
Abstract
Nanoparticles are emerging as a cutting-edge technology to improve crop agricultural input efficiency and reduce biotic and abiotic stresses. In viticulture, nanoparticles hold promise for the sustainable application of an elicitor (methyl jasmonate, MeJ), allowing a considerable dosage reduction. Herein, the influence of the foliar application of free MeJ (10 mM) and MeJ nanoformulation (ACP-MeJ, 1 mM MeJ) on Tempranillo grape amino acids content over two vintages (2019 and 2020) was evaluated. While both MeJ treatments provided a significant increase of the amino nitrogen and yeast assimilable nitrogen in the must in 2019, there were no significant differences on these parameters in 2020. In 2019, MeJ treatment enhanced the synthesis of most of the amino acids included in this study, while ACP-MeJ promoted the formation of six amino acids. Hence, the content of total amino acids, with and without proline, was higher after applying MeJ than in the control musts. However, these values were higher for control must than for MeJ samples in 2020. The multivariable analysis confirmed that the vintage factor had a more prominent effect on the overall parameters of the musts. This strong influence of the vintage could be related to the higher rainfall in 2020.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Nanomaterials have received an ever-increasing attention in the field of agrochemicals due to their exceptional properties, including their large surface area, higher chemical/thermal stability and tunable unique physicochemical characteristics (i.e., structure, solubility, surface reactivity) [1, 2]. Owing to these outstanding properties, nano-agrochemical delivery system has great potential for facilitating the uptake and translocation of nutrients in plants, improving the efficacy of agrochemicals, and consequently alleviating environmental pollution and promoting food security [3]. Recently, a novel nanoelicitor has been designed through the loading of biomimetic amorphous calcium phosphate nanoparticles, similar to those found in bone, with methyl jasmonate (ACP-MeJ) [4]. The nanoelicitor provided a sustainable release and protection against thermal degradation of MeJ, ensuring elicitor activity over longer period of times on the surface of the leaves and reducing by ten times the required dosage [4].
This nanoelicitor has been applied in the vineyard of two red grape varieties, Monastrell and Tempranillo, and the results, published to date, are collected in six scientific articles, focused on its effect on: (i) grape: Monastrell nitrogen composition [5] and Tempranillo phenolic composition [6]; (ii) wine: stilbenes content in Monastrell [4], volatile compounds in Monastrell [7] and a broad characterization of volatile, nitrogen and phenolic composition in Tempranillo [8]; and (iii) grape and wine: nitrogen composition in Monastrell [9].
Regarding Monastrell grape variety, it has been observed that the application of MeJ, free or in nano-form, increased the total content of amino acids in grapes, although to a greater extent when used conventionally, i.e., as free [9]. However, Pérez-Álvarez et al. [5], when applying ACP-MeJ in this same grape variety, did not observe influence on the nitrogen composition of the grapes. As for Monastrell wine, the use of MeJ and ACP-MeJ in the vineyard increased the content of total stilbenes [4] and total amino acids [9], while the influence on the wine volatile composition depended on the family of compounds and the vintage: total acids increased in one of the vintages, and total alcohols and esters were not affected in any of the vintages, while terpenoids increased when applying conventional MeJ in all vintages [7].
Regarding the Tempranillo variety, the treatments in the vineyard with MeJ and ACP-MeJ improved the content of phenolic compounds in the grapes, the effect being dependent on the vintage, so that, in a vintage, MeJ increased the total content of anthocyanins and ACP-MeJ that of hydroxycinnamic acids (HCA); while, in the second vintage, the application of MeJ had no effect on the phenolic composition of the grapes and, however, ACP-MeJ increased the total content of flavonols, flavanols, gallic acid, and stilbenes [6]. Regarding the Tempranillo wine [8], its phenolic composition was little affected by the treatments with MeJ and ACP-MeJ carried out in the vineyard, since only the content of gallic acid and total HCA showed changes; furthermore, the total amino acids content was little affected; and volatile compounds decreased their concentration when applying both treatments in the first vintage, while in the second one, these compounds were not affected by ACP-MeJ, while esters and acids decreased again when applying MeJ, without effect on alcohols.
Therefore, the study of the nitrogen composition of Tempranillo grapes, after the application of MeJ and ACP-MeJ in the vineyard, is a key issue, since there are no previous studies on this topic, unlike in the Monastrell grape variety [5, 9]. Nitrogen compounds are essential for the correct development of alcoholic fermentation [10, 11]. Moreover, these compounds are precursors of the main fermentative volatile compounds [12, 13], and have been especially affected by the applications in the Tempranillo vineyard of MeJ and ACP-MeJ [8], although the same effect was not observed when the treatments were applied in Monastrell [7], despite the important difference in the nitrogen composition of the grapes [9]. For all these reasons, the aim of this work was to study the influence of MeJ and ACP-MeJ foliar treatments on Tempranillo grape amino acids content over two vintages.
Materials and methods
Foliar treatments and must samples
Tempranillo (Vitis vinifera L.) variety grown in Finca La Grajera, Logroño, La Rioja, Spain (42° 26′ 25″ North, Latitude; 2° 30′ 56″ West, Longitude; 456 m above sea level) in 2019 and 2020 vintages was used. Vineyard had grafted onto R-110 rootstock and trained to a vertical shoot positioned trellis system, had been planted in 1997. Climate data were recorded by the Agroclimatic Information Service of La Rioja (SIAR) installed close to field. The collected data were the rain accumulated from the beginning of April until 1st of September, being 247.8 L/m2 in 2019 and 217.8 L/m2 in 2020; and the average maximum, mean, and minimum temperatures, being 27.0 °C, 13.8 °C, and 3.7 °C, respectively, in 2019, and 26.3 °C, 13.8 °C, and 3.7 °C, respectively, in 2020.
The experiment involved the foliar application of free methyl jasmonate (MeJ) and amorphous calcium phosphate nanoparticles doped with this elicitor (ACP-MeJ). The synthesis and characterization of ACP-MeJ to determine the composition structure and morphology were carried out as previously described elsewhere [4]. To carry out the treatments, aqueous solutions were prepared with a concentration 10 mM of MeJ (according to previous works) [14, 15] and 1 mM of ACP-MeJ [5, 6], using Tween® 80 (Sigma-Aldrich, Madrid, Spain) as wetting agent (1 mL/L). Control plants were sprayed with Tween® 80 water solution. The foliar applications were carried out twice, at veraison and 7 days later on. The treatments were arranged in a complete randomized block design, with ten vines for replication and treatment, and were done in triplicate.
Grapes from all grapevines and treatments were picked at their optimum technological maturity, and they were destemmed and crushed to obtain the musts. Then, the general parameters were determined, and aliquots of each must sample were frozen at − 20° C for later analysis of nitrogen composition.
General enology parameters’ determination
The must enological parameters were analyzed using the official methods of OIV [16]: ºBrix, probable alcohol, pH, and total acidity. Glucose, fructose, malic acid, and nitrogen fractions [ammonium nitrogen, amino nitrogen, and yeast assimilable nitrogen (YAN)] were determined using a Miura One enzymatic equipment (TDI, Barcelona, Spain).
As the treatments were performed in triplicate, the results of these parameters are shown as the average of three analyses (n = 3).
Analysis of amino acids in the musts by HPLC–DAD
The amino acids determination was performed by the method described by Garde-Cerdán et al. [17]. Briefly, the derivatization of nitrogen compounds was performed by reaction of 750 μL of methanol (Merck, Darmstadt, Germany), 1.75 mL of borate buffer 1 M (pH 9), 1 mL of sample, 30 μL of diethyl ethoxymethylenemalonate (DEEMM) (Sigma-Aldrich, Madrid, Spain), as reagent of derivatization, and 20 μL of 2-aminoadipic acid (internal standard) (Sigma-Aldrich). The derivatization was carried out in a tube over 30 min in an ultrasound bath. Then, the sample was heated at 75 °C for 2 h to degrade the excess DEEMM and other by-products.
The analyses were performed on a Shimadzu Nexera X2 Ultra High-Performance Liquid Chromatograph (UHPLC) (Shimadzu, Kyoto, Japan) equipped with an automatic liquid sampler, and a diode array detector (DAD). Chromatographic separation was performed in an ACE HPLC column (C18-HL) (Aberdeen, Scotland) particle size 5 μm (250 mm × 4.6 mm). Amino acids were eluted under the conditions described by Garde-Cerdán et al. [18]. Phase A, 25 mM acetate buffer, pH 5.8, with 0.4 g of sodium azide; phase B, 80:20 (v/v) mixture of acetonitrile and methanol (Merck). DAD monitored at 280, 269, and 300 nm was used to detection. The volume of sample injected was 50 μL. The target compounds were identified according to the retention times and the UV–Vis spectral characteristics of corresponding standards (Sigma-Aldrich) derivatizated. Quantification was carried out using the calibration graphs of the respective standards in 0.1 N HCl, which underwent the same process of derivatization that the samples.
The treatments were performed in triplicate, so the results of free amino acids correspond to the average of three analyses (n = 3).
Statistical analysis
The statistical elaboration of the data was performed using SPSS Version 21.0 statistical package for Windows (SPSS, Chicago, USA). General parameters and nitrogen compound data were processed using the variance analysis (ANOVA) (p ≤ 0.05). Differences between samples were compared using the Duncan test at 95% probability level. Also, a multivariate factorial analysis (with treatment and vintage as factors) was performed considering enological parameters and nitrogen compounds in grapes. Discriminant analysis was performed to classify the different samples according to their nitrogen composition.
Results and discussion
General parameters in the musts
Table 1 shows the general enological parameters and nitrogen fractions in the control and in the samples from applications with free methyl jasmonate (MeJ) and MeJ loaded on ACP nanoparticles (ACP-MeJ), in 2019 and 2020 vintages. In the first vintage, there were no significant differences in the weight of 100 berries, while the control presented greater glucose and fructose content, which translated into higher ºBrix and probable degree, than the musts from the MeJ foliar application (Table 1). Wang et al. [19] found a reduction in ºBrix, glucose, and fructose after MeJ treatment in Gewürztraminer grape variety, pointing to an elicitor repressive effect on berries maturation. Must samples from ACP-MeJ treatment did not present significant differences with control and MeJ ones for these enological parameters. Regarding pH, total acidity, and malic acid, only total acidity was greater in the MeJ musts than in the control samples (Table 1). D'Onofrio et al. [20] observed that MeJ treatment diminished total acidity of Sangiovese grape variety. Regarding the nitrogen fractions, there was no difference between samples for ammonium nitrogen, while amino nitrogen and YAN were higher in the samples from the MeJ treatment than in the control one, and without significant differences with the ACP-MeJ samples (Table 1). In all cases, the samples had sufficient YAN content to avoid fermentation problems [21].
Nevertheless, in the second vintage, in any of the general parameters and nitrogen fractions of the musts were found significant differences (Table 1), in agreement with other works that have used this elicitor in free or conventional form in Tempranillo cultivar [14, 22, 23]. The vines on which the treatments were applied were the same in both vintages; thus, the different response in the general enological parameters and in the nitrogen fractions due to the vintage could be due to higher rainfall in August 2020, 32.9 L/m2, compared to 11.5 L/m2 in 2019.
Influence of the foliar MeJ and ACP-MeJ treatments on must amino acids’ content
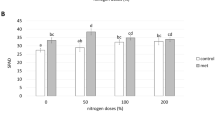
Table 2 shows the results of free amino acids content in control, MeJ and ACP-MeJ grapes, in both vintages, and Fig. 1 shows the concentration of total amino acids and total amino acids without proline in these samples.

Total amino acids and total amino acids without proline (mg/L) in musts from control, methyl jasmonate (MeJ), and nanoparticles doped with MeJ (ACP-MeJ) foliar treatments, in 2019 and 2020 vintages. All parameters listed with their standard deviation (n = 3). For each vintage and nitrogen parameter, different letters indicate significant differences between samples (p ≤ 0.05)
In the first vintage, 2019, the concentration in the musts of all the amino acids, with the exception of two of them, increased with the application of MeJ; while the treatment with ACP-MeJ had less influence on the amino acid composition, since only affected to six of the amino acids present in the samples, increasing their content (Table 1). Cysteine was the only amino acid whose content decreased with both treatments. These results were reflected in a higher concentration of total amino acids in the musts, from both treatments, than in the control, being the increase greater when applying the conventional MeJ (Fig. 1a); while the content of total amino acids without proline was only affected by applying free MeJ (Fig. 1b).
The results observed in the second vintage, 2020, were very different from those found in the first one, 2019. Several amino acids were found in the musts in lower concentration when applying MeJ than in the control samples, i.e., glutamine, histidine, glycine, threonine + citrulline, valine, isoleucine + tryptophan, leucine, and phenylalanine, and only one of them, lysine, increased its content with this treatment (Table 1); thus, the content of total amino acids and total amino acids without proline was lower after applying MeJ than in the control musts (Fig. 1a and b). However, when the foliar treatments were carried out with ACP-MeJ, practically, all the amino acids were found in similar concentrations as in the control, except for threonine + citrulline, valine, cysteine, leucine, and phenylalanine (Table 1), so the content of total amino acids and total amino acids without proline did not show significant differences with the control one (Fig. 1a and b).
The different response of the plant to the application of MeJ in both formats, free and nano, in each vintage could be due to several factors. First, the nitrogen needs of the plant, measured as nitrogen content in the berry, in 2019 were higher than in 2020, so that the plant responded more receptively in the first year of the trial, and it did in a more pronounced way when applying MeJ in a higher dosage, that is, in a conventional way than in nano-form (10 mM versus 1 mM) (Fig. 1). Second, MeJ is slowly released when applied in nano-form [4]; therefore, there could be a memory effect in the plants, so that in the second year, it behaved in a similar way to free MeJ (Fig. 1). This result was in agreement with Gil-Muñoz et al. [9], who also observed that, in the first year of application, the musts from the treatment with free MeJ had a higher content of amino acids than ACP-MeJ; while in the second year, the amino acid concentrations were lower, or similar, in ACP-MeJ than in MeJ. However, the application of free MeJ in the second year of our study was negative from the point of view of the must nitrogen composition respect to the control, which could be due, as indicated above, to the fact that the plants did not have the same nitrogen needs, since the content, in the control, of total amino acids in 2020 was practically double that in 2019 (Fig. 1).
The two most representative amino acids of grapes are arginine and proline, since they are two of the most abundant amino acids [24]. In addition, arginine is the best nitrogen source, after ammonium, for yeasts [17, 21], while proline is not metabolized by yeasts under typical vinification conditions, that is, in the absence of oxygen and in the presence of good nitrogen sources [25, 26]. Therefore, these two amino acids are related to assimilable and non-assimilable nitrogen, respectively, being the proline/arginine ratio a parameter that can indicate adequate or inadequate initial conditions, in terms of nitrogen available to yeasts during fermentation. Both treatments, MeJ and ACP-MeJ, decreased this ratio compared to the control in 2019, especially when applying free MeJ, while this parameter was not affected in 2020. Consequently, both treatments clearly improved the nitrogen available to the yeasts in 2019, which was the year with the lowest concentration of amino acids in the control must (Table 2 and Fig. 1).
Other important amino acids for being precursors of fermentative aromatic compounds, i.e., threonine, tyrosine, valine, methionine, isoleucine, tryptophan, leucine, and phenylalanine, that is, nitrogen compounds that determine the fermentative bouquet of wine and its organoleptic quality, were also affected by the treatments carried out in the vineyard, as previously mentioned (Table 2). Higher alcohols can be formed anabolically from sugars as well as catabolically from amino acids via the Ehrlich pathway [27]. In 2019, the content of alcohols in the control wines was higher than that of the wines from the MeJ and ACP-MeJ treatments [8], which was probably due to the fact that the sugar content was higher in the control must (Table 1), indicating that the main route of formation of these compounds was the anabolic pathway. Since there were no differences in sugar content between MeJ and ACP-MeJ must samples (Table 1), the differences, respect to the control wines, in the content of alcohols were higher in MeJ than in ACP-MeJ wines [8], probably due to the higher content of amino acids in the MeJ musts (Table 2). However, in 2020, as there were no differences in the sugar content between the control musts and those treated with MeJ and ACP-MeJ (Table 1), practically no differences were observed in the content of higher alcohols in the wines [8], and when there were differences, their content in MeJ wines was lower, since these musts had a lower content of several amino acids (Table 2). Given that several alcohols are precursors of acetate esters and the control wines had the highest alcohol content in 2019, since they had a greater amount of sugars (Table 1), what was said for higher alcohols corresponds to what was observed for esters [8].
Multivariable analysis
Table 3 shows the results of the factorial analysis (treatment, vintage, and their interaction) of the general parameters and nitrogen fractions of the musts. None of the parameters studied was affected by the treatment applied foliarly in the vineyard. Therefore, the vintage factor had the most impact on the overall parameters of the musts, so that, regardless of treatment, the weight of 100 berries and the ammonium nitrogen content were higher in 2020 than in 2019; while total acidity, fructose, and malic acid were higher in 2019 than in 2020 (Table 3). There was no interaction between the two factors for any of the general parameters studied, but did for nitrogen fractions.
Table 4 presents the results of the factorial analysis (treatment, vintage, and their interaction) of the must amino acids. The content in grapes of most of these compounds was affected by both factors. Regardless of vintage, the content of total amino acids and total amino acids without proline was higher in the samples treated with MeJ than in the control and in those from the treatments with ACP-MeJ, without significant differences between them (Table 4). This result was due to the fact that practically all the amino acids, with the exception of glutamic acid, asparagine, glutamine, GABA, valine, cysteine, and phenylalanine, in the case of the control, and aspartic acid, glutamic acid, asparagine, glutamine, arginine, GABA, and cysteine, in the case of ACP-MeJ, were in higher concentration in the MeJ musts (Table 4). It should be noted that Pérez-Álvarez et al. [5] also found no differences due to the treatment factor between the control and the samples from the treatment with ACP-MeJ. As for the effect of the vintage, regardless of the treatment, it was observed that all amino acids, except histidine, proline, methionine, and leucine, showed significant differences depending on the vintage, and therefore also in the total content of amino acids, with and without proline (Table 4). It should be noted that practically all of them, with the exception of aspartic acid, threonine + citrulline, and cysteine, had a higher concentration in 2020 than in 2019, being the total content of these nitrogen compounds higher in the second vintage (Table 4). Given that the vines used for this trial were the same in both vintages and that the vineyard was not fertilized, the differences between vintages could be due to the aforementioned, that is, to the fact that in August 2020, it rained more than in 2019, allowing a higher absorption by the plant of the nitrogen available in the soil. The main soil environmental factor affecting the nutrient flow is the soil water potential. A lack of water makes the soil water potential drop; therefore, the nitrogen moves slowly, and its absorption and transport are reduced [28]. There was interaction between the two factors for all amino acids, except glutamic acid (Table 4).
Figure 2 shows the discriminant analysis carried out with the amino acids concentration of the different samples in 2019 (Fig. 2a), in 2020 (Fig. 2b), and considering both vintages (Fig. 2c). In 2019 and 2020, it was observed that Function 1 (99.6% and 96.8%, respectively) allows a very good separation of the samples according to the treatment performed (control, MeJ, ACP-MeJ). In both vintages, control and ACP-MeJ are quite separated from MeJ (Fig. 2a and b), but with opposite behavior, more amino acids in 2019 and less in 2020 in the samples from the MeJ application, according to the results obtained (Table 4 and Fig. 1). If the global study is considered (all treatments and both vintages), it can be observed the existence of four differentiated groups, separated by both Functions (Function 1: 70.2%; Function 2: 22.9%). Function 1 separates the samples by vintage, the 2020 samples on the right and the 2019 samples on the left, due to the higher content of nitrogen compounds in the second vintage; while Function 2 separates them by treatments, in both vintages, the MeJ is clearly differentiated from the control and ACP-MeJ (Fig. 1c), according to their nitrogen composition.

Discriminant analysis of amino acids’ content (mg/L) in musts from control, methyl jasmonate (MeJ), and nanoparticles doped with this elicitor (ACP-MeJ) treatments, in a 2019, b 2020, and c 2019 and 2020 vintages
Conclusions
The effect of methyl jasmonate (MeJ) and methyl jasmonate-doped nanoparticle (ACP-MeJ) treatments on the nitrogen composition of Tempranillo grapes during two vintages were evaluated. The amino nitrogen and yeast assimilable nitrogen were higher in the grapes treated with MeJ and ACP-MeJ than in the control ones in 2019. However, in the second vintage, no significant differences were observed in any of the must general parameters and nitrogen fractions studied. With respect to the amino acids’ content, MeJ treatment enhanced the concentration of all of them, except from glutamic acid and γ-aminobutyric acid, in 2019. ACP-MeJ treatment only increased the concentration of six of them, prompting to a total amino acid concentration lower than MeJ treatment (applying ten times higher MeJ dosage), but higher than control sample. Nevertheless, the content of total amino acids and total amino acids without proline was lower after applying MeJ treatments (MeJ and ACP-MeJ) than in the control musts in 2020. The multivariable analysis revealed that all amino acids, except histidine, proline, methionine, and leucine, show significant differences depending on the vintage and regardless of the treatment. A prominent effect of the vintage on the overall parameters of must could be related to the higher rainfall in 2020, considering that MeJ is an elecitor able to trigger plant defense responses against abiotic stress (i.e., drought).
Data availability
All data included in this manuscript are available upon request by contacting with the corresponding authors.
References
Tilman D, Cassman KG, Matson PA, Naylor R, Polasky S (2002) Agricultural sustainability and intensive production practices. Nature 418:671–677. https://doi.org/10.1038/nature01014
Lowry GV, Avellan A, Gilbertson LM (2019) Opportunities and challenges for nanotechnology in the agri-tech revolution. Nat Nanotechnol 14:517–522. https://doi.org/10.1038/s41565-019-0461-7
An C, Sun C, Li N, Huang B, Jiang J, Shen Y, Wang C, Zhao X, Cui B, Wang C, Li X, Zhan S, Gao F, Zeng Z, Cui H, Wang Y (2022) Nanomaterials and nanotechnology for the delivery of agrochemicals: strategies towards sustainable agriculture. J Nanobiotechnol 20:11. https://doi.org/10.1186/s12951-021-01214-7
Parra-Torrejón B, Ramírez-Rodríguez GB, Giménez-Bañón MJ, Moreno-Olivares JD, Paladines-Quezada DF, Gil-Muñoz R, Delgado-López JM (2021) Nanoelicitors with prolonged retention and sustained release to produce beneficial compounds in wines. Environ Sci Nano 8:3524–3535. https://doi.org/10.1039/d1en00504a
Pérez-Álvarez EP, Rubio-Bretón P, Intrigliolo DS, Parra-Torrejón B, Ramírez-Rodríguez GB, Delgado-López JM, Garde-Cerdán T (2022) Year, watering regime and foliar methyl jasmonate doped nanoparticles treatments: effects on must nitrogen compounds in Monastrell grapes. Sci Hortic 297:110944. https://doi.org/10.1016/j.scienta.2022.110944
Garde-Cerdán T, Sáenz de Urturi I, Rubio-Bretón P, Marín-San Román S, Baroja E, Ramírez-Rodríguez GB, Delgado-López JM, Pérez-Álvarez EP (2023) Foliar application of methyl jasmonate and methyl jasmonate supported on nanoparticles: incidence on grape phenolic composition over two seasons. Food Chem 402:134244. https://doi.org/10.1016/j.foodchem.2022.134244
Giménez-Bañón MJ, Moreno-Olivares JD, Paladines-Quezada DF, Bleda-Sánchez JA, Fernández-Fernández JI, Parra-Torrejón B, Delgado-López JM, Gil-Muñoz R (2022) Effects of methyl jasmonate and nano-methyl jasmonate treatments on Monastrell wine volatile composition. Molecules 27:2878. https://doi.org/10.3390/molecules27092878
Pérez-Álvarez EP, Sáenz de Urturi I, Rubio-Bretón P, Marín-San Román S, Murillo-Peña R, Parra-Torrejón B, Ramírez-Rodríguez GB, Delgado-López JM, Garde-Cerdán T (2022) Application of elicitors, as conventional and nano forms, in viticulture: effects on phenolic, aromatic and nitrogen composition of Tempranillo wines. Beverages 8:56. https://doi.org/10.3390/beverages8030056
Gil-Muñoz R, Giménez-Bañón MJ, Moreno-Olivares JD, Paladines-Quezada DF, Bleda-Sánchez JA, Fernández-Fernández JI, Parra-Torrejón B, Ramírez-Rodríguez GB, Delgado-López JM (2021) Effect of methyl jasmonate doped nanoparticles on nitrogen composition of Monastrell grapes and wines. Biomolecules 11:1631. https://doi.org/10.3390/biom11111631
Martínez-Moreno R, Quirós M, Morales P, Gonzalez R (2014) New insights into the advantages of ammonium as a winemaking nutrient. Int J Food Microbiol 177:128–135. https://doi.org/10.1016/j.ijfoodmicro.2014.02.020
Bouzas-Cid Y, Trigo-Córdoba E, Falqué E, Orriols I, Mirás-Avalos JM (2018) Influence of supplementary irrigation on the amino acid and volatile composition of Godello wines from the Ribeiro Designation of Origin. Food Res Int 111:715–723. https://doi.org/10.1016/j.foodres.2018.05.074
Garde-Cerdán T, Ancín-Azpilicueta C (2008) Effect of the addition of different quantities of amino acids to nitrogen-deficient must on the formation of esters, alcohols, and acids during wine alcoholic fermentation. LWT Food Sci Technol 41:501–510. https://doi.org/10.1016/j.lwt.2007.03.018
Lytra G, Miot-Sertier C, Moine V, Coulon J, Barbe J-C (2020) Influence of must yeast-assimilable nitrogen content on fruity aroma variation during malolactic fermentation in red wine. Food Res Int 135:109294. https://doi.org/10.1016/j.foodres.2020.109294
Garde-Cerdán T, Portu J, López R, Santamaría P (2016) Effect of methyl jasmonate application to grapevine leaves on grape amino acid content. Food Chem 203:536–539. https://doi.org/10.1016/j.foodchem.2016.02.049
Garde-Cerdán T, Gutiérrez-Gamboa G, Baroja E, Rubio-Bretón P, Pérez-Álvarez EP (2018) Influence of methyl jasmonate foliar application to vineyard on grape volatile composition over three consecutive vintages. Food Res Int 112:274–283. https://doi.org/10.1016/j.foodres.2018.06.048
OIV (2009) Compendium of internationals methods of wine and must analysis. OIV, Paris
Garde-Cerdán T, Martínez-Gil AM, Lorenzo C, Lara JF, Pardo F, Salinas MR (2011) Implications of nitrogen compounds during alcoholic fermentation from some grape varieties at different maturation stages and cultivation systems. Food Chem 124:106–116. https://doi.org/10.1016/j.foodchem.2010.05.112
Garde-Cerdán T, Lorenzo C, Lara JF, Pardo F, Ancín-Azpilicueta C, Salinas MR (2009) Study of the evolution of nitrogen compounds during grape ripening. Application to differentiate grape varieties and cultivated systems. J Agric Food Chem 57:2410–2419. https://doi.org/10.1021/jf8037049
Wang J, VanderWeide J, Yan Y, Tindjau R, Pico J, Deluc L, Zandberg WF, Castellarin SD (2022) Impact of hormone applications on ripening-related metabolites in Gewürztraminer grapes (Vitis vinifera L.): the key role of jasmonates in terpene modulation. Food Chem 388:132948. https://doi.org/10.1016/j.foodchem.2022.132948
D’Onofrio C, Matarese F, Cuzzola A (2018) Effect of methyl jasmonate on the aroma of Sangiovese grapes and wines. Food Chem 242:352–361. https://doi.org/10.1016/j.foodchem.2017.09.084
Bell S-J, Henschke PA (2005) Implications of nitrogen nutrition for grapes, fermentation and wine. Aust J Grape Wine Res 11:242–295. https://doi.org/10.1111/j.1755-0238.2005.tb00028.x
Portu J, Santamaría P, López-Alfaro I, López R, Garde-Cerdán T (2015) Methyl jasmonate foliar application to Tempranillo vineyard improved grape and wine phenolic content. J Agric Food Chem 63:2328–2337. https://doi.org/10.1021/jf5060672
Portu J, López R, Baroja E, Santamaría P, Garde-Cerdán T (2016) Improvement of grape and wine phenolic content by foliar application to grapevine of three different elicitors: methyl jasmonate, chitosan, and yeast extract. Food Chem 201:213–221. https://doi.org/10.1016/j.foodchem.2016.01.086
Stines AP, Grubb J, Gockowiak H, Henschke PA, Høj PB, Van Heeswijck R (2000) Proline and arginine accumulation in developing berries of Vitis vinifera L. in Australian vineyards: Influence of vine cultivar, berry maturity and tissue type. Aust J Grape Wine Res 6:150–158. https://doi.org/10.1111/j.1755-0238.2000.tb00174.x
Arias-Gil M, Garde-Cerdán T, Ancín-Azpilicueta C (2007) Influence of addition of ammonium and different amino acid concentrations on nitrogen metabolism in spontaneous must fermentation. Food Chem 103:1312–1318. https://doi.org/10.1016/j.foodchem.2006.10.037
Pinu FR, Edwards PJB, Gardner RC, Villas-Boas SG (2014) Nitrogen and carbon assimilation by Saccharomyces cerevisiae during Sauvignon blanc juice fermentation. FEMS Yeast Res 14:1206–1222. https://doi.org/10.1111/1567-1364.12222
Ayräpää T (1971) Biosynthetic formation of higher alcohols by yeasts. Dependence on the nitrogenous nutrient level of the medium. J Inst Brew 77:266–275. https://doi.org/10.1002/j.2050-0416.1971.tb06945.x
Gong P, Zhang Y, Liu H (2022) Effects of irrigation and N fertilization on 15N fertilizer utilization by Vitis vinifera L. Cabernet Sauvignon in China. Water 14:1205. https://doi.org/10.3390/w14081205
Acknowledgements
S. M.-S.-R. and R. M.-P. thank Gobierno de La Rioja and INIA, respectively, for their predoctoral contracts. G.B. R.-R. also acknowledges Junta de Andalucía for her postdoctoral contract within the PAIDI 2020 program (DOC_01383). E.P. P.-Á. thanks the Ministerio de Ciencia, Innovación y Universidades for her Juan de la Cierva-Incorporación contract.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This work has been carried out thanks to funding from the Ministerio de Ciencia, Innovación y Universidades under Project Nos. RTI2018-096549-B-I00 and RTI-2018-095794-A-C22.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Compliance with ethics requeriments
This article does not contain any studies with human or animal subjects.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Garde-Cerdán, T., Sáenz de Urturi, I., Marín-San Román, S. et al. Influence of foliar treatments with methyl jasmonate and methyl jasmonate-doped nanoparticles on nitrogen composition of Tempranillo grapes during two vintages. Eur Food Res Technol 249, 1187–1197 (2023). https://doi.org/10.1007/s00217-023-04206-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00217-023-04206-z