
Abstract
Climate change is altering interactions among plants and pollinators. In alpine ecosystems, where snowmelt timing is a key driver of phenology, earlier snowmelt may generate shifts in plant and pollinator phenology that vary across the landscape, potentially disrupting interactions. Here we ask how experimental advancement of snowmelt timing in a topographically heterogeneous alpine-subalpine landscape impacts flowering, insect pollinator visitation, and pathways connecting key predictors of plant-pollinator interaction. Snowmelt was advanced by an average of 13.5 days in three sites via the application of black sand over snow in manipulated plots, which were paired with control plots. For each forb species, we documented flowering onset and counted flowers throughout the season. We also performed pollinator observations to measure visitation rates. The majority (79.3%) of flower visits were made by dipteran insects. We found that plants flowered earlier in advanced snowmelt plots, with the largest advances in later-flowering species, but flowering duration and visitation rate did not differ between advanced snowmelt and control plots. Using piecewise structural equation models, we assessed the interactive effects of topography on snowmelt timing, flowering phenology, floral abundance, and pollinator visitation. We found that these factors interacted to predict visitation rate in control plots. However, in plots with experimentally advanced snowmelt, none of these predictors explained a significant amount of variation in visitation rate, indicating that different predictors are needed to understand the processes that directly influence pollinator visitation to flowers under future climate conditions. Our findings demonstrate that climate change-induced early snowmelt may fundamentally disrupt the predictive relationships among abiotic and biotic drivers of plant-pollinator interactions in subalpine-alpine environments.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Plant-pollinator mutualisms are crucial for the persistence of most flowering plants and many insects. These mutualisms can be disrupted by climate change-driven shifts in the timing of life history events, such as flowering and emergence, that result in phenological mismatches between plants and insect pollinators (Kudo and Ida 2013; Kudo and Cooper 2019). For example, climate change can alter the cues triggering spring flowering and insect emergence differently, and/or plants and insect pollinators can differ in their responses to changing climatic cues (Kudo and Cooper 2019; Stemkovski et al. 2020). Despite the fact that plant-pollinator interactions in alpine ecosystems may be particularly vulnerable to climate change (Inouye 2020), few studies of plant and pollinator phenological responses to climate change have been conducted in these systems. In the alpine, air temperature and the timing of snowmelt are critical cues shaping flowering onset and bee emergence (Totland and Alatalo 2002; Stemkovski et al. 2020). In addition to decreased winter snowpack and increased spring temperatures associated with climate change, earlier snowmelt is driven by the deposition of anthropogenic dust on the surface of snow, which decreases albedo (Deems et al. 2013). Earlier snowmelt timing can cause alpine plant species to emerge from winter dormancy and to flower earlier, influencing pollinator visitation rates (Totland 1993).
Advanced snowmelt is likely to have direct impacts on both plants and insect pollinators at the same time it increases the risk of phenological mismatch. In some montane systems, earlier-flowering species have longer flowering durations, which can increase reproductive output (Pardee et al. 2019). However, advanced flowering may have direct, negative impacts on plant reproduction by reducing flowering durations for some functional groups, such as succulents and cushion plants (Prevéy et al. 2019; Jabis et al. 2020). Early flowering can also expose plants to late spring and early summer frost events, causing buds and flowers to be damaged by frost and reducing reproductive success (Inouye 2008; Pardee et al. 2019). Similarly, harsh, early spring alpine conditions can affect foraging success of insect pollinators. Although some bumble bees and moths have adaptations that permit flying and foraging under low air temperatures and strong winds, such conditions may preclude other species from visiting flowers early in the season (Pyke et al. 2011). Additionally, advanced-flowering plants may experience altered soil moisture patterns, which could affect floral cues and pollinator attraction (Gezon et al. 2016). For example, advanced snowmelt and drought have been shown to decrease flower size and nectar production (Powers et al. 2022). Thus, even if plants and insects overlap spatially and temporally under advanced snowmelt conditions, changes in floral traits and insect behavior may lead to a decoupling of plant-pollinator interactions.
The impacts of snowmelt shifting earlier as a result of anthropogenic change in the alpine will play out across a backdrop of complex topography. Topographic heterogeneity is likely to influence smaller-scale patterns of snowmelt timing across alpine landscapes, contributing to variation in plant and pollinator phenological responses (Bueno de Mesquita et al. 2018; Inouye 2020). Topography modulates soil moisture (Litaor et al. 2008), and topography-driven variation in soil moisture can, in turn, affect plant phenology and floral traits (Suárez et al. 2011). In the context of plant-pollinator interactions, microhabitats that become snow-free at different times confer phenological heterogeneity across the alpine landscape, which could decrease the risk of phenological mismatch between plants and pollinators at the community level (Graae et al. 2018). Topographic complexity, in combination with differential sensitivity to snowmelt timing among plant species, could lead to co-flowering of novel assemblages of plants, altering competition and pollination success (Sargent and Ackerly 2008; CaraDonna et al. 2014). Topography is therefore critical to assessing how climate change will affect plant-pollinator interactions in alpine ecosystems.
In this study, we used a large-scale experimental manipulation of snowmelt timing in a topographically complex alpine environment to investigate how advanced snowmelt affected plant-pollinator interactions. In particular, we asked how experimental advancement of snowmelt timing affected (1) flowering onset and duration, (2) floral abundance, and (3) pollinator visitation rate. We also asked how these responses varied with topographic variation that contributes to natural variation in snowmelt timing, using structural equation modeling to examine interacting factors shaping snowmelt timing, flowering phenology, and pollinator visitation rate. We predicted that topography would influence snowmelt timing, which subsequently would influence flowering phenology and floral abundance, and floral abundance would influence pollinator visitation rate. We further predicted that the strength of the relationships among these interacting factors would differ for plots with advanced snowmelt and control plots.
Methods
Study system
This work was performed in 2020 at Niwot Ridge (40.05411, -105.5891), located in the Colorado Rocky Mountains, USA. Four sites (named Audubon, Lefty, East Knoll, Trough) were used within this study to represent subalpine and alpine environments ranging from 3380 to 3500 m in elevation, dominated by low-growing forbs. Each of our four sites was within an array of five sites that were part of an “early spring” experiment done as part of the Niwot Ridge Long-Term Ecological Research Program, wherein 40 m × 10 m control plots were paired with advanced plots of the same size. In advanced plots, a thin layer of inert black sand was spread on the surface of the snow in late spring, prior to snowmelt at a rate of 227 kg of sand per 40 m x 10 m plot. Sand was spread in control plots following snowmelt to control for any effects of the sand on the structure or temperature of the soil surface. Previous work has demonstrated that this sand has little effect on soil microbes. Black sand has also been used to effectively advance snowmelt timing in a high-elevation forest (Blankinship et al. 2018), and, similarly, locally-sourced dust has been used to advance snowmelt in the alpine tundra (Steltzer et al. 2009). This procedure was initiated in 2018, and repeated in the summers of 2019 and 2020 such that 2020 data collection occurred during the third year of black sand application.
We established five subplots in each of the two plots (control and advanced) at each of three sites (Audubon, East Knoll, Trough) and six subplots in each of the two plots at one site (Lefty), for a total of 21 control subplots and 21 advanced subplots (Fig. S1). These subplots (2 m × 1 m) were paired between control and advanced plots such that each pair had a similar elevation (mean difference ± SD in elevation between pairs = 0.20 ± 0.61 m), and were arranged to capture topographic heterogeneity within each plot. The principal measure of topography used was topographic position index (TPI). TPI compares the elevation of a central point to points around it within a given radius or neighborhood; positive TPI values indicate a peak or ridge occurs at that point in the landscape at a given scale, while negative values indicate that a valley occurs around that point in the landscape at a given scale (Oldfather et al. 2016). The TPI of the subplots at a 15-m radius ranged from − 1.11 to 0.68. To minimize the impacts of any runoff snowmelt into our plots from adjacent areas, which could have impacted soil moisture, we established subplots at least 1 m away from the perimeter of plots.
Data collection
To assess whether the black-sand treatment influenced flowering phenology of forbs, we counted the total number of flowers in subplots twice per week beginning with flowering onset and ending when flowering ceased. A total of 50 forb and 1 woody species were present across all sites (Table S1). Many of the forbs in this system produce flowers that senesce within 3–4 d, and those that have longer floral longevity (e.g., Geum rossii (Rosaceae)) occur at similar densities in subplots across treatments. From our flower count data, we created a metric of floral abundance by taking the cumulative sum of flowers across all flowering species at all time points per subplot.
To assess pollinator visitation to flowers, we performed 15-min observations of the entire flowering community twice per week in subplots where flowers were present, which represented approximately ten weeks of the summer in each site. We conducted observations only when cloud cover was less than 50% and winds were under 24 km per h. We recorded the identities of insects and plants upon observing contact of insects with anthers or stigmas and collected the first two insect visitors of each morphospecies per observation period for identification (Table S1, Table S2). To ensure equal sampling effort, the total amount of observation time per treatment was approximately the same at 3,615 min in control plots and 3,690 min in advanced plots across all sites. Within each site, we counted flowers in subplots of each treatment the same number of times across the season and therefore used the total cumulative visits to flowers across the entire flowering season at the subplot level as our metric of pollinator visitation rate.
To assess the topography of each subplot, we used a digital elevation model (DEM) for Niwot Ridge from OpenTopography (Anderson et al. 2013). Using the R (R Core Team 2022) package raster and function terrain (Hijmans 2023), we calculated elevation, aspect, and slope. We also calculated TPI at a 1.5-m and 15-m radius to represent topography (Oldfather et al. 2016).
Data analysis
To explore the overall impacts of the black-sand treatment on the timing of snowmelt, we fit a linear mixed-effects model (LMM) for each site using snowmelt timing as the response variable, treatment as the predictor, and subplot identity (with subplots paired by elevation) as a random effect.
To explore how snowmelt timing influenced flowering onset, we fit a LMM with flowering onset of individual plant species in advanced snowmelt subplots as our response variable, flowering onset of individual plant species in control subplots as our predictor variable, and plant species identity as a random effect. We tested the normality of residuals using the R package “DHARMa” (Hartig 2022) and found that they were normally distributed. We then determined whether this linear fit differed from a null expectation of no difference in flowering onset between treatments (i.e., deviated from a 1:1 line) using the package “emmeans” (Lenth 2023). Finally, we calculated the difference in flowering onset for plants in advanced vs. control subplots, then fitted a LMM with the difference as the response variable, flowering onset in control subplots as the predictor variable, and plant species as a random effect. We used the R package “lme4” (Bates et al. 2015) for all LMMs.
To examine the influence of treatment on floral abundance, flowering duration, and pollinator visitation rate, we fit separate LMMs with each response variable, treatment as the predictor variable, and paired subplots nested in sites as a random effect. Flowering duration was calculated as the difference in days between the first and last day that a subplot had flowers of any summer-flowering species. We tested the normality of residuals using the R package “DHARMa” (Hartig 2022) and found that they were normally distributed.
Finally, to examine the relationships among topography, snowmelt timing, flowering phenology, floral abundance, and pollinator visitation rate, we constructed piecewise structural equation models (pSEMs) with the R packages vegan and piecewiseSEM (Lefcheck 2016; Oksanen et al. 2022). The base pSEM model was created using inferences derived from the literature, exploratory analyses, and variation inflation factor (VIF) analyses. Predictor variables were selected based on covariance, which was determined using Pearson correlation with R package Hmisc, and then assessed using VIF analyses (Harrell 2023; Zuur et al. 2009). We excluded predictors that led to a VIF > 2.0 to reduce the effect of collinearity (Berglund et al. 2013) and performed model selection by choosing models with the lowest Akaike Information Criterion using the R package “AIC” (Shipley 2013). Variables that did not lead to VIF scores over 2.0 were added based on tests of directed separation (Lefcheck 2016). We tested the normality of residuals of all linear models using the R package “DHARMa” (Hartig 2022).
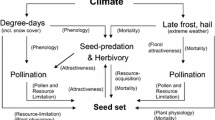
The base pSEM model (Fig. 1) predicted that, at the subplot level, TPI at a 15-m radius impacted the timing of snowmelt, that snowmelt timing influenced flowering onset, that flowering onset influenced flowering duration and the day of peak pollinator visitation rate to flowers, and that floral abundance influenced pollinator visitation rate. Site was used as a random effect in each individual linear model. To compare model structure between treatments, we created two separate models using the same base pSEM model: one with data from control subplots, and one with data from advanced snowmelt subplots. In both models, we used gamma distributions with a log link function for models that had temporal measurements as the response variable, which included models predicting snowmelt timing, flowering onset, and day of peak pollinator visitation rate (Bolker 2008). We found that residuals of three of twelve total models were marginally significantly non-normal. In the control subplot pSEM, models with non-normal residuals were those explaining flowering onset and flowering duration (p = 0.03 and p = 0.02, respectively). In the advanced subplot pSEM, the only model with non-normal residuals was the model explaining flowering onset (p = 0.01). We re-fitted these models with subsets of the data to achieve normality and found that the qualitative results did not change. We report the results from the models with the complete datasets.

The base piecewise structural equation model (pSEM) used to construct pSEMs for both control and advanced snowmelt plots. The model predicts that topographic position index (TPI) at a radius of 15 m impacts snowmelt timing, that snowmelt timing influences flowering onset, that flowering onset influences flowering duration and the day of peak pollinator visitation rate, that flowering duration influences floral abundance, and that floral abundance influences the pollinator visitation rate across the entire flowering duration
Results
Black-sand treatment advanced snowmelt in plots in three sites (Audubon, Lefty, and Trough) by 11.8, 13.0, and 16.8 days, respectively, but there was no significant effect of treatment in site East Knoll (Table 1, Fig. S2). Because snowmelt timing was not affected by the addition of black sand in this site, and because forbs began flowering in this site before we started to monitor subplots for flowering, we excluded this site in all subsequent data analysis. Thus, analyses of flowering onset, floral abundance, flowering duration, and pollinator visitation rate, as well as all pSEMs, use data from three sites (Audubon, Lefty, Trough) comprising 16 paired subplots in six plots.
Observations of flower visitation were conducted twice per week for approximately ten weeks for a total of 7,305 min across all sites, during which we observed a total of 2,589 insect-flower interactions and collected 347 flower-visiting insects representing 103 morphospecies, 36 families, and 6 orders (Table S2). Across all sites, Diptera represented 79.3% of visitors, Hymenoptera represented 15.5% of visitors, Hemiptera represented 3.2% of visitors, Lepidoptera represented 1.7% of visitors, Coleoptera represented 0.3% of visitors, and Orthoptera represented 0.1% of visitors. In the sites used in the construction of pSEMs (all those except East Knoll), Diptera represented 82.9% of visitors, Hymenoptera represented 11.8% of visitors, Hemiptera represented 3.5% of visitors, Lepidoptera represented 1.8% of visitors, and Coleoptera represented 0.1% of visitors. Across sites, pollinators visited a total of 37 different plant species in 16 families (Table S1). Sites and the subplots within them supported different species of flowering plants, driven largely by the topographic heterogeneity of the landscape and its effects on snowmelt and soil moisture (Litaor 2008). Among the 4 sites, East Knoll melted out and flowered first (melting out on May 12 and flowering on May 23 in the control treatment in 2020) and represents a dry meadow community. Audubon and Lefty melted out and flowered later (melting out June 1 and June 3, and flowering on June 17 and 24, respectively, in the control treatment in 2020) and primarily represent dry and moist meadow communities. Trough melted out and flowered latest (melting out on June 29 and flowering on July 8 in the control treatment in 2020) and primarily contains snowbed and moist meadow communities. At all sites, subplots lower in elevation melted out and flowered 7 to 33 days later than those higher in elevation and thus tend to represent snowbed and moist meadow communities. At East Knoll, the most visited plant was Geum rossii (31.4% of visits), and the second most visited plant was Arenaria fendleri (Caryophyllaceae, 16.6% of visits). At Audubon, the most visited plant was also G. rossii (72.0% of visits), and the second most visited plant was Potentilla diversifolia (Rosaceae, 6.5% of visits). At Lefty, the most visited plant was again G. rossii (32.1% of visits), and the second most visited plant was Solidago simplex (Asteraceae, 25.5% of visits). At Trough, the most visited plant was Ligusticum tenuifolium (Apiaceae, 41.2% of visits), and the second most visited plant was G. rossii (33.7% of visits).
Flowering onset of plants in control subplots was a significant predictor of flowering onset in advanced subplots (Fig. 2a), with an estimated coefficient that differed significantly from 1 (the null expectation; LMM [estimate ± SE]: 0.73 ± 0.05, t111 = 13.54, p < 0.001; Fig. 2a). Further, the difference in flowering onset for plants in advanced vs. control subplots was positively correlated with flowering onset (LMM: 0.27 ± 0.05, t111 = 4.89, p < 0.001; Fig. 2b).

Flowering onset in control plots predicts flowering onset in response to advanced snowmelt. (A) Regression shown in red, and null expectation of no difference in flowering onset between treatments (1:1 line) shown in black. (B) Difference in flowering onset for plants in advanced vs. control subplots, with a regression line shown in black
Black-sand treatment had a significant, positive effect on floral abundance, but did not significantly influence flowering duration or pollinator visitation rate (Table 1).
We obtained good fits for pSEMs for both the control (Fisher’s C 34.19, AIC = -121.05, p = 0.08, df = 24; Fig. 3a, Table S3) and advanced subplots (Fisher’s C 29.05, AIC = -91.09, p = 0.41, df = 28; Fig. 3b, Table S3). In both pSEMs, TPI at 15 m was significantly negatively correlated with snowmelt timing, snowmelt timing was significantly positively correlated with flowering onset, and TPI at 15 m was significantly positively correlated with floral abundance, indicating that snow melted later in depressions and that floral abundance was lower on ridges. However, these three paths are the only significant interactions shared by both models. The pSEM for control subplots showed a high degree of agreement with our base model: flowering onset had a significant negative effect on flowering duration, flowering duration had a significant positive effect on floral abundance, and floral abundance had a significant positive effect on pollinator visitation rate. Further, flowering onset and duration had significant positive effects on the day of peak pollinator visitation rate, and the day of peak pollinator visitation rate had a significant positive effect on pollinator visitation rate (Figs. 1 and 3a, Table S3). In contrast, the pSEM for advanced subplots showed few significant paths, and there were no significant predictors of flowering duration, day of peak pollinator visitation rate, or pollinator visitation rate (Fig. 3b, Table S3).

The final piecewise structural equation models for (A) control and (B) advanced snowmelt subplots. Both models had an acceptable fit (p > 0.05) and each individual regression used site as a random effect. Green arrows indicate a significant, positive effect; yellow arrows indicate a significant, negative effect; and gray arrows indicate a non-significant interaction. Values adjacent to arrows represent the estimated strength of the relationship, and R2 values indicate the proportion of variance explained by the model for the associated response variable
Discussion
Our large-scale manipulation of snowmelt timing in an alpine ecosystem demonstrated that advanced snowmelt is associated with higher floral abundance and earlier flowering onset, particularly for species that flower later in the season, and that novel predictors are needed to understand the processes that directly influence pollinator visitation to flowers under climate change. Thus, despite the fact that flowering duration and pollinator visitation rate did not differ between advanced snowmelt and control plots, the relationships between abiotic and biotic drivers of visitation were altered with earlier snowmelt, reducing our ability to identify the mechanisms by which climate change will affect plant and pollinator communities.
As expected, flowering onset was earlier in plots with advanced snowmelt (Fig. 2). Indeed, our pSEMs for both control and advanced snowmelt plots showed that flowering onset was positively correlated with snowmelt timing (Fig. 3). However, when snowmelt was advanced, earlier-flowering species showed a smaller advance in flowering onset than did later-flowering species (Fig. 2a). Thus, the magnitude of advance in flowering onset in advanced snowmelt plots was positively related to flowering onset in control plots (Fig. 2b).
Experiments in both alpine and subalpine ecosystems have tended to show the opposite, with early-flowering species experiencing the greatest phenological advancement in response to experimentally advanced snowmelt (Dunne et al. 2003; Petraglia et al. 2014). However, a meta-analysis revealed that the phenologies of late-flowering species in the coldest tundra sites were most strongly impacted by warming (Prevéy et al. 2019). The ability to grow and flower rapidly in response to snowmelt may be an important adaptation in late-melting habitats because of the abbreviated growing season (Totland and Alatalo 2002). Therefore, snowmelt – and associated cues such as light availability and soil moisture – may cue the initiation of plant growth and thus drive flowering onset in mid- and late-season flowering species. Early-flowering species in both alpine and montane systems, on the other hand, may face harsher conditions if they flower early in response to advanced snowmelt, resulting in negative fitness consequences (Inouye 2008; Prevéy et al. 2019). Thus, late-flowering species may respond to cues such as the amount of solar energy, growing degree days, and number of frost days more strongly than early-flowering species (Bienau et al. 2015).
Given our finding that plants that flower later in the season showed greater advances in flowering onset, the assemblage of flowering species available to pollinators at a given time may change under climatic conditions that result in earlier snowmelt. For example, Forrest et al. (2010) showed that snowmelt timing in subalpine meadows altered the flowering times and synchrony of historically co-flowering species that share pollinators. Under future climates, the flowering phenologies of species that flower in early- and mid-season may be more synchronous in our study sites, altering competitive and facilitative interactions (Sargent and Ackerly 2008). Indeed, because co-flowering synchrony can affect plant-pollinator interactions (Kraft and Ackerly 2014), shifts in the relative timing of flowering at the community level will likely affect plant reproduction. Further, plots with advanced snowmelt had significantly higher floral abundance than control plots. In contrast, in a sub-alpine, montane habitat, Delphinium nuttallianum (Ranunculaceae) produced fewer total flowers in years with low snow accumulation, likely because flowers were exposed to colder temperatures (Inouye and McGuire 1991). It appears that advanced snowmelt did not incur damage to flowers in our study system, and the positive effect of experimentally advanced snowmelt on floral abundance may have been driven by longer flowering duration, though the effect of treatment on flowering duration was not statistically significant. Additionally, because many alpine plants exhibit developmental preformation, or the initiation of organs at least one growing season before maturation (Diggle 1997; Meloche and Diggle 2001), the two previous years of black sand application, advanced snowmelt, and artificially-lengthened growing season may have cumulatively increased floral abundance (Prather et al. 2023). Indeed, inflorescences of Geum rossii, which received 46.0% of all pollinator visits, require 3 years from initiation to maturity (Meloche and Diggle 2001). If advanced snowmelt differentially increases floral abundances among species, thereby changing relative floral abundances, competition among plants for pollination services may change.
Although our treatment did not generate differences in flowering duration or pollinator visitation rate, advanced snowmelt altered the interrelationships between flowering onset, duration, and abundance and pollinator visitation and phenology (Fig. 3). In control plots, later flowering onset was associated with shorter flowering duration. This relationship between flowering onset and flowering duration was absent in plots with experimentally advanced snowmelt, and studies in other alpine and subalpine ecosystems have similarly found that flowering duration was not affected by treatments combining advanced snowmelt and warming (Semenchuk et al. 2016; Jabis et al. 2020). Similarly, in control plots only, flowering duration was significantly positively correlated with floral abundance in subplots, and later flowering onset was associated with later dates of peak visitation. Finally, the strongest predictor of pollinator visitation rate to control subplots was floral abundance. In contrast, pollinator visitation rate was unpredictable in advanced snowmelt plots. Given a wealth of evidence that floral abundance can influence pollinator visitation (e.g., Eckhart et al. 2006; Shibata and Kudo 2020), the lack of relationship between floral abundance and pollinator visitation rate in advanced snowmelt plots suggests that factors other than those examined here could play a role in driving plant-pollinator interactions under future climates. For example, harsh weather in the early spring may prevent certain pollinator taxa from visiting flowers but favor other taxa (Pyke et al. 2011). Further, advanced snowmelt may alter floral signals and rewards by changing soil moisture, thereby changing insect pollinator behavior (Gezon et al. 2016; Powers et al. 2022). Alternatively, advanced snowmelt may result in decreased phenological synchrony between plants and pollinators (Kudo and Ida 2013; Kudo and Cooper 2019). However, in some systems, the phenological advancement of bees and plants in response to global temperature increases has occurred at a similar pace, indicating that certain taxa may be less likely to become phenologically mismatched (Bartomeus et al. 2011).
The spatial extent of our study enabled us to ask whether topography interacted with the black-sand treatment to influence snowmelt timing. Topographic position index (TPI) at 15 m was positively correlated with later snowmelt in both control and advanced snowmelt plots (Fig. 3), corroborating research showing that topography influences snow depth and melt timing (Bueno de Mesquita et al. 2018; Östman 2018). Valleys likely accumulate more snow and may thus have higher soil moisture, influencing flowering phenology (Bueno de Mesquita et al. 2018). Our findings add further evidence that topographic heterogeneity influences the hydrology of alpine ecosystems (Grünewald et al. 2013). Improving our understanding of how topography influences snow accumulation and plant phenology is critical to predicting how climate change will impact alpine communities.
In isolating the effects of advanced snowmelt across a large area, our study did not attempt to incorporate multiple aspects of climate change. In particular, we note that our study did not address increases in temperature that are predicted to occur with climate change (Diaz et al. 2003). Warming impacts vegetative and reproductive phenology differently (Collins et al. 2021) and shortens flowering duration in tundra systems (Prevéy et al. 2019), and direct warming would likely have further accelerated snowmelt and altered our findings. In addition, we focused here on community-level visitation rates, rather than examining whether the composition of insect visitors differed between control and advanced snowmelt plots. Given that shifts in flowering phenology are known to alter the relative frequencies of interaction with different pollinators, affecting reproductive output (Rafferty and Ives 2012), such analyses would be valuable. Finally, we found that the site with earliest snowmelt experienced no effect of black-sand treatment on snowmelt timing, possibly due to its low total snow accumulation, suggesting that points on the landscape with less snow may respond differently to warming.
This study is the first to demonstrate that experimental advancement of snowmelt can fundamentally change the relationships among the biotic factors that influence plant-pollinator interactions. We provide evidence that under novel climate conditions, models fail to explain the rate of pollinator visitation, indicating that different variables, such as pollinator behavior and phenology, may be needed to predict plant-pollinator interaction rates in future climates. Further, our manipulation of snowmelt timing is, to our knowledge, the largest in extent that has been executed in an alpine system (Livensperger et al. 2006; Steltzer et al. 2009; Wipf et al. 2009; Cornelius et al. 2013; Petraglia et al. 2014; Sherwood et al. 2017; Frei and Henry 2021). By experimentally advancing snowmelt across this landscape, we gained novel insight into how climate change will differentially affect flowering phenology and alter mechanisms driving plant-pollinator interactions.
Data availability
Data associated with this study is available via the Environmental Data Initiative (Rose-Person et al. 2023). https://doi.org/10.6073/pasta/920fe947b69eea415bdca0af3b3e33c9.
Reference
Anderson R (2013) Lidar Data, Niwot Ridge Long-Term Ecological Research Site, Colorado. Distributed by OpenTopography. https://doi.org/10.5069/G9FJ2DQM. Accessed: 2021-09-18
Bartomeus I, Ascher JS, Wagner D, Danforth BN, Colla S, Kornbluth S, Winfree R (2011) Climate-associated phenological advances in bee pollinators and bee-pollinated plants. PNAS 108(51):20645–20649. https://doi.org/10.1073/pnas.1115559108
Bates D, Maechler M, Bolker B, Walker S (2015) Ime4: Fitting Linear Mixed-Effects Models Using Lme4. https://arxiv.org/abs/1406.5823
Berglund E, Lytsy P, Westerling R (2013) Adherence to and beliefs in lipid-lowering medical treatments: a structural equation modeling approach including the necessity-concern framework. Patient Educ Couns 91(1):105–112. https://doi.org/10.1016/j.pec.2012.11.001
Bienau MJ, Kröncke M, Eiserhardt WL, Otte A, Graae BJ, Hagen D, Eckstein RL (2015) Synchronous flowering despite differences in snowmelt timing among habitats of Empetrum hermaphroditum. Acta Oecol 69:129–136. https://doi.org/10.1016/j.actao.2015.10.005
Blankinship JC, McCorkle EP, Meadows MW, Hart SC (2018) Quantifying the legacy of snowmelt timing on soil greenhouse gas emissions in a seasonally dry montane forest. Glob Change Biol 24(12):5933–5947. https://doi.org/10.1111/gcb.14471
Bolker BM (2008) Ecological models and data in R. Princeton University Press, New Jersey
Bueno de Mesquita CP, Tillmann LS, Bernard CD, Rosemond KC, Molotch NP, Suding KN (2018) Topographic heterogeneity explains patterns of vegetation response to climate change. Arct Antarct Alp Res 50(1):e1504492. https://doi.org/10.1080/15230430.2018.1504492
CaraDonna PJ, Iler AM, Inouye DW (2014) Shifts in flowering phenology reshape a subalpine plant community. PNAS 111(13):4916–4921. https://doi.org/10.1073/pnas.1323073111
Collins CG, Elmendorf SC, Hollister RD, Henry GH, Clark K, Bjorkman AD, Suding KN (2021) Experimental warming differentially affects vegetative and reproductive phenology of tundra plants. Nat Commun 12(1):3442. https://doi.org/10.1038/s41467-021-23841-2
R Core Team (2023) _R: A Language and Environment for Statistical Computing_. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Cornelius C, Leingärtner A, Hoiss B, Krauss J, Steffan-Dewenter I, Menzel A (2013) Phenological response of grassland species to manipulative snowmelt and drought along an altitudinal gradient. JXB 64(1):241–251. https://doi.org/10.1093/jxb/ers321
Deems JS, Painter TH, Barsugli JJ, Belnap J, Udall B (2013) Combined impacts of current and future dust deposition and regional warming on Colorado River Basin snow dynamics and hydrology. Hydrol Earth Syst Sci 17(11):4401–4413. https://doi.org/10.5194/hess-17-4401-2013
Diaz HF, Eischeid JK, Duncan C, Bradley RS (2003) Variability of freezing levels, melting season indicators, and snow cover for selected high-elevation and continental regions in the last 50 years. In: Diaz HF (ed) Climate Variability and Change in High Elevation Regions: Past, Present & Future, Adv Glob Chang Res 15:33–52. https://doi.org/10.1007/978-94-015-1252-7_3
Diggle PK (1997) Extreme preformation in alpine Polygonum viviparum: an architectural and developmental analysis. Am J Bot 84(2):154–169. https://doi.org/10.2307/2446077
Dunne JA, Harte J, Taylor KJ (2003) Subalpine meadow flowering phenology responses to climate change: integrating experimental and gradient methods. Ecol Monogr 73(1):69–86. https://doi.org/10.1890/0012-9615(2003)073[0069:SMFPRT]2.0.CO;2
Eckhart VM, Rushing NS, Hart GM, Hansen JD (2006) Frequency–dependent pollinator foraging in polymorphic Clarkia xantiana ssp. xantiana populations: implications for flower colour evolution and pollinator interactions. Oikos 112(2):311–327. https://doi.org/10.1111/j.0030-1299.2006.14289.x
Forrest J, Inouye DW, Thomson JD (2010) Flowering phenology in subalpine meadows: does climate variation influence community co-flowering patterns? J Ecol 91(2):431–440. https://doi.org/10.1890/09-0099.1
Frei ER, Henry GH (2021) Long-term effects of snowmelt timing and climate warming on phenology, growth, and reproductive effort of Arctic tundra plant species. Arct Sci 8(3):700–721. https://doi.org/10.1139/as-2021-0028
Gezon ZJ, Inouye DW, Irwin RE (2016) Phenological change in a spring ephemeral: implications for pollination and plant reproduction. Glob Change Biol 22(5):1779–1793. https://doi.org/10.1111/gcb.13209
Graae BJ, Vandvik V, Armbruster WS, Eiserhardt WL, Svenning JC, Hylander K, Lenoir J (2018) Stay or go–how topographic complexity influences alpine plant population and community responses to climate change. Perspect Plant Ecol Evol Syst 30:41–50. https://doi.org/10.1016/j.ppees.2017.09.008
Grünewald T, Stötter J, Pomeroy JW, Dadic R, Moreno Baños I, Marturià J, Lehning M (2013) Statistical modelling of the snow depth distribution in open alpine terrain. Hydrol Earth Syst Sci 17(8):3005–3021. https://doi.org/10.5194/hess-17-3005-2013
Harrell F Jr (2023) Hmisc: Harrell Miscellaneous. R package version 4.8-0. https://CRAN.R-project.org/package=Hmisc
Hartig F (2022) _DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models_. R package version 0.4.6, https://CRAN.R-project.org/package=DHARMa
Hijmans R (2023) _raster: Geographic Data Analysis and Modeling_. R package version 3.6–23. https://CRAN.R-project.org/package=raster
Inouye DW (2008) Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Ecology 89(2):353–362. https://doi.org/10.1890/06-2128.1
Inouye DW (2020) Effects of climate change on alpine plants and their pollinators. Ann NY Acad Sci 1469(1):26–37. https://doi.org/10.1111/nyas.14104
Inouye DW, McGuire AD (1991) Effects of snowpack on timing and abundance of flowering in Delphinium nelsonii (Ranunculaceae): implications for climate change. Am J Bot 78(7):997–1001. https://doi.org/10.1002/j.1537-2197.1991.tb14504.x
Jabis MD, Winkler DE, Kueppers LM (2020) Warming acts through earlier snowmelt to advance but not extend alpine community flowering. J Ecol 101(9):e03108. https://doi.org/10.1002/ecy.3108
Kraft NJ, Ackerly DD (2014) Assembly of plant communities. Ecol Lett 8:67–88
Kudo G, Cooper EJ (2019) When spring ephemerals fail to meet pollinators: mechanism of phenological mismatch and its impact on plant reproduction. Proc R Soc B: Biol Sci 286(1904):20190573. https://doi.org/10.1098/rspb.2019.0573
Kudo G, Ida TY (2013) Early onset of spring increases the phenological mismatch between plants and pollinators. J Ecol 94(10):2311–2320. https://doi.org/10.1890/12-2003.1
Lefcheck JS (2016) piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol Evol 7(5):573–579. https://doi.org/10.1111/2041-210X.12512
Lenth R (2023) emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.8.8. https://CRAN.R-project.org/package=emmeans
Litaor MI, Williams M, Seastedt TR (2008) Topographic controls on snow distribution, soil moisture, and species diversity of herbaceous alpine vegetation, Niwot Ridge, Colorado. J Geophys Res Biogeosci 113. https://doi.org/10.1029/2007JG000419
Livensperger C, Steltzer H, Darrouzet-Nardi A, Sullivan PF, Wallenstein M, Weintraub MN (2016) Earlier snowmelt and warming lead to earlier but not necessarily more plant growth. Ann Bot Plants 8. https://doi.org/10.1093/aobpla/plw021
Meloche CG, Diggle PK (2001) Preformation, architectural complexity, and developmental flexibility in Acomastylis rossii (Rosaceae). Am J Bot 88(6):980–991. https://doi.org/10.2307/2657079
Oksanen J, Simpson G, Blanchet F, Kindt R, Legendre P, Minchin P, O’Hara R, Solymos P, Stevens M, Szoecs E, Wagner H, Barbour M, Bedward M, Bolker B, Borcard D, Carvalho G, Chirico M, De Caceres M, Durand S, Evangelista H, FitzJohn R, Friendly M, Furneaux B, Hannigan G, Hill M, Lahti L, McGlinn D, Ouellette M, Ribeiro Cunha E, Smith T, Stier A, Ter Braak C, Weedon J (2022) _vegan: Community Ecology Package_. R package version 2.6-4. https://CRAN.R-project.org/package=vegan
Oldfather MF, Britton MN, Papper PD, Koontz MJ, Halbur MM, Dodge C, Ackerly DD (2016) Effects of topoclimatic complexity on the composition of woody plant communities. https://doi.org/10.1093/aobpla/plw049. Ann Bot Plants
Östman SAH (2018) Plant-pollinator interactions in the alpine: Landscape heterogeneity acts as a potential buffer against climate-change induced mismatch in the pollinator-generalist Ranunculus acris. Master’s Thesis, The University of Bergen
Pardee GL, Jensen IO, Inouye DW, Irwin RE (2019) The individual and combined effects of snowmelt timing and frost exposure on the reproductive success of montane forbs. J Ecol 107(4):1970–1981. https://doi.org/10.1111/1365-2745.13152
Petraglia A, Tomaselli M, Petit Bon M, Delnevo N, Chiari G, Carbognani M (2014) Responses of flowering phenology of snowbed plants to an experimentally imposed extreme advanced snowmelt. J Plant Ecol 215:759–768. https://doi.org/10.1007/s11258-014-0368-1
Powers JM, Briggs HM, Dickson RG, Li X, Campbell DR (2022) Earlier snow melt and reduced summer precipitation alter floral traits important to pollination. Glob Change Biol 28(1):323–339. https://doi.org/10.1111/gcb.15908
Prather RM, Dalton RM, barr b, Blumstein DT, Boggs CL, Brody AK, Inouye DW, Irwin RE, Martin JGA, Smith RJ, Van Vuren DH, Wells CP, Whiteman HH, Inouye BD, Underwood N (2023) Current and lagged climate affects phenology across diverse taxonomic groups. Proc R Soc B: Biol Sci 290(1990). https://doi.org/10.1098/rspb.2022.2181
Prevéy JS, Rixen C, Rüger N, Høye TT, Bjorkman AD, Myers-Smith IH, Wipf S (2019) Warming shortens flowering seasons of tundra plant communities. Nat Ecol Evol 3(1):45–52. https://doi.org/10.1038/s41559-018-0745-6
Pyke GH, Inouye DW, Thomson JD (2011) Activity and abundance of bumble bees near Crested Butte, Colorado: diel, seasonal, and elevation effects. Ecol Entomol 36(4):511–521. https://doi.org/10.1111/j.1365-2311.2011.01295.x
Rafferty NE, Ives AR (2012) Pollinator effectiveness varies with experimental shifts in flowering time. Ecology 93(4):803–814. https://doi.org/10.1890/11-0967.1
Rose-Person A, Spasojevic M, Forrester C, Bowman W, Suding K, Oldfather M, Rafferty N (2023) Pollinator visitation, flower count, and seed set in black sand plots, 2020. Ver 1. https://doi.org/10.6073/pasta/920fe947b69eea415bdca0af3b3e33c9. Environmental Data Initiative
Sargent RD, Ackerly DD (2008) Plant–pollinator interactions and the assembly of plant communities. Trends Ecol Evol 23(3):123–130. https://doi.org/10.1016/j.tree.2007.11.003
Semenchuk PR, Gillespie MA, Rumpf SB, Baggesen N, Elberling B, Cooper EJ (2016) High Arctic plant phenology is determined by snowmelt patterns but duration of phenological periods is fixed: an example of periodicity. Environ Res Lett 11(12):125006. https://doi.org/10.1088/1748-9326/11/12/125006
Sherwood JA, Debinski DM, Caragea PC, Germino MJ (2017) Effects of experimentally reduced snowpack and passive warming on montane meadow plant phenology and floral resources. Ecosphere 8(3):e01745. https://doi.org/10.1002/ecs2.1745
Shibata A, Kudo G (2020) Floral abundance and bee density affect species-specific foraging patterns of alpine bumble bees. Arthropod Plant Interact 14:771–783. https://doi.org/10.1007/s11829-020-09787-0
Shipley B (2013) The AIC model selection method applied to path analytic models compared using ad-separation test. J Ecol 94(3):560–564. https://doi.org/10.1890/12-0976.1
Steltzer H, Landry C, Painter TH, Anderson J, Ayres E (2009) Biological consequences of earlier snowmelt from desert dust deposition in alpine landscapes. PNAS 106(28):11629–11634. https://doi.org/10.1073/pnas.0900758106
Stemkovski M, Pearse WD, Griffin SR, Pardee GL, Gibbs J, Griswold T, Irwin RE (2020) Bee phenology is predicted by climatic variation and functional traits. Ecol Lett 23(11):1589–1598. https://doi.org/10.1111/ele.13583
Suárez LH, Pérez F, Armesto JJ (2011) Strong phenotypic variation in floral design and display traits of an annual tarweed in relation to small-scale topographic heterogeneity in semiarid Chile. Int J Plant Sci 172(8):1012–1025. https://doi.org/10.1086/660105
Totland Ø (1993) Pollination in alpine Norway: flowering phenology, insect visitors, and visitation rates in two plant communities. Canad J Bot 71(8):1072–1079. https://doi.org/10.1139/b93-124
Totland Ø, Alatalo JM (2002) Effects of temperature and date of snowmelt on growth, reproduction, and flowering phenology in the arctic/alpine herb, Ranunculus glacialis. Oecologia 133(2):168–175. https://doi.org/10.1007/s00442-002-1028-z
Wipf S, Stoeckli V, Bebi P (2009) Winter climate change in alpine tundra: plant responses to changes in snow depth and snowmelt timing. Clim Change 94(1–2):105–121. https://doi.org/10.1007/s10584-009-9546-x
Zuur AF, Mira A, Carvalho F, Ieno EN, Saveliev AA, Smith GM, Walker NJ (2009) Negative binomial GAM and GAMM to analyse amphibian roadkills. In: Zuur AF, Ieno EN, Walker N, Smith GM (eds) Mixed effects models and extensions in ecology with R. Springer, New York, NY, pp 383–397
Acknowledgements
We thank Cliff Bueno de Mesquita for his substantial contributions to the initiation and development of the black sand addition experiment. We thank E. Joy, K. Gates, N. Campbell, C. Cosma, E. Hall, E. Hu, J. Dao, A. Li, and K. Sevilla for their immense contribution to data collection. We also thank J. G. Smith for her unwavering support in the field and with plant identification. In addition, we thank J. Resasco for his help establishing methodology, S. Elmendorf for her assistance with data interpretation, and K. Hess for creating a safe workplace. We thank J. Morse, T. Lampard, H. Brandes, and S. Aplet for their exceptional work in establishing and maintaining the Extended Growing Season Experiment at the Niwot Ridge. Finally, we thank the editors for organizing this special issue, and we thank three anonymous reviewers and D. Inouye for constructive comments that improved an earlier version of this manuscript.
Funding
This study received substantial funding from the Niwot Ridge LTER VII program (NWT VII: NSF DEB − 1637686). The authors have no competing interests to declare that are relevant to the content of this article.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by Annika Rose-Person with assistance from Nicole Rafferty, Marko Spasojevic, Chiara Forrester, and Meagan Oldfather. The first draft of the manuscript was written by Annika Rose-Person and Nicole Rafferty, and all authors commented on previous versions of the manuscript.
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rose-Person, A., Spasojevic, M.J., Forrester, C. et al. Early snowmelt advances flowering phenology and disrupts the drivers of pollinator visitation in an alpine ecosystem. Alp Botany (2024). https://doi.org/10.1007/s00035-024-00315-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00035-024-00315-x