of non-scientific volunteers (citizen scientists) who can contribute through structured protocols, opportunistic data, or crowdsourcing initiatives (Baumbach and Dunbar 2017; Becken et al. 2019). Collaboration provides mutual benefits, by reducing science-related costs and increasing the amount of information collected, while citizen scientists receive a sense of participation and become familiar with research and scientific knowledge (Lucrezi et al. 2018). In the marine realm, some of most popular citizen scientists are SCUBA divers, who have access to a variety of information (e.g. abiotic data, species presence, and photos), most of which can be used in ecological studies, such as species monitoring and climate change studies (Martin et al. 2016; Lucrezi et al. 2018).
The green turtle, C. mydas, and the hawksbill turtle, E. imbricata, use the Brazilian coast as both nesting and foraging grounds, and both are found on national and international lists of threatened species. However, most feeding grounds in this area are understudied (Santos et al. 2011; IUCN 2022). Both species suffer from anthropogenic threats, including interaction with debris and fishing, boat strikes, and fibropapillomatosis (Monteiro et al. 2016; Farias et al. 2019; Tagliolatto et al. 2020). Despite the potential for sea turtle research along the Brazilian coast, few published articles use photo-ID in underwater surveys (Reisser et al. 2008; Proietti et al. 2014a; Calmanovici et al. 2018). Even with the high potential recreational use of Brazilian coastal areas, few studies have taken advantage of citizen-science activities to access sea turtles (Calmanovici et al. 2018; Vieira et al. 2020).
Arraial do Cabo is a Brazilian municipality, and part of a sustainable use conservation unit, the Marine Extractive Reserve of Arraial do Cabo, where only traditional fishing is allowed. The reserve does not have a ‘no-take’ area, and general enforcement is rather ineffective. Although the region is a high-density foraging ground for C. mydas, and home to E. imbricata (Mello-Fonseca et al. 2021), sea turtles are not a target species of local conservation strategies. The clear water and rich marine biodiversity of Arraial do Cabo enables the occurrence of diverse aquatic tourist activities, such as recreational dives and boat trips. In Arraial do Cabo, 13 local dive operators perform approximately 25,000 dives throughout the year, especially during holidays, in winter and in the summer period (Pedrini et al. 2007; Giglio et al. 2017a). These dives are conducted along the rocky reefs at depths ranging from 3 to 11 m. The density of sea turtles makes these animals easy to find during diving operations, thereby facilitating the acquisition of photos and videos that are valuable for citizen-science monitoring (Giglio et al. 2018).
This study aimed to estimate the number of individuals, frequency of encounter, site fidelity, and residence of C. mydas and E. imbricata in a subtropical rocky reef foraging ground with great tourist appeal using online dive forms and photo-ID. This is the first study to combine citizen science monitoring and photo-ID to describe sea turtle spatio-temporal trends on the Brazilian coast. We also attempted to search for the temporal prevalence and incidence of fibropapillomatosis. In particular, we highlight the importance of applying photo-ID from citizen-scientists data in touristic areas.
Materials and methods
Study site
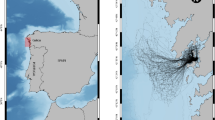
The study was conducted in Arraial do Cabo (22°57ʹS, 42°01ʹW) on the southeastern coast of Brazil, which is characterized by a subtropical rocky reef ecosystem influenced by seasonal upwelling events with average water temperature ranging from 18ºC to 22ºC (Cordeiro et al. 2020). The physical barrier formed by the Cabo Frio and Porcos Islands creates two distinct regions regarding the influence of upwelling and waves: the exposed and the sheltered region (Fig. 1). The exposed region has lower temperatures (average ~ 18 °C) and a predominance of warm-temperate affinity species, while the sheltered region has an average temperature of 22 ºC and tropical affinity species dominance (Guimaraens and Coutinho 1996; Ferreira et al. 2001). Most of the scuba dives that occur in Arraial do Cabo are limited to the sheltered region and are concentrated in 4 points (Porcos Island, Cardeiros, Anequim and Abóbora Bay) (Pedrini et al. 2007; Giglio et al. 2017a) (Fig. 1). The exposed region has a maximum depth of 25 m, while the sheltered region has an average depth of 6 m, with a maximum depth of 15 m. These reefs are predominantly rocky shores composed of a rich epilithic algal community (EAC), macroalgae, and the zoanthid Palythoa caribaeorum, with a lower presence of corals and other invertebrates. In comparison, the exposed region has a higher macroalgae coverage, however they share the same dominant group, Rhodophyta (Mello-Fonseca et al. 2021).

a Map of Rio de Janeiro state in Brazil, indicating the municipality of Arraial do Cabo. b Map of Arraial do Cabo with indications of the most visited dive sites according to Pedrini et al. (2007): AQ (Anequim), AB (Abóbora Bay), CA (Cardeiros), PO (Porcos Island) and location of intentional captures: PG (Praia Grande), PA (Praia dos Anjos), PP (Praia do Pontal)
Data input
To obtain the data, a citizen-science program named “Mar de Tartaruga” (Turtle Sea) was created in November 2019. A social media account (Instagram; www.instagram.com) was used to identify and contact dive operators, divers, and underwater photographers from Arraial do Cabo. The hashtag “Mergulho em Arraial do Cabo” (Diving in Arraial do Cabo) was used on the Instagram search tool to find other recreational and professional divers who were willing to participate. Moreover, the same term was used on YouTube (www.youtube.com) and Vimeo (www.vimeo.com) to find additional underwater photographers.
Social media profiles were pre-selected (54 Instagram pages) by the presence of at least one underwater image or video of sea turtles in Arraial do Cabo. All divers were contacted to request permission to use the images and were introduced to the citizen-science project. When individuals were interested in participating, they were invited to share any former or unposted digital archives accompanied by additional information (date, location, and general observations). Therefore, the database consisted of observations dating back to before this study began (i.e. before 2021). We maintained contact with the citizen scientists through Instagram, or Whatsapp Messenger app, if contact was requested, to share an online diving form to facilitate data acquisition. The diving form had 11 questions and space for media file uploading (Table S1).
We also used photos from a parallel monitoring program to increase our database. Sea turtles were intentionally hand-captured in snorkeling surveys (July and August 2021) at three locations (Praia Grande, Praia dos Anjos, and Praia do Pontal) (Fig. 1). Animals were measured at the maximum curved carapace length (CCL) with a flexible tape (0.1 cm precision), weighed using a digital hanging scale (0.1 kg precision) and photographed for photo-ID. All captured individuals (n = 13) were juveniles of C. mydas (CCL ranged from 32.0 to 58.4 cm, mean = 38.5 cm).
Photo-ID
All images were reviewed for suitability for the comparison process. The criteria to select images included: lateralized images, good definition, and absence of epibionts or any object interfering in the visualization of the lateral head scales. Images without dates or locations were excluded.
Photo-ID was performed in the pattern-recognition algorithm “Hotspotter” (Crall et al. 2013), available on the platform “Internet of Turtles” (iot.wildbook.org). The photos were submitted to the Internet of Turtles and went through the automatic process of identifying the side present in the photo (left or right), with validation by the researcher, and detection of the lateral head scales pattern. To determine which side is present in the photos (right or left), Internet of Turtles machine learning considered the position of the turtle in the photo. If the turtle was facing to the right, it was considered a right side photo, if the turtle was facing to the left, it was considered a left side photo. We acknowledge the possibility of misidentifications caused by mirrored images at the time of capture, storage or posting, but this was the only classification method available. Comparison was made by the Hotspotter algorithm between pairs of images, considering the side shown in the picture (Fig. 2). After, 30 images with the best compatibility probability were manually validated. In case of doubt about compatibility, flippers scales were manually used for confirmation.

Example of photo-ID from one individual, the areas that are highlighted display potentially matched features of the left side. a Sighting of 12/01/2014. b Sighting of 05/10/2021
Fibropapillomatosis (FP)
To determine the presence or absence of FP, only photos with non-retracted neck regions and at least one flipper were evaluated. Prevalence was calculated by dividing the total number of individuals with FP by the total number of individuals evaluated for each year and the total interval (2006–2021) (Tagliolatto et al. 2016). Tumor regression was recorded whenever reduction of tumors was identified in resighted individuals. To assess tumor regression, individuals’ images were visually compared, using the largest length of the tumor and having the length of the turtle’s eye as a reference to identify proportional changes in tumor size. Although observations were limited to only a fraction of sea turtle bodies (i.e., the side shown in the image), this is the first data on FP for free-living sea turtles at Arraial do Cabo.
Data analysis
Here, the term “sighting” refers to the event where photos were taken of one or both sides of the individual. To avoid overestimating the number of individuals, sightings were separated into 3 groups according to the side view in the photos: (A) individuals with right side photos, (B) individuals with left side photos, and (C) individuals with both sides photos (A intersection B). Thus, the minimum number of unique individuals was group ‘A’ or ‘B’, considering the group with the highest number of distinct turtles, and the maximum number of unique individuals was the sum of all unique individuals (group A + group B−group C). The minimum and the maximum number of unique individuals was calculated for the total interval (2006–2021) and for each year. Only the minimum number of unique individuals for the total interval were used to calculate the residence time and FP prevalence. Residence time was estimated as the interval between the individua's first and last sighting. Whenever the exact day was not available, the sighting was standardized to the 15th day of the month. When the location of the images taken at dive sites on Cabo Frio Island was inaccurate, it was considered and labelled as Cabo Frio Island. Only one person (Isabella Neves-Ferreira) made the entire selection of photos and analysis.
Results
Data input
In total from 576 sightings, 641 good quality images were obtained for photo-ID, dated between April 2006 and October 2021. Of these, 447 photos were acquired through social media screening, 168 from voluntary submissions, and 26 from snorkeling intentional hand-captures. Only 26 photos did not have the exact day and were considered as sightings on the 15th day of the month. The species Chelonia mydas (80.97% of the records) was the most representative species. Over the 15 years, there was an increase in the number of photos, with the highest number of C. mydas images in 2020 (n = 145) and E. imbricata in 2019 (n = 25) (Fig. 3a). Geographic information (i.e., name of the diving site) was available for 219 sightings, with the majority (66.21%) occurring in the sheltered region of Arraial do Cabo, where dives are more frequent (Fig. 3b).

Sighting information for Chelonia mydas and Eretmochelys imbricata in Arraial do Cabo, Brazil. a Number of photos over the years. b Sighting locations for Chelonia mydas (left) and for Eretmochelys imbricata (right), the circle size is proportional to the number of sightings. The circles in the middle of Cabo Frio Island indicate data collected at any of the island’s dive points in the sheltered region, but without defined location. CM: C. mydas, EI: E. imbricata
Between November 2019 and October 2021, 19 diving forms were received, totaling 26.4 h of diving at 9 dive sites. All dives had at least one turtle sighting. Divers reported 45 observations of C. mydas resulting in an encounter rate of 1.7 turtle·hour−1. Sightings of E. imbricata were comparatively less frequent, with a total of 19, resulting in an encounter rate of 0.72 turtle·hour−1.
Photo-ID
The minimum number of unique individuals of C. mydas was 174, of which 132 had one side photo and 42 had both side photos. E. imbricata presented 32 distinct individuals, of which, 23 individuals had only one side photo and 9 had both side photos. Thus, in the total interval (2006–2021), the ratio between the number of individuals of C. mydas and E. imbricata was 5:1. Among years, the ratio of C. mydas to E. imbricata individuals was not constant and ranged from 2:3 in 2014 to 8:1 in 2020 (Fig. 4b). The greatest number of individuals was found in the year with higher number of photos of C. mydas (2020 with 59 individuals), and in the third year with the highest number of photos of E. imbricata (2020 with 7 individuals) (Fig. 3a and 4b). The number of individuals reached a maximum of 302 individuals for C. mydas and 46 for E. imbricata, considering the maximum number of unique individuals (group A + group B−group C) (Fig. 4a).

Information of the photo-ID process. a Number of individuals identified in the total range. b Number of individuals identified per year for Chelonia mydas (top) and for Eretmochelys imbricata (bottom). In color (yellow and purple) are the minimum number of unique individuals confirmed for each species, with the values in black. In gray is the maximum possible number of individuals. To calculate the minimum values, we considered the highest value of unique individuals between group ‘A’ and group ‘B’, where group ‘A’ are individuals with right side photos and group ‘B’ are individuals with left side photos. The maximum number of unique individuals was the sum of all unique individuals (group A + group B−group C), where group ‘C’ is individuals with both side photos (A intersection B). CM: C. mydas, EI: E. imbricata
Considering the minimum number of unique individuals, 25.8% (n = 45) of C. mydas and 21.8% (n = 7) of E. imbricata were resighted. However, these individuals were present in 58.6% (n = 210) of all C. mydas images and 71.5% (n = 73) of all E. imbricata images. The median interval between each resighting event was 3.7 months (mean = 8 ± 10.2 months) for C. mydas and 1.2 months (mean = 4 ± 8.6 months) for E. imbricata. The median interval between the first and last individual sighting was 1.7 years (min.: 0.1 months–max.: 6.7 years) for C. mydas and 2.4 years (min.: 0.8 months–max.: 5.6 years) for E. imbricata (Fig. 5). The timeline of these individuals and the last registered dive site can be found in Supplementary Information—Fig. S1 and S2. Among the resighted individuals, five stood out for being seen more than once over consecutive years at the same or at nearby dive sites, and two individuals were seen at distant dive sites (Table 1).

Information of resighted individuals in Arraial do Cabo, Brazil. Interval, in years, between the first and last record of each individual for Chelonia mydas (top) and for Eretmochelys imbricata (bottom). The dotted line represents the median. Records with no defined day were assigned to the the 15th day of the month
Fibropapillomatosis (FP)
A total of 293 photos were used (229 of C. mydas and 64 of E. imbricata) for the FP’ analysis. Photos were from 119 individuals of C. mydas and 24 individuals of E. imbricata. There was no FP detection on E. imbricata. Tumors were observed in 20 individuals (36 photos) of C. mydas. The highest number of new individuals identified with the disease was in 2018 (8 individuals), and in 2019 there were no new occurrences of FP. The prevalence of the disease between 2006 and 2021 was 13.99%. Excluding 2010, which had only one photo evaluated, the highest value was found in 2018 with 24.39%, followed by 2016 with 20.00%. FP site-specific regression was observed in two individuals (10.00%), one in 2018 and the other in 2019 (Table S2). Tumor progression was observed in another individual after a 9 month interval.
Discussion
Through the analysis of hundreds of photos retrieved from social media and from photos provided by citizen scientists, we found that most turtles on the rocky reefs of Arraial do Cabo are C. mydas. We identified individuals that are residents for up to 6 years and have a high degree of site fidelity. Although the method used does not allow a full body analysis, our results show that C. mydas is affected by FP, and we were also able to report cases of site-specific tumor regression.
Citizen science
Information from citizen scientists helped extend our understanding of the spatial and temporal distribution of sea turtles in the subtropical rocky reefs studied. However, opportunistic data only allowed us to obtain baseline information on spatial and temporal patterns, since there is no standardization in the sampling. Sampling was irregularly collected over the years, and this influences the number of individuals identified and the probability of resighting, as more photographs increase the chances of finding new individuals or resighting individuals already in the database. There was also a bias towards photos from the sheltered region, where most dives occur, which may have influenced the reported C. mydas and E. imbricata ratio. Despite all these biases, results from other studies support the use of data collected by citizen scientists. For example, to gather information on leatherback food items (Nordstrom et al. 2019), to analyze anthropogenic pressure on sea turtles (Papafitsoros et al. 2020), and to investigate the distribution and movements of individuals (Chassagneux et al. 2013; Baumbach and Dunbar 2017; Baumbach et al. 2019). Therefore, citizen-science monitoring programs can provide valuable and robust data to support scientific research.
The high number of photos acquired in a short period indicates the potential for exploring photo-ID for sea turtle monitoring in the region. Despite the intermittent temporal distribution of photos in our results, older registers were essential to establishing baseline information on the presence of turtles over multiple years. The number of images from social media screening and the contact with the public highlights the potential of social media to increase data collection. It is noteworthy that the use of social media in Brazil is recent, so the collection of image collection from social media screening was limited to 15 years before present, with the highest number of images between 2018 and 2020. In 2020, the COVID-19 pandemic began just after the first quarter of the year. However, the increasing trend in the number of photos continued. In Brazil, this period is dedicated to summer holidays, when diving activities are frequent in Arraial do Cabo. However, in 2021 the number of dives decreased throughout the year and therefore the number of images had an interruption in the upward trend. Another topic to be mentioned is that data collection occurred until October 2021, at the beginning of the relaxation of public health measures to containing the pandemic. Therefore, 2021 did not include the summer period, which contributed to the low number of photos.
Given the complex logistics of collecting sea turtle data underwater and the need for more information on sea turtles at foraging grounds (Wildermann et al. 2018), extracting as much information as possible with low cost and less animal manipulation is highly valuable for research purposes. Photos and videos can provide a snapshot of individuals’ characteristics. We used the images for individual temporal and spatial description and for the temporal evaluation of FP. Considering the existing limitations (image quality, sampling bias, and body area variability), media files can also be used for other approaches, including injury, biofouling and behavior monitoring, as already evaluated with digital media (Goatley et al. 2012; Hall and McNeill 2013; Ciccione et al. 2015; Wright et al. 2020; Hanna et al. 2021). Despite its great utility and simple application, photo-ID, particularly with citizen-science data, lacks the manipulation necessary for other data collection (health checks, genetics, precise measurements). However, even in surveys that require direct capture, photo-ID can be used in parallel with flipper tagging as another possible marking technique.
Diver propensity to fill out online forms must be considered for new citizen-science activities. Here, divers were more willing to share images, an activity more common in underwater tourism, than to take time to fill out dive forms. Beyond efforts to maintain constant interaction with volunteers and attract new ones (Martin et al. 2016), the experience during this work showed that incorporating new practices should be carefully planned together with the volunteers to increase acceptance. In our case, a closer interaction with the participants was required to reinforce the importance of the dive forms and feedback. In the future, scheduled meetings with stakeholders may facilitate data acquisition, as well as try to provide printed forms to dive operators.
Spatial and temporal description
The higher frequency of C. mydas in relation to E. imbricata corroborates previous studies carried out in Brazil (Baptistotte 2007; Werneck et al. 2018; Tagliolatto et al. 2020). Despite the low number of diving forms, all those analyzed provided information of at least one turtle sighting. However, the sighting rates (1.7 turtles·hour−1 for C. mydas and 0.72 turtle·hour−1 E. imbricata) were lower than those previously found for the same study area (8.61 C. mydas and 0.87 E. imbricata·hour−1) (Mello-Fonseca et al. 2021). Diving activity occurs between 3 and 11 m, and overlaps with the habitat use of both species, therefore sea turtles can be easily observed during recreational dives. Arraial do Cabo is a high-density area of juvenile C. mydas, where most individuals forage in shallow habitats, down to 6 m (Mello-Fonseca et al. 2021). Although the habitat use of E. imbricada in Arraial do Cabo is unknown, in other foraging grounds, individuals prefer shallow areas for foraging and resting, ranging from 2 to 20 m (Houghton et al. 2003; Blumenthal et al. 2009; Selby et at. 2019). Thus, differences in the sighting rates may reflect the low number of available forms (19 diving forms), the size of the surveyed area, and method used, as volunteers do not conduct dedicated observations. Moreover, the diver-turtle interaction is biased towards individuals with a possible neutral behavior to the divers’ approach, missing those using deeper depths, hiding in caves or even more unapproachable individuals (Papafitsoros 2015; Hayes et al. 2016).
The low number of resighted individuals may indicate transient individuals using Arraial do Cabo as a migratory corridor. Resighting can be influenced by individuals with a larger home range, related to spatial variability that causes individuals to displace between patches and different habitats for foraging and sleeping (Makowski et al. 2006; Scales et al. 2011; Christiansen et al. 2017; Wood et al. 2017) and under seasonal variations in temperatures, sea turtles can also undertake displacements to warmer regions, temporarily changing home range (Metz et al. 2020). Furthermore, mortality caused by small-scale fishing and interaction with debris may influence the resighting rate of individuals, as these threats have already been recorded for juvenile turtles in Arraial do Cabo (Tagliolatto et al. 2020). As we used opportunistic data, it is possible that recorded dives were inconsistent throughout the year, and this may also explain the low number of resighted individuals, resulting in longer intervals between resighting events.
Assuming the temporal gaps in our data, some turtles exhibited considerable residence time. The results show turtles occupying the region for at least 6 years. Since the annual contribution was not constant, it was not possible to define minimum and maximum residence time. In an area located 315 km south of our study area (Ubatuba-SP), the majority of C. mydas (93.4%, n = 141) were resighted in less than 1 year (maximum of ≅ 2.3 years) during an 8-year study (Gallo et al. 2006). On the tropical oceanic island of Fernando de Noronha, the mean recapture time for E. imbricata was 3.9 years (min. 2 days, max. 14 years) during a long-term study (31 years) (Bellini et al. 2019). Patterns of fidelity can be explained by habitat quality, which include optimal temperature and prey availability (Metz et al. 2020). Chelonia mydas is essentially herbivorous but may opportunistically feed on jellyfish, fishes and molluscs (Awabdi et al. 2013; Di Beneditto et al. 2017), while the diet of E. imbricata may include sessile invertebrates, such as sponges, corals and zoanthids (Berube et al. 2012; Martins et al. 2020). Arraial do Cabo sustains a high abundance of macroalgae (Brasileiro et al. 2009) and a rich benthic community (Ferreira et al. 1998; Rogers et al. 2014; Aued et al. 2018), with both tropical and warm-temperate affinity components. These characteristics favor the occurrence of a high density of turtles (Mello-Fonseca et al. 2021), with individuals residing for years, pointing to Arraial do Cabo as a potentially important development foraging ground for both species (Awabdi et al. 2013; di Beneditto et al. 2017).
The life cycles of sea turtles typically cross geopolitical boundaries, making population conservation initiatives an international issue and a challenge that requires synergistic efforts, including work on the nesting and foraging grounds. Although the natal origin of individuals was not the focus of this study, mixed stocks in Brazil are generally composed of turtles from Brazil, the Caribbean, Mid-Atlantic Islands, and West Africa (Bjorndal et al. 2006; Proietti et al. 2014b). Over the last few decades, nesting grounds in many countries have received some sort of conservation status, resulting in a measure of successful population growth (Mazaris et al. 2017). Arraial do Cabo is a foraging ground with multiple anthropogenic impacts, such as intense SCUBA diving, artisanal fishing, boat traffic and pollutants (Giglio et al. 2017a, b; Lima et al. 2018). In the current research, turtles were seen at the same or at nearby dive sites where some individuals were constantly resighted. Sea turtles have been reported to have high fidelity to small areas, either for foraging or resting (Berube et al. 2012; Christiansen et al. 2017; Petit et al. 2020), while recreational dives are limited to specific dive points (Pedrini et al. 2007). Therefore, divers are probably observing the same individuals and the resident turtles are continuously exposed to anthropogenic pressures (Papafitsoros et al. 2020). Although individuals may become accustomed to non-intrusive observers (von Brandis et al. 2010), boat strikes and divers’ approach may affect turtles behavior and metabolism (Hayes et al. 2016; Wright et al. 2020). As a development foraging ground, it is critical to propose significant management measures to protect sea turtles in Arraial do Cabo, for instance, zoning and turtle-diver interaction protocols.
Fibropapillomatosis (FP)
The infection prevalence rate of FP between 2006 and 2021 was comparatively lower than many other foraging grounds and, in our site demonstrated interannual fluctuation (Table S2 and 2). These results must be interpreted with caution, as the method used did not allow a full body analysis, so that prevalence value may actually be higher than reported here. FP has been related to eutrophic conditions of coastal environments (dos Santos et al. 2010) and may be associated with habitats where sea surface temperature is high (> 30 °C) (Dujon et al. 2021). Arraial do Cabo is an urbanized municipality with a variety of pollution sources. For instance, the sheltered region hosts a harbor, marina, pluvial channel, sewage discharge, and intense boat traffic, which are likely to sustain high incidences of infected turtles. However, we still lack knowledge on how internal water circulation can disperse contaminants (Godoy et al. 2013), and how upwelling lower temperatures can buffer the prevalence of the disease (Dujon et al. 2021). Furthermore, the high incidence of recruits and small juveniles (≤ 40 cm SCL for both species) (Mello-Fonseca et al. 2021) can support the low prevalence rate, since they may have just arrived in the near-shore area and may only recently had contact with the virus (Aguirre and Lutz 2004).
Individuals have different propensities to present FP (Alfaro-Núñez et al. 2014), so changes in the composition of individuals may be associated with annual prevalence variation. Although Arraial do Cabo appears to have a comparatively lower FP prevalence than other study areas, indicating a possible healthy foraging ground, the overall regression rate was lower than other foraging grounds (Table 2). As many turtles are not resighted, it was not possible to assess more progression or regression cases. It is also worth mentioning that we had limited images for each animal, which only allowed us to confirm regression or progression in certain regions of the animals. Therefore, our analysis does not clarify whether FP was regressing or progressing elsewhere on the individuals. FP is a major cause of death among sea turtles in southeastern Brazil (Tagliolatto et al. 2020) and requires dedicated monitoring to arrest disease evolution.
Conclusion
We demonstrated that social media can be useful for monitoring charismatic megafauna, such as sea turtles. Using photo-ID, it was possible to detect resident individuals occupying Arraial do Cabo for several years and to record cases of FP, including tumor regression. Since Arraial do Cabo has several threats to sea turtles, we highlight the need for further studies to fill the gaps considering habitat use and anthropogenic impacts on them. We also emphasize the need for significant management measures to protect both species and the marine ecosystem. A photo-ID approach using data submitted by citizen scientists provided sea turtle estimates in a foraging ground using a non-invasive method, underpinning benefits, such as low cost, and promoting communication with the public about science and conservation. This combination can be applied to other locations to obtain similar kinds of data and increase knowledge of sea turtle demographics and population trends.
Data availability
The database generated and analyzed during the current study is available from the corresponding author upon reasonable request.
References
Adnyana W, Ladds P, Blair D (1997) Observations of fibropapillomatosis in green turtles (Chelonia mydas) in Indonesia. Aust Vet J 75:737–742. https://doi.org/10.1111/j.1751-0813.1997.tb12258.x
Aguirre AA, Lutz PL (2004) Marine turtles as sentinels of ecosystem health: is fibropapillomatosis an indicator? EcoHealth 1:275–283. https://doi.org/10.1007/s10393-004-0097-3
Alfaro-Núñez A, Frost BM, Bojesen AM, Rasmussen I, Zepeda-Mendoza L, Tange OM, Gilbert MTP (2014) Global distribution of Chelonid fibropapilloma-associated herpesvirus among clinically healthy sea turtles. BMC Evol Biol 14:206. https://doi.org/10.1186/s12862-014-0206-z
Araujo G, Montgomery J, Pahang K, Labaja J, Murray R, Ponzo A (2016) Using minimally invasive techniques to determine green sea turtle Chelonia mydas life-history parameters. J Exp Mar Biol Ecol 483:25–30. https://doi.org/10.1016/j.jembe.2016.06.004
Aued AW, Smith F, Quimbayo JP, Cândido DV, Longo GO, Ferreira CEL, Witman JD, Floeter SR, Segal B (2018) Large-scale patterns of benthic marine communities in the Brazilian Province. PLoS ONE 13:e0198452. https://doi.org/10.1371/journal.pone.0198452
Awabdi DR, Siciliano S, di Beneditto APM (2013) First information about the stomach contents of juvenile green turtles, Chelonia mydas, in Rio de Janeiro, south-eastern Brazil. Mar Biodivers Rec 6:e5. https://doi.org/10.1017/S1755267212001029
Baptistotte C (2007) Caracterização espacial e temporal da fibropapilomatose em tartarugas marinhas da costa brasileira. PhD Thesis, University of São Paulo. https://doi.org/10.11606/T.91.2007.tde-11032008-153152
Baumbach DS, Dunbar SG (2017) Animal mapping using a citizen-science web-based GIS in the Bay Islands, Honduras. Mar Turtle Newsl 152:16
Baumbach DS, Anger EC, Collado NA, Dunbar SG (2019) Identifying sea turtle home ranges utilizing citizen-science data from novel web-based and smartphone GIS applications. Chelonian Conserv Biol 18:133. https://doi.org/10.2744/CCB-1355.1
Becken S, Connolly RM, Chen J, Stantic B (2019) A hybrid is born: integrating collective sensing, citizen science and professional monitoring of the environment. Ecol Inform 52:35–45. https://doi.org/10.1016/j.ecoinf.2019.05.001
Bellini C, Santos A, Patrício A, Bortolon L, Godley B, Marcovaldi M, Tilley D, Colman L (2019) Distribution and growth rates of immature hawksbill turtles Eretmochelys imbricata in Fernando de Noronha, Brazil. Endang Species Res 40:41–52. https://doi.org/10.3354/esr00979
Berube MD, Dunbar SG, Rützler K, Hayes WK (2012) Home range and foraging ecology of juvenile hawksbill sea turtles (Eretmochelys imbricata) on inshore reefs of Honduras. Chelonian Conserv Biol 11:33–43. https://doi.org/10.2744/CCB-0898.1
Bjorndal KA, Bolten AB, Moreira L, Bellini C, Marcovaldi MÂ (2006) Population structure and diverazilianbrazilian green turtle rookeries based on mitochondrial DNA sequences. Chelonian Conserv Biol 5:262–268. https://doi.org/10.2744/1071-8443(2006)5[262:PSADOB]2.0.CO;2
Blumenthal JM, Austin TJ, Bothwell JB et al (2009) Diving behavior and movements of juvenile hawksbill turtles Eretmochelys imbricata on a Caribbean coral reef. Coral Reefs 28:55–65. https://doi.org/10.1007/s00338-008-0416-1
Bolten AB (2003) Variation in sea turtle life history patterns: neritic vs. oceanic developmental stages. In: Lutz PL, Musick JA, Wyneken J (eds) The biology of sea turtles, 2nd edn. CRC Press, Florida, pp 243–257
Bourlat SJ, Borja A, Gilbert J, Taylor MI, Davies N, Weisberg SB, Griffith JF, Lettieri T, Field D, Benzie J, Glöckner FO, Rodríguez-Ezpeleta N, Faith DP, Bean TP, Obst M (2013) Genomics in marine monitoring: New opportunities for assessing marine health status. Mar Pollut Bull 74:19–31. https://doi.org/10.1016/j.marpolbul.2013.05.042
Brjáileiro PS, Yoneshigue-Valentin Y, Bahia RDG, Reis RP, Amado Filho GM (2009) Algas marinhas bentônicas da região de Cabo Frio e arredores: síntese do conhecimento. Rodriguésia 60:39–66. https://doi.org/10.1590/2175-7860200960103
Calmanovici B, Waayers D, Reisser J, Clifton J, Proietti M (2018) I3S Pattern as a mark-recapture tool to identify captured and free-swimming sea turtles: an assessment. Mar Ecol Prog Ser 589:263–268. https://doi.org/10.3354/meps12483
Carpentier AS, Jean C, Barret M, Chassagneux A, Ciccione S (2016) Stability of facial scale patterns on green sea turtles Chelonia mydas over time: a validation for the use of a photo-identification method. J Exp Mar Biol Ecol 476:15–21. https://doi.org/10.1016/j.jembe.2015.12.003
Carter SJB, Bell IP, Miller JJ, Gash PP (2014) Automated marine turtle photograph identification using artificial neural networks, with application to green turtles. J Exp Mar Biol Ecol 452:105–110. https://doi.org/10.1016/j.jembe.2013.12.010
Casale P, Mazaris AD, Freggi D, Vallini C, Argano R (2009) Growth rates and age at adult size of loggerhead sea turtles (Caretta caretta) in the Mediterranean Sea, estimated through capture-mark-recapture records. Sci Mar 73:589–595. https://doi.org/10.3989/scimar.2009.73n3589
Casale P, Freggi D, Salvemini P (2017) Tag loss is a minor limiting factor in sea turtle tagging programs relying on distant tag returns: the case of Mediterranean loggerhead sea turtles. Eur J Wildl Res 63:12. https://doi.org/10.1007/s10344-016-1059-0
Castelblanco-Martínez DN, Blanco-Parra MP, Charruau P, Prezas B, Zamora-Vilchis I, Niño-Torres CA (2019) Detecting, counting and following the giants of the sea: a review of monitoring methods for aquatic megavertebrates in the Caribbean. Wildl Res 46:545. https://doi.org/10.1071/WR19008
Chassagneux A, Jean C, Bourjea J, Ciccione S (2013) Unraveling behavioral patterns of foraging hawksbill and green turtles using photo-identification. Mar Turtle Newsl 137:1–5
Christiansen F, Esteban N, Mortimer JA, Dujon AM, Hays GC (2017) Diel and seasonal patterns in activity and home range size of green turtles on their foraging grounds revealed by extended Fastloc-GPS tracking. Mar Biol 164:10. https://doi.org/10.1007/s00227-016-3048-y
Ciccione S, Jean C, Carpentier A, Barret M (2015) Cause and healing of a sea turtle injury revealed by photoidentification. Indian Ocean Turtle Newslett 21:10–12
Cordeiro CAMM, Harborne AR, Ferreira CEL (2020) The biophysical controls of macroalgal growth on subtropical reefs. Front Mar Sci 7:1–14. https://doi.org/10.3389/fmars.2020.00488
Crall JP, Stewart CV, Berger-Wolf TY, Rubenstein DI, Sundaresan SR (2013) otSpotter—Patterned species instance recognition. In: 2013 IEEE Workshop on Applications of Computer Vision (WACV). p 230–37. https://doi.org/10.1109/WACV.2013.6475023
Crossin G, Cooke S, Goldbogen J, Phillips R (2014) Tracking fitness in marine vertebrates: current knowledge and opportunities for future research. Mar Ecol Prog Ser 496:1–17. https://doi.org/10.3354/meps10691
de Farias DSD, de Alencar AEB, Bomfim ADC, Fragoso ABDL, Rossi S, Moura GJB, Gavilan SA, Silva FJDL (2019) Marine turtles stranded in northeastern Brazil: composition, spatio-temporal distribution, and anthropogenic interactions. Chelonian Conserv Biol 18:105. https://doi.org/10.2744/CCB-1309.1
di Beneditto APM, Siciliano S, Monteiro LR (2017) Herbivory level and niche breadth of juvenile green turtles (Chelonia mydas) in a tropical coastal area: insights from stable isotopes. Mar Biol 164:13. https://doi.org/10.1007/s00227-016-3044-2
dos Santos R, Martins A, Torezani E, Baptistotte C, Farias J, Horta P, Work T, Balazs G (2010) Relationship between fibropapillomatosis and environmental quality: a case study with Chelonia mydas off Brazil. Dis Aquat Organ 89:87–95. https://doi.org/10.3354/dao02178
Dujon AM, Schofield G, Venegas RM, Thomas F, Ujvari B (2021) Sea turtles in the cancer risk landscape: a global meta-analysis of fibropapillomatosis prevalence and associated risk factors. Pathogens 10:1295. https://doi.org/10.3390/pathogens10101295
Dunbar SG, Ito HE, Bahjri K, Dehom S, Salinas L (2014) Recognition of juvenile hawksbills Eretmochelys imbricata through face scale digitization and automated searching. Endang Species Res 26:137–146. https://doi.org/10.3354/esr00637
Dunbar SG, Anger EC, Parham JR, Kingen C, Wright MK, Hayes CT, Safi S, Holmberg J, Salinas L, Baumbach DS (2021) HotSpotter: using a computer-driven photo-id application to identify sea turtles. J Exp Mar Biol Ecol 535:151490. https://doi.org/10.1016/j.jembe.2020.151490
Ferreira CEL, Gonçalves JEA, Coutinho R, Peret AC (1998) Herbivory by the dusky damselfish Stegastes fuscus (Cuvier, 1830) in a tropical rocky shore: effects on the benthic community. J Exp Mar Biol Ecol 229:241–264. https://doi.org/10.1016/S0022-0981(98)00056-2
Ferreira CEL, Goncçalves JEA, Coutinho R (2001) Community structure of fishes and habitat complexity on a tropical rocky shore. Environ Biol Fish 61:353–369. https://doi.org/10.1023/A:1011609617330
Gallo BMG, Macedo S, de Giffoni BB, Becker JH, Barata PCR, Gallo BMG, Barata PCR, Macedo S, Becker JH (2006) Sea turtle conservation in Ubatuba, southeastern Brazil, a feeding area with incidental capture in coastal fisheries. Chelonian Conserv Biol 5:93. https://doi.org/10.2744/1071-8443(2006)5[93:STCIUS]2.0.CO;2
Gatto CR, Rotger A, Robinson NJ, Santidrián Tomillo P (2018) A novel method for photo-identification of sea turtles using scale patterns on the front flippers. J Exp Mar Biol Ecol 506:18–24. https://doi.org/10.1016/j.jembe.2018.05.007
Giglio VJ, Ternes MLF, Mendes TC, Cordeiro CAMM, Ferreira CEL (2017a) Anchoring damages to benthic organisms in a subtropical scuba dive hotspot. J Coast Conserv 21:311–316. https://doi.org/10.1007/s11852-017-0507-7
Giglio VJ, Bender MG, Zapelini C, Ferreira CEL (2017b) The end of the line? Rapid depletion of a large-sized grouper through spearfishing in a subtropical marginal reef. Perspect Ecol Conserv 15:115–118. https://doi.org/10.1016/j.pecon.2017.03.006
Giglio VJ, Luiz OJ, Chadwick NE, Ferreira CEL (2018) Using an educational video-briefing to mitigate the ecological impacts of scuba diving. J Sustain Tour 26:782–797. https://doi.org/10.1080/09669582.2017.1408636
Gillis AJ, Wildermann NE, Ceriani SA, Seminoff JA, Fuentes MM (2020) Evaluating different spatial scales of forage item availability to determine diet selection of juvenile green turtles (Chelonia mydas). Mar Biol 167:1–17. https://doi.org/10.1007/s00227-020-03782-y
Goatley CHR, Hoey AS, Bellwood DR (2012) The role of turtles as coral reef macroherbivores. PLoS ONE 7:e39979. https://doi.org/10.1371/journal.pone.0039979
Godoy JM, Souza TA, Godoy MLDP, Moreira I, Carvalho ZL, Lacerda LD, Fernandes FC (2013) Groundwater and surface water quality in a coastal bay with negligible fresh groundwater discharge: Arraial do Cabo, Brazil. Mar Chem 156:85–97. https://doi.org/10.1016/j.marchem.2013.05.004
Guimaraens MA, Coutinho R (1996) Spatial and temporal variation of benthic marine algae at the Cabo Frio upwelling region, Rio de Janeiro, Brazil. Aquat Bot 52:283–299. https://doi.org/10.1016/0304-3770(95)00511-0
Hall AG, McNeill JB (2013) Inferring sea turtle recapture rates using photographic identification. Herpetol Rev 44:561–569
Hanna ME, Chandler EM, Semmens BX, Eguchi T, Lemons GE, Seminoff JA (2021) Citizen-sourced sightings and underwater photography reveal novel insights about green sea turtle distribution and ecology in southern California. Front Mar Sci. https://doi.org/10.3389/fmars.2021.671061
Hayes CT, Baumbach DS, Juma D, Dunbar SG (2016) Impacts of recreational diving on hawksbill sea turtle (Eretmochelys imbricata) behaviour in a marine protected area. J Sustain Tour 25:79–95. https://doi.org/10.1080/09669582.2016.1174246
Hebblewhite M, Haydon DT (2010) Distinguishing technology from biology: a critical review of the use of GPS telemetry data in ecology. Philos Trans R Soc B 365:2303–2312. https://doi.org/10.1098/rstb.2010.0087
Hirama S, Ehrhart LM (2007) Description, prevalence and severity of green turtle fibropapillomatosis in three developmental habitats on the east coast of Florida. Florida Sci 70:435–448
Houghton JD, Callow MJ, Hays GC (2003) Habitat utilization by juvenile hawksbill turtles (Eretmochelys imbricata, Linnaeus, 1766) around a shallow water coral reef. J Nat Hist 37:1269–1280. https://doi.org/10.1080/00222930110104276
IUCN (2022) The IUCN red list of threatened species. In: Version 2021-3. https://www.iucnredlist.org. Accessed 09 Nov 2022
Kanive PE, Rotella JJ, Jorgensen SJ, Chapple TK, Hines JE, Anderson SD, Block BA (2019) Size-specific apparent survival rate estimates of white sharks using mark–recapture models. Can J Fish Aquat Sci 76:2027–2034. https://doi.org/10.1139/cjfas-2018-0142
Katsanevakis S, Weber A, Pipitone C, Leopold M, Cronin M, Scheidat M, Doyle T, Buhl-Mortensen L, Buhl-Mortensen P, D’Anna G et al (2012) Monitoring marine populations and communities: methods dealing with imperfect detectability. Aquat Biol 16:31–52. https://doi.org/10.3354/ab00426
Lima SR, Barbosa JMS, Padilha FGF, Saracchini PGV, Braga MA, Leite JS, Ferreira AMR (2018) Physical characteristics of free-living sea turtles that had and had not ingested debris in Microregion of the Lakes, Brazil. Mar Pollut Bull 137:723–727. https://doi.org/10.1016/j.marpolbul.2018.10.032
Lucrezi S, Milanese M, Palma M, Cerrano C (2018) Stirring the strategic direction of scuba diving marine citizen science: a survey of active and potential participants. PLoS ONE 13:1–28. https://doi.org/10.1371/journal.pone.0202484
Machado GS, Mas GH, Vidal WA, Monteiro-Neto C, Lobo-Hajdu G (2013) Evidence of regression of fibropapillomas in juvenile green turtles Chelonia mydas caught in Niterói, southeast Brazil. Dis Aquat Organ 102:243–247. https://doi.org/10.3354/dao02542
Makowski C, Seminoff JA, Salmon M (2006) Home range and habitat use of juvenile Atlantic green turtles (Chelonia mydas L.) on shallow reef habitats in Palm Beach, Florida, USA. Mar Biol 148:1167–1179. https://doi.org/10.1007/s00227-005-0150-y
Martin V, Smith L, Bowling A, Christidis L, Lloyd D, Pecl G (2016) Citizens as scientists: what influences public contributions to marine research? Sci Commun 38:495–522. https://doi.org/10.1177/1075547016656191
Martins RF, Andrades R, Nagaoka SM, Martins AS, Longo LL, Ferreira JS, Bastos KV, Joyeux JC, Santos RG (2020) Niche partitioning between sea turtles in waters of a protected tropical island. Reg Stud Mar Sci 39:101439. https://doi.org/10.1016/j.rsma.2020.101439
Mazaris AD, Schofield G, Gkazinou C, Almpanidou V, Hays GC (2017) Global sea turtle conservation successes. Sci Adv 3:e1600730. https://doi.org/10.1126/sciadv.1600730
Mello-Fonseca J, Cordeiro C, Ferreira C (2021) Spatial distribution of sea turtles on South Atlantic subtropical reefs. Mar Ecol Prog Ser 678:125–138. https://doi.org/10.3354/meps13860
Metz TL, Gordon M, Mokrech M, Guillen G (2020) Movements of juvenile green turtles (Chelonia mydas) in the nearshore waters of the northwestern Gulf of Mexico. Front Mar Sci 7:1–17. https://doi.org/10.3389/fmars.2020.00647
Monteiro DS, Estima SC, Gandra TBR, Silva AP, Bugoni L, Swimmer Y, Seminoff JA, Secchi ER (2016) Long-term spatial and temporal patterns of sea turtle strandings in southern Brazil. Mar Biol 163:247. https://doi.org/10.1007/s00227-016-3018-4
Nielsen JM, Clare EL, Hayden B, Brett MT, Kratina P (2017) Diet tracing in ecology: Method comparison and selection. Methods Ecol Evol 9:278–291. https://doi.org/10.1111/2041-210X.12869
Nordstrom B, James M, Martin K, Worm B (2019) Tracking jellyfish and leatherback sea turtle seasonality through citizen science observers. Mar Ecol Prog Ser 620:15–32. https://doi.org/10.3354/meps12978
Papafitsoros K (2015) In-water behaviour of the loggerhead sea turtle (Caretta caretta) under the presence of humans (Homo sapiens) in a major Mediterranean nesting site. In: Kaska Y, Sönmez B, Türkecan O, Sezgin Ç (eds) Book of abstracts of the 35th annual symposium on sea turtle biology and conservation. MACARTpress, Turkey, p 47
Papafitsoros K, Panagopoulou A, Schofield G (2020) Social media reveals consistently disproportionate tourism pressure on a threatened marine vertebrate. Animal Conserv. https://doi.org/10.1111/acv.12656
Pedrini ADG, Maneschy FA, Costa C, Almeida ER, Costa EA (2007) Gestão ambiental em áreas protegidas x estatísticas de mergulho na RESEX marinha de Arraial do Cabo, RJ. OLAM Ciência Tecnologia Rio Claro/SP Brasil 2:270–294
Petit I, González C, Gusmao J, Álvarez-Varas R, Hinojosa I (2020) Resting dynamics and diel activity of the green turtle (Chelonia mydas) in Rapa Nui, Chile. Chelonian Conserv Biol 19:124. https://doi.org/10.2744/CCB-1374.1
Proietti MC, Reisser J, Secci ER (2014a) Foraging by immature hawksbill sea turtles at Brazilian islands. Mar Turtle Newslett 135:4–6
Proietti MC, Reisser J, Marins LF, Rodriguez-Zarate C, Marcovaldi MA, Monteiro DS, Pattiaratchi C, Secchi ER (2014b) Genetic structure and natal origins of immature hawksbill turtles (Eretmochelys imbraziliann Brazilian waters. PLoS ONE 9:e88746. https://doi.org/10.1371/journal.pone.0088746
Reisser J, Proietti M, Kinas P, Sazima I (2008) Photographic identification of sea turtles: method description and validation, with an estimation of tag loss. Endang Species Res 5:73–82. https://doi.org/10.3354/esr00113
Rogers R, Correal GO, Oliveira TC et al (2014) Coral health rapid assessment in marginal reef sites. Mar Biol Res 10:612–624. https://doi.org/10.1080/17451000.2013.841944
Rossi S, Sánchez-Sarmiento AM, dos Santos RG, Zamana RR, Prioste FES, Gattamorta MA, Ochoa PFC, Grisi-Filho JHH, Matushima ER (2019) Monitoring green sea turtles in Brazilian feeding areas: relating body condition index to fibropapillomatosis prevalence. J Mar Biol Assoc Uk 99:1879–1887. https://doi.org/10.1017/S0025315419000730
Santos AS, Almeida AP, Santos AJB et al (2011) Plano de ação nacional para a conservação das tartarugas marinhas. Instituto Chico Mendes de Conservação da Biodiversidade ICMBio, Brasília
Scales L, Lewis JA, Lewis JP, Castellanos D, Godley BJ, Graham RT (2011) Insights into habitat utilisation of the hawksbill turtle, Eretmochelys imbricata (Linnaeus, 1766), using acoustic telemetry. J Exp Mar Biol Ecol 407:122–129. https://doi.org/10.1016/j.jembe.2011.07.008
Schofield G, Katselidis KA, Dimopoulos P, Pantis JD (2008) Investigating the viability of photo-identification as an objective tool to study endangered sea turtle populations. J Exp Mar Biol Ecol 360:103–108. https://doi.org/10.1016/j.jembe.2008.04.005
Schofield G, Klaassen M, Papafitsoros K, Lilley MKS, Katselidis KA, Hays GC (2020) Long-term photo-id and satellite tracking reveal sex-biased survival linked to movements in an endangered species. Ecology 101:1–10. https://doi.org/10.1002/ecy.3027
Selby TH, Hart KM, Smith BJ, Pollock CG, Hillis-Starr Z, Oli MK (2019) Juvenile hawksbill residency and habitat use within a Caribbean marine protected area. Endang Species Res 40:53–64. https://doi.org/10.3354/esr00975
Shaver D, Walker J, Backof T (2019) Fibropapillomatosis prevalence and distribution in green turtles Chelonia mydas in Texas (USA). Dis Aquat Organ 136:175–182. https://doi.org/10.3354/dao03403
Su CM, Huang CT, Cheng IJ (2015) Applying a fast, effective and reliable photographic identification system for green turtles in the waters near Luichiu Island, Taiwan. J Exp Mar Biol Ecol 467:115–120. https://doi.org/10.1016/j.jembe.2015.03.003
Tabuki K, Nishizawa H, Abe O, Okuyama J, Tanizaki S (2021) Utility of carapace images for long-term photographic identification of nesting green turtles. J Exp Mar Biol Ecol 545:151632. https://doi.org/10.1016/j.jembe.2021.151632
Tagliolatto A, Guimarães S, Lobo-Hajdu G, Monteiro-Neto C (2016) Characterization of fibropapillomatosis in green turtles Chelonia mydas (Cheloniidae) captured in a foraging area in southeastern Brazil. Dis Aquat Organ 121:233–240. https://doi.org/10.3354/dao03020
Tagliolatto AB, Goldberg DW, Godfrey MH, Monteiro-Neto C (2020) Spatio-temporal distribution of sea turtle strandings and factors contributing to their mortality in south-eastern Brazil. Aquat Conserv Mar Freshwat Ecosyst 30:331–350. https://doi.org/10.1002/aqc.3244
Tourani M (2021) A review of spatial capture–recapture: ecological insights, limitations, and prospects. Ecol Evol 12:e8468. https://doi.org/10.1002/ece3.8468
Vieira EA, de Souza LR, Longo GO (2020) Diving into science and conservation: recreational divers can monitor reef assemblages. Perspect Ecol Conserv 18:51–59. https://doi.org/10.1016/j.pecon.2019.12.001
von Brandis RG, Mortimer JA, Reilly BK (2010) In-water observations of the diving behaviour of immature hawksbill turtles, Eretmochelys imbricata, on a coral reef at D’Arros Island, Republic of Seychelles. Chelonian Conserv Biol 9:26–32. https://doi.org/10.2744/CCB-0815.1
Werneck MR, Almeida D, Baldassin P, Guimarães S, Nunes LA, Lacerda PD, Oliveira ALM (2018) Sea turtle beach monitoring program in Brazil. In: Aguillón-Gutiérrez D (ed) Reptiles and amphibians. IntechOpen, London, pp 23–47
Wildermann N, Gredzens C, Avens L et al (2018) Informing research priorities for immature sea turtles through expert elicitation. Endang Species Res 37:55–76. https://doi.org/10.3354/esr00916
Williams JL, Pierce SJ, Rohner CA, Fuentes MMPB, Hamann M (2017) Spatial distribution and residency of green and loggerhead sea turtles using coastal reef habitats in southern Mozambique. Front Mar Sci. https://doi.org/10.3389/fmars.2016.00288
Wood LD, Brunnick B, Milton SL (2017) Home range and movement patterns of subadult hawksbill sea turtles in southeast Florida. J Herpetol 51:58–67. https://doi.org/10.1670/15-133
Wright MK, Baumbach DS, Collado N, Safi SB, Dunbar SG (2020) Influence of boat traffic on distribution and behavior of juvenile hawksbills foraging in a marine protected area in Roatán, Honduras. Ocean Coast Manag 198:105379. https://doi.org/10.1016/j.ocecoaman.2020.105379
Acknowledgements
The authors thank all the dive operators, divers and underwater photographers who participated in this work, sending photos and diving forms. We also thank the anonymous reviewers for their careful reading of our manuscript and their insightful suggestions. CELF is continually supported by grants from Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Funding
This study was financed in part by the Coordination for the Improvement of Higher Education Personnel – Brazil (CAPES) – Finance Code 001, through a scholarship granted to Isabella Neves-Ferreira.
Author information
Authors and Affiliations
Contributions
Conceptualization: IN-F and JM-F; formal analysis and investigation: IN-F; writing—original draft preparation IN-F; writing—review and editing: JM-F and CF; resources: CF; supervision: JM-F and CF.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest that would bias publication of this manuscript.
Ethics approval
Permission to handle the turtles was given by the ICMBio Institute (SISBio no 78142-1). All procedures performed are in accordance with the ethical standards of Centro TAMAR—ICMBio, Ministry of the Environment.
Additional information
Responsible Editor: P. Casale .
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Neves-Ferreira, I., Mello-Fonseca, J. & Ferreira, C.E.L. Photo-identification shows the spatio-temporal distribution of two sea turtle species in a Brazilian developmental foraging ground. Mar Biol 170, 83 (2023). https://doi.org/10.1007/s00227-023-04226-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-023-04226-z