
Abstract
There is a great scientific curiosity to discover all environments sheltering microalgae, especially those with exceptional characteristics from coldest to hottest ones, the purpose remains to explore the potential of the native microalgae flora and the research for new bioactive compounds. This study aimed to isolate a polysaccharide-producing microalga from an extreme ecosystem and to evaluate its capacity to inhibit the α-d-glucosidase enzyme. Chlorella strain is isolated from hypersaline Lake in the Algerian desert. The exopolysaccharide extraction was performed by the concentration of free-cell supernatant in a rotary evaporator. The infrared analysis showed a characteristic footprint of carbohydrates with particular functional groups, such as sulfate. Gas chromatography–mass spectrometry has revealed a hetero-exopolysaccharide composed of galactose 35.75%, glucose 21.13%, xylose 16.81%, fructose 6.96%, arabinose 5.10%, and glucuronic acid 2.68%. The evaluation of the anti-hyperglycemic activity demonstrated a significant α-d-glucosidase inhibition of 80.94 ± 0.01% at 10 mg mL−1 with IC50 equal to 4.31 ± 0.20 mg mL−1. This study opens a vast prospect to use exopolysaccharides as natural nutraceutical or food additive.
Graphical Abstract

Similar content being viewed by others

Introduction
The scientific community continues exploring microalgae molecules from new environments, aiming to find bioactive compounds with interesting properties to use in biotechnology. Microalgae are photosynthetic organisms known for their extraordinary diversity, and natural origin, making them a good choice for a variety of applications (Colusse et al. 2022).
Green microalgae have been receiving increased attention since they present the most diverse group of microalgae. Among Chlorophycean genera; Chlorella is a small, single-celled, green, eukaryotic microalga, with a size around 2–15 μm in diameter. These unicellular green microalgae are present in all water habitats (fresh and marine water), exhibiting a cosmopolitan occurrence (Wong et al. 2015). Despite being a simple alga, Chlorella is a rich source of a variety of molecules implicated in a large spectrum of innovative applications such as bioremediation, biofuels, food, feed, biopolymers, and aquaculture (Abreu et al. 2023).
Chlorella is a potent microalga that possesses excellent potential, some species have been approved for human consumption by the European Food Safety Authority (No 258/97)Footnote 1 (Niccolai et al. 2019). Its supplementation in mammals, including humans, has been reported to exhibit various pharmacological activities (Neumann et al. 2018; Ramos-Romero et al. 2021). Therefore, Chlorella species have been added to different commercial food products as dietary supplements. It has been incorporated as whole biomass to ameliorate the techno-functional and nutritional properties in pasta, yogurt, cheese, cookies, and water/oil emulsions (Caporgno and Mathys 2018).
Chlorella is among the few eukaryotic microalgae that synthesize substances with a broad spectrum of antibacterial/antifungal activity (Mashhadinejad et al. 2016; Dinev et al. 2021; Shaima et al. 2022), Chlorella marina produces lycopene (Mtaki et al. 2020), Chlorella stigmatophora produces polysaccharides (Sathasivam et al. 2019), Chlorella pyrenoidosa produces flavonoids (Yadavalli et al. 2022), Chlorella protothecoides produces sporopollenin (He et al. 2016), Chlorella fusca produces sporopollenin (Priyadarshani and Rath 2012). Furthermore, Chlorella produces a plethora of biomolecules with relevant properties for human health and food industry, such as polysaccharides (PSs). PSs are considered the primary active component of Chlorella owning diverse health-promoting potentials and food functional qualities, like those associated with antioxidation, anticoagulation, immunomodulation, anti-hyperlipidemia, antitumor, neuroprotection, and anti-asthmatic effect (Sheng et al. 2007; Chen et al. 2014, 2016; Barboríková et al. 2019; Wan et al. 2020; Mousavian et al. 2022; Tang et al. 2023).
Studies on the anti-diabetic activity of PSs derived from Chlorella are rare, most work done concerns the supply of Chlorella as whole biomass in rats and the evaluation of its activity (Jong-Yuh and Mei-Fen 2005; Senthilkumar and Ashokkumar 2012; Yuan et al. 2020; Ramos-Romero et al. 2021; Xiong et al. 2022). Qiu et al. (2022) have described the anti-hyperglycemic, antioxidant, and anti-inflammatory potential of PSs extracted from Chlorella pyrenoidosa in aging-related diabetes mice model. The suggested findings demonstrated the inhibition of high glucose levels through induction of insulin secretion, and the prevention of oxidation by improving the secretion of antioxidant enzymes (Qiu et al. 2022). The mechanisms by which Chlorella may protect humans from diabetes and related risk factors are mainly unclear, the common explanation given is that the protective effect against diabetes by inducing insulin secretion, but also by increasing the circulating glucose uptake in the liver and muscles (Bito et al. 2020; Ramos-Romero et al. 2021).
Most studies have used Chlorella as whole biomass in animal models for the assessment of the biological activities, thus, a synergetic effect between all compositional compounds occurs in the observed potential, and the main molecule responsible for the activity is not defined. Herein, there is a serious need to explore the bioactivity of each single metabolite. Therefore, this paper is a contribution to the evaluation of the antihyperglycemic activity through the assessment of α-d-glucosidase enzyme inhibition by exopolysaccharide (EPS) derived from a wild Chlorella sp. isolated from a hypersaline Lake (Chott Ain El-Beida), Ouargla-Algeria. The Lake is of international importance since it has been classified as an internationally protected wetland according to RAMSAR Convention 1971. The native microalgae flora in Algeria is still under-investigated, this is the first report on microalgae’s EPS derived from a local strain. To the best of our knowledge, no papers were reported on the evaluation of α-d-glucosidase inhibition by exopolysaccharide-producing Chlorella sp. from Saharan biotopes.
Materials and methods
Measurement of physicochemical parameters
A water sample was taken from Chott Aïn El-Beida lake Ouargla-Algeria (31°57′45.3″N 5°22′30.7″E) January 29th 2019, and the physicochemical parameters of water were measured using a multi-parameter analyzer; the parameters were: the hydrogen potential (pH 0.00–14.00), the temperature (°C), the salinity (g L−1), the conductivity (m S−1 cm−1), the dissolved oxygen (ppm) and the total dissolved solids (ppt) (HANNA HI 9828).
Isolation and microscopic observation
The isolation of microalgae was accomplished by transferring the water sample into a solid Bold’s Basal media (BBM), at 1.5% of agar; macronutrients (g L−1 dH2O) NaNO3 25, CaCl2.2H2O 2.5, MgSO4.7H2O 7.5, K2HPO4 7.5, KH2PO4 17.5, NaCl 2.5, EDTA-alkaline solution EDTA 50, KOH 31, Acidified Iron Solution FeSO4.7H2O 4.98, H2SO4 1 mL, Boron Solution H3BO3 11.42, Trace Metal Solution ZnSO4.7H2O 8.82, MnCl2.4H2O 1.44, MoO3 0.71, CuSO4 5H2O 1.57, Co(NO3)2 6H2O 0.49 with a final pH of 6.6 ± 0.2. Incubation of Petri dishes was done at 25 °C ± 02, 16 h–8 h light–dark photoperiod cycle and irradiance of 100 µmol photons m−1 s−1 during 2 weeks. After the apparition of colonies, every single colony was purified on BBM agar plate until getting axenic culture. The bacterial contamination were prevented using streptomycine sulfate within the agar plate (Raja et al. 2007). The morphological features were observed using the optical microscope (S/N-EU 1900816, euromex, BioBlue Lab), and the scanning electron microscope (SEM) Quattro ESEM, Thermo Fisher Scientific–US, with a field emission gun.
Growth conditions and exopolysaccharide extraction
The cultivation was carried out in BBM medium at 25 °C ± 02, under a white fluorescent light provided in 16 h–8 h light–dark photoperiod cycle, with an intensity of 150 µmol photons m−1 s−1. After cultivation, the culture were centrifuged at 4000 g for 30 min, the resulted free cell supernatant was concentrated by rotary evaporator (Heidolph Hei-VAP, Germany), at 45 °C for 100 ppm to about 1/4 of its initial volume, then, precipitated with cold ethanol (1:4 w/v) overnight at − 20 °C. Another centrifugation was done at 6000 g for 10 min, the pellet was then collected, lyophilized (Alpha2-4 LSCbasic freeze dryer) and stored at 4 °C.
Exopolysaccharide characterization
Biochemical analysis
The phenol–sulfuric acid technique was used to determine the total sugar content (TSC). TSC was calculated from the glucose calibration curve (Dubois et al. 1956). The protein content was determined using the Bradford assay with some modifications as reported in Berges et al. (1993); 1 mL (instead of 0.1 mL) of the standard, and 1 mL (instead of 3 mL) of Bradford reagent, then calculated from the BSA calibration curve. The modified Folin-Ciocalteu technique was also used to determine the total phenolic content (TPC). The gallic acid calibration curve was used to calculate the TPC (Bradford 1976; Berges et al. 1993; Zakaria et al. 2017; Kaliwal 2019).
Scanning electron microscopy coupled with energy dispersive X-ray analysis
The lyophilized powder of the crude extract was analyzed using a SEM (SEM, SIGMA 500/VP, ZEISS) at a voltage of 20 kV. The EPS is a non-conductor material, thus it was deposited onto carbon tape and gold-sputtered prior to analysis. The elemental composition on the EPS surface was determined using the Energy Dispersive X-Ray Analysis (EDX) technique (Fimbres-Olivarría et al. 2016; Olasehinde et al. 2019).
Fourier transform infra-red analysis
Fourier transform infrared (FT-IR) analysis was employed in the transmission mode at a range of 400–4000 cm−1, using a Perkin Elmer spectrometer at a resolution of 8 cm−1. The sample was compressed in a cold 150 Mpa isostatic press (CIP) with 23 ± 2 mg of KBr, to attain a 200–250 μm thick pellet (Ferreira et al. 2020).
Gas chromatography mass spectrometry
The monosaccharide composition was determined using gas chromatography mass spectrometry (GC–MS) ‘Agilent 6890 Series GC System coupled to an Agilent 5973 Network, Shimadzu, GCMS-QP2020 NX’. GC–MS is a widely applied technique in the identification of polysaccharide composition due to high accuracy and repeatability (Zhang and Zhou 2017). Briefly, 10 mg of crude EPS were hydrolyzed using 1 mL of trifluoroacetic acid 2 M at 120 °C for 1 h 30 min. The liquid was then evaporated at 60 °C by nitrogen stream. Then, the hydrolysate was subjected to derivatization using (BSTFA/TMCS; 99%/1%), under Argon gas, the mixture was incubated at room temperature for 2 h to make trimethylsilyl-O-glycosides, which were then solubilized in dichloromethane after the reagent was evaporated. Next, the obtained derivation was injected to the separation column OPTIMA 1MS (30 m × 0.32 mm id, 0.25 μm) with a helium flow rate of 2.3 mL/min. Firstly, temperature was at 100 °C for 3 min. Secondly, an increment of 8 °C/min up to 200 °C for 1 min was used before a final increment of 5 °C/min up to 250 °C. The electronic impact (EI, 70 eV) ionization method was applied with the trap temperature set at 150 °C and the target ion was fixed at 40–800 m/z. Twelve mono-sugars were used as standards (Arabinose Ara, Rhamnose Rha, Galactose (Gal), Glucose (Glc), Galacturonic acid (GalA), Xylose (Xyl), Fucose (Fuc), Mannose (Man), Glucuronic acid (GlcA), Ribose (Rib), Glucosamine (GlcN) and Galactosamine (GalN) (Sigma-Aldrich).
In vitro assessment of anti-α-d-glucosidase activity
The test measures the in vitro inhibition potential of the α-d-Glucosidase enzyme by the crude EPS, it is based on the measurement of the released ρ-nitrophenyl-α-d-glucopyranoside (ρ-NPG). Briefly, 0.5 mL of the α-d-glucosidase enzyme 0.2 U mL−1 (Sigma-Aldrich) was added to 0.1 mL of the EPS extract–previously suspended in H2O– and was incubated at 37 °C for 15 min under gentle stirring. Next, 0.125 mL (4 mM) of ρ-NPG was added, then, incubated at 37 °C for 20 min. Afterward, 1 mL of Na2CO3 (0.2 M) was supplied to stop the reaction. Finally, the absorbance was read at λ = 405 nm using a spectrophotometer (6850 UV/Vis, JENWAY) (Bisht et al. 2013; Qian et al. 2015). The acarbose (Sigma-Aldrich) was used as a positive control. The percentage of inhibition was calculated as follows:
ABS: absorbance.
Statistical analysis
Every measurement was made in triplicate, and results were expressed as means ± SD. The linear regression was used to determine the IC50 of the α-d-Glucosidase inhibitory effect of EPS extract using XLSTAT 2022 (trial version, Add in soft Inc., Boston, MA, USA).
Results and discussions
Physicochemical parameters measurement of Lake water
The measured parameters on Lake water of Chott Ain El-Beida are: potential of hydrogen (pH), temperature (°C), salinity (g L−1), electrical conductivity (mS−1 cm−1), dissolved oxygen (ppm) and total dissolved solids (ppt). These parameters give information on water characteristics and the conditions surrounding the microalgae flora in this ecosystem. The main observation concerns the high salinity (51.46 g L−1) and conductivity (74.70 mS−1 cm−1) of water (Table 1).
Water physicochemical parameters provide some information on the microalgal native flora of the marine community, and aiding in the interpretation of different algal classes occurrence in their environment. The degree of salinity (Sn) makes it possible to classify water according to 4 categories: freshwater (Sn < 0.5 g L−1), freshwater to brackish (0.5 to 5 g L−1), brackish to salty water (18 to 30 g L−1) and salt water (Sn > 30 g L−1) (Hecker et al. 1996). Therefore, the water of Chott Aïn El-Beida Lake water is defined as hypersaline water. Saharan environments have been investigated as having little biological interest and limited biodiversity. However, this notion has changed recently, research has shown that arid habitats are home to unique species which adapt to the harshest ecosystems by modulating their physiology to acquire more resistance capacities (Guezoul et al. 2013).
Microscopic observation and biochemical analysis
A strain was isolated from Chott Aïn El-Beida Lake, situated in the Algerian desert. The microscopic observations and morphological features revealed the small green cells ranging between 2–10 µm in diameter that belong to Chlorella sp. (Fig. 1).

(a) Chlorella sp., (S1) cells observed under a light microscope × 100, (b) Chlorella sp. (S1) cells observed under a scanning electron microscope
The biochemical analysis demonstrates different fractions of the extract (Table 2).
It is known that each fraction of the microalgae’s cell differs depending on the strain type and their physiological reactions to biotic and abiotic parameters (Barkia et al. 2019). In this study, the total sugar, the total protein, and the polyphenols amounts of the crude EPS were close to those obtained in other studies (Chen et al. 2016; Trabelsi et al. 2016; Kamble et al. 2018; Yu et al. 2019; Casas-arrojo et al. 2021), who described a total carbohydrate fraction of 48.53%, 57.53%, 52%, 52.74%, and 78%, and a total protein of 15.96%, 8.29%, 12%, 0.75%, and 2% from Porphyridium cruentum, Chlorella vulgaris, Chlorella pyrenoidosa, Graesiella sp., and Chlamydomonas reinhardtii, respectively.
Exopolysaccharide characterization
The extracted EPS consisted of a white-colored powder, with fine and granulated parts, the SEM micrographs showed an amorphous solid state, and granule forms, with compact and porous structure of irregular shape (Fig. 2a, b). The EDX analysis confirms the presence of sulfur (Fig. 2c). Sulfur is a common element in microalgal EPS, it is implicated in the functional groups, and contributes to the negative charges of these biomolecules. It was reported that the EPS produced by 120 marine microalgae were mostly sulfated (Raposo et al. 2014; Liu et al. 2016).

(a and b) Micrographs of the lyophilized EPS powder derived from Chlorella sp., obtained by SEM at different magnifications (A × 2.5k, B × 5k), (c) EDX showing different constitutive elements on the surface of the crude EPS
Sulfate quantity and its position in the EPS affects positively its bioactivity. Among the possible explanations is that the presence of a sulfate group in the EPS activates the hydrogen on the anomeric carbon, which increases the EPS’s hydrogen supply capacity and hence its potentiality. Also, a high sulfate content can increase the polysaccharide’s water solubility and improves its biological activity (Zhong et al. 2019). Sometimes an abnormally high sulfate content may affect the structure; the increasing in the degree of sulfation disrupts the EPS’s triple-helical structure, thus, reducing its capacity to supply hydrogen, resulting in less potentiality of the EPS (Liu et al. 2018).
In contrast to polysaccharides obtained from various organisms, microlgae-derived polysaccharides, referred to as sulfated polysaccharides due to the presence of sulfate esters, they exhibit diverse biological properties such as anticoagulant, anti-tumor, antidiabetic, anti-microbial, anti-radiation, anti-inflammatory effects… These sulfated polysaccharides have demonstrated efficacy in preventing the accumulation of free radicals and reactive chemical species, thereby serving as a protective mechanism against oxidative and radical stress agents (Arunkumar et al. 2021, 2023). The effectiveness of these polysaccharides is intricately tied to factors such as their degree and position of sulfation, sugar composition, molecular weight, and the treatment process (Delattre et al. 2016; Guehaz et al. 2024). Sulfate groups are incorporated into the sugar structure’s backbone to withstand harsh marine conditions such as high salinity, resulting in modifications to their polymeric structure, transforming into sulfated polysaccharides that exhibit significant biological activity and find extensive commercial applications (Muthukumar et al. 2021). The metabolism of the synthesis of the polysaccharide inside the microalgal cells, which enzymes are implicated, and how enzymes participate in the maturation of the final structure of the polysaccharides is yet to be clear. These mechanisms have a crucial relation to the phylogenic origin, however, herein further research is needed to elucidate the sulfation step in polysaccharides derived from microalgae, and how the degree and the position of sulfation is controlled.
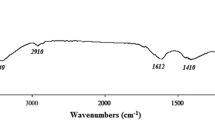
The FT-IR revealed many peaks ranging from 3466 to 518 cm−1 (Fig. 3). The broad intense absorption peak around 3466 cm−1 and 2920 cm−1 characterizes the stretching vibration of O–H and C-H in the sugar residues, respectively (Sun et al. 2014; Song et al. 2018). The bands around 1635 cm−1 and 1358 cm−1 were possibly attributed to the carboxyl groups due to the presence of glucuronic acid (Xia et al. 2014). The absorption observed at 1168 cm−1 and 1074 cm−1 may be due to C–O–C stretching of glycosidic bands, and the presence of C-O–H side groups of the pyrane ring in the extracts, respectively (Wang et al. 2014; Chokshi et al. 2016). The weak band at 1263 cm−1 and the band at 860 cm−1 are assigned to the S = O and C-O-S stretching vibrations of the sulfate groups, respectively (Abd El Baky et al. 2014; Fimbres-Olivarría et al. 2016; Yuan et al. 2020). Bands at 984 cm−1 and 950 cm−1 were generally attributed to C–C and C-O stretching vibrations in pyranose rings and indicating the presence of polysaccharides as the major component (Malinowska et al. 2018).

FT-IR spectrum of crude EPS extracted from Chlorella sp
Various methods were used for the characterization of the polysaccharides derived from microalgae and cyanobacteria, however, GC–MS remain the best method of choice (Delattre et al. 2016; Mehta and Shah 2021). The GC–MS technique revealed a heteropolysaccharide composed mainly of galactose 35.75%, glucose 21.13%, xylose 16.81%, rhamnose 11.57%, (Table 3). Jakhu et al. (2021) have reported similar results concerning the dominant sugars; galactose 33.43%, glucose 22.10%, and xylose11.86% when the product was derived from Chlorella sp. strain (Jakhu et al. 2021). According to Capek et al. 2020; galactose was the dominant hexose in the EPS isolated from Chlorella vulgaris with 37.9% (Capek et al. 2020). Also, galactose is the main constituent in the EPS of Dictyosphaerium chlorelloides with 42% (Halaj et al. 2022). Koçer et al. (2021) have studied the composition of the EPS of Chlorella minutissima and Chlorella sorokiniana; again, the main sugars were galactose 51.28%, 45.89% and glucose 25.07%, 28.68% respectively, the EPS from both strains contained a weak fucose content 0.12% and 1.33% respectively (Koçer et al. 2021). Kokarakis et al. (2022) reported different EPS composition from Chlorella sp.; glucosamine 23.4%, galactosamine 17.8%, rhamnose 25.5%, Arabinose 10.3%, glucose 8.9%, xylose 5.5% and mannose 2.7% (Kokarakis et al. 2022).
In vitro assessment of anti-α-d-glucosidase activity
The EPS extract was assessed for the in vitro inhibition of the α-d-Glucosidase enzyme, the results present the inhibition percentages of both acarbose and crude EPS at different dilutions (Fig. 4). The EPS inhibits α-d-Glucosidase enzyme in dose-dependent manner. The inhibition increases from 6.2% at 0.25 mg mL−1 to reach 80.94% at maximal concentration of 10 mg mL−1.

Inhibition of α-d-Glucosidase enzyme by crude EPS derived from Chlorella sp
The half-maximal inhibitory concentration (IC50) was determined and made with that of acarbose (Table 4). The IC50 equals to 4.31 ± 0.20 mg mL−1 compared 0.39 ± 0.04 mg mL−1 of acarbose.
Our findings are close to those made on the methanolic extract of Arthrospira platensis, that reported a strong α-glucosidase inhibitory activity of 97.42% and an IC50 value of 9.56 mg mL−1 (Gheda et al. 2021). Likewise, ethyl acetate extracted from Nannochloropsis oculata exhibited a significant inhibitory action on the α-glucosidase enzyme by 80.42% at the concentration of 1 g mL−1 with an IC50 value of 178.53 μg mL−1 (Deepa et al. 2022).
Moreover, Priatni et al. (2016) have assessed the α-d-glucosidase inhibition performed by EPS from 10 different Indonesian cyanobacterial producers. Although most of the EPSs exhibited inhibitory activity, there values were lower compared to the results from the current study. The highest inhibitory levels were exhibited by EPSs from Pseudanabaena sp. (14.02%) and Chroococcus sp. (13.0%) isolates, while products from Phormidium sp. and Oscillatoria limnetica did not exhibit any inhibitory activity. Additionally, Qi and Kim (2017) have investigated the α-d-glucosidase activity of carotenoids derived from Chlorella ellipsoidea. The extracts displayed potent inhibitory effect against α-d-glucosidase with a non-competitive inhibition (Qi and Kim 2017). In addition, fucoxanthin from Phaeodactylum tricornutum exerted a weak inhibitory activity against rat-intestinal α-glucosidase 32.18% at a concentration of 1 g mL−1 in a dose-dependent manner with an IC50 value of 4.75 µg mL−1 (Arthitaya et al. 2019).
Conclusion
While the utilization of microalgae or their byproducts as food alternatives is not yet competitive, this research underscores the importance of exploring harsh environments that may harbor native strains producing bioactive compounds crucial for developing nutritious and health-promoting foods, especially in the context of diabetes.
This preliminary investigation presents findings on the extraction of an EPS derived from Chlorella sp. strain found in the hypersaline Chott Aïn El-Beida Lake within the Algerian Sahara. The study highlights the notable potential of this EPS as an anti-hyperglycemic agent. The composition of the EPS includes galactose, glucose, xylose, rhamnose, fucose, arabinose, and glucuronic acid, accompanied by sulfate, methyl, and carboxyl functional groups. The observed efficacy is attributed to the sulfate and glucuronic acid content.
Although this study contributes valuable insights into the anti-hyperglycemic activity of Chlorella sp.-derived EPS, further research is imperative to fully characterize the EPS’s structure and optimize growth conditions. Additionally, assessing the EPS’s potential through in vivo investigations using animal models, exploring additional parameters, and elucidating the mechanistic processes involved in diabetes modulation will be worthwhile.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Notes
(European Union, Novel Food catalogue & No 2015/2283, EFSA).
References
Abd El Baky H, Hanaa El Baz KF, El-Latife SA (2014) Induction of sulfated polysaccharides in Spirulina platensis as response to nitrogen concentration and its biological evaluation. J Aquac Res Dev 5:1–8. https://doi.org/10.4172/2155-9546.1000206
Abreu AP, Martins R, Nunes J (2023) Emerging applications of Chlorella sp. and Spirulina (Arthrospira) sp. Bioengineering 10:1–34. https://doi.org/10.3390/bioengineering10080955
Arthitaya K, Taehwan KIMA, Moo KIMS (2019) Inhibitory activities of microalgal fucoxanthin against α-amylase, α-glucosidase, and glucose oxidase in 3T3-L1 cells linked to type 2 diabetes. J Oceanol Limnol 37:928–937
Arunkumar K, Raja R, Kumar VBS et al (2021) Antioxidant and cytotoxic activities of sulfated polysaccharides from five different edible seaweeds. J Food Meas Charact 15:567–576. https://doi.org/10.1007/s11694-020-00661-4
Arunkumar K, Sreena KS, Moosa M et al (2023) Cytotoxic characterization of optically negative Codium fragile polysaccharide against HeLa and MCF cell lines. Bioact Carbohydr Diet Fibre 29:100341. https://doi.org/10.1016/j.bcdf.2022.100341
Barboríková J, Šutovská M, Kazimierová I et al (2019) Extracellular polysaccharide produced by Chlorella vulgaris—chemical characterization and anti-asthmatic profile. Int J Biol Macromol 135:1–11. https://doi.org/10.1016/j.ijbiomac.2019.05.104
Barkia I, Saari N, Manning SR (2019) Microalgae for high-value products towards human health and nutrition. Mar Drugs 17:1–29. https://doi.org/10.3390/md17050304
Berges JA, Fisher AE, Harrison PJ (1993) A comparison of Lowry, Bradford and Smith protein assays using different protein standards and protein isolated from the marine diatom Thalassiosira pseudonana. Mar Biol 115:187–193. https://doi.org/10.1007/BF00346334
Bisht S, Kant R, Kumar V (2013) International Journal of Biological Macromolecules α-D-Glucosidase inhibitory activity of polysaccharide isolated from Acacia tortilis gum exudate. Int J Biol Macromol 59:214–220. https://doi.org/10.1016/j.ijbiomac.2013.04.057
Bito T, Okumura E, Fujishima M, Watanabe F (2020) Potential of Chlorella as a dietary supplement to promote human health. Nutrients 12:1–21. https://doi.org/10.3390/nu12092524
Bradford MM (1976) A rapid and sensitive method for the quantitation microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Capek P, Matulová M, Šutovská M et al (2020) Chlorella vulgaris α-L-arabino-α-L-rhamno-α, β-D-galactan structure and mechanisms of its anti-inflammatory and anti-remodelling effects. Int J Biol Macromol 162:188–198. https://doi.org/10.1016/j.ijbiomac.2020.06.151
Caporgno MP, Mathys A (2018) Trends in microalgae incorporation into innovative food products with potential health benefits. Front Nutr 5:1–10. https://doi.org/10.3389/fnut.2018.00058
Casas-arrojo V, Decara J, Arrojo-Agudo MLA et al (2021) Immunomodulatory, antioxidant activity and cytotoxic effect of sulfated polysaccharides from Porphyridium cruentum. Biomolecules 11(4):488
Chen PB, Wang HC, Liu YW et al (2014) Immunomodulatory activities of polysaccharides from Chlorella pyrenoidosa in a mouse model of Parkinson’s disease. J Funct Foods 11:103–113. https://doi.org/10.1016/j.jff.2014.08.019
Chen YX, Liu XY, Xiao Z et al (2016) Antioxidant activities of polysaccharides obtained from Chlorella pyrenoidosa via different ethanol concentrations. Int J Biol Macromol 91:505–509. https://doi.org/10.1016/j.ijbiomac.2016.05.086
Chokshi K, Pancha I, Ghosh T et al (2016) Green synthesis, characterization and antioxidant potential of silver nanoparticles biosynthesized from de-oiled biomass of thermotolerant oleaginous microalgae: Acutodesmus dimorphus. RSC Adv 6:72269–72274. https://doi.org/10.1039/c6ra15322d
Colusse GA, Carneiro J, Duarte MER et al (2022) Advances in microalgal cell wall polysaccharides: a review focused on structure, production, and biological application. Crit Rev Biotechnol 42:562–577. https://doi.org/10.1080/07388551.2021.1941750
Deepa PK, Subramanian A, Manjusha WA (2022) Phytochemical screening and evaluation of antidiabetic activity of the marine microalgae: Nannochloropsis sp. Int J Life Sci Pharma Res 10:36–40. https://doi.org/10.22376/ijpbs/lpr.2020.10.4.l36-40
Delattre C, Pierre G, Laroche C, Michaud P (2016) Production, extraction and characterization of microalgal and cyanobacterial exopolysaccharides. Biotechnol Adv 34:1159–1179. https://doi.org/10.1016/j.biotechadv.2016.08.001
Dinev T, Tzanova M, Velichkova K et al (2021) Antifungal and antioxidant potential of methanolic extracts from Acorus calamus l., Chlorella vulgaris beijerinck, Lemna minuta kunth and Scenedesmus dimorphus (turpin) kützing. Appl Sci 11:4745. https://doi.org/10.3390/app11114745
Dubois M, Gilles KA, Hamilton JK et al (1956) Colorimetric method for determination of sugars and related substances. Anal Chem 28:350–356. https://doi.org/10.1021/ac60111a017
Ferreira AS, Ferreira SS, Correia A, Vilanova M (2020) Reserve, structural and extracellular polysaccharides of Chlorella vulgaris: a holistic approach. Algal Res 45:101757. https://doi.org/10.1016/j.algal.2019.101757
Fimbres-Olivarría D, López-Elías JA, Carvajal-Millán E et al (2016) Navicula sp. sulfated polysaccharide gels induced by Fe (III): rheology and microstructure. Int J Mol Sci 17(8):1238. https://doi.org/10.3390/ijms17081238
Gheda SF, Abo-Shady AM, Abdel-Karim OH, Ismail GA (2021) Antioxidant and antihyperglycemic activity of Arthrospira platensis (Spirulina platensis) methanolic extract: in vitro and in vivo study. Egypt J Bot 61:71–93. https://doi.org/10.21608/ejbo.2020.27436.1482
Guehaz K, Boual Z, Abdou I et al (2024) Microalgae’s polysaccharides, are they potent antioxidants ? Critical review. Arch Microbiol 206:14. https://doi.org/10.1007/s00203-023-03738-y
Guezoul O, Chenchouni H, Sekour M et al (2013) An avifaunal survey of mesic manmade ecosystems “Oases” in algerian hot-hyperarid lands. Saudi J Biol Sci 20:37–43. https://doi.org/10.1016/j.sjbs.2012.10.001
Halaj M, Matulová M, Capek P (2022) Structural features of biologically active extracellular polysaccharide produced by green microalgae Dictyosphaerium chlorelloides. Int J Biol Macromol 214:152–161. https://doi.org/10.1016/j.ijbiomac.2022.05.197
He X, Dai J, Wu Q (2016) Identification of sporopollenin as the outer layer of cell wall in microalga Chlorella protothecoides. Front Microbiol 7:1–11. https://doi.org/10.3389/fmicb.2016.01047
Hecker N, Costa LT, Farinha JC, Vevès T (1996) Inventaires des Zones Humides Méditerranéennes, Collecte des données, vol 2. Publication MedWet, Wetlands International/Instituto da Natureza, p 99
Jakhu S, Sharma Y, Sharma K et al (2021) Production and characterization of microalgal exopolysaccharide as a reducing and stabilizing agent for green synthesis of gold-nanoparticle: a case study with a Chlorella sp. from Himalayan high-altitude psychrophilic habitat. J Appl Phycol 33:3899–3914. https://doi.org/10.1007/s10811-021-02580-3
Jong-Yuh C, Mei-Fen S (2005) Potential hypoglycemic effects of Chlorella in streptozotocin-induced diabetic mice. Life Sci 77:980–990. https://doi.org/10.1016/j.lfs.2004.12.036
Kaliwal LPBB (2019) Microalga Scenedesmus bajacalifornicus BBKLP-07, a new source of bioactive compounds with in vitro pharmacological applications. Bioprocess Biosyst Eng 42(6):979–994. https://doi.org/10.1007/s00449-019-02099-5
Kamble P, Cheriyamundath S, Lopus M, Sirisha VL (2018) Chemical characteristics, antioxidant and anticancer potential of sulfated polysaccharides from Chlamydomonas reinhardtii. J Appl Phycol 30:1641–1653. https://doi.org/10.1007/s10811-018-1397-2
Koçer AT, İnan B, Kaptan Usul S et al (2021) Exopolysaccharides from microalgae: production, characterization, optimization and techno-economic assessment. Braz J Microbiol 52:1779–1790. https://doi.org/10.1007/s42770-021-00575-3
Kokarakis EJ, Nazos TT, Mavroudakis L et al (2022) Structural and physicochemical characterization of an aminosugar-rich exopolysaccharide isolated from a Chlorella sp. Algal Res 68:102881. https://doi.org/10.1016/j.algal.2022.102881
Liu L, Pohnert G, Wei D (2016) Extracellular metabolites from industrial microalgae and their biotechnological potential. Mar Drugs 14:191. https://doi.org/10.3390/md14100191
Liu Y, Tang Q, Duan X et al (2018) Antioxidant and anticoagulant activities of mycelia polysaccharides from Catathelasma ventricosum after sulfated modification. Ind Crops Prod 112:53–60. https://doi.org/10.1016/j.indcrop.2017.10.064
Malinowska E, Klimaszewska M, Strączek T et al (2018) Selenized polysaccharides—biosynthesis and structural analysis. Carbohydr Polym 198:407–417. https://doi.org/10.1016/j.carbpol.2018.06.057
Mashhadinejad A, Zamani H, Sarmad J et al (2016) Effect of growth conditions and extraction solvents on enhancement of antimicrobial activity of the microalgae Chlorella vulgaris. Pharm Biomed Res 2:65–73. https://doi.org/10.18869/acadpub.pbr.2.4.65
Mehta D, Shah D (2021) Cyanobacteria and microalgae growing on monuments of UNESCO World Heritage site Champaner Pavagadh, India: biofilms and their exopolysaccharide composition. Arch Microbiol 203:3425–3433. https://doi.org/10.1007/s00203-021-02334-2
Mousavian Z, Safavi M, Azizmohseni F et al (2022) Characterization, antioxidant and anticoagulant properties of exopolysaccharide from marine microalgae. AMB Express 12:27. https://doi.org/10.1186/s13568-022-01365-2
Mtaki K, Kyewalyanga MS, Mtolera MSP (2020) Assessment of antioxidant contents and free radical-scavenging capacity of Chlorella vulgaris cultivated in low cost media. Appl Sci 10:1–11. https://doi.org/10.3390/app10238611
Muthukumar J, Chidambaram R, Sukumaran S (2021) Sulfated polysaccharides and its commercial applications in food industries—a review. J Food Sci Technol 58:2453–2466. https://doi.org/10.1007/s13197-020-04837-0
Neumann U, Derwenskus F, Gille A et al (2018) Bioavailability and safety of nutrients from the microalgae Chlorella vulgaris, nannochloropsis oceanica and phaeodactylum tricornutum in C57BL/6 mice. Nutrients 10:65. https://doi.org/10.3390/nu10080965
Niccolai A, Chini Zittelli G, Rodolfi L et al (2019) Microalgae of interest as food source: biochemical composition and digestibility. Algal Res 42:101617. https://doi.org/10.1016/j.algal.2019.101617
Olasehinde TA, Mabinya LV, Olaniran AO, Okoh AI (2019) Chemical characterization of sulfated polysaccharides from Gracilaria gracilis and Ulva lactuca and their radical scavenging, metal chelating, and cholinesterase inhibitory activities. Int J Food Prop 22:100–110. https://doi.org/10.1080/10942912.2019.1573831
Priatni S, Budiwati TA, Ratnaningrum D et al (2016) Antidiabetic screening of some Indonesian marine cyanobacteria collection. Biodiversitas 17:642–646. https://doi.org/10.13057/biodiv/d170236
Priyadarshani I, Rath B (2012) Commercial and industrial applications of micro algae—a review. J Algal Biomass Util 3(4):89–100
Qi J, Kim SM (2017) Characterization and immunomodulatory activities of polysaccharides extracted from green alga Chlorella ellipsoidea. Int J Biol Macromol 95:106–114. https://doi.org/10.1016/j.ijbiomac.2016.11.039
Qian J, Bai Y, Tang J, Chen W (2015) Antioxidation and α-glucosidase inhibitory activities of barley polysaccharides modi fi ed with sulfation. LWT Food Sci Technol 64:104–111. https://doi.org/10.1016/j.lwt.2015.05.034
Qiu Y, Gao X, Chen R et al (2022) Metabolomics and biochemical insights on the regulation of aging-related diabetes by a low-molecular-weight polysaccharide from green microalga Chlorella pyrenoidosa. Food Chem X 14:100316. https://doi.org/10.1016/j.fochx.2022.100316
Raja R, Hema Iswarya S, Balasubramanyam D, Rengasamy R (2007) PCR-identification of Dunaliella salina (Volvocales, Chlorophyta) and its growth characteristics. Microbiol Res 162:168–176. https://doi.org/10.1016/j.micres.2006.03.006
Ramos-Romero S, Torrella JR, Pagès T et al (2021) Edible microalgae and their bioactive compounds in the prevention and treatment of metabolic alterations. Nutrients 13:1–16. https://doi.org/10.3390/nu13020563
Raposo MFDJ, De Morais AMMB, De Morais RMSC (2014) Influence of sulphate on the composition and antibacterial and antiviral properties of the exopolysaccharide from Porphyridium cruentum. Life Sci 101:56–63. https://doi.org/10.1016/j.lfs.2014.02.013
Sathasivam R, Radhakrishnan R, Hashem A, Abd-Allah EF (2019) Microalgae metabolites: a rich source for food and medicine. Saudi J Biol Sci 26:709–722. https://doi.org/10.1016/j.sjbs.2017.11.003
Senthilkumar T, Ashokkumar N (2012) Impact of Chlorella pyrenoidosa on the attenuation of hyperglycemia-mediated oxidative stress and protection of kidney tissue in streptozotocin-cadmium induced diabetic nephropathic rats. Biomed Prev Nutr 2:125–131. https://doi.org/10.1016/j.bionut.2012.01.006
Shaima AF, Mohd Yasin NH, Ibrahim N et al (2022) Unveiling antimicrobial activity of microalgae Chlorella sorokiniana (UKM2), Chlorella sp. (UKM8) and Scenedesmus sp. (UKM9). Saudi J Biol Sci 29:1043–1052. https://doi.org/10.1016/j.sjbs.2021.09.069
Sheng J, Yu F, Xin Z et al (2007) Preparation, identification and their antitumor activities in vitro of polysaccharides from Chlorella pyrenoidosa. Food Chem 105:533–539. https://doi.org/10.1016/j.foodchem.2007.04.018
Song H, He M, Gu C et al (2018) Extraction optimization, purification, antioxidant activity, and preliminary structural characterization of crude polysaccharide from an arctic Chlorella sp. Polymers (basel) 10:292. https://doi.org/10.3390/polym10030292
Sun Y, Wang H, Guo G et al (2014) The isolation and antioxidant activity of polysaccharides from the marine microalgae Isochrysis galbana. Carbohydr Polym 113:22–31. https://doi.org/10.1016/j.carbpol.2014.06.058
Tang H, Huang J, Yuan Q et al (2023) A regular Chlorella mannogalactan and its sulfated derivative as a promising anticoagulant: structural characterization and anticoagulant activity. Carbohydr Polym 314:120956. https://doi.org/10.1016/j.carbpol.2023.120956
Trabelsi L, Chaieb O, Mnari A et al (2016) Partial characterization and antioxidant and antiproliferative activities of the aqueous extracellular polysaccharides from the thermophilic microalgae Graesiella sp. BMC Complement Altern Med 16:1–10. https://doi.org/10.1186/s12906-016-1198-6
Wan XZ, Ai C, Chen YH et al (2020) Physicochemical characterization of a polysaccharide from green microalga Chlorella pyrenoidosa and its hypolipidemic activity via gut microbiota regulation in rats. J Agric Food Chem 68:1186–1197. https://doi.org/10.1021/acs.jafc.9b06282
Wang HB, Wu SJ, Liu D (2014) Preparation of polysaccharides from cyanobacteria Nostoc commune and their antioxidant activities. Carbohydr Polym 99:553–555. https://doi.org/10.1016/j.carbpol.2013.08.066
Wong CY, Teoh ML, Phang SM et al (2015) Interactive effects of temperature and uv radiation on photosynthesis of Chlorella strains from polar, temperate and tropical environments: differential impacts on damage and repair. PLoS One 10:1–14. https://doi.org/10.1371/journal.pone.0139469
Xia S, Gao B, Li A et al (2014) Preliminary characterization, antioxidant properties and production of chrysolaminarin from marine diatom Odontella aurita. Mar Drugs 12:4883–4897. https://doi.org/10.3390/md12094883
Xiong W, Chen J, He J et al (2022) Anti-diabetic potential of Chlorella Pyrenoidosa-based mixture and its regulation of gut microbiota. Plant Foods Hum Nutr 77:292–298. https://doi.org/10.1007/s11130-022-00968-1
Yadavalli R, Ratnapuram H, Motamarry S et al (2022) Simultaneous production of flavonoids and lipids from Chlorella vulgaris and Chlorella pyrenoidosa. Biomass Convers Biorefin 12:683–691. https://doi.org/10.1007/s13399-020-01044-x
Yu M, Chen M, Gui J et al (2019) Preparation of Chlorella vulgaris polysaccharides and their antioxidant activity in vitro and in vivo. Int J Biol Macromol 137:139–150. https://doi.org/10.1016/j.ijbiomac.2019.06.222
Yuan Q, Li H, Wei Z et al (2020) Isolation, structures and biological activities of polysaccharides from Chlorella: a review. Int J Biol Macromol 163:2199–2209. https://doi.org/10.1016/j.ijbiomac.2020.09.080
Zakaria SM, Mustapa Kamal SM, Harun MR et al (2017) Extraction of antioxidants from Chlorella sp. using subcritical water treatment. IOP Conf Ser Mater Sci Eng 206:012035. https://doi.org/10.1088/1757-899X/206/1/012035
Zhang B, Zhou Y (2017) Carbohydrate composition analysis in Xylem. Meth Mol Biol 1544:213–222. PMID: 28050838. https://doi.org/10.1007/978-1-4939-6722-3_15
Zhong Q, Wei B, Wang S et al (2019) The antioxidant activity of polysaccharides derived from marine organisms: an overview. Mar Drugs 17(12):674. https://doi.org/10.3390/md17120674
Acknowledgements
The authors thank Professor Chafaa Smail, Department of Biology, Batna 2 University, Batna, Algeria, for allowing access to the laboratory of the department.
Funding
Open access funding provided by Università degli Studi di Firenze within the CRUI-CARE Agreement. No funds, grants, or other support was received.
Author information
Authors and Affiliations
Contributions
K. G. Methodology, Investigation, Writing -Original Draft, Z. B. Design, Supervision. A. T. and H. M. Conceptualization, Resources. H. B, G. P. and P. M. Investigation, Formal Analysis. A. A. Supervision, Validation, Review and Editing.
Corresponding author
Ethics declarations
Conflict of interest
Authors declare that they have no competing interest that could influence this work.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Financial interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Communicated by Yusuf Akhter.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Guehaz, K., Boual, Z., Telli, A. et al. A sulfated exopolysaccharide derived from Chlorella sp. exhibiting in vitro anti-α-d-Glucosidase activity. Arch Microbiol 206, 218 (2024). https://doi.org/10.1007/s00203-024-03940-6
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00203-024-03940-6