
Abstract
Objective
Acute respiratory distress syndrome (ARDS) in young infants is linked with a pulmonary inflammatory response part of which are increased interleukin-8 (IL-8) levels and migration of polymorphonuclear leukocytes (PMNL) into lung tissue. A topical application of an antibody against IL-8 might therefore decrease PMNL migration and improve lung function.
Design
Randomized, controlled, prospective animal study.
Setting
Research laboratory of a university children’s hospital.
Subjects and interventions
Anesthetized, mechanically ventilated newborn piglets (n=22) underwent repeated airway lavage to remove surfactant and to induce lung inflammation. Piglets then received either surfactant alone (S, n=8), or a topical antibody against IL-8 admixed to surfactant (S+IL-8, n=8), or an air bolus injection (control, n=6).
Measurements and results
After 6 h of mechanical ventilation following intervention, oxygenation [S 169±51 (SD) vs S+IL-8 139±61 mmHg] and lung function (compliance: S 1.3±0.4 vs S+IL-8 0.9±0.4 ml/cmH2O/kg; extra-vascular lung-water: S 27±9 vs S+IL-8 52±28 ml/kg) were worse in the S+IL-8 group because reactive IL-8 production [S 810 (median, range 447–2323] vs S+IL-8 3485 (628–16180) pg/ml; P<0.05) with facilitated migration of PMNL into lung tissue occurred. Moreover, antibody application caused augmented chemotactic potency of IL-8 [linear regression of migrated PMNL and IL-8 levels: S r2=0.30 (P=ns) vs S+IL-8 r2=0.89 (P=0.0002)].
Conclusion
Topical anti-IL-8 treatment after lung injury increases IL-8 production, PMNL migration, and worsens lung function in our piglet lavage model. This effect is in contrast to current literature using pre-lung injury treatment protocols. Our data do not support anti-IL-8 treatment in young infants with ARDS.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Acute respiratory distress syndrome (ARDS) with impaired lung function and gas exchange is a condition eventually seen in newborn and young infants suffering from sepsis, meconium aspiration pneumonia, persistent pulmonary hypertension, inhalation syndrome, and bacterial or viral pneumonia [1, 2, 3]. ARDS has been defined as a severe lung disease with acute onset, bilateral infiltrates on chest radiograph, absence of left atrial hypertension, and a PaO2/FiO2-ratio ≤200 [4]. It is now well appreciated that ARDS hits patients of all ages including newborn and young infants with the conditions mentioned above [5]. ARDS can be thought of as the end result of an inflammatory process of the capillary-alveolar unit affecting the capillary-alveolar permeability, epithelial integrity, and type I and II pneumocyte function.
A key event of lung inflammation is the migration of polymorphonuclear leukocytes (PMNL) into lung tissue [6] which is mediated by a series of events such as rolling, adherence, and penetration of the capillary endothelium and migration into lung regions affected by the pro-inflammatory stimulus. PMNL are known to release a variety of enzymes in the lung such as oxidants, proteases (elastase, collagenase), leukotriens, and platelet factor, all together inducing damage to type I and II pneumocytes, functional compromise of surfactant, lung edema, and epithelial necrosis [7]. Mediators of chemotactic attraction for PMNL are the chemokines, an inhomogenous group of molecules such as IL-8, C5a, PAF, and LTB4, with IL-8 probably being the most potent chemoattractant factor [8].
Increased concentrations of pro-inflammatory cytokines attracting PMNL have been found in the tracheal aspirates from adult patients with ARDS [9, 10]. Kurdowska et al. [11] reported that a major portion of IL-8 in bronchoalveolar fluid in the ARDS lung is associated with anti-IL-8 autoantibody, thus inhibiting IL-8 interaction with its specific receptor on PMNL. This reaction seems to be part of the endogenous healing process of the lung. Several groups of authors have described intravenous anti-IL-8 application in different animal models of acute lung injury before induction of lung injury, with a favorable effect on mortality, lung function, and gas exchange [12, 13, 14, 15, 16].
The rationale of this study was to examine the effect of a monoclonal IL-8 antibody given topically by admixture to surfactant in a newborn piglet model of acute lung injury through repeated airway lavage, after induction of lung injury. Our intention was to better mimic the clinical situation of a young infant with established ARDS and admitted to an intensive care unit, when consideration of the unique immunologic situation of young infants [17, 18] would lead to preference of a topical rather than systemic treatment with drugs hampering the immunologic response.
The hypothesis of this study was that topical treatment with an antibody against IL-8 would reduce PMNL migration, followed by improvement in gas exchange, and lung function.
Materials and methods
Animal preparation
The experimental protocol was approved by the local Review Board for the Care of Animal Subjects in accordance with the German law for animal protection (BGBI 1, page 1319) and the European Community guidelines (86/609/EC).
Twenty-two newborn piglets (mixed country breed) of either sex from four litters were studied between day 2 and 10 of age, weighing 1.6–4.6 kg. Initially 0.025 mg/kg atropine, 10 mg/kg ketamine, and 1 mg/kg midazolam hydrochloride were administered intramuscularly. An oral intubation with an uncuffed 3.0 mm endotracheal tube (ETT) was performed after additional 5 mg/kg propofol intravenously. Anesthesia and muscle paralysis were maintained by 16 mg/kg/h propofol and 0.17 mg/kg/h pancuronium. To prevent leakage the ETT was tightly secured in place by a peritracheal ligature. Catheters were inserted into the right common carotid artery and into the left internal jugular vein.
Mechanical ventilation and airway lavage
Mechanical ventilation was provided by two time-cycled pressure limited infant ventilators (Babylog 1, Dräger, Lübeck, Germany). The following ventilator settings were used: FiO2 = 0.6, positive end-expiratory pressure (PEEP) = 4 mbar, flow = 8 l/min, inspiratory time = 0.4 s, ventilator rate = 35 bpm before lavage and 45 bpm during and immediately after lavage. Peak inspiratory pressure (PIP) was adjusted to keep tidal volume (VT) at approximately 6 ml/kg.
We used a modification of the original lavage protocol described by Lachmann et al. [19]. Each lavage involved the instillation and removal of 30 ml/kg of warmed normal saline via the ETT carried out over a 30-s period. After the first lavage PEEP was increased to 8 mbar and the ventilator rate to 45 bpm to ensure adequate oxygenation and ventilation and to fasten surfactant removal and alveolar collapse [20]. The first six lavages were performed in changing side positions. Airway lavage was repeated every 3–5 min until both the PaO2 decreased to approximately 40 mmHg, and a minimum PIP of ≥18 mbar was required to maintain VT at 6 ml/kg. Prior to baseline measurements the original PEEP level of 4 mbar was reestablished.
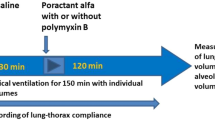
Experimental protocol
Measurements of gas exchange (PaO2 and PaCO2), lung function indices (dynamic compliance of the respiratory system (Crs), tidal volume (VT) and functional residual capacity (FRC)), and extravascular lung water (EVLW) were made before airway lavage, after airway lavage at baseline, and at 30 min, 1 h, 2 h, 3 h, 4 h, and 6 h after intervention (i.e., surfactant administration or air bolus). Ventilator rate was maintained at 45 bpm until completion of surfactant administration and was then switched back to 35 bpm for the remainder of the study. Animals of the control group were continued to be ventilated at 45 bpm.
At baseline the animals were randomized to one of the following protocols:
-
intervention group 1 (S) receiving 6 ml/kg surfactant and an unspecific antibody (n=8)
-
intervention group 2 (S+IL-8) receiving 6 ml/kg surfactant and an antibody against IL-8 (n=8)
-
a control group (C) receiving 6 ml/kg of air (n=6).
All animals of the two intervention groups received 100 mg/kg Curosurf (equivalent to a volume of 6 ml/kg) via a second lumen of the ETT within 2 min [21] and without interrupting mechanical ventilation.
Piglets of the two intervention groups were studied in pairs of two with the same sex and age. Control animals were randomized in three pairs of two. One piglet of the S+IL-8 group was later excluded from statistical analysis for possible airway infection.
Surfactant preparation and antibody
The surfactant preparation used in this study was Curosurf (Nycomed, Unterschleissheim, Germany) at a concentration of 20 mg/ml after dilution with normal saline to promote its physiologic response [22]. In the S group 1 ml of phosphate-buffered saline was added containing 500 µg of unspecific mouse IgG1 (ANC-278–010, Ancell, Bayport, USA). In the S+IL-8 group 500 µg of an anti-porcine IL-8 antibody (monoclonal mouse IgG1 from murine hybridoma cells, immunized with E.coli-derived porcine IL-8 and purified by protein G affinity chromatography; MAB 535, R&D Systems, Wiesbaden, Germany) was added to PBS. Absence of bacterial endotoxin in immunoglobulin products was confirmed by Limulus amoebocyte lysate assay (Pyrosperse, Cambrex Bio Science, Verviers, Belgium).
Measurement of gas exchange, lung function, and hemodynamic indices
PaO2 and PaCO2 were measured from blood samples taken from the carotid arterial catheter. To calculate Crs, VT, and FRC we used a multiple-breath nitrogen washout technique for lung volumes and the single-breath least squares regression method for lung mechanics as previously described [23].
Hemodynamic monitoring was performed using a PC 8000 PiCCO monitor (Pulsion, Munich, Germany). Cardiac output monitoring was accomplished using the transpulmonary indicator dilution technique (cold bolus of 3 ml normal saline <8 °C) and calculation of the area under the curve by the Stuart-Hamilton method:
where Tb is the blood temperature before cold injection, Ti is the temperature of the cold bolus, Vi is the cold bolus volume, C is a constant composed of specific weights and warmths of blood and cold bolus, and ∫ΔTb*dt is the area under the thermodilution curve.
Extravascular lung water (EVLW) is equivalent to the extravascular thermoaccessible volume in the lung assessed by the mean transit time method (needle to needle volume):
where ITTV is the intrathoracic thermoaccessible volume, and ITBV is the intrathoracic blood volume [24].
Clinical care
A heating pad was used to maintain a constant core temperature of the piglets between 38 °C and 39 °C. Each piglet received an infusion of D5 W in 1/3 normal saline at a fluid intake of 80 ml/kg/day.
Histologic processing and dry-wet ratios
After killing, lungs were removed from the thorax and the circulation was flushed with normal saline to remove blood. The right lower lobe was then put on a continuous distending pressure and fixed with 4% formaldehyde. Two tissue probes (approximately 20×20×4 mm) were taken from central and peripheral locations and cut at a width of 3 µm for staining with hematoxilin-eosin using standard methods. The following score was employed: 0=not at all; 1=mild; 2=moderate; 3=severe (also using steps of 0.5 for intermediate ranking) describing pathologic criteria such as alveolar collapse, lobar atelectasis, edema, interstitial infiltration, infiltration with PMNL, and necrosis of small airways.
Right middle lobe specimens of lung tissue were used to determine the degree of lung flooding in vitro. The specimens were dried in an oven at 60 °C overnight to calculate dry-wet-ratios.
In vitro testing of surfactant from BAL
To assess any detrimental effect of the used immunoglobulins on surfactant function, original surfactant preparations and BAL fluid gathered after 6 h of mechanical ventilation were tested for their minimal surface tensions using a modified Wilhelmy balance (E. Biegler, Mauerbach, Austria) applying 500 µl samples to the surface of a warmed saline-filled trough.
BAL and blood cell count
Native BAL fluid was gathered by gentle suction of the syringe which was used to push normal saline at body temperature into the airways of the piglets. BAL fluid from the left lung only (15 ml/kg) was used for measurements after 6 h of mechanical ventilation. BAL fluid was filtered to remove gross particles and was then centrifuged for 4 min at 4 °C and 2,500 rpm. Cellular material was resuspended into PBS for differential cell count.
Blood samples for differential blood cell counts were taken from the carotid catheter before the first lavage and after 6 h of mechanical ventilation before sacrifice of the piglets.
Cytokine measurements
IL-8 concentrations in BAL were determined using the QuantikineP porcine IL-8/CXCL8 immunoassay (R&D Systems, Wiesbaden, Germany). IL-6 and TNF-α were measured by bioassay. The IL-6 assay involved the use of a mouse hybridoma cell line (7TD1) which depends on the presence of IL-6 for its growth. The TNF-α assay based on the sensitivity of a porcine kidney cell line (PK15 clone 15) to cytotoxic effects of porcine TNF-α.
Migratests
The potency of lavage fluid (at 6 h after intervention) to induce migration of porcine granulocytes (chemotactic activity) was measured using the Migratest 24T (Becton Dickinson, Heidelberg, Germany) according to manufacturer instructions. Because of the varying chemotactic effect of surfactant preparations in the literature and the impact of concentration [25] we chose lavage fluid gathered after 6 h of mechanical ventilation reflecting the surfactant concentration in the individual lung.
Statistical methods
All mean values are expressed ±1SD. To establish comparability of the three experimental groups at baseline, we used a Kruskal-Wallis test which was also used for assessing differences in cytokine concentrations (Fig. 5) and differential cell counts (Table 1). Repeated measures analysis of variance (ANOVA) was used to assess differences between the two intervention groups in change over time (Fig. 1, Fig. 2, Fig. 3). A Mann Whitney U-test was used to evaluate differences in dry-wet-ratio, surface tension, histology score and cytokine levels (Fig. 4, Fig. 5) between the two intervention groups. These scores are displayed as box and whisker plots with the boxes extending from the 25th to the 75th percentile. Linear regression was used to analyze the relationship between IL-8 concentrations and PMNL-migration (Fig. 6) and are expressed as r2. Significant differences between groups were assumed to be present at values of P<0.05.

ANOVA (comparison of the two intervention groups only): PaO2 (P=0.65) and PaCO2 (P=0.16). Note: a different ventilator rate was used in the C group (45 instead of 35 bpm). Means±SD.

ANOVA (comparison of the two intervention groups only): Crs (P=0.02) and FRC (P<0.001). Means±SD.

ANOVA (comparison of the two intervention groups only): EVLW (P<0.0001). Means±SD.

Histological criteria assessing atelectasis, edema, and tissue infiltration for all three groups. Significant differences between the two intervention groups (Mann Whitney U-test) could not be detected. Median, boxes extending from the 25th to the 75th percentile, whiskers showing the highest and lowest values.

Cytokine levels in BAL fluid with the first lavage and after 6 h of mechanical ventilation. Kruskal-Wallis test showing no differences between all three groups with the first lavage; after 6 h significant differences for IL-8 only between S and S+IL-8 groups (P=0.03; Mann Whitney U-test) and S and C groups (P=0.02). Median, boxes extending from the 25th to the 75th percentile, whiskers showing the highest and lowest values. *Extreme value: 1,234 U/ml.

Migration of PMNL through a leukocyte separation medium using BAL fluid from lung lavage after 6 h of mechanical ventilation. Linear regression of migrated PMNL and IL-8 levels revealing a strong association in the S+IL-8 group only (r2=0.89, P=0.0002); S: r2=0.30 (P=ns); C: r2=0.53 (P=0.09).
Power analysis
To determine sample size we chose the main outcome parameter, PMNL in BAL after 6 h of mechanical ventilation, and used the following formula:
with
-
Zα = 1.96 using an α (type I error) = 0.05
-
Zβ = -0.842 using a β (type II error) = 0.2
-
σ = 50 PMNL/µl as an estimated standard deviation (SD)
-
Δ = 50 PMNL/µl as an clinical important difference in PMNL density in lavage fluid.
Therefore, we used eight piglets in the two intervention groups (and—to save animals—six piglets in the control group).
Results
Comparability of study groups
Twenty-two piglets (11 pairs of same sex and age) from four litters were consecutively studied after randomization at the completion of airway lavage. One piglet of the S+IL-8 group was excluded for reasons mentioned above. Significant differences between groups could be ruled out for weight, the number of lavages employed (S: 12.6±5.6; S+IL-8: 13.3±5.9; C: 11.8±3.7) and the final PIP (S: 22.8±4.1; S+IL-8: 23.0±3.3; C: 23.0±3.7 mbar) suggesting even damage to airways and lung tissue.
At baseline (completion of airway lavage) there were no statistically significant differences between the three groups for PaO2, PaCO2, Crs, VT, FRC, and EVLW.
Clinical parameters
Figure 1, Fig. 2 and Fig. 3 display the clinical parameters of gas exchange, lung function and hemodynamics over a study period of 6 h after lavage. Significant differences between the two intervention groups for all parameters in change over time (PaO2 and PaCO2 excepted) could be detected.
Histology, dry-wet-ratio, and surface tension
Figure 4 displays six histological criteria for evaluation of atelectasis, edema, and cell infiltration into lung tissue yielding decreased infiltration (however, not significant) in the S group by this semi-quantitative approach. The dry-wet-ratio was: S 11.2±3.9%; S+IL-8 10.5±6.8%; C 9.6±0.5% (S vs S+IL-8: P=ns). Minimal surface tensions of surfactant from BAL fluid after 6 h of mechanical ventilation were: S 25±3 mN/m; S+IL-8 25±2 mN/m, C 43±9 mN/m (S vs S+IL-8: P=ns). Minimal surface tensions of surfactant incubated in a cuvette and mixed with either unspecific IgG1 or IL-8 antibody after 6 h were: S 8.0±0.6 mN/m; S+IL-8 6.1±2.8 mN/m, indicating no influence of the applied immunoglobulins on surface properties of the diluted surfactant preparation.
Leukocytes in BAL and blood
Cell counts from BAL fluid at first lavage and peripheral blood did not reveal any statistical differences. After 6 h of mechanical ventilation, piglets of the S group showed lower complete cell counts and lower absolute PMNL counts in BAL fluid (Table 1). We observed a decreased percentage of PMNL and an increased percentage of macrophages in the C group. Differential cell counts from blood (also Table 1) showed differences in absolute PMNL counts almost reaching statistical significance (P=0.05).
Cytokine assays
For the three cytokines IL-6, TNF-α and IL-8 no differences between groups with the first lavage could be determined (Fig. 5). After 6 h of mechanical ventilation, IL-8 levels in the S+IL-8 group were significantly higher than in the S group [S: median 810 (range 447–2323) pg/ml, S+IL-8: 3485 (628–16180), P=0.03]; and S group levels higher than C group values [C: 352 (159–752); P=0.02]. No differences could be detected for IL-6 and TNF-α.
Migratests
Linear regression revealed a strong r2 for BAL fluid of the S+IL-8 group piglets and an uncoupling of chemotactic potency of IL-8 on PMNL migration in the S group (Fig. 6).
Discussion
The results of our study demonstrate that topical administration of a monoclonal IgG1 anti-porcine IL-8 antibody admixed to surfactant results in an increased expression of IL-8, released into the airways of newborn piglets after repeated airway lavage. This reaction was followed by an enhanced migration of PMNL into lung tissue and alveoli. We observed lung tissue infiltration with PMNL and other cells which occurred hand in hand with impaired gas exchange, lung mechanics and lung volumes, and fluid lungs as assessed by EVLW and dry-wet-ratio. On the other hand, surfactant alone (i.e., without admixture of a specific antibody) downregulated the inflammatory process, not at the level of cytokine expression but at the level of cell infiltration. Surfactant alone also caused superior improvement in lung function when compared to surfactant + antibody treatment. The admixture of a topical antibody against IL-8 did not interfere with surfactant function as demonstrated by equal minimal surface tension of surfactant from BAL fluid taken after 6 h of mechanical ventilation.
Clearly, there is only limited experience with the IL-8 antibody which was used in our experiment. To neutralize porcine IL-8 -induced myeloperoxidase release from human neutrophils, Schröder et al. [26] have shown that an ND50 of approximately 3–10 µg/ml was needed to suppress myeloperoxidase production. The topical dose of the antibody was calculated by the following equation: average body weight (2.8 kg) × average FRC (26 ml/kg) × median neutralizing dose (7 µg/ml), yielding approximately a dose of 500 µg per piglet.
In contrast to our hypothesis, IL-8 expression was magnified by lung tissue in the S+IL-8 group. Moreover, in vitro migration of PMNL by migratests was strongly related in the S+IL-8 group only. We suggest that the antibody’s activity was probably neutralized by a burst expression of IL-8, thus leaving enough unbound IL-8 to unfold its activity. To find out whether this compensatory reaction was specific for the lung after a topical treatment, we tested some more piglets undergoing the same lung lavage protocol followed by a systemic application of the IL-8 antibody with increasing doses: 0.1 µg/kg, 1 µg/kg, and 10 µg/kg. The following IL-8 levels could be detected in BAL fluid after 6 h of mechanical ventilation: 0.1 µg/kg (median and range): 10,160 pg/ml (3,785–18,540); 1 µg/kg 16,290 pg/ml (1,633–21,500); 10 µg/kg: 65,780 pg/ml (40,850–90,710). The corresponding serum levels were: 0.1 µg/kg: 0 pg/ml (0–32); 1 µg/kg: 32 pg/ml (0–134); 10 µg/kg: 651 pg/ml (212–1091). We therefore conclude that topical as well as systemic application of a monoclonal anti-porcine IL-8 antibody elicits an upward regulation in the transcription of IL-8 which counteracts the properties of this antibody as originally shown in cell culture media.
Another interesting finding of our study was the surfactant effect on IL-8 induced PMNL migration. As shown on Table 1, a trend to lower relative and absolute PMNL concentrations in the BAL fluid was observed in those piglets from the S group. We suggest that the difference in PMNL infiltration into lung tissue as assessed by histology and PMNL concentration in BAL fluid (although statistically not significant) was linked with improved parameters of gas exchange, lung function, and hemodynamics. In particular, all parameters influenced by lung edema (Crs, EVLW, and dry-wet-ratio) were clearly improved in the S group when compared to the S+IL-8 group. Even though IL-8 concentrations in BAL fluid were least in those piglets of the C group, PMNL concentrations were least in the S group revealing an uncoupling of IL-8 effect and chemotactic activity on PMNL migration (S: r2=0.30; C: r2=0.53; Fig. 6). This means that the anti-inflammatory effect of surfactant does not exert its action on the level of chemokine production, but on PMNL migration by an unknown mechanism.
Some studies support our findings of uncoupled IL-8 concentrations and PMNL migration into lung tissue when giving surfactant for lung disease. Adult patients suffering from ventilator-associated pneumonia were studied to evaluate the effect of nebulized synthetic phospholipids (Exosurf) on IL-8 levels in tracheal aspirates [27]. IL-8 levels increased in the intervention group (mean: surfactant 318, vs control 156 ng/ml BAL fluid) whereas percentage of PMNL decreased concomitantly (absolute numbers of PMNL in BAL fluid were not given in this study.) In a premature lamb model with RDS it could be shown that mRNA levels for IL-8 and PMNL infiltration into lung tissue were not matched sufficiently regardless of the type of surfactant used [28]. Putative explanations might be that increased expression of IL-8 receptors desensitize C5a and LTB4 receptor pathways hindering PMNL to follow the dominant signal such as IL-8 [29].
Production and release of IL-8 in the lung takes place in a variety of different cell types such as epithelial cells of large and small airways, type I pneumocytes, and alveolar macrophages [30, 31]. As soon as it is built in the cells, IL-8 diffuses to the epithelium of small capillaries and unfolds its chemotactic properties on PMNLs passing by [14]. IL-8 concentrations and PMNL concentrations are well-matched in BAL fluid from adult patients with ARDS [32] with IL-8 levels being highest in those patients who do not survive [33]. Anti-IL-8 autoantibodies are produced by adult patients with ARDS to form anti-IL-8:IL-8 complexes which predominantly belong to IgG3 and IgG4 subclasses [11]. IgA autoantibodies are also known to be complexed with IL-8 as seen in severe asthma [34]. High IL-8 and anti-IL-8 IgM autoantibody concentrations have been measured in the tracheal aspirates of mechanically ventilated premature infants at risk of chronic lung disease [35], whereas a significant increase in anti-IL-8 IgG autoantibodies could not be detected [36] suggesting immature IgG secretion in the preterm infant [17].
Several groups of authors have addressed the option to suppress the pro-inflammatory effects of IL-8 in animal models of acute lung injury. Folkesson et al. [14] used acid aspiration in rabbits and observed decreased lung damage and mortality. PMNL migration could be lessened by IL-8 antibodies given to rabbits after experimental ischemia and reperfusion [12]. Induction of lung disease in rats after IgG-immune complex administration could be blunted by an IL-8 antibody with the effect of less capillary permeability and extravasal red blood cells [13]. Accumulation of PMNL in BAL could be decreased in rabbits after lung trauma with repeated collapse and reexpansion [16]. Endotoxin-induced lung injury in rabbits was relieved by IL-8 antibody resulting in improved gas exchange and less mortality [15]. All authors used monoclonal anti-IL-8 antibodies of the IgG type as also used in our study. However, there are three important differences between these studies and our experiment: the application of a systemic antibody against IL-8 was made before induction of lung injury in adult animals, which might have resulted in a completely different effect as demonstrated above.
Several limitations of this study must be addressed: a) our piglet model of repeated airway lavage is characterized by lung edema, infiltration with blood-borne cells, impaired lung mechanics and lung volumes, surfactant deficiency and dysfunction. The model mimics newborn and infant ARDS, but the vascular injury such as perivascular cell injury [37] and the full inflammatory process [38], however, may not be completely reproduced in this model. Secondary injuries such as baro- and volutrauma, very high oxygen concentrations for ventilation, nosocomial infections of the lung, and associated multiple organ failure are not represented. Conventional ventilation with low lung volume strategies using a rather low PEEP level (e.g. 2 mbar as shown by Rotta et al. [39] and Suh et al. [40]) would have even increased the inflammatory response with additional PMNL infiltration, cytokine expression, and capillary-alveolar leak; however our aim was to duplicate standard ventilatory protocols as used in our intensive care unit. b) the presence of an IL-8 antibody in BAL fluid might affect the ability of an ELISA assay to detect IL-8 and to raise IL-8 concentrations factitiously; c) the appearance of early anti-IL-8 autoantibody production was not addressed; d) dosing of the IL-8 antibody was calculated from our piglets’ FRC and the neutralizing dose from a single experimental study with cell cultures [26]. Information on kinetics of the antibody and interactions with different surfactant components are not known.
In conclusion, this study demonstrates that topical application of an antibody against IL-8 leads to increased expression of IL-8 from lung tissue which is linked with impaired gas exchange, lung function, and hemodynamics due to a more extensive degree of PMNL migration into lung tissue. Surfactant application alone blunts IL-8 activity and reduces PMNL migration with advantages for all parameters mentioned above. Surfactant alone does not reduce IL-8 expression by the lung. In contrast to current literature, this paradoxical effect might be the consequence of a topical anti-IL-8 application after induction of lung injury in an immature lung. Contrary to our hypothesis, treatment of infant ARDS with a topical anti-IL-8 antibody does not seem to be a treatment option according to the results obtained in this study.
References
Lotze A, Mitchell BR, Bulas DI, Zola EM, Shalwitz RA, Gunkel JH, and the Survanta in Term Infants Group (1998) Multicenter study of surfactant (beractant) use in the treatment of term infants with severe respiratory failure. J Pediatr 132:40–47
LeVine AM, Lotze A, Stanley S, Stroud C, O’Donnell R, Whitsett J, Pollack MM (1996) Surfactant content in children with inflammatory lung disease. Crit Care Med 24:1062–1067
Marraro GA, Luchetti M, Galassini EM, Abbiati G (1999) Natural surfactant supplementation in ARDS in pediatric age. Minerva Anestesiol 65:92–97
Bernard GR, Artigas A, Brigham KL, Carlet J, Falke K, Hudson L, Lamy M, Legall JR, Morris A, Spragg R (1994) The American-European Consensus Conference on ARDS. Definitions, mechanisms, relevant outcomes, and clinical trial coordination. Am J Respir Crit Care Med 149:818–824
Anderson MR (2003) Update on pediatric respiratory distress syndrome. Respir Care 48:261–276
Rossi AG, Haslett C (1998) Inflammation, cell injury, and apoptosis. In: Said SI (ed) Proinflammatory and antiinflammatory peptides. Lung biology in health and disease. Marcel Dekker, New York, pp 9–24
Ware LB, Matthay MA (2000) The acute respiratory distress syndrome. N Engl J Med 342:1334–1349
Vaddi K, Keller M, Newton RC (1997) The chemokine factsbook. Academic Harcourt Brace, San Diego, pp 23–37
Meduri GU, Headley S, Kohler G, Stentz F, Tolley E, Umberger R, Leeper K (1995) Persistent elevation of inflammatory cytokines predicts a poor outcome in ARDS. Plasma IL-1beta and IL-6 levels are consistent and efficient predictors of outcome over time. Chest 107:1062–1073
Meduri GU, Kohler G, Headley S, Tolley E, Streutz F, Postlethwaite A (1995) Inflammatory cytokines in the BAL of patients with ARDS; persistent elevation over time predicts poor outcome. Chest 108:1303–1314
Kurdowska A, Miller EJ, Noble JM, Baughman RP, Matthay MA, Brelsford WG, Cohen AB (1996) Anti-IL-8 autoantibody in alveolar fluid from patients with the adult respiratory distress syndrome. J Immunol 157:2699–2706
Sekido N, Mukaida N, Harada A, Nakanishi I, Watanabe Y, Matsushima K (1993) Prevention of lung reperfusion injury in rabbits by a monoclonal antibody against interleukin-8. Nature 365:654–657
Mulligan MS, Jones ML, Bolanowski MA, Baganoff MP, Deppeler CL, Meyers DM, Ryan US, Ward PA (1993) Inhibition of lung inflammatory reactions in rats by an anti-human IL-8 antibody. J Immunol 150:5585–5595
Folkesson HG, Matthay MA, Hebert C, Broaddus VC (1995) Acid aspiration-induced lung injury in rabbits is mediated by IL-8 dependent mechanisms. J Clin Invest 96:107–116
Yokoi K, Mukaida N, Harada A, Watanabe Y, Matsushima K (1997) Prevention of endotoxemia-induced acute respiratory distress syndrome-like lung injury in rabbits by a monoclonal antibody to IL-8. Lab Invest 76:375–384
Nakamura M, Fujishima S, Sawafuji M, Ishizaka A, Oguma T, Soejima K, Matsubara H, Tasaka S, Kikuchi K, Kobayashi K, Ikeda E, Sadick M, Hebert CA, Aikawa N, Kanazawa M, Yamaguchi K (2000) Importance of interleukin-8 in the development of reexpansion lung injury in rabbits. Am J Respir Crit Care Med 161:1030–1036
Andersson U, Bird G, Britton S, Palacios R (1981) Humoral and cellular immunity in humans studied at the cell level from birth to two years of age. Immunological Rev 57:5–38
Metinko A (2004) Neonatal pulmonary host defense mechanisms. In: Polin RA, Fox WW, Abman SH (eds) Fetal and neonatal physiology. Saunders, Philadelphia, pp 1620–1673
Lachmann B, Robertson B, Vogel J (1980) In vivo lung lavage as an experimental model of the respiratory distress syndrome. Acta Anaesthesiol Scand 24:231–236
Krause MF, Hoehn T (1999) Decrease in lung volume depends on end-expiratory pressure in a rabbit model of airway lavage. Respiration 66:259–264
Krause MF, Schulte-Mönting J, Hoehn T (1998) Rate of surfactant administration influences lung function and gas exchange in a surfactant-deficient rabbit model. Pediatr Pulmonol 25:196–204
Van der Bleek J, Plötz F, Van Overbeek F, Heikamp A, Beekhuis H, Wildevuur C, Okken A, Bambang Oetomo S (1993) Distribution of exogenous surfactant in rabbits with severe respiratory failure: the effect of volume. Pediatr Res 34:154–158
Krause MF, Jäkel C, Haberstroh J, Schulte-Mönting J, Leititis JU, Orlowska-Volk M (2001) Alveolar recruitment promotes homogenous surfactant distribution in a piglet model of lung injury. Pediatr Res 50:34–43
Schiffmann H, Erdlenbruch B, Singer D, Singer S, Herting E, Hoeft A, Buhre W (2002) Assessment of cardiac output, intravascular volume status, and extravascular lung water by transpulmonary indicator dilution in critically ill neonates and infants. J Cardiothorac Vasc Anesth 16:592–597
Scholtes U, Wiegand N, Zwirner J, Herting E (2002) Influence of porcine natural modified surfactant on chemotaxis and oxidative metabolism of polymorphonuclear leukocytes. Immunobiol 205:290–302
Schröder JM, Mrowietz U, Morita E, Christophers E (1987) Purification and partial biochemical characterization of a human monocyte-derived, neutrophil-activating peptide that lacks interleukin 1 activity. J Immunol 139:3474–3483
Baughman RP, Henderson RF, Whitsett J, Gunther KL, Keeton DA, Waide JJ, Zaccardelli DS, Pattishall EN, Rashkin MC (2002) Surfactant replacement for ventilator-associated pneumonia: a preliminary report. Respiration 69:57–62
Ikegami M, Jobe AH (2002) Injury responses to different surfactants in ventilated premature lamb lungs. Pediatr Res 51:689–695
Ali H, Richardson RM, Haribabu B, Snyderman R (1999) Chemoattractant receptor cross-desensitization. J Biol Chem 274:6027–6030
Nakamura H, Yoshimura K, Jaffe HA, Crystal RG (1991) Interleukin-8 gene expression in human bronchial epithelial cells. J Biol Chem 266:19611–19617
Rolfe MW, Kunkel SL, Standiford TJ, Chensue SW, Allen RM, Evanoff HL, Phan SH, Strieter RM (1991) Pulmonary fibroblast expression of interleukin-8: a model for alveolar macrophage-derived cytokine networking. Am J Respir Cell Mol Biol 5:493–501
Chollet-Martin S, Jourdain B, Gibert C, Elbim C, Chastre J, Gougerot-Pocidalo MA (1996) Interactions between neutrophils and cytokines in blood and alveolar spaces during ARDS. Am J Respir Crit Care Med 153:594–601
Kiehl MG, Ostermann H, Thomas M, Muller C, Cassens U, Kienast J (1998) Inflammatory mediators in bronchoalveolar lavage fluid and plasma in leukocytopenic patients with septic shock-induced acute respiratory distress syndrome. Crit Care Med 26:1194–1199
Shute JK, Vrugt B, Lindley IJD, Holgate ST, Bron A, Aalbers R, Djukanovic R (1997) Free and complexed interleukin-8 in blood and bronchial mucosa in asthma. Am J Respir Crit Care Med 155:1877–1883
Takasaki J, Ogawa Y (2001) Anti-interleukin-8 autoantibody in the tracheobronchial aspirate of infants with chronic lung disease. Pediatr Int 43:48–52
Takasaki J, Ogawa Y (2000) Anti-IL-8 autoantibodies in the tracheobronchial aspirate of infants in the development of chronic lung disease. Acta Neonatol Jpn 36:1–5
Nielsen JB, Sjöstrand UH, Edgren EL, Lichtwarck-Aschoff M, Svensson BA (1991) An experimental study of different ventilatory modes in piglets in severe respiratory distress induced by surfactant depletion. Intensive Care Med 17:225–233
Von der Hardt K, Kandler MA, Fink L, Schoof E, Dötsch J, Brandenstein O, Bohle RM, Rascher W (2004) High frequency oscillatory ventilation suppresses inflammatory response in lung tissue and microdissected alveolar macrophages in surfactant depleted piglets. Pediatr Res 55:339–346
Rotta AT, Gunnarsson B, Fuhrman BP, Hernan LJ, Steinhorn DM (2001) Comparison of lung protective ventilation strategies in a rabbit model of acute lung injury. Crit Care Med 29:2176–2184
Suh GY, Koh Y, Chung MP, An CH, Kim H, Jang WY, Han J, Kwon OJ (2002) Repeated derecruitments accentuate lung injury during mechanical ventilation. Crit Care Med 30:1848–1853
Acknowledgments
The surfactant preparation used in this study (Curosurf) was generously provided by Nycomed, Unterschleissheim, Germany. The authors are indebted to Jürgen Schulte-Mönting for statistical advice.
Author information
Authors and Affiliations
Corresponding author
Additional information
Financial support: supported by Hübner-Stiftung im Stifterverband, Essen; and Deutsche Forschungsgemeinschaft, Bonn, grant KR 1863/1-1
Rights and permissions
About this article
Cite this article
Ankermann, T., Wiemann, T., Reisner, A. et al. Topical interleukin-8 antibody attracts leukocytes in a piglet lavage model. Intensive Care Med 31, 272–280 (2005). https://doi.org/10.1007/s00134-004-2464-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00134-004-2464-5