
Abstract
Key message
The bs5 resistance gene against bacterial spot was identified by map-based cloning.
Abstract
The recessive bs5 gene of pepper (Capsicum annuum L.) conditions a non-hypersensitive resistance trait, characterized by a slightly swollen, pale green, photosynthetically active leaf tissue, following Xanthomonas euvesicatoria infection. The isolation of the bs5 gene by map-based cloning revealed that the bs5 protein was shorter by 2 amino acids as compared to the wild type Bs5 protein. The natural 2 amino acid deletion occurred in the cysteine-rich transmembrane domain of the tail-anchored (TA) protein, Ca_CYSTM1. The protein products of the wild type Bs5 and mutant bs5 genes were shown to be located in the cell membrane, indicating an unknown function in this membrane compartment. Successful infection of the Bs5 pepper lines was abolished by the 6 bp deletion in the TM encoding domain of the Ca_CYSTM1 gene in bs5 homozygotes, suggesting, that the resulting resistance might be explained by the lack of entry of the Xanthomonas specific effector molecules into the plant cells.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Plants are regularly invaded by beneficial or pathogenic micro-organisms, mainly bacteria and fungi (Wille et al. 2019). Endosymbiotic Rhizobia, endomycorrhizal fungi, endophytic, epiphytic and rhizospheric microbes that live inside or outside their host plants all form beneficial associations (Afzal et al. 2019). On the other hand, pathogenic micro-organisms establish harmful interactions which are detrimental to the host plant except when the plant is resistant against the invader. Resistant plants have evolved defense mechanisms which are related to animal innate immunity, but plants have an arsenal of diverse recognition mechanisms as opposed to the animal adaptive immune system (Dodds and Rathjen 2010).
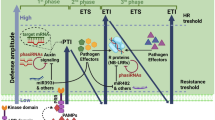
The plant immune system possesses two strategies to detect and fight against pathogens. At first, plants evolved an external receptor system in which pattern recognition receptors (PRRs) can recognize and bind conserved microbial elicitor molecules, called microbial-associated molecular patterns (MAMPs) or pathogen-associated molecular patterns (PAMPs). Stimulation of PRRs by elicitors leads to MAMP-triggered immunity (MTI) or PAMP-triggered immunity (PTI). MTI/PTI is considered to be the primary layer of inducible defense (basal resistance) against pathogenic intruders (Dixon et al. 2000; Jones and Dangl 2006). The detection of certain PAMPs/MAMPs can induce a hypersensitive response (HR), of programmed cell death, at the point of pathogen intrusion (Klement et al. 1964; Flor 1971; Balint-Kurti 2019), although, at present, most PAMPs/MAMPs thus far identified do not induce cell death following perception (Ingle et al. 2006). PTI is more commonly associated with induction of the production of a range of antimicrobial compounds and cell wall thickening, known as callose depositions, in the vicinity of the detected PAMP (Ingle et al. 2006).
Plants also evolved an intracellular receptor system, in which resistance proteins (R proteins) recognize, directly or indirectly, pathogen virulence molecules called effectors (protein products of avirulence genes; Keen 1990; Van der Biezen and Jones 1998). The resulting complex induces effector-triggered immunity (ETI) events that lead to rapid defense response (Jones and Dangl 2006; Boller and Felix 2009; Dodds and Rathjen 2010; Dolatabadian 2020; Harris et al 2020; Wang et al 2022). In contrast to MTI/PTI, in the large majority of cases, ETI is associated with HR (Balint-Kurti 2019). HR is the result of an interaction between a dominant resistance gene (R gene) in the host with a dominant corresponding avirulence gene (Avr gene) in the pathogen. This interaction is referred to as the gene-for-gene relationship, a concept discovered by H. H. Flor and elaborated by other colleagues (Klement et al. 1964; Flor 1971; Higgins et al. 1998; Heath 2000; Balint-Kurti 2019). Resistance is conferred only if both R-gene, present in either homo- or heterozygous genetic configuration, and the corresponding Avr gene are present in the same interaction. Many R-gene and Avr genes have been cloned (Kourelis and van der Hoorn 2018; Garcia-Ruiz et al. 2021; Calle García et al. 2022).
PTI/MTI is generally effective against non-adapted pathogens in a phenomenon called non-host resistance, whereas ETI is active against adapted pathogens. However, these relationships are not exclusive and depend on the elicitor molecules present in each infection. Successful pathogens are able to suppress PTI responses and thereby multiply and cause disease (Heath 2000; Panstruga and Moscou 2020).
Plants are vulnerable to pathogenic microbes when the pathogen produces a virulence factor and the plant can not develop a resistance response but possess dominant susceptibility genes (S genes). In this compatible interaction plants become seriously diseased resulting in an abnormal physiological process that disrupts the plant's normal structure, growth, function, or other activities. Recessive S gene alleles, conferring resistance, have been identified following mutagenesis or as natural variants (e.g., barley mlo, xa13 and eIF4, respectively, conferring resistance to powdery mildew, Xanthomonas (X) bacteria, and potyvirus; Yuan et al. 2009; Kusch and Panstruga 2017; Schmitt-Keichinger 2019). Since recessive S gene alleles conditioning resistance are more durable than major resistance (R) genes (Stall et al. 2009; Palloix et al. 2009), they are more valuable resources in disease resistant crop breeding and that is the case for pepper breeding (Garcia-Ruiz et al. 2021).
Pepper, Capsicum annuum L. (Ca) and tomato, Solanum lycopersicum L. plantations around the world are under continuous threat by the bacterial pathogen, Xanthomonas euvesicatoria (Xe), the causal agent of bacterial spot disease (Stall et al. 2009). An epidemic of bacterial spot disease can cause significant loss in pepper and tomato production when elevated temperature combines with high humidity. To fight against this deleterious pathogen in pepper, dominant (Bs1, Bs2, Bs3, Bs4C, Bs7) and recessive (bs5, bs6, bs8) resistance genes have been identified in wild type pepper species (Ronald and Staskawicz 1988; Jones et al. 2002; Römer et al. 2007; Tai et al. 1999; Potnis et al. 2012; Strauß et al. 2012; Sharma et al. 2022) and introgressed into cultivars (Vallejos et al. 2010). Unfortunately, there is no durable natural resistance determinant against Xe in tomato available for breeding (Stall et al. 2009). Dominant Bs1, Bs2, Bs3, Bs4C and Bs7 genes confer unstable resistance because new races of Xe breakdown the resistance trait (Stall et al. 2009). Nevertheless, these dominant genes have been cloned and characterized thoroughly; however, the recessive genes bs5, bs6 and bs8 remained in obscurity, though the investigation of the molecular basis of recessive genes is relevant in understanding the molecular basis of virulence (Garcia-Ruiz et al. 2021) and the infection process leading to cell and tissue necrosis after infection. In this paper, we describe the map-based cloning and characterization of the bs5 gene and its protein product conditioning recessive resistance trait against Xe in pepper.
Materials and methods
Bacterium and plant material used in this study
For the determination of disease symptoms, pepper plants were inoculated by pathogenic Xe strain 78, (abbreviated as Xe78) obtained from the microbial collection of Corvinus University of Budapest, Hungary, kindly provided by Laszlo Palkovics. Using specific primer pairs in a PCR amplification experiment, it was demonstrated that Xe78 does not carry genes for avrBS1, avrBS2, avrBS3 and avrBS4. In addition, implementing a standard identification protocol using differential hosts Xe78 could be classified as Race 10 (Kurowski et al. 2010). Xe species were previously designated as X. campestris pv. vesicatoria (Jones et al. 2006). X. perforans (strain NCPPB No.4321) was kindly provided by Jeffrey B. Jones, University of Florida, Gainesville, Florida, USA. X. gardneri (kindly provided by Dr. Aleksa Obradovic, University of Belgrade). The following plant materials were used in this study: (i) Xe susceptible (XeS) material: Ca cv. Early CalWonder (ECW) was kindly provided by Jeffrey B. Jones, University of Florida, Gainesville, Florida, USA. (Jones et al. 2002); Ca cv. Feherozon (CaFo), a commertially available Hungarian „Sweet Bell pepper” cultivar (https://www.kertimag.hu/bogyosok/feherozon.html). (ii) Xe resistant (XeR) material: Ca cv. 50R (ECW50R) was kindly provided by Jeffrey B. Jones, University of Florida, Gainesville, Florida, USA. (Vallejos et al. 2010); Ca var. T1 (CaT1), a single seed plant from accession no. PI163192 of AVRDC—The World Vegetable Center Gene Bank, Shanhua, Taiwan (http://avrdc.org/); Ca var. DH99-269 (CaDH269) was kindly provided by G. Csilléry, Budakert Ltd, Budapest, Hungary (Szarka 2008; Csillery et al. 2004); Ca cv Global (CaGl) is a commercially available Hungarian cultivar, https://sites.google.com/site/fpkutato/fajtaink/cseresznyepaprika-fajtk). The genotypes of the above plants are given in Fig. S1.
Plant growth conditions and crossing
Before use, pepper seeds were kept at 4 °C for one to two weeks for vernalization, then sown, geminated and grown under greenhouse conditions in 4 inch pots containing “Florimo®” general flower soil (http://www.florimo.hu/termekeink/viragfoldek/) at 25 °C for 16/8 h light/dark period until flowering. To generate F1 hybrids, maternal flowers were emasculated and manually crossed with pollen from the paternal parent. F2 and F3 seeds were collected from self-pollinated plants.
Xe infection and evaluation of disease symptoms
For the determination of the XeR/XeS phenotype of individual plants, leaf infections with Xe78 bacteria were performed. Bacteria were grown in Nutrient Broth (Sigma-Aldrich Co.) at 28 °C until stationary phase, pelleted by centrifugation and suspended in sterile tap water at a concentration of 105 CFU/mL. Inoculation was done with 1–2 × 104 Xe78 bacteria per leaf. The abaxial side of young leaves (8-leaf stage) was infiltrated into the intercellular space of the leaves with a syringe fitted to a rubber tube. Infected plants were grown under greenhouse conditions as described above. The evaluation of the disease symptoms was as follows: Xe resistant plant leaves showed pale green discoloration, healthy, but swollen tissue (Fig. 1) following 6 days after inoculation (DAI) and in later stage as well (Szarka and Csilléry 1995; Szarka et al. 2006; Szarka 2008). Abscission of the leaves was not observed. Xe susceptible plant leaves exhibited yellowing, watersoaking after 4–6 DAI, complete necrosis after 6–12 DAI and abscission of the infected leaves after two weeks.

Disease symptoms of leaves following infection with Xe78. Leaf phenotype of parents (CaT1, CaFo) and F1 (CaT1 x CaFo) hybrid plants after 6 days post inoculation with Xe78. Blue arrows show resistant, while red arrows show the susceptible leaf phenotype. The inoculation procedure and phenotypes of the leaves is described in “Materials and methods”
Allelism testing for bs5 using reference pepper line
To ascertain whether or not the XeR plants were in fact carrying a recessive bs5 allele, allelism tests for bs5 were performed using ECW50R as reference line. In ECW50R XeR is conditioned by a single, homozygous recessive gene bs5 (Vallejos et al. 2010). To this end, ECW50R as paternal parent was crossed with XeR CaT1, CaGl, CaDH269, respectively, as maternal parents. F1 seeds were germinated, grown and tested for XeR after inoculation with Xe78 bacteria. All F1 progenies were Xe resistant, demonstrating that the recessive gene controlling XeR in these plants is allelic to bs5. Genotyping of the bs5 allele in the plant used in this study is shown in Fig. S1.
Construction of pepper BAC library and chromosome walking
A Bacterial Artificial Chromosome (BAC) library was constructed from the XeR pepper line CaDH269. This line developed the same resistance symptoms after Xe78 infection as described above for ECW50R and CaT1. In addition, the allelism test and the phenotype of the self-pollinated progeny demonstrated that this plant also carried a bs5 allele in homozygous configuration. Young leaves from CaDH269 were collected and a BAC library was constructed by BIO SandT Inc. (Montreal, Canada). The bacterial suspension of the BAC library was plated and single colonies were transferred to the cells of 96-well microtiter plates containing 100 μl Nutrient Broth (Sigma-Aldrich Co.) per cell. In total of 380,000 clones were collected and the microtiter plates were arranged in three dimensional array (2 × 3 × 24 plates of 96 wells = 13,824) of which 72 pools were collected (each pool contained 24 × 24 = 576 clones). In this way multiplex PCR could be used to identify single BAC clones with just 72 PCR amplifications and agarose gel-electrophoresis. During chromosomal walking, BAC end specific markers were either used: (i) to isolate overlapping novel BAC clones, (ii) to re-confirm overlapping relationship with the previously identified BAC clones (e.g., primer pairs M_50b4-40 were used to verify the overlap with BAC-326d1, while primer pairs M_50b4-OP were used to identify BAC-137d8, etc.), or (iii) to verify the map position of the bs5 gene (to avoid miss-orientation).
Total DNA isolation, PCR amplification and electrophoresis of amplicons
Total DNA was isolated from approximately 100 mg young leaf tissue using the ZenoGene40 Plant DNA Isolation Kit (ZENON Biotechnology Ltd, Szeged, Hungary) according to the supplier’s instructions. PCR amplification was performed using standard conditions in Taq Buffer (ZENON Biotechnology Ltd, Szeged, Hungary) in the presence of Taq DNA polymerase (Dupla-Taq™, ZENON Biotechnology Ltd, Szeged, Hungary), 0.4 μM primers, 1.5 mM MgCl2 and 5–10 ng total DNA as template. PCR cycles were carried out, following an initial denaturation step for 3 min at 94 °C, at 94 °C for 1 min, annealing at 45–68 °C (depending on the primer annealing temperature) for 1 min, extension at 72 °C for 1 min over 35 cycles and terminated with 7 min incubation at 72 °C. PCR amplification products were separated in 1–3% agarose gels depending on the expected length of the PCR fragment in 1 × Tris acetate/EDTA buffer. Fragments undistinguishable by length in the gel were subjected to SSCP (Single Strand Conformation Polymorphism; Orita et al. 1989) analysis. Buffer, gels and staining were prepared and performed by standard procedures. Primers used throughout this study are listed in Table S1. Primer names have a “Pr_” prefix and an “F” or “R” suffix. Genetic markers are referred to as the name of the primers (without “F/R” suffix) having an “M_” prefix.
Preparation of RNA, reverse transcription and transcriptome analysis by RNA-Seq
Young leaves from sensitive and resistant pepper plants were ground in liquid nitrogen then RNA was extracted using the SV Total RNA isolation kit (Promega Corporation, Madison, USA). To reduce the genomic DNA contamination, on-column DNase treatment was carried out. The purification was accomplished according to the manufacturer's instructions. Integrity of the RNA was checked by the Agilent 2100 Bioanalyzer system (Agilent Santa Clara, CA, USA). For the generation of constructs of Bs5 and bs5, cDNA was synthesized according to the supplier’s instructions using the Maxima Reverse Transcriptase (Thermo Fisher Scientific, Waltham, MA).
Before library preparation for total transcriptome analysis, ribosomal RNAs were removed using the Ribo-Zero rRNA removal kit (Epicentre, Madison, USA). Library preparation and RNA sequencing (RNA-Seq) were performed by using the dedicated kits and the total RNA sequencing was carried out using the SOLiD4 sequencer (Life Technologies, Carlsbad, CA, USA) by the Seqomics Ltd. (Mórahalom, Hungary).
Generating constructs
For reverse complementation experiment, a 3 kb long genomic region with 1 kb native promoter and 440 bps terminator region of Bs5 were amplified using CaFo genomic DNA as template with specific oligonucleotides extended with the sequences of PstI restriction sites at their 5′ ends (Fig. S2). PstI digested PCR fragments were inserted into PstI site of pCAMBIA1303 binary vector. Hygromycin B phosphotransferase gene was exchanged with the aminoglycoside phosphotransferase (NeoR) gene using XhoI restriction sites for generating the same clones with kanamycin resistance.
For GFP-Bs5-fusion constructs, Bs5 coding sequences were amplified from pepper cDNA using Bs5 specific primers containing NheI and PmlI restriction endonuclease sites (Fig. S3). Template cDNAs were prepared from total RNA of Bs5 sensitive CaFo and bs5 resistant CaT1 plants using RevertAid Premium Reverse Transcriptase (Fermentas, Lithuania) with oligo dT20 primer according to the manufacturer's protocol. The amplified Bs5 and bs5 cDNA fragments were cloned into pGEMT Easy vector (Promega Corporation, Madison, USA) and then the NcoI-NheI fragment from pCAMBIA1302 containing the mgfp5 gene was inserted in front of Bs5 variants. NcoI and blunt-ended SalI (filled with Klenow enzyme) fragments containing the GFP-Bs5 and GFP-bs5 fusion proteins were inserted into NcoI and blunt ended BstEII site of the pCAMBIA1302 binary vector.
Transient gene expression in N. benthamiana epidermal cells and microscopy analysis
The coding region of the CaFo-Bs5 and CaT1-bs5 cDNAs were cloned at the C-terminal end of the green fluorescent protein (GFP) of pCambia-1302 (see Fig S3). The resulting plasmids pCambia-1302-GFP-Bs5 and pCambia-1302-GFP-bs5 were introduced from E. coli into Agrobacterium tumefaciens strain C58 by tri-parental mating and were infiltrated into young Nicotiana benthamiana leaves along an E. coli strain expressing P19 RNA silencing suppressor. After 48 h, protoplasts were prepared from the infected area using the protoplast isolation method (Nagy and Maliga 1976) and the subcellular localization of GFP fluorescent signal was investigated by Zeiss (Zeiss LSM 510 META) and Olympus (Olympus Fluoview FV1000 Confocal laser scanning microscope) confocal microscopes.
Transformation of pepper plants
The Hungarian pepper cultivar CaGI carrying the recessive bs5 gene in homozygous configuration was used in transformation experiments to reverse-complement the mutation in the bs5 genes (the bs5, but not the Bs5 allele was present in CaGl as demonstrated by allelism test and by PCR based genotyping Fig. S1). In these reverse-complementation experiments transgenic plants carry an extra copy of the wild type Bs5 gene together with the two copies of the resident bs5 alleles. The ectopic expression of the Bs5 gene in the transgenic plants will supply wild type Bs5 functions consequently it is expected that the resistance trait of CaGl will be reversed to susceptibility since Bs5 gene is dominant over bs5 (see above, the genetics of this trait show, that F1 plants with Bs5/bs5 genetic configurations are susceptible to Xe78). To this end, CaGl leaves (8 leaf state) were treated by At ShooterGRifR [pCambia-1303(HygR)-Bs5], At ShooterGRifR [pCambia-1303(NeoR)-Bs5] and At ShooterGRifR (pCambia-1303) as a control according to the transformation protocol described by Mihalka et al. (2003). For structural and functional map of plasmid constructs (see Fig. S2).
Expression of the Bs5, bs5 and Ca_CYSTM2 genes
Expression of Bs5, bs5, Ca_CYSTM2 and WD40 genes were investigated by whole transcriptome shotgun sequencing (RNA Seq, using SOLiD™ System, http://www.lifetechnologies.com) of RNA isolated from bulked leaf tissue of 8 resistant and 8 susceptible plants of CaFoT1-F2 plants (the same ones used for mapping, see “Materials and methods”). Reads were aligned to the corresponding TC-s. RNA was isolated form Xe resistant and susceptible leaf samples at 0 (samples 53 and 55) and 24 h (samples 54 and 56) post infection, respectively. The number of reads was: 20,365,803; 42,128,939; 27,750,667 and 25,427,924 for samples 53, 54, 55 and 56, respectively. There was at least tenfold difference in expression of Bs5 and Ca_CYSTM2 in leaves (Z. Szabó unpublished observations). In addition to RNA Seq, EST database of NCBI was also used to extract EST sequences from different tissues.
Results
Selecting Xe resistant pepper plant and allelism testing for bs5
In a search for durable XeR pepper plants, several seed accessions from the pepper seed stock collection of The World Vegetable Center Genebank (http://avrdc.org/) were screened in pathogeniticity tests. A single pepper plant, denoted CaT1 exhibited robust resistance following inoculation with Xe78. Inoculation with Xe78 resulted in typical water-soaked lesions on the leaves of XeS plants but XeR pepper leaves showed only slight leaf changes (Fig. 1). In addition, CaT1 plants were also resistant to X. perforans, but were susceptible to X. gardneri (data not shown, see also Sharma et al. 2022). The resistant leaf phenotype of the Xe infected CaT1 plants resembled that of ECW50R (Vallejos et al. 2010), CaDH269 (Csillery et al. 2004; Szarka et al. 2006) and CaGl (https://sites.google.com/site/fpkutato/fajtaink/cseresznyepaprika-fajtk).
To ascertain whether the above plants were carrying bs5 allele in homozygous configuration, F1 allelism tests were performed using ECW50R, a reference plant (a bs5/bs5 homozygote, Jones et al. 2002; Vallejos et al. 2010) as paternal parent, and crossed with CaT1, CaDH269, and CaGl plants, respectively. F1 progeny were tested for Xe phenotype after inoculation with Xe78 bacteria and – as a result—all F1 plants were XeR, demonstrating that the recessive gene controlling Xe resistance in CaT1, CaDH269, and CaGl, respectively, were allelic to bs5. Moreover, the homozygosity of plants CaT1, CaDH269, CaGl and ECW50R were further confirmed for the bs5 locus afterward (Fig. S1) using a bs5-specific molecular marker, M_bs5g developed based on the cloned bs5 gene (see below).
Genetic mapping of the bs5 gene
In order to identify the bs5 gene, map-based cloning was initiated. To establish F2 segregation populations, F1 plants were generated by crossing the XeR CaT1 plant (♀) with the XeS CaFo(♂) and F1 progenies—a total of 46 individuals—were tested for disease symptoms after inoculation of the leaves with Xe78 bacteria. F1 plants and the paternal parent (CaFo) showed the characteristic XeS symptoms of bacterial spot disease (water-soaked lesions, complete necrosis and abscission of infected leaves) while maternal parent (CaT1) was XeR (Fig. 1). Based on these results, we concluded that: (i) similarly to the former observations, the susceptibility to Xe78, conditioned by the wild type Bs5 allele, was a dominant trait over the resistant bs5 allele; (ii) both bs5 and Bs5 were in homozygous configuration in the parental lines; (iii) the F1 plants were derived from crossing and not from self-pollination because no F1 progeny inherited the XeR (maternal) phenotype. The hybrid nature of the F1 plants was also confirmed with the M_472g4_OP molecular marker (Fig. S4). These results were in agreement with the former findings that the resistance to bacterial spot disease was conditioned by the recessive gene bs5 (Jones et al. 2002). To identify the chromosomal location of the bs5 gene, F2 individuals, derived from 46 self-mated F1 plants and termed collectively CaT1Fo F2, were used for genetic mapping. A total of 4662 F2 plants were scored for disease symptoms of which 3516 and 1146 were XeS and XeR, respectively. These figures did not deviate significantly from the expected 3:1 ratio (χ2 = 0.44, p = 0.51), therefore we confirmed the hypothesis that the XeR trait is conditioned by a single recessive gene, bs5 in this diploid pepper population.
Rough genetic mapping was carried out using 8 XeS and 8 XeR selected individuals of the F2 segregation population. Total DNA isolated from fresh leaves were subjected to PCR amplification using previously published eleven genetic markers (Lee et al. 2004). The genetic analysis of these eleven pepper specific Simple Sequence Repeat (SSR) markers positioned the bs5 gene between markers M_CaCY and M_AF244121-A at the upper end of Linkage Group 3 of the genetic map of pepper (Lee et al. 2004). M_CaCY is equivalent to the pepper RFLP marker CaPR10 (Lee et al. 2004) and to the tomato marker M-T1065; (http://www.sgn.cornell.edu/marker/SGN-M1736/details). We extended the mapping population to 4662 F2 individuals to determine the map positions more precisely. The fine mapping revealed that the genetic linkage between marker M_CaCY and bs5 was about 0.20 ± 0,01 centimorgan. Since marker M_AF244121-A was more distant, the marker M_CaCY was a good starting point for chromosome walking toward bs5 (Fig. 2A).

Genetic and physical map around the bs5 locus. A, Primary and overlapping BAC clones of the bs5 contig are colored green. Positions of markers (red characters in italics), used for positioning the resistant phenotype conditioned by the bs5 gene, are shown black arrows; three overlapping BAC clones (50b4, 137d4 and 472g4) make up the bs5 region. B, The chromosome segments of three F2 plant individuals carrying tightly linked recombination breaks around bs5 are illustrated; colors represent combined genotypes of the two sister-chromatids (Kiss et al. 1998); magenta and green represent resistant and susceptible genotypes, respectively. C, gene content within and around the bs5 locus flanked by two genetic markers M_P6 and M_WD40; genes and their relative orientations and intergenic regions are shown by horizontal thick arrows and thin black bars, respectively; length of the genes and intergenic regions are to scale. D, structure of the bs5 gene; the length of the exons (E) and introns (I) are shown in base pairs (bp). E1 and E3 are divided by thin vertical bars into 5′ or 3′ untranslated regions (UTR) and coding regions, respectively; length of the exons and introns are in relative scale. OP and -40 ends of the BAC clones are highlighted by vertical bar and closed circles, respectively
Chromosomal walking identifies candidate gene for Bs5/bs5
The molecular marker M_CaCY was used to isolate the primary BAC clone from the BAC library constructed from pepper CaDH269 plants (see “Materials and methods”). Overlapping BAC clones were isolated using markers generated from the end sequences of the identified BAC clones. These steps were reiterated and resulted in the bs5 contig of about 600-kilobase covering the bs5 region (Fig. 2A). Genetic mapping with markers developed based on BAC end sequences delimited the bs5 gene between markers M_50b4-40 and M_1045d3-OP defining the bs5 region (Fig. 2A). Based on the sequences of BAC clones BAC-50b4 and BAC-1045d3 (overlapping the bs5 region), additional genetic markers were developed and used for genetic mapping that further narrowed down the position of the bs5 gene. This analysis identified three recombinant plants, CaT1Fo-F2-1290, CaT1Fo-F2-2241 and CaT1Fo-F2-3812 (Fig. 2B), which helped to delimit the bs5 gene in the bs5 locus flanked by two markers, M_P6 and M_WD40, respectively (Fig. 2C). The sequence of BAC-50b4 between the markers M_P6 and M_WD40 revealed that the bs5 locus covered 14,654 bp (see partial sequence of BAC-50b4 GenBank accession no.: OM681616). The corresponding Bs5 locus was also sequenced from the XeS parent CaFo resulting in 14,734 bp length sequence (GenBank accession no.: OM681615). To identify the genetic alteration responsible for the resistance trait, the sequences of the Bs5 and bs5 loci, respectively, were aligned and coding regions were compared (Fig. S5). BLASTN search of the pepper specific Expressed Sequence Tags (EST) database of NCBI (http://www.ncbi.nlm.nih.gov/; taxid:4071) revealed two protein coding sequences, Ca_WD40 (encoding a protein with WD40 repeats) and Ca_CYSTM1 (coding for a tail-anchored cystine-rich transmembrane module, CYSTM) genes (Fig. 2C). From the EST sequences of the two genes, TC sequences, Ca_WD40-TC and Ca_CYSTM1-TC, respectively, were generated (Fig. S6) and aligned to the sequence of Bs5 and bs5 locus. Nucleotide polymorphisms were searched for in these two genes to identify mutations distinguishing between the resistant and susceptible alleles. For Ca_WD40 gene, besides several single nucleotide polymorphisms (SNPs) in the non-coding sequence, a single nucleotide change of C to A at position 11 305 was found in the coding sequence between the XeS and XeR alleles, but this alteration did not cause a change of amino acid (Fig. S5). In addition, the analysis of the expression of Ca_WD40 in publicly available RNAseq and EST databases (https://ncbi.nlm.nih.gov/sra/?term=capsicum+annuum+RNA-Seq; https://blast.ncbi.nlm.nih.gov/Blast.cgi) showed similar transcriptional activity, therefore Ca_WD40 was considered unlikely to be responsible for the XeR trait. For the Ca_CYSTM1 gene, several SNPs in the 3′ untranslated region (UTR) and a 6 bp deletion in the third exon were detected between the Bs5 and bs5 allele (Fig. S5). Since the 6 bp deletion occurred in the coding sequence of the XeR allele we suggested that this mutant variant of Ca_CYSTM1, denoted Ca_cystm1 could be a good candidate for the bs5 gene of pepper.
Genetic complementation of the bs5 mutation
In order to prove that Ca_CYSTM1 and Ca_cystm1 corresponded to Bs5 and bs5 genes, genetic complementation was carried out. Because the wild type Bs5 allele—conferring Xe susceptibility—is a dominant character, it seemed reasonable to convert the resistant phenotype to sensitivity by introducing the wild type allele into the XeR pepper line CaGl. The sequence of the Ca_CYSTM1 gene controlled by its native promoter was cloned into pCambia vectors carrying either the hygromycin phosphotransferase resistance (HygR) or the neomycin phosphotransferase resistance (NeoR) marker gene (Fig. S2) and introduced into the bs5/bs5 homozygous CaGl plants using the ‘shooter’ Agrobacterium transformation system (Mihalka et al. 2003). Leaves developed on transformed plants were tested for the presence of isopentenyl transferase (ipt), HygR, NeoR, Ca_CYSTM1 and Ca_cystm1 genes, respectively, as well as for flower development, mature seed formation and Xe susceptibility/resistance (Table S2). Control transformants transformed with empty vector (C1, C2, C3) developed normal roots and could grow to maturity, set seeds and were resistant to Xe (Table S2 and Fig. 3B). Positive transformants (TR116 and TR121 series) carrying the ectopic Ca_CYSTM1 gene could only be maintained and developed to full-grown plants when they were grafted to a non-transformed CaGl rootstock. The Xe78-infected leaves on Ca_CYSTM1 transformants, confirmed for the presence of marker genes as well as Ca_CYSTM1 and Ca_cystm1 alleles,, respectively, showed the characteristic susceptible symptoms, water-soaked lesions and defoliation (Table S2 and Fig. 3A). We found that transgenic plants carrying the ectopic CYSTM1 gene were not healthy, in all cases cork-like outgrowth occurred on the stems (Fig. 4A), tissue necrosis occurred on stems and leaves, mortality was high, and in all but one case the plants were sterile; independent transformants are shown in Fig. 4B–H.

Leaf phenotype of a Ca_CYSTM1 transformed (A) and a control (B) plant after inoculation by Xe78 (inoculation sites are encircled). For TR116.2.82 and C1, see Table S2

Transformed plants. CaGI plants (which are bs5/bs5 homozygous, Xe resistant) were genetically modified by introducing the Bs5 gene under its native promoter. Five HygR (TR116.2.56; TR116.2.82; TR116.4.77; TR116.4.97; TR116.4.103; TR116.4.154) and two NeoR (TR121.2.19; TR121.2.82) independent transformant shoots were selected on agar matrix after transformation with pCYSTM1::CYSTM1 constructs. After grafting (A), plants were transferred to soil in pots and grown in the greenhouse. Cork-like outgrowth is shown by red arrow. Panels from C to H show independent transformed plants. H shows transformant TR116.2.82, which produced fruits and mature seed
From the map-based cloning results and the genetic complementation experiments, we concluded that the 6 bp deletion found in the Ca_CYSTM1 gene was in fact responsible for the XeR trait in pepper, consequently Ca_CYSTM1 and Ca_cystm1 gene corresponded to Bs5 and bs5 genes, respectively. Therefore, the term Bs5 and bs5 will be used hereafter.
Expression of the CYSTM genes in pepper
Expression of Bs5 and bs5 genes was investigated by whole transcriptome shotgun sequencing (RNA Seq) of RNA isolated from leaf tissue of 8 XeR and 8 XeS bulk plants of the CaFoT1-F2 population (the same ones used for preliminary mapping). This analysis revealed that both Bs5 and bs5 genes were expressed in leaf tissue (data not shown). In addition, Bs5 is expressed in other tissues as is supported by EST sequences (Table S3) and RNA Seq data of Sequence Read Archives of NCBI (e.g., SRX15644350-SRX15644367, SRX13447132-SRX13447138, SRX13447140-SRX13447147, SRX9188483-SRX9188486). It is worth mentioning that there is no nucleotide difference between the genomic, EST and RNASeq sequences concerning Bs5 gene despite the sequences originated from different pepper cultivars e.g., (ECW50R, CaT1, CaFo, Ca cv. Bukang, Ca cv. Nokkwang and Ca cv. Hang Keun, respectively).
The pepper genome contains a CYSTM paralog of Bs5
A similarity search using Bs5-TC as query identified an additional homologous gene, Ca_CYSTM2 encoding another CYSTM protein, Ca_CYSTM2 (Fig. S6, Fig. S7, Fig. S8). The Ca_CYSTM2 gene is located on the other side of the bs5 gene compared with Ca_WD40 positioned in opposite orientation indicating the duplication of the CYSTM genes in pepper. ESTs for Ca_CYSTM2 gene were obtained from the pepper EST database (taxid:4072) of NCBI (Table S3) and the alignment of Bs5-TC and Ca_CYSTM2-TC sequences at nucleotide level (Fig. S7) revealed extensive similarity throughout the coding region (256 nucleotides out of 273 were identical; 93.8% homology). It is interesting to note, that, out of the 17 nucleotide changes, seven were non-synonymous and ten were synonymous mutations (Fig. S7) consistent with Ca_CYSTM2 being under strong purifying selection (Lawrie et al. 2013). However, the ~ 300 bp promoter region of Bs5 and Ca_CYSTM2 displayed considerable differences that might indicate distinct transcriptional activity of the two Ca_CYSTM genes.
The bs5 protein is deficient in the predicted transmembrane domain
Bs5 and bs5 genes code for peptides of 92 and 90 residues, respectively (Fig. 5). Similarity search of the predicted peptide sequence indicated that both are tail-anchored cystine-rich transmembrane module (CYSTM) proteins (Venancio et al. 2010). A transmembrane (TM) domain was predicted at the C-terminus of the Bs5 protein (Fig. 6) composed of 20 AA residues indicating membrane localization. The mutant bs5 encodes a shorter version of Bs5 lacking two residues in the predicted TM domain (Fig. 5 and Fig. 6) which probably affects the function of the bs5 peptide. Similarity search at the protein level identified homologous genes in other Solanaceae (Fig.S9) and more distant plant species but even a homologous peptide was found among human translated sequences (Fig. 6).

Alignment of cDNA and deduced protein sequence of Bs5 and bs5 alleles. The sequences of Bs5-mRNA and bs5-mRNA were taken from Fig. S6, and represent coding, 5′- and 3′-UTR sequences of Bs5 and bs5 alleles, respectively. The sequences of Bs5-mRNA and bs5-mRNA were verified also by sequencing both parental genomic DNAs and coding sequences (see text). Except for the six base pair deletion, bs5 has the same amino acid sequence as Bs5. Start and Stop codons are underlined. Deletion is highlighted by red dashes (-). poly(A) represents the polyA tail. Unmatched nucleotides are highlighted with red characters. The primers aligned to the sequence were used to synthetize cDNA. The direction of primers is indicated by black arrowheads and by 5′ marking

Multiple alignment of CYSTM proteins from selected organisms, and secondary structure prediction. Protein sequences were obtained from the protein databanks as described below. Alignment was done manually by the help of aligned coding sequences. TM domains were predicted by the “Split Server Prediction” software: http://split.pmfst.hr/split/4/. The 2 amino acid deletion in bs5 (second row) is highlighted be two dashes. The amino acids were highlighted by colors according to the scheme by Lesk (http://www.bioinformatics.nl/~berndb/aacolour.html) as follows: small nonpolar (orange), G; hydrophobic (green), C, P, Y; polar (magenta), Q; negatively charged (red), D, E; positively charged (blue), K,R. The names of the sequences are labeled as follows: the first four letters denote the abbreviated name of the organism (first letter stands for the first letter of the genus name, the next three are the first three letters of the species name). After this the name of the CYSTM proteins were given. The species abbreviations are: Ca: Capsicum annuum; Nb: Nicotiana benthamiana; Sl: Solanum lycorpersicum; At: Arabidopsis thaliana; Hs: Homo sapiens. The protein sequences were obtained for: Bs5, bs5, Ca_CYSTM2, this study; Slyc_CYSTM1 and Slyc_CYSTM2: solgenomics.net reference no.: Solyc09g098310.3.1 and Solyc09g098300.3.1, respectively; Nb_CYSTM1 and Nb_CYSTM2, see Fig S9, respectively; At_WIH1 and At_WIH2: NCBI Reference Sequence: NP_569052.1 and NP_181673.1, respectively; Hs CYSTM1: https://www.uniprot.org/uniprot/Q9H1C7. Abbreviation of the secondary structure predictions (https://predictprotein.org/): E, extended or other; H, Alpha-helix
The Bs5-GFP fusion protein was localized to the plasma membrane
Tail-Anchored proteins can have a variety of membrane locations (Abell et al. 2003; Venancio and Aravind 2010; Abell and Mullen 2011). To define what membrane the Bs5/bs5 proteins are located in subcellularly, constructs coding for green fluorescent protein (GFP) fused to the N-termini of Bs5 or bs5 peptides were generated (Fig. S3). The constructs were transiently expressed in Nicotiana benthamiana leaves, protoplasts were isolated from the infected tissues and the signal of the GFP fusion proteins of Bs5/bs5 was visualized by confocal laser scanning microscope. High-resolution optical images captured from different depths of the sample demonstrated that N-terminal GFP-Bs5 and GFP-bs5 fusion proteins are targeted into the plasma membrane by slightly different efficiency (Fig. 7, Panel A and Panel B).

Confocal microscopy of N-terminal GFP-Bs5 (A) and GFP-bs5 (B) fusion proteins. For GFP fusion constructs, Bs5 and bs5 coding sequences were amplified from cDNA using specific primers, Pr_Bs5c F7, Pr_Bs5c R7 and Pr_Bs5c F, Pr_bs5c R7T, respectively, containing NheI and PmlI restriction endonuclease sites (Fig. S3) and cloned at the C-terminal end of the green fluorescent protein (GFP) of pCambia-1302. The resulting plasmids pCambia-1302-GFP-Bs5 and pCambia-2302-GFP-bs5 (see legend to Fig S3) were infiltrated into young Nicotiana benthamiana leaves. After 48 h, protoplasts were prepared from the infected area using the protoplast isolation method described by Nagy and Maliga (1976) and the subcellular localization of GFP fluorescence was investigated by Zeiss (Zeiss LSM 510 META) and Olympus (Olympus Fluoview FV1000) Confocal laser scanning microscopes
Discussion
In this paper, we described the map-based cloning of the bs5 gene of pepper which conditions recessive resistance against Xanthomonas euvesicatoria. In pepper plants carrying the bs5 allele in homozygous configuration, the leaf tissue at the site of infection, remains alive and functioning. This phenotype is substantially different from that of the programmed cell death-based hypersensitive response (HR) characterized by tissue necrosis (Klement et al. 1964; Balint-Kurti 2019). In addition, bs5 based resistance is stable; no breakdown has yet been observed. This is an important feature of recessive alleles of susceptibility genes which confer resistance.
Reverse genetic complementation was taken to substantiate bs5 as responsible for resistance. In these complementation experiments bs5 homozygous CaGl plants were transformed with wild the type Bs5 allele. Transformants with ectopic Bs5 displayed abnormal phenotypes in contrast to the control transformants (without the Bs5 gene), which were normal in appearance. The reason for the altered habitus of the Bs5 transformants is not known. One could hypothesize that attenuated function of the Bs5/bs5 proteins might be responsible for the abnormal phenotype as a consequence of gene silencing. RNA interference provoked by the ectopic Bs5 transgene could plausibly lower the expression of the Bs5 and bs5 genes. This same mechanism could also reduce the amount of the CaCYSTM2 gene product, since there is extensive nucleotide sequence similarity between that of Bs5 and CaCYSTM2 genes (Fig S7). The simultaneous silencing of Bs5, bs5 and CaCYSTM2, might be responsible for insufficient function(s) of the corresponding proteins leading to morphological changes. wih1, wih2 and uvi15 all encode mutant CYSTM proteins homologous to Bs5 (Fig. 5; Venancio and Aravind 2010) and a similar stunted character with a high percentage of sterility and abnormal growth was demonstrated for Arabidopsis thaliana plants carrying wih1/wih2 double mutations (Lieber et al. 2011). Likewise, sensitivity to ultraviolet light, defective sporulation, and loss of viability during growth of the Schizosaccharomyces pombe uvi15 mutant has also been described (Lee et al. 1994, 1995). Taking into account the abnormal mutant phenotypes one could conclude, that CYSTM proteins have a more general function than stress tolerance (Lee et al. 1995; Beilharz et al. 2003; Li et al. 2009; Venancio and Aravind 2010; Hillenmeyer et al. 2010; Feng et al. 2011). The available CYSTM null mutants in Arabidopsis wih1/wih2 double mutants (Lieber et al. 2011) and in Schizosaccharomyces pombe uvi15 mutant, as well as the Bs5 transformed pepper lines (see above), were all viable but of decreased fitness indicating that CYSTM proteins are required for normal growth.
DNA sequence analysis of the wild type Bs5 gene revealed that Bs5 has three exons and codes for a short protein of 92 amino acids (Fig. S5, Fig. 6). The Bs5 protein belongs to the tail-anchored (TA), cysteine-rich transmembrane module (CYSTM) protein family (Kutay et al. 1993; Lee et al. 1995; Beilharz et al. 2003; Li et al. 2009; Venancio and Aravind 2010; Abell and Mullen 2011; Feng et al. 2011; Lieber et al. 2011). The bs5 allele has the same exon–intron structure and encodes a similar protein to Bs5 except for the lack of two leucine amino acids in the TM domain (Fig. 5, 6). This shorter protein seems to be responsible for the resistance trait in bs5 pepper lines.
Tail-Anchored proteins are targeted to membranes in such a way that the N-terminal ends face the cytosol (Paul et al. 2013) and the C-terminal part is inserted into the membrane. Taking into consideration this feature, N-terminal GFP-Bs5 protein fusions were generated and used to transiently express these constructs in N. benthamiana (Nb) leaves. Although Nb was a heterologous system, but is in the same family as pepper, we expected that Bs5/bs5 proteins should be targeted into the same membrane in Nb as in Ca, since AAs in the N-terminal and the TM domain direct TA proteins into the membrane system (Borgese et al. 2003). From the alignment of the CYSTM proteins, one can see that the first 24 AAs and the last 16 AAs of Bs5 and NbCYSTM, respectively are identical (Fig. 6). According to the microscopic images, both GFP-Bs5 and GFP-bs5 protein were localized in the cell membrane, which is in agreement with previous publications for the location of CYSTM proteins (Borgese et al. 2003; Abell et al. 2007; Kriechbaumer et al. 2009). On the other hand, GFP-Bs5 and GFB-bs5 proteins, respectively, did not show exactly the same intensity and localization pattern of GFP, since GFP-Bs5 protein localized more or less entirely in the cell membrane while in the case of GFP-bs5 membrane targeting was disturbed to some extent and GFP signals displayed a fainter and more diffuse signal (Fig. 7). Confocal microscopy of the GFP-Bs5-transformed leaves also revealed several dark green spots near and around the membrane, suggestive of some kind of compartmentalization and vesicular trafficking of the GFP-Bs5 protein (Fig. 7A). These dark green spots were almost missing in the case of the GFP-bs5 fusion (Fig. 7B). The lack of the two adjacent leucines in bs5 therefore disturbed the normal membrane targeting process and abolished the dark green spots characteristic of GFP-Bs5. The above findings were in agreement with results described in previous publications showing that CYSTM TA proteins were cell membrane located in a wide range of organisms (Xu et al. 2018; https://www.uniprot.org/uniprot/Q9H1C7).
It is interesting to note, that in spite of high degree of protein sequence similarity (or identity) (Fig. 6) and similar expression in leaves of both CaCYSTM2 and bs5, CaCYSTM2 did not complement bs5 with respect to resistance in bs5 plants (ECW50R, CaT1, CaDH269 and CaGl). Non-complementation might be attributed to the three AAs difference, LLQ in bs5 and FME in CaCYSTM2, respectively at the beginning of the TM domain (Fig. 6). From this, one may conclude that CaCYSTM2 is not a functional paralogue of Bs5/bs5.
The molecular function of the CYSTM proteins is unknown. One can only speculate that the TA proteins should be involved in those biological processes—such as biotic or abiotic stresses, plant development (Lee et al. 1995; Venancio and Aravind 2010; Xu et al. 2018; Lieber et al. 2011)—which one can measure, visually see and/or deduce from the phenotype and properties of the mutant organisms (e.g., phenotypes of A. thaliana wih1/wih2, S. pombe uvi15 and Ca bs5 mutants, respectively). AA sequence resemblance, interaction with other proteins as well as the localization of the CYSTM proteins, however, may leave some more room for cautious speculation. The cytoplasmic domain of Bs5 contains glycine-tyrosine-proline-proline repeats (GYPP-R). These repeats are found in several proteins and represent one class of proline rich proteins which can participate in protein–protein interactions with high specificity (Matsushima et al. 1990; Williamson 1994; Brownawell and Creutz 1997; Kay et al. 2000; Strugnell et al. 2005; Hu et al. 2009; Creutz 2009; Li et al. 2009; Feng et al. 2011). The C-terminal TM domain indicates, and experimental evidence verifies, that CYSTM TA proteins are membrane bound (see also Fig. 7). It is plausible to suppose that CYSTM proteins may interact with other proteins. If identified, interacting partners might shed light on the biological process in which the complex participates (Huang et al. 2013). For example, the human CYSTM1 protein (https://www.uniprot.org/uniprot/Q9H1C7) (and see also Fig. 5) seems to interact with two proteins, BAG3 (https://www.uniprot.org/uniprot/O95817) (a molecular chaperone regulator) and SYT16 (https://www.uniprot.org/uniprot/Q17RD7) (Synaptotagmin-16), which were identified in yeast two-hybrid system. BAG3 may be involved in chaperone binding, while SYT16 participates in vesicular trafficking and exocytosis of secretory vesicles in non-neuronal tissues of mammals (Wolfes and Dean 2020). Besides, the TM domain of the CYSTM proteins adopts an α-helix conformation in the membrane, and as a consequence cysteine residues face in different orientations allowing the establishment of disulfide or hydrogen bonds with residues of neighboring proteins, as was demonstrated for Syntaxin A1, a CYSTM TA protein (https://www.uniprot.org/uniprot/Q16623; Gregoret, et al. 1991; Cohen et al. 2007; Witt 2008; Bachnoff et al. 2013). Syntaxins and annexins function in vesicle fusion process (Brownawell and Creutz 1997; Vardar et al. 2016). As a conclusion, it cannot be excluded, that the Bs5 protein, one way or another, participates in vesicular trafficking (e.g., endocytosis).
It has been demonstrated that during Xe infection effector molecules from Xe enter plant cells and are delivered through translocons which are the pore forming apparatus of the Type Three Secretion System (TTSS) (Kim and Hartmann 1985; Schornack et al. 2008; Kay and Bonas 2009; Büttner and Bonas 2010). Translocons, are formed in the plant cell membrane and built up by proteins encoded by the pathogen (Sory and Cornelis 1994; Jones and Dangl 2006; Block et al. 2008; Büttner and He 2009; Galan et al. 2014). This arrangement necessitates the interaction of proteins from both partners. If a plant protein involved in this interaction is altered by a mutation in such a way that the effector delivery is disturbed, then the plant may become resistant to Xe. It is possible, that the wild type Bs5 protein is involved in the transport of effectors—in one way or another—into the plant cells and its mutant bs5 allele will be non-functional in effector delivery. Hampered effector entry was shown in bs5 plants compared with the susceptible pepper lines (Ortega et al. 2019), therefore we hypothesize that bs5 plants are resistant against Xe due to the lack of entry of Xe effectors into the host cell. Nevertheless, it is plausible to suppose that resistance to Xe may be generated by inducing deletions in orthologous CYSTM genes using genome editing approach (Osakabe and Osakabe 2015). The same 6 bp deletions could result in Xe resistant plants in those plant species, e.g., in tomato (Solanum lycopersicum), which are closely related to pepper and in which no durable natural resistance exists against Xanthomonas euvesicatoria.
Data availability
All data relevant to this study have been provided in the manuscript, or in the supplemental online materials. Sequence data of the Bs5 and bs5 loci described in this article can be found in the GenBank/EMBL sequence collection under the accession numbers OM681616 and OM681615, respectively. All plant materials in this study are available for research purposes from the corresponding author by request.
Change history
08 May 2023
A Correction to this paper has been published: https://doi.org/10.1007/s00122-023-04366-2
References
Abell BM, Jung M, Oliver JD, Knight BC, Tyedmers J, Zimmermann R, High S (2003) Tail-anchored and signal-anchored proteins utilize overlapping pathways during membrane insertion. J Biol Chem 278:5669–5678. https://doi.org/10.1074/jbc.M209968200
Abell BM, Rabu C, Leznicki P, Young JC, High S (2007) Post-translational integration of tail-anchored proteins is facilitate by defined molecular chaperons. J Cell Science 120:1743–1751. https://doi.org/10.1242/jcs.002410
Abell BM, Mullen RT (2011) Tail-anchored membrane proteins: exploring the complex diversity of tail-anchored-protein targeting in plant cells. Plant Cell Rep 30:137–151. https://doi.org/10.1007/s00299-010-0925-6
Afzal I, Shinwaria ZK, Sikandar S, Shahzadc S (2019) Plant beneficial endophytic bacteria: mechanisms, diversity, host range and genetic determinants. Microbiol Res 221:36–49. https://doi.org/10.1016/j.micres.2019.02.001
Bachnoff N, Cohen-Kutner M, Trus M, Atlas D (2013) Intra-membrane signaling between the voltage-gated Ca2+-channel and cysteine residues of syntaxin 1A coordinates synchronous release. Sci Rep 3:1620. https://doi.org/10.1038/srep01620
Balint-Kurti P (2019) The plant hypersensitive response: concepts, control and consequences. Mol Plant Pathol 20:1163–1178. https://doi.org/10.1111/mpp.12821
Beilharz T, Egan B, Silver PA, Hofmann K, Lithgow T (2003) Bipartite signals mediate subcellular targeting of tail-anchored membrane proteins in Saccharomyces cerevisiae. J Biol Chem 278:8219–8223. https://doi.org/10.1074/jbc.M212725200
Block A, Li G, Fu ZQ, Alfano JR (2008) Phytopathogen type III effector weaponry and their plant targets. Curr Opin Plant Biol 11:396–403. https://doi.org/10.1016/j.pbi.2008.06.007
Boller T, Felix GA (2009) Renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu Rev Plant Biol 60:379–406. https://doi.org/10.1146/annurev.arplant.57.032905.105346
Borgese N, Colombo S, Pedrazzini E (2003) The tale of tail-anchored proteins: cpmiong from the cytosol and looking for a membrane. J Cell Biol 161:1013–1019. https://doi.org/10.1083/jcb.200303069
Brownawell AM, Creutz CE (1997) Calcium-dependent binding of sorcin to the N-terminal domain of synexin (Annexin VII). J Biol Chem 272:22182–22190. https://doi.org/10.1074/jbc.272.35.22182
Büttner D, He SY (2009) Type III protein secretion in plant pathogenic bacteria. Plant Physiol 150:1656–1664. https://doi.org/10.1104/pp.109.139089
Büttner D, Bonas U (2010) Regulation and secretion of Xanthomonas virulence factors. FEMS Microbiol Rev 34:107–133. https://doi.org/10.1111/j.1574-6976.2009.00192
Calle García J, Guadagno A, Paytuvi-Gallart A, Saera-Vila A, Amoroso CG, D’Esposito D, Andolfo G, Aiese Cigliano R, Sanseverino W, Ercolano MR (2022) PRGdb 4.0: an updated database dedicated to genes involved in plant disease resistance process. Nucleic Acids Res 50:D1483–D1490. https://doi.org/10.1093/nar/gkab1087
Cohen R, Marom M, Atlas D (2007) Depolarization-evoked secretion requires two vicinal transmembrane cysteines of syntaxin 1A. PLoS ONE 2:e1273. https://doi.org/10.1371/journal.pone.0001273
Creutz CE (2009) Novel protein ligands of the annexin A7 N-terminal region suggest pro-beta helices engage one another with high specificity. Gen Physiol Biophys 28:F7–F13
Csillery G, Szarka S, Sardi E, Mityko J, Kapitany J, Nagy B, Szarka J (2004) The unity of plant defense. In: Proceedings of the 12th Eucarpia meeting on genetics and breeding of capsicum and eggplant, Noordwijkerhout, Netherlands pp 147–153
Ditta G, Stanfield S, Corbin D, Helinski D (1980) Broad host range DNA cloning system for gram-negative bacteria: construction of a gene bank of Rhizobium meliloti. Proc Natl Acad Sci USA 77:7347–7351. https://doi.org/10.1073/pnas.77.12.7347
Dixon MS, Golstein C, Thomas CM, van der Biezen EA, Jones JDG (2000) Genetic complexity of pathogen perception by plants: the example ofRcr3, a tomato gene requiredspecifically by Cf-2. Proc Natl Acad Sci USA 97:8807–8814. https://doi.org/10.1073/pnas.97.16.8807
Dodds P, Rathjen J (2010) Plant immunity: towards an integrated view of plant–pathogen interactions. Nat Rev Genet 11:539–548. https://doi.org/10.1038/nrg2812
Dolatabadian A (2020) Plant-microbe interaction. Biology 10(1):15. https://doi.org/10.3390/biology10010015
Flor HH (1971) Current Status of the Gene-for-Gene Concept. Annu Rev Phytopathol 9(275):296. https://doi.org/10.1146/annurev.py.09.090171.001423
Feng Y, Peng H, Liang S (2011) Molecular analysis of the PGYRP (proline-, glycine- and tyrosine-rich protein) gene family in soybean. Mol Biol Rep 38:2739–2750. https://doi.org/10.1007/s11033-010-0419-1
Galan JE, Lara-Tejero M, Marlovits TC, Wagner S (2014) Bacterial type III secretion systems: specialized nanomachines for protein delivery into target cells. Annu Rev Microbiol 68:415–438. https://doi.org/10.1146/annurev-micro-092412-155725
Garcia-Ruiz H, Szurek B, Van den Ackerveken G (2021) Stop helping pathogens: engineering plant susceptibility genes for durable resistance. Curr Opin Biotechnol 70:187–219. https://doi.org/10.1016/j.copbio.2021.05.005
Gregoret LM, Rader SD, Fletterick RJ, Cohen FE (1991) Hydrogen bonds involving sulfur atoms in proteins. Proteins 9:99–107. https://doi.org/10.1002/prot.340090204
Harris JM, Balint-Kurti P, Bede JC, Day B, Gold S, Goss EM, Grenville-Briggs LJ, Jones KM, Wang A, Wang Y, Mitra RM, Hoon Sohn K, Alvarez ME (2020) What are the top 10 unanswered questions in molecularplant-microbe interactions? MPMI 33:1354–1365. https://doi.org/10.1094/MPMI-08-20-0229-CR
Heath MC (2000) Nonhost resistance and nonspecific plant defenses. Curr Opin Plant Biol 3:315–319. https://doi.org/10.1016/s1369-5266(00)00087-x
Higgins VJ, Lu H, Xing T, Gelli A, Blumwald E (1998) The gene-for-gene concept and beyond: Interactions and signals. Can J Plant Path 20:150–157. https://doi.org/10.1080/07060669809500420
Hillenmeyer ME, Ericson E, Davis RW, Nislow C, Koller D, Giaever G (2010) Systematic analysis of genome-wide fitness data in yeast reveals novel gene function and drug action. Genome Biol 11:R30. https://doi.org/10.1186/gb-2010-11-3-r30
Huang X, Dai FF, Gaisano G, Giglou K, Han J, Zhang M, Kittanakom S, Wong V, Wei L, Showalter AD, Sloop KW, Stagljar I, Wheeler MB (2013) The identification of novel proteins that interact with the GLP-1 receptor and restrain its activity. Mol Endocrinol 27:1550–1563. https://doi.org/10.1210/me.2013-1047
Ingle RA, Carstens M, Denby KJ (2006) PAMP recognition and the plant−pathogen arms race. BioEssays 28:880–889. https://doi.org/10.1002/bies.20457
Jones JB, Minsavage GV, Roberts PD, Johnson RR, Kousik CS, Subramanian S, Stall RE (2002) A non-hypersensitive resistance in pepper to the bacterial spot pathogen is associated with two recessive genes. Phytopathology 92:273–277. https://doi.org/10.1094/PHYTO.2002.92.3.273
Jones JB, George H, Lacy GH, Bouzar H, Stall RE, Schaad NW (2006) Reclassification of the Xanthomonads associated with bacterial spot disease of tomato and pepper. Syst Appl Microbiol 27:755–762. https://doi.org/10.1078/0723202042369884
Jones JDG, Dangl JL (2006) The plant immune system. Nature 444:323–329. https://doi.org/10.1038/nature05286
Kay BK, Williamson MP, Sudol M (2000) The importance of being proline: the interaction of proline-rich motifs in signaling proteins with their cognate domains. FASEB J 14:231–241
Kay S, Bonas U (2009) How Xanthomonas type III effectors manipulate the host plant. Curr Opin Microbiol 12:37–43. https://doi.org/10.1016/j.mib.2008.12.006
Keen NT (1990) Gene-for-gene complementarity in plant-pathogen interactions. Annu Rev Genet 24:447–463. https://doi.org/10.1146/annurev.ge.24.120190.002311
Kiss GB, Kereszt A, Kiss P, Endre G (1998) Colormapping: a non-mathematical procedure for genetic mapping. Acta Biol Hung 49:47–64
Kim BS, Hartmann RW (1985) Inheritance of a gene (Bs3) conferring hypersensitive reistance to Xanthomonas campestris pv. vesicatoria in pepper (Capsicum annuum). Plant Dis 69:233–235
Kriechbaumer V, Shaw R, Mukherjee J, Bowsher CG, Harrison A-M, Abell BM (2009) Subcellular distribution of tail-anchored proteins in Arabidopsis. Traffic 10:1753–1764. https://doi.org/10.1111/j.1600-0854.2009.00991.x
Klement Z, Farkas GL, Lovrekovich L (1964) Hypersensitive reaction induced by phytopathogenic bacteria in the tobacco leaf. Phytopathology 54:474–477
Kourelis J, van der Hoorn RAL (2018) Defended to the nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell 30:285–299. https://doi.org/10.1105/tpc.17.00579
Kurowski C, Conn K Himmel P (2010) Guidline for identification of pepper bacterial leaf spot races using differential hosts. In: APS-ISF collaboration to implement a system to standardize identification of plant pathogen race and strains based on host differentials, pp 1–5
Kusch S, Panstruga R (2017) mlo-Based resistance: an apparently universal “Weapon” to defeat powdery mildew disease. Mol Plant Microbe Interact 30:179–189. https://doi.org/10.1094/MPMI-12-16-0255-CR
Kutay U, Hartmann E, Rapoport TA (1993) A class of membrane proteins with a C-terminal anchor. Trends Cell Biol 3:72–75. https://doi.org/10.1016/0962-8924(93)90066-a
Lawrie DS, Messer PW, Hershberg R, Petrov DA (2013) Strong purifying selection at synonymous sites in D. melanogaster. PLoS Genet 9:1003527. https://doi.org/10.1371/journal.pgen.1003527
Lee JK, Park EJ, Chung HK, Hong SH, Joe CO, Park SD (1994) Isolation of UV-inducible transcripts from Schizosaccharomyces pombe. Biochem Biophys Res Commun 202:1113–1119. https://doi.org/10.1006/bbrc.1994.2043
Lee JK, Kim M, Choe J, Seong RH, Hong SH, Park SD (1995) Characterization of uvi15+, a stress-inducible gene from Schizosaccharomyces pombe. Mol Gen Genet 246:663–670. https://doi.org/10.1007/BF00290711
Lee JM, Nahm SH, Kim YM, Kim BD (2004) Characterization and molecular genetic mapping of microsatellite loci in pepper. Theor Appl Genet 108:619–627. https://doi.org/10.1007/s00122-003-1467-x
Li H, Yang J, Wang Y, Chen Z, Tu S, Feng L, Zhu Y, Li Y (2009) Expression of a novel OSPGYRP (rice proline-, glycine- and tyrosine-rich protein) gene, which is involved in vesicle trafficking, enhanced cold tolerance in E. coli. Biotechnol Lett 31:905–910
Lieber D, Lora J, Schrempp S, Lenhard M, Laux T (2011) Arabidopsis WIH1 and WIH2 genes act in the transition from ic to reproductive cell fate. Curr Biol 21:1009–1017. https://doi.org/10.1016/j.cub.2011.05.015
Matsushima N, Kretsinger CCE, RH, (1990) Polyproline, beta-turn helices. Novel secondary structures proposed for the tandem repeats within rhodopsin, synaptophysin, synexin, gliadin, RNA polymerase II, hordein, and gluten. Proteins 7:125–155. https://doi.org/10.1002/prot.340070204
Mihalka V, Balazs E, Nagy I (2003) Binary transformation systems based on “shooter” mutants of Agrobacterium tumefaciens: a simple, efficient and universal gene transfer technology that permits marker gene elimination. Plant Cell Rep 21:778–784. https://doi.org/10.1007/s00299-003-0597-6
Nagy JI, Maliga P (1976) Callus induction and plant regeneration from mesophyll protoplasts of Nicotiana sylvestris. Z Pflanzenphysiol 78:453–455
Orita M, Iwahana H, Kanazawa H, Hayashi K, Sekiya T (1989) Detection of polymorphisms of human DNA by gel electrophoresis as single-strand conformation polymorphisms. Proc Natl Acad Sci USA 86:2766–2770. https://doi.org/10.1073/pnas.86.8.2766
Ortega A, Schultik A, Pham J, Cho M-J, Doudna J, Staskawicz B (2019) Cas9 genome editing to establish bacterial spot resistance in tomato. IS-MPMI XVIII Congress, Abstracts of Poster Presentations, July 14-18, Glasgow Scotland
Osakabe Y, Osakabe K (2015) Genome editing with engineered nucleases in plants. Plant Cell Physiol 56:389–400
Palloix A, Ayme V, Moury B (2009) Durability of plant major resistance genes to pathogens depends on the genetic background, experimental evidence and consequences for breeding strategies. New Phytol 183:190–199. https://doi.org/10.1111/j.1469-8137.2009.02827.x
Panstruga R, Moscou MJ (2020) What is the molecular basis of nonhost resistance? MPMI 33:1253–1264. https://doi.org/10.1094/MPMI-06-20-0161-CR
Paul P, Simm S, Blaumeiser A, Scharf KD, Fragkostefanakis S, Mirus O, Schleiff E (2013) The protein translocation systems in plants - composition and variability on the example of Solanum lycopersicum. BMC Genomics 14:189. https://doi.org/10.1186/1471-2164-14-189
Potnis N, Minsavage G, Smith JK, Hurlbert JC, Norman D, Rodrigues R, Stall RE, Jones JB (2012) Avirulence proteins AvrBs7 from Xanthomonas gardneri and AvrBs1.1 from Xanthomonas euvesicatoria contribute to a novel gene-for-gene interaction in pepper. Mol Plant Microbe Interact 25:307–320. https://doi.org/10.1094/MPMI-08-11-0205
Quackenbush J, Liang F, Holt I, Pertea G, Upton J (2000) The TIGR gene indices: reconstruction and representation of expressed gene sequences. Nucleic Acids Res 28:141–145. https://doi.org/10.1093/nar/28.1.141
Ronald PC, Staskawicz BJ (1988) The avirulence gene avrBs1 from Xanthomonas campestris pv. vesicatoria encodes a 50-kD protein. Mol Plant Microbe Interact 1:191–198
Römer P, Hahn S, Jordan T, Strauss T, Bonas U, Lahaye T (2007) Plant pathogen recognition mediated by promoter activation of the pepper Bs3 resistance gene. Science 318:645–648. https://doi.org/10.1126/science.1144958
Schmitt-Keichinger C (2019) Manipulating cellular factors to combat viruses: a case study from the plant eukaryotic translation initiation factors eIF4. Front Microbiol 10:17. https://doi.org/10.3389/fmicb.2019.00017
Schornack S, Minsavage GV, Stall RE, Jones JB, Lahaye T (2008) Characterization of AvrHah1, a novel AvrBs3-like effector from Xanthomonas gardneri with virulence and avirulence activity. New Phytol 179:546–556. https://doi.org/10.1111/j.1469-8137.2008.02487.x
Sharma A, Minsavage GV, Gill U, Hutton S, Jones JB (2022) Identification and mapping of bs8, a novel locus conferring resistance to bacterial spot caused by Xanthomonas gardneri. Phytopathology. https://doi.org/10.1094/PHYTO-08-21-0339-R
Sory MP, Cornelis GR (1994) Translocation of a hybrid YopE-adenylate cyclase from Yersinia enterocolitica into HeLa cells. Mol Microbiol 14:583–594. https://doi.org/10.1111/j.1365-2958.1994.tb02191.x
Stall RE, Jones JB, Minsavage GV (2009) Durability of resistance in tomato and pepper to xanthomonads causing bacterial spot. Annu Rev Phytopathol 47:265–284. https://doi.org/10.1146/annurev-phyto-080508-081752
Strauß T, van Poecke RMP, Strauß A, Römer P, Minsavage GV, Singh S, Wolf C, Strauß A, Kim S, Lee H-A, Yeom S-I, Parniske M, Stall RE, Jones JB, Choi D, Prins M, Thomas Lahaye T (2012) RNA-seq pinpoints a Xanthomonas TAL-effector activated resistance gene in a large-crop genome. Proc Natl Acad Sci USA 109:19480–19485. https://doi.org/10.1073/pnas.1212415109
Strugnell JM, Norman MD, Jackson JA, Drummond AJ, Cooper A (2005) A Molecular phylogeny of the coleoid cephalopods (Mollusca: Cephalopoda) using a multigene approach; the effect of data partitioning on resolving phylogenies in a Bayesian framework. Mol Phylogenet Evol 37:426–441. https://doi.org/10.1016/j.ympev.2005.03.020
Szarka E (2008) Biochemical and genetic examinations of the general defense system of plants, PhD thesis, Department of Genetics and Plant Breeding, Corvinus University of Budapes, Hungary
Szarka J, Csilléry G (1995) Defense systems against Xanthomonas campestris pv. veicatoria in pepper. In: Proceeding of 9th Eucarpia meeting on genetics and breeding of Capsicum and Eggplant, Budapest, Hungary, 21–26 Aug, pp 184–187
Szarka J, Toldi O, Szarka E, Remenyik J, Csilléry G (2006) General defense reaction in the plant kingdom. Acta Agron Hung 54:221–232. https://doi.org/10.1556/AAgr.54.2006.2.9
Tai TH, Dahlbeck D, Clark ET, Gajiwala P, Pasion R, Whalen MC, Stall RE, Staskawicz BJ (1999) Expression of the Bs2 pepper gene confers resistance to bacterial spot disease in tomato. Proc Natl Acad Sci USA 96:14153–14158. https://doi.org/10.1073/pnas.96.24.14153
Vallejos CE, Jones V, Stal RE, Jones JB, Minsavage GV, Schultz DC, Rodrigues R, Olsen LE, Mazourek M (2010) Characterization of two recessive genes controlling resistance to all races of bacterial spot in peppers. Theor Appl Genet 121:37–46. https://doi.org/10.1007/s00122-010-1289-6
Van der Biezen EA, Jones JD (1998) Plant disease-resistance proteins and the gene-for-gene concept. Trends Biochem Sci 23:454–456. https://doi.org/10.1016/s0968-0004(98)01311-5
Vardar G, Chang S, Arancillo M, Wu YJ, Trimbuch T, Rosenmund C (2016) Distinct functions of syntaxin-1 in neuronal maintenance, synaptic vesicle docking, and fusion in mouse. Neurons J Neurosci 36:7911–7924. https://doi.org/10.1523/JNEUROSCI.1314-16.2016
Venancio TM, Aravind L (2010) CYSTM, a novel cysteine-rich transmembrane module with a role in stress tolerance across eukaryotes. Bioinformatics 26:149–152. https://doi.org/10.1093/bioinformatics/btp647
Wang Y, Rory N, Pruitt RN, Nürnberger T, Wang Y (2022) Evasion of plant immunity by microbial pathogens. Nat Rev Microbiol. https://doi.org/10.1038/s41579-022-00710-3
Wille L, Messmer MM, Studer B, Hohmann P (2019) Insights to plant-microbe interactions provide opportunities to improve resistance breeding against root diseases in grain legumes. Plant Cell Environ 42:20–40. https://doi.org/10.1111/pce.13214
Williamson MP (1994) The structure and function of proline-rich regions in proteins. Biochem J 297:249–260. https://doi.org/10.1042/bj2970249
Witt D (2008) Recent developments in disulfide bond formation. Synthesis 16:2491–2509. https://doi.org/10.1055/s-2008-1067188
Wolfes AC, Dean C (2020) The diversity of synaptotagmin isoforms. Curr Opin Neurobiol 63:198–209. https://doi.org/10.1016/j.conb.2020.04.006
Xu Y, Yu Z, Zhang D, Huang J, Wu C, Yang G, Yan K, Zhang S, Zheng C (2018) CYSTM, a novel non-secreted cysteine-rich peptide family, iInvolved in environmental stresses in Arabidopsis thaliana. Plant Cell Physiol 59:423–438. https://doi.org/10.1093/pcp/pcx202
Yuan M, Chu Z, Li X, Xu C, Wang S (2009) Pathogen-induced expressional loss of function is the key factor in race-specific bacterial resistance conferred by a recessive R gene xa13 in rice. Plant Cell Physiol 50:947–955. https://doi.org/10.1093/pcp/pcp046
Acknowledgements
We thank Dr. T.H.N. Ellis, Dr. P. Salamon, G. Csilléry and R.D. Farkas for their help during the work and critical reading of the manuscript, A. Ferhan for his help in the confocal microscopy, C. Iliescu, M. Szivós, K. Miró, I. Harsányi, A. Bukovinszky, T. Földi for technical assistance.
Funding
Open access funding provided by Hungarian University of Agriculture and Life Sciences. This work was supported by ABC-MATE, Godollo, Hungary; the Jedlik Ányos Research Fund, Grant No. NKFP-4-2005-0015; the OTKA, Grants No.: NK-43167 and K-111935; the ABSTRESS FP7-KBBE-2011–5, Project No.: 289562 and the LEGATO FP7-KBBE.2013.1.2-02, Project No.: 613551 of the European Union.
Author information
Authors and Affiliations
Contributions
ZSZ and MB: performed the genetical mapping; ZSZ, MCS and AD: participated in the transformation experiment; ZSZ, AD and GYBK: participated in the RNA seq experiment; ZSZ, AD and PK: participated in the transient expression and confocal microscopy. ZSZ and GYBK: wrote the paper.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Yiqun Weng.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
122_2023_4340_MOESM1_ESM.tif
Supplementary file 1: Fig. S1 Genotypes of the plants used in allelism tests Marker M_bs5g were used to genotype the plants for Bs5/bs5 alleles. Total DNA isolated from leaves of the plants was used as templates. After PCR amplification using Pr_bs5g F1 and Pr_bs5g R1, fragments were separated in agarose gel, and visualized bands were converted to genotypes. Upper and lower fragments were amplified from Bs5 and bs5 alleles, respectively. In heterozygotes where the two alleles were amplified the heteroduplex fragments run slower, therefore the upper band is fuzzy. The resistant and susceptible phenotypes of the self-pollinated progenies of the above plants verified the heterozygous and homozygous genetic configuration (data not shown). The genotypes of the plants for Bs5/bS5 alleles using M_bs5g marker are as follows: ECW50R, bs5/bs5; T1xECW50R F1, bs5/bs5; CaT1, bs5/bs5; T1xFo F1, Bs5/bs5; CaFo, Bs5/Bs5; CaDH269 (bs5/bs5); CaGl (bs5/bs5), ECW, (Bs5/Bs5).
122_2023_4340_MOESM2_ESM.tif
Supplementary file 2: Fig. S2. Structural and functional map of Bs5 genomic constructs for Agrobacterium mediated transformation. Abreviations: HygR, hygromycin phosphotransferase gene; NeoR, neomycin phosphotransferase gene; UTR, Untranslated region. GUS, β-glucuronidase gene, GFP, Green Fluorescence Protein gene, Nos, Nopaline synthase terminator, 35S, CAMV 35S gene, term, transcription terminator. Arrows indicate the direction of transcription, RB and LB, Right and Left border sequence, respectively
122_2023_4340_MOESM3_ESM.pdf
Supplementary file 3: Fig. S3 N-terminal GFP-Bs5, GFP-bs5 constructs used for confocal microscopy. The double stranded cDNA sequence of Bs5 and bs5 was amplified from single stranded cDNA synthetized from polyA+ RNA isolated from young leaves of CaFo and CaT1 parental plants, respectively. For amplification, primer pairs Pr_Bs5c F7, Pr_Bs5c R7, and Pr_Bs5c F7, Pr_bs5 R7T, respectively, were used. Isolation of polyA + RNA and cDNA synthesis were carried out with Promega SV total RNA Kit and RevertAid First Strand cDNA Synthesis Kit. Double stranded cDNA sequences of CaFo-Bs5 and CaT1-bs5 were cloned into pGem-T Easy vector. From these constructs, CaFo Bs5 and CaT1 bs5 cDNA sequences were cloned in frame after the structural gene of green fluorescent protein (GFP) of pCambia-2302 vector using NheI, SalI and NheI, BstEII restriction enzymes (SalI and BstEII sticky ends were made blunt ended by Mung Bean nuclease. Panel A and B shows the functional and relevant restriction map of pCambia-2302 CaFo Bs5 and pCambia-2302 CaT1 bs5, respectively. The nucleotide and amino acid (AA) sequences of the junctions at the GFP – Bs5, and GFP—bs5 genes/proteins are shown in Panel C and D, respectively. Primer sequences are underlined.
122_2023_4340_MOESM4_ESM.tif
Supplementary file 4: Fig. S4 Demonstration of the hybrid nature of the F1 plants from cross CaT1 (♀) x CaFo (♂) using molecular marker M_472g4-OP. Fragment length polymorphism was detected after PCR amplification and agarose gel electrophoresis. Total DNA isolated from leaves of the F1 plants were amplified with primers Pr_472g4-OP F and Pr_472g4-OP R. Amplification products were separated in 3% agarose gel, and visualized bands were converted to genotypes. Upper and lower fragments were amplified from Bs5 and bs5 alleles, respectively. MM, 100 bp molecular ladder (Fermentas). F1 hybrid plants are numbered from 1- 46. CaT1, Ca var. T1; CaFo, Ca cv. Feherozon
122_2023_4340_MOESM5_ESM.pdf
Supplementary file 5: Fig. S5 Alignment of the nucleotide sequence of the Bs5 and bs5 loci. The sequence of the bs5 locus was determined by sequencing EcoRI and HindIII subclones as well as by high throughput SOLiD™ sequencing of BAC-50b4. The wild type Bs5 locus was sequenced from amplified genomic DNA of plant CaFo using specific primer pairs. These primer pairs were designed in such a way that the amplified fragments were overlapped with each other. The sequences of the bs5 and the Bs5 loci were deposited to the GenBank (GenBank accession no.: OM681616; GenBank accession no.: OM681615, respectively). The sequence of Bs5-TC, CaWD40-TC and primers for markers M_P6, M_WD40, M_bs5g, M_Bs5g were also aligned to the genomic sequence. CaBs5-TC and CaWD40-TC sequences were presented by red characters. Start and Stop codons were underlined. 5′-UTR/3′-UTR and coding regions were highlighted by lower and upper case letters, respectively. Introns and missing nucleotides (deletions) are indicated by dashes. The direction of primers is indicated by black arrows and by marking the 5′ end. Nucleotide alterations and indels in the sequence of bs5 gene were highlighted by turquoise-blue background color. poly(A), Polyadenylation tail
122_2023_4340_MOESM6_ESM.pdf
Supplementary file 6: Fig. S6 Tentative consensus sequence of the CYSTM and WD40 genes in pepper. The > 14 kb partial genomic sequence of BAC-50b4 between the flanking genetic markers (M_P6, M_WD40) was used to search homologous sequences in the NCBI “Expressed Sequence Tags” (EST) databases using BLASTN option (http://www.ncbi.nlm.nih.gov/). EST sequences of the five genes were recruited from the Capsicum annuum (taxid:4072) EST database of NCBI (http://www.ncbi.nlm.nih.gov/) using the Bs5 region of BAC-50b4 (see Fig. 1). The individual ESTs (see Table S3) were aligned (using the SeqMan program of the DNA STAR Software; http://www.dnastar.com/), and the consensus sequence generated by the program was taken as the Tentative Consensus (TC; Quackenbush et al. 2000) sequences for Bs5, Bs5-TC (Panel A), for WD40, CaWD40-TC (Panel B) and for Ca_CYSTM2-TC (Panel C). No nucleotide difference was found between the genomic, CYSTM1-TC and CYSTM1-mRNA sequences despite the genomic and EST sequences originated from different pepper cultivars (CaFo, Ca cv. Bukang, Ca cv. Nokkwang and Ca cv. Hang Keun, respectively). Aligning CYSTM1-mRNA and cystm1-mRNA revealed the 6 bp difference between the two sequences
122_2023_4340_MOESM7_ESM.pdf
Supplementary file 7: Fig. S7 Alignment of Ca_CYSTM2-TC and Bs5-TC. The sequences were taken from Fig. S3, and represents coding, 5′- and 3′-UTR sequences, respectively. Single letter coded amino acids (AA) were deduced from Ca_CYSTM2-TC sequence. Start and Stop codons were underlined. 5′-UTR/3′-UTR and coding regions were highlighted by upper and lower case letters, respectively. Unmatched nucleotides are highlighted with red characters. Nucleotide changes in the coding region which were in the third position of the codon and resulted in no AA change were underlined. The first and second AA in a pair marked deduced AAs from the Ca_CYSTM2-TC and Bs5-TC sequence, respectively. Amino acid similarity highlighted in the paired AA by color: hydrophobic, green; small nonpolar, orange; polar, magenta
122_2023_4340_MOESM8_ESM.pdf
Supplementary file 8: Fig. S8 Alignment of the promoter and 5′ UTR region of Bs5/bs5 and Ca_CYSTM2 genes. More than 300 bp sequence upstream of the start codon was aligned by the help of dot plot matrix based homology search. Sequences before the start codon are in lower case letters. The length of the 5′ UTR sequences were defined according to the longest cDNA at the 5′ end. 5′ UTR sequences are in italics. Identical nucleotides are shown by black characters, mismatch nucleotides are in red. Bs5 and bs5 sequences were 100% identical in this region and displayed as Bs5/bs5. “pr” after the name of the genes stands for promoter
122_2023_4340_MOESM9_ESM.pdf
Supplementary file 9: Fig. S9. The coding and protein sequence of the CYSTM genes of Nicotiana benthamiana. The coding sequences of Nb_CYSTM1 and Nb_CYSTM2 were retrieved from the SolGene database (https://solgenomics.net/) contigs Niben101Scf02563Ctg059 and Niben101Scf02915Ctg016/Niben101Scf02915Ctg017, respectively. The coding sequence was translated to obtain the deduced protein sequence
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Szabó, Z., Balogh, M., Domonkos, Á. et al. The bs5 allele of the susceptibility gene Bs5 of pepper (Capsicum annuum L.) encoding a natural deletion variant of a CYSTM protein conditions resistance to bacterial spot disease caused by Xanthomonas species. Theor Appl Genet 136, 64 (2023). https://doi.org/10.1007/s00122-023-04340-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00122-023-04340-y