
Abstract
Numerous disease-causing parasites must invade host cells in order to prosper. Collectively, such pathogens are responsible for a staggering amount of human sickness and death throughout the world. Leishmaniasis, Chagas disease, toxoplasmosis, and malaria are neglected diseases and therefore are linked to socio-economical and geographical factors, affecting well-over half the world’s population. Such obligate intracellular parasites have co-evolved with humans to establish a complexity of specific molecular parasite–host cell interactions, forming the basis of the parasite’s cellular tropism. They make use of such interactions to invade host cells as a means to migrate through various tissues, to evade the host immune system, and to undergo intracellular replication. These cellular migration and invasion events are absolutely essential for the completion of the lifecycles of these parasites and lead to their for disease pathogenesis. This review is an overview of the molecular mechanisms of protozoan parasite invasion of host cells and discussion of therapeutic strategies, which could be developed by targeting these invasion pathways. Specifically, we focus on four species of protozoan parasites Leishmania, Trypanosoma cruzi, Plasmodium, and Toxoplasma, which are responsible for significant morbidity and mortality.
Similar content being viewed by others

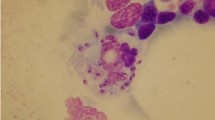
Mechanisms of host-cell invasion in Leishmania
Introduction
Leishmania are obligate intracellular protozoa that infect mononuclear phagocytes. The parasites exist in two morphological forms, as an intracellular aflagellated amastigote in the vertebrate hosts, such as humans, dogs, lizards or rodents, and as a motile flagellated promastigote in the invertebrate sand fly vector. Six major Leishmania species (L. tropica, L. major, L. donovani, L. infantum, L. braziliensis, and L. mexicana) cause the three main forms of the disease in humans, dermal cutaneous leishmaniasis, visceral leishmaniasis (VL), and muco-cutaneous leishmaniasis. The form and severity of the disease greatly depend on the infecting Leishmania species and the immune status of the host [1].
Life cycle
Leishmania amastigotes are taken up by a female sand fly from an infected vertebrate host during a blood meal. Within the gut of the sand fly, Leishmania amastigotes transform via intermediate non-infective procyclic promastigotes into infective metacyclic promastigotes [2]. Infective metacyclic promastigotes are introduced intradermally into the pool of blood of the vertebrate host by the feeding-infected sand fly [3]. Within a short span of time, neutrophils are recruited to site of sand fly bite, where they engulf the promastigotes providing them with temporary shelter [4]. Infected neutrophils or free parasites are then taken up by professional phagocytes (dendritic cells and macrophages), which migrate away from the site of the sand fly inoculation site [5]. Leishmania promastigotes survive in the parasitophorous vacuoles where they transform into amastigotes and replicate, eventually overburdening the infected cell leading to rupture. Extracellular amastigotes re-infect local phagocytes leading to subsequent cycles of infection. The transmission cycle is complete when infected phagocytes or extracellular parasites are taken up by feeding sandflies. Amastigotes then transform into promastigotes in the sand fly midgut and then further differentiate to become infective metacyclic promastigotes, which are able to infect a new vertebrate host [6]. Leishmania can also be transmitted through blood transfusion from previously infected to naive human hosts [7, 8].
Mediated uptake by host cells at the site of sand fly inoculation
When an infected sand fly releases Leishmania promastigotes into the skin, they evade immediate host immune response and destruction by seeking refuge in different types of host cells, which includes dermal macrophages (Fig. 1), fibroblasts, keratinocytes, and dendritic cells (DCs). Metacyclic promastigotes evade immediate destruction by deactivating the host’s complement system [9, 10]. In particular, lipophosphoglycan (LPG), which is highly expressed on the surface of metacyclic promastigotes, interferes with the insertion of membrane attack complex [11] and promastigote specific kinases deactivate the classical and alternative complement pathway by phosphorylating complement proteins [12]. Furthermore, the surface-membrane metalloprotease, glycoprotein 63 (known as gp63, leishmanolysin, or major surface protease), cleaves C3b attached to its surface, to an inactive form, C3bi, which facilitates the complement receptor 3 (CR3)-mediated entry into the host cells [13]. Leishmania promastigotes gain entry into the dermal macrophages by utilizing the host CR3, which prevents the parasites from being killed by circulating phagocytes and non-immune serum. Additionally, in dermal macrophages, which lack the respiratory burst machinery, promastigotes have enhanced ability to transform into and multiply as amastigotes compared to other macrophage populations [14]. Similarly, both Leishmania promastigotes and amastigotes are actively ingested by the skin fibroblasts [15]. These cells provide a safe environment for Leishmania for up to 7 days post-infection, as they produce low levels of nitric oxide even in the presence of lipopolysaccharide and interferon-γ, compared to macrophages [15]. The saliva of sand flies has been shown to have hemostatic and immunomodulatory effects in the host, which affect the infectivity and pathogenesis of Leishmania [16]. While many of these effects are common to the saliva from Old (Phlebotomus spp.) and New (Lutzomyia spp.) World sand flies, there also some effects that are different. Sand fly saliva contains a multitude of components, the most well described is the vasodilator protein called Maxadilan [17, 18]. Sand fly saliva increases the migration of inflammatory cells to the site of deposition, enhancing the interaction of Leishmania with potential host cells [19]. The saliva from Lutzomyia longipalpis and Maxadilan alone decrease TNF-α, IL-10, and increase IL-6, IL-8, and IL-12 production in LPS stimulated human macrophages [20]. Maxadilan diminishes DTH responses in mice and inhibits T cell activation [20–22], and Phlebotomus saliva up-regulates Th2 responses and down-regulates Th1 responses in L. major infection [23]. The saliva of Phlebotomus papatasi down-regulates NO production in LPS- or IFN-g-activated macrophages [23, 24]. Repeat exposure of hosts to sand fly saliva induced antibody formation and cell-mediated responses to saliva proteins. Interestingly, pre-exposure to sand fly saliva has been shown to be protective to subsequent Leishmania infection [25, 26], bringing forth the possibility is using saliva components as a vaccine [27–31]. Work is ongoing in order to understand how the responses to saliva protect against leishmaniasis or if the results of these studies in mice are applicable to humans [32].

Mechanisms of host-cell invasion in Leishmania
Keratinocytes are epithelial cells in the skin that form an important initial barrier between the host and the environment. The cellular interaction between keratinocytes and Leishmania is critical in delivering the initial triggers for Th-cell differentiation, although these are not target cells for parasite entry. Moreover, early cytokine expression of IL-1β, osteopontin, IL-12, IL-4, and IL-6 occurs in the keratinocytes of parasite-resistant mice strains following inoculation of Leishmania promastigotes. Among these cytokines, IL-6 and IL-4 from epidermal keratinocytes are essential for resistance and generation of protective Th1 immunity [33]. On the other hand, DCs present in the epidermis are highly motile and capable of crawling between interstitial tissue spaces serving as efficient pathogen sensors [34]. These cells internalize Leishmania and transport them to lymph nodes for priming of Leishmania specific T cells [35]. Interestingly, mice depleted of epidermal DCs had smaller lesions and reduced number of disease promoting T regulatory cells highlighting the role of DCs in promoting host susceptibility [36].
Mediated uptake by neutrophils
Neutrophils are the first cells to arrive at the site of infection within a few minutes of a sand fly bite [37]. This early recruitment of neutrophils might depend on specific cues provided by the release of alarmins (signal for tissue damage), chemokines and cytokines [38, 39]. Neutrophils are crucial cells for early defense against infections as they can kill obligate intracellular pathogens via reactive oxygen species (ROS) [40, 41], neutrophil elastase (NE), or neutrophil extracellular traps (NET) [42]. However, once phagocytic killing has been evaded or silenced, neutrophils can serve as host cells for Leishmania promastigotes [37]. Human neutrophils can readily phagocytose L. major promastigotes but amastigote recognition or uptake has not been detected by extracellular neutrophils [43]. Lipophoshoglycan and tartrate-resistant acid phosphatase present on the cell surface of Leishmania promastigotes inhibits lysosome fusion and the respiratory burst and superoxide anion production in neutrophils [44–46]. Leishmania promastigotes release chemotactic lipid Leishmania chemotactic factor (LCF), which attracts neutrophils [47] and interacts with Lipoxin A4 receptors (ALX), which deactivates the neutrophil oxidative burst [48]. Neutral elastase, a serine peptidase released by neutrophils, is deactivated by an inhibitor of serine peptidase present in Leishmania, which is crucial for intracellular parasite survival [49]. Moreover, virulent inoculum of Leishmania promastigotes contains a large subpopulation of apoptotic parasites, which bind to host annexin A5, which induces TGF-β and consequently mediates the silencing of phagocytes and survival of the viable subpopulation [50, 51].
Promastigotes can effectively modulate the short lifespan of neutrophils by delaying or accelerating apoptosis to their advantage. Neutrophils from human blood or murine peritoneal cavity have delayed apoptosis upon Leishmania infection [50–52]. On the contrary, neutrophils from infected dermis exhibit a marked increase in apoptosis, indicated by the enhanced expression of phosphatidylserine (PS). Interestingly, this increase in PS expression marks the infected neutrophils to be phagocytosed by dermal DCs [53]. Similarly, infected neutrophils are also phagocytosed by macrophages indirectly delivering promastigotes forming the basis for the Trojan horse model of macrophage infection [51]. However, recent in vivo studies have shown that neutrophils do not directly transfer their parasites to the macrophages. Instead, the apoptotic neutrophils lyse releasing the apoptotic and viable promastigotes, which are then engulfed by the macrophages [37, 53].
Mediated uptake by macrophages
Macrophages serve as one of the critical host cells for Leishmania parasites and it is therein where parasites undergo amastigotes transformation and replicate and deactivate the microbicidal machinery of the host cell. The C3bi (generated by C3b proteolysis by gp63) opsonized metacyclic promastigotes bind to macrophage receptor CR3 facilitating silent entry into the macrophages [13, 54]. Additionally, gp63 binds directly to fibronectin receptors through at least two domains [55, 56] and may also utilize this receptor by forming a bridge between gp63-bound fibronectin [57]. High-mannose type glycans on the surface of promastigotes also facilitate parasite binding through mannose-fucose receptors [58]. Gp63 may also bind directly to CR3 to promote invasion [59]. After initial attachment, Leishmania promastigotes initiate caveolae-dependent phagocytosis for entering host macrophages [43]. Leishmania LPG is inserted into phagosome cholesterol-rich lipid microdomains, which causes exclusion of the exocytosis regulator, synaptotagmin V, which prevents the acquisition of cathepsin D and the vesicular proton-ATPase, in the newly formed Leishmania-containing vacuoles (Fig. 1). This creates an intracellular phagosomal niche that fails to acidify and trigger microbicidal machinery, providing sufficient time for promastigotes to differentiate into amastigotes [60]. Furthermore, vacuoles containing promastigotes interact poorly with late endosomes and lysosomes, as observed by the delayed recruitment of LAMP-1 and Rab7. These events result in the delay of phagosome maturation, which interferes with the recruitment of signal transducers and vesicle trafficking to developing phagosomes [61, 62]. This is characterized by the formation of F-actin coat around the phagosome due to LPG-mediated deactivation of protein kinase C (PKC)α [63] and abnormal retention of actin polymerization machinery (Arp2/3,Wiskott Aldrich Syndrome Protein, myosin and α-actinin) and Cdc42 [64]. This also leads to the LPG-dependent exclusion of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase cytosolic components, \({\text{p47}}^{\text{phox}}\)and \({\text{p67}}^{\text{phox}}\), from the promastigote containing phagosome resulting in deactivation of ROS generation [65]. As promastigotes transform into amastigotes, the parasitophorous vacuoles become acidic (pH 4.7–5.2) [66, 67] and acquire lysosomal hydrolases and late endosomal/lysosomal proteins such as Rab7 and LAMP-1 [67]. Amastigotes multiply within these phagolysosomes until being released from ruptured macrophages and then enter neighboring macrophages through a non-caveolae pathway after ligating Fc-γ and PS receptors [1]. Amastigotes survive in phagolysosomes using several strategies that include: (1) deactivation of the ROS generation machinery [68, 69]; (2) attenuation of the PKC activity [70]; (3) suppression of STAT1 signaling [71]; (4) dysregulation of JAK/STAT signaling by depleting membrane cholesterol [72]; (5) activation of Leishmania specific ecto-protein phosphatases, cysteine proteinases or host cellular phosphotyrosine phosphatases or by inducing ceramide generation [73–77] (6) interference with the strength of CD40 cross-linking [78, 79]; (7) inhibition of mTOR complex 1 along with concomitant activation of translational repressor 4E-BP1, which promotes Leishmania proliferation [80]. In addition, the phagolysosomes containing Leishmania are enlarged due to increased expression of lysosomal trafficking regulator gene (LYST/Beige) that protects the parasites from oxidative damage by diluting the microbicidal effects of nitric oxide [81].
Furthermore, Leishmania also interferes with the strength of CD40 cross-linking or reciprocally activates MAPK phosphatases (MKP), MKP-1, and MKP-3 resulting in antagonistic regulation of ERK1/2 and p38MAPK, which governs the production of IL-10 and IL-12 by Leishmania-infected macrophages [78, 79].
Therapeutic considerations
The current anti-leishmanial drugs antimonials, amphotericin B, pentamidine, and miltefosine have limited therapeutic application due to toxicity and increasing multiple drug resistance. Novel therapeutic approaches are needed and those based on thwarting the mechanisms crucial for cellular invasion by Leishmania may hold promising results. Targeting MAPK signaling pathway using the specific MKP-1 inhibitor, triptolide, could be an effective therapeutic option for controlling L. major infection [79]. Similarly, AS-605240, a PI3Kγ inhibitor, is another promising therapeutic candidate as it confers resistance against L. mexicana infection and showed similar protection as sodium stibogluconate [82]. Furthermore, cysteine proteases, important for intracellular survival of Leishmania amastigotes, are targeted by cystatin, a natural cysteine protease inhibitor, for curing experimental VL and could be developed as potential therapeutic candidate [83, 84]. Since Leishmania depletes membrane cholesterol and disrupts lipid rafts in host macrophages, therapies based on restoration of cholesterol levels [72, 85, 86] and raft-associated proteins [87, 88] are other promising strategies.
Mechanisms of host-cell invasion by Trypanosoma cruzi
Introduction
The lifecycle of Trypanosoma cruzi was first described in the early 1900s by the Brazilian physician Carlos Chagas, who while working on malaria, suspected that insects infesting local rural dwellings transmitted human disease. Upon dissection he found motile flagellated parasites within the gastrointestinal tracts of these triatomine insects (family Reduviidae). He named the parasite after his mentor Prof. Oswaldo Cruz and determined that these were infective to mammals through experimental infection of monkeys, and further established they caused human disease by finding the parasite in the blood and tissues of acutely and chronically infected patients. In doing so, Chagas has been the only scientist that has completely described the epidemiology, clinical syndrome, etiology, vector and complete lifecycle of a microbial agent. Named after him, Chagas disease (also known as American trypanosomiasis), describes the clinical syndromes caused by infection with T. cruzi [89].
Life cycle of Trypanosoma cruzi
The life cycle of T. cruzi is complex and interesting. Transmission of T. cruzi occurs mainly by deposition of infective metacyclic trypomastigotes (MT) present in the urine and feces of hematophagous bugs from the family Reduviidae, of the genera Triatoma, Rhodnius, and Panstrongylus. The development of replicative non-infective epimastigote forms into infective MTs during their transit in the insect gastrointestinal tract is essential to the transmission to human hosts. Once MTs are deposited in the feeding wound they can invade nucleated hosts cells or be engulfed by professional phagocytes (macrophages and DCs) [90]. Human infection by the oral ingestion also occurs through the stomach epithelial invasion of MTs present in insect contaminated preparations of fruit juice [91–95]. After MT host-cell infection, parasites are initially housed in parasitophorous vacuoles but then escape into the host-cell cytoplasm and differentiate into aflagellated amastigotes. Amastigotes replicate in the cytoplasm until they are triggered to differentiate into motile flagellated trypomastigotes, which escape host cells and disseminate in the blood and lymph. The triggers for intracellular differentiation of amastigotes into trypomastigotes are unknown. Blood-stream trypomastigotes lead to further infection by binding to and invading nucleated non-phagocytic cells and are also phagocytized by macrophages, both similar to the initiation of infection that occurs by insect-derived MTs. Amastigotes are also infective and can disseminate in infected hosts. The life cycle is complete when trypomastigotes or amastigotes are ingested by reduviid bugs feeding on mammalian hosts, wherein they differentiate back into replicative epimastigotes. The mechanisms by which T. cruzi life cycle forms invade human cells are discussed in more detail below. While insect transmission is the primary mode of parasite acquisition, oral and transplacental routes can also occur and infection through transplanted blood and tissue from chronically infected individuals is increasing worldwide and has prompted expansion of screening practices of donated products [96, 97].
Mechanisms of host-cell invasion
T. cruzi can actively invade multiple non-phagocytic cells. The MTs and bloodstream trypomastigotes (also termed tissue-culture derived trypomastigotes, or TCT, produced in vitro by cultivation of parasites in tissue culture and provide a model of bloodstream forms) are the main infective forms of the parasite and will be referred to as trypomastigotes collectively throughout the rest of this review unless otherwise specified. Infection by amastigotes can also lead to productive infection although the mode of infection by these forms is not entirely understood. Epimastigotes are also infective in vitro but are not widely thought to contribute significantly to natural infection. The process of invasion by trypomastigotes is complex and dynamic and involves the interplay of multiple parasite and host factors. The bulk of studies looking at invasion have been done in vitro with a variety of different host-cell systems. This is complicated by the fact that different parasite strains infect cells by different mechanisms. Parasites can also infect macrophages by phagocytosis or through parasite invasion, as they do in non-phagocytic cells. There is controversy as to which of these is the main mode of parasite entry, however both are operative simultaneously. The information below provides a general overview of the main concepts concerning T. cruzi host-cell invasion. The readers may want to refer to several excellent and comprehensive reviews by pioneers in this field for further information [98–101].
Non-phagocytic cell infection
In vitro analysis of parasite invasion of non-phagocytic cells has resulted in the definition of several distinct steps (Fig. 2). Trypomastigotes initiate infection by interacting with the host-cell surface membrane by way of several parasite surface-membrane protein ligands. The host-cell receptors for many of these ligands have not yet been identified. Many members (gp82, gp90, gp85/TS, gp30) of the gp85/trans-sialidase (TS) super-family are implicated in the initiation of infection by trypomastigotes [102, 103]. Gp82, and the related gp30, are MT-specific surface proteins that upon engagement of an unknown receptor(s) mediate bidirectional Ca2+ release in both the parasite and host cell [104–106]. On the parasite side, this interaction triggers phospholipase C (PLC) activation generating diacylglycerol (DAG) and inositol triphosphate (IP3), which further activates protein kinase C and release of Ca2+ from the endoplasmic reticulum. In host-cell targets, PLC is activated, which ultimately release Ca2+ from intracellular stores via IP3-signaling. Recent data indicates that phosphatidylinositol 3-kinase (PI3K) and the mammalian target of rapamycin (mTOR) are also triggered during this process, however the downstream events beyond this are not well elucidated [106]. In some parasite strains, which express gp33/50, a mucin-related protein, host-cell Ca2+ release is triggered leading to invasion similar to but not as robustly as that mediated by gp82 [107]. Gp90 expressed in MTs of some parasite strains is a negative regulator of cell invasion and does not trigger Ca2+ mobilization or intracellular signaling [107, 108]. Gp82, −30, −90, and −35/50 are expressed to different degrees in various T. cruzi strains, which underscores the complex heterogeneity of the mechanisms of parasite invasion [109]. Bloodstream trypomastigotes appear to invade cells in a similar fashion but through a different set of interactions. Host-cell PLC activation occurs through triggering of the host bradykinin receptor, in an interesting mechanism in which parasite secrete cruzipain, a cysteine protease, which cleaves host kininogen to liberate bradykinin [110, 111]. Similar to MT invasion, PLC activation generates IP3, which triggers downstream release of host Ca2+. Ca2+ release is also mediated by action of a proteolytic cleavage product generated by oligopeptidase B activity on an unknown parasite protein substrate [112–114]. Parasite surface-membrane trans-sialidase (TS) promotes invasion of bloodstream forms through transfer of host surface sialic acid residues onto various parasite mucins-proteins, which are a large family of heavily glycosylated GPI-linked surface proteins [115–117]. Engagement of these sialylated parasite proteins trigger host-cell signaling via activation in a number of different ways including PI3K/Akt-, MAP-kinase and ERK- pathways [118–120].

Mechanisms of host-cell invasion by Trypanosoma cruzi
Parasite binding of extracellular matrix proteins (ECM) (fibronectin, laminin and collagen) and proteoglycans (heparin, heparin sulfate) is also implicated in invasion [121–124]. The gp85/TS superfamily of proteins been shown to bind multiple ECM components [123]. Also notable is a 60-kDa surface protein, termed Penetrin, which binds heparin and heparin sulfate in a sialic acid-independent manner that promotes invasion [125, 126], and may also associate with ECM components. Recently, homologs of the leishmanial surface protease, gp63, have been described in T. cruzi, and it appears as they may be involved in parasite–host cell binding [127, 128], however is it unclear if they are molecularly distinct from Penetrin.
Binding and invasion of host cells by parasites trigger several different, yet overlapping parasite entry pathways. Exploitation each of these is how by T. cruzi is such a successful pathogen at infecting multiple cell types. Each of these pathways, which culminate in the encasement of parasites within lysosomal-membrane-based vacuoles, which are necessary for subsequent events leading to productive infection [129].
In the “lysosomal-dependent pathway”, lysosomes are recruited to the site of parasite binding in a Ca2+-dependent manner. The exocytosis of lysosomes occurs by their fusion with the surface membrane abutting the parasite, which causes release of acid sphingomyelinase that generates ceramide through cleavage of membrane sphingomyelin [130]. The formation of ceramide in the outer leaflet of the membrane induces endocytosis of the wounded membrane. Parasites take advantage of this by becoming internalized in these lysosomal-based endosomes. This process may require the Ca2+-binding protein synaptotagmin VI and is probably powered by kinesin movement on microtubules that requires Ca2+-calmodulin [131, 132], both correlating with the necessary role of Ca2+ in this process. In the “lysosomal-independent pathway” parasites are initially housed in surface-membrane containing phosphoinositides, which eventually fuse with the lysosomes at later times post-infection [133, 134]. The protein dynamin, a GTPase, which associates with PI3K, may be involved in formation and dynamics of vacuole maturation [135, 136]. Prior to lysosomal fusion parasite entry into cells is reversible and parasites can escape the host cell. Thus fusion of these plasma membrane-derived vacuoles with lysosomes is a key step that leads to productive infection [137].
A third pathway has been proposed in which parasites exploit the autophagocytic pathway, which is operative under starvation conditions [138–140]. In this pathway, parasites invade cells and are initially housed in autophagocytic vacuoles, which eventually fuse with lysosomes to create an acidic autophagolysosomal compartment. These vacuoles are thought to be rich in nutrients for parasite survival and polyamines, which promote differentiation of trypomastigotes into amastigotes [141]. Overall, this last model is probably a modified version of the lysosomal-independent pathway specialized for conditions of starvation. The functional significance of actin and whether it is required or deleterious to parasite invasion in these pathways is likely a function of the ligands utilized for entry by different parasite forms [142].
Extracellular amastigotes are also infective to both phagocytic and non-phagocytic cells. Amastigotes of different strains appear to actively invade non-phagocytic host cells through recruitment of actin at the site of parasite attachment [142–144]. T. cruzi infection also occurs through the oral route where is has been responsible for several outbreaks [91, 95]. This occurs through parasite contamination of fruit juice (e.g., acai and guava fruit) from crushing of parasite-laden triatomines present on berries during processing. Consumption of contaminated juice delivers MTs into the stomach and duodenum where they invade the gastric mucosal lining. Parasite gp82 is a chief ligand for invasion and the proteolytic degradation of the anti-invasive ligand gp90 further facilitates parasite entry [145–147]. Parasite strains not expressing gp82 invade via the related gp30, which also triggers Ca2+ signaling. Parasite strains expressing gastric protease-susceptible gp90 are relatively more infective via the oral route than those expressing protease-resistant gp90 [148].
Phagocytic cell infection
Macrophages are also targets for parasite infection, especially at the initiation of infection when MTs are deposited in the tissue by insects. In addition, amastigotes released from infected cells also infect macrophages. Both stages are taken up by phagocytosis, initially housed in parasitophorous vacuoles, where they can be killed by generation of nitric oxide and peroxynitrite [149–151]. The maturation of parasitophorous vacuoles into phagolysosomes is important for parasite survival, which is required for entry into the host cytoplasm where they avoid exposure from microbicidal radicals. In the cytoplasm parasite differentiate and replicate as they do in non-phagocytic cells. The initial entry of parasites into macrophages is probably by way of both an actin-dependent phagocytic process and an active invasion process similar as those described above in non-phagocytic cells. Recent studies indicate that the integrity of membrane rafts composed of cholesterol, ganglioside GM, flotillin and caveolins are important in trypomastigote and amastigote infection of macrophages [152–154]. It is unclear whether, in the case of macrophages, this affects only active-invasion and/or the phagocytic parasite pathway. However, lipid membrane raft disruption diminishes infection of non-phagocytic cell invasion by trypomastigotes probably by disaggregation of proteins important for signaling pathways that initiate the invasion process.
Therapeutic considerations
The drugs currently used to treat Chagas disease include the nitroheterocyclics compounds benznidazole and nifurtimox, which are indicated in acute and intermediate stage T. cruzi infection. The mechanism of action of nifurtimox is not completely understood but is probably due to the induction of oxidative radical formation within the parasite and/or lowering of thiol levels through the production of nitroso-intermediate due to reduction of the nitro-group of the compound by the parasite [155–157]. The action of benznidazole is thought to act by reduction of macromolecule production through interactions of its nitro-group [158, 159]. The efficacy of these drugs in chronic Chagas disease is somewhat controversial and trials are underway to determine if there is any benefit of treatment at this late stage [160–162]. These drugs are significantly toxic, yet are the mainstay of anti-parasitic therapy because there are no other alternatives approved for human use at present. Inhibitors of cruzipain, an important cysteine protease of T. cruzi, are promising candidates for future therapy [163–165]. This protease is essential for viability of all stages of T. cruzi and is important as several key steps in the lifecycle, including parasite invasion (see above), cell division and differentiation [166]. Preliminary studies indicate that the vinyl sulfone inhibitor, K11777, is a therapeutic candidate and that other compounds that inhibit this enzyme may find there way into the pipeline of new investigational drugs.
Mechanisms of host-cell invasion by apicomplexan parasites
Introduction
Apicomplexan parasites are a unique group of protists united by the presence of an organelle called the apicoplast as well as the apical complex, a structure specifically involved in host-cell invasion. Although there are many organisms that fall under the phylum Apicomplexa, this section of the review will focus primarily on Plasmodium falciparum, the causative agent of cerebral malaria, with additional discussion of Toxoplasma gondii invasion, the causative agent of Toxoplasmosis.
Malaria is an infectious disease that has plagued man for millennia. Despite enormous struggles at prevention and treatment, currently over 250 million cases occur annually, resulting in an estimated 700,000 deaths, mostly African children [167]. Humans get infected with P. falciparum when bit by an infected mosquito, as this is the definitive host. The impact of this disease on the at-risk population is staggering and new, targeted approaches to prevent and cure it are desperately needed. T. gondii can infect any mammal, with humans being an accidental host. T. gondii can only sexually reproduce in the intestines of cats, the parasite’s definitive host, making every other host an intermediate host. In the United States over 60 million people carry the parasite but generally only those with compromised immune systems and pregnant women develop symptoms of disease [168]. Humans get infected by ingesting parasite-contaminated food or water or by ingesting infected and undercooked meat.
Although we understand a great deal regarding the biology of the parasites responsible for malaria and toxoplasmosis, it is only relatively recently that many of the molecular mechanisms by which the parasite recognizes and invades or traverses through host cells have been elucidated. There is a striking complexity of cellular traversal and invasion mechanisms utilized by Apicomplexa parasites for entry into the host cell. For Plasmodium, we focus on the two mammalian cell types into which the parasites invade, namely the liver hepatocyte and the erythrocyte. Toxoplasma is known to invade all nucleated cells, both non-phagocytic and phagocytic cells. For both species, we discuss notable parasite proteins and organelles involved in each of these invasion processes and highlight the host molecules known to be involved. As these processes represent critical events in the parasite’s life cycle, we finish with a brief section describing a number of ongoing efforts to target invasion in an attempt to combat these menacing diseases.
As this is meant to be a general review of Apicomplexan invasion, we regrettably have had to limit the depth of our description. Many excellent reviews exist that are more focused on various aspects of this topic and we direct the reader to these [169–175].
Plasmodium life cycle
Five species of Apicomplexan parasites of the genus Plasmodium infect humans to cause malaria and, like many parasites, they develop through discrete stages, each particularly adapted to one of its two hosts (the human and the female anopheline mosquito). These include P. falciparum, P. vivax, P. malariae, P. ovale and, shown recently to infect humans, P. knowlesi [176]. A striking feature of these parasites is their ability to invade a wide variety of cell types during this complex life cycle. This cycle can be broken down into eight steps: (1) In the midgut of an infected female mosquito, the definitive host, diploid zygotes migrate through the chitinous, cell-free peritrophic matrix and penetrate the midgut epithelium, taking up residence between the epithelium and the midgut basal lamina; (2) The zygotes then develops into oocysts that mature and release motile sporozoites that travel through the mosquito’s hemocoel, attaching to and traversing through salivary gland cells to access the interior of the gland; (3) Upon the female mosquito taking a blood meal from the mammalian host, sporozoites are deposited in dermal tissue and use gliding motility to reach dermal vessels, crossing the endothelium into the bloodstream; (4) Sporozoites then rapidly (within minutes) migrate to and cross the sinusoids of the liver by traversal through resident macrophages, called Kupffer cells. Once in the liver parenchyma, sporozoites trans-migrate through a number of hepatocytes by a membrane wounding process, after which they eventually invade a hepatocyte through invagination of the hepatocyte membrane; (5) Within the hepatocyte, sporozoites develop into exoerythrocytic forms that differentiate and replicate to produce haploid merozoites, which are then released into the peripheral bloodstream; (6) Merozoites invade erythrocytes, initiating the erythrocytic cycle of infection. During the erythrocytic stages, parasites replicate and progress through the following developmental stages within erythrocytes: ring, trophozoite and schizont. Merozoites egress from the erythrocyte and re-invade uninfected erythrocytes in a 48–72-h cycle, dependent on the species; (7) During this cycle of development and re-invasion of erythrocytes, some parasites transform into sexual gametes; (8) Gametocytes are taken up by a mosquito, and fertilize to become zygotes, thus completing the life cycle [176].
Plasmodium invasion of the hepatocyte
Invasion of a liver hepatocyte is perhaps best thought of as an ordered process of both extracellular and intracellular steps with the inhibition of any of these resulting in a block in infection. After traversing the dermal epithelium and entry to the bloodstream, sporozoites are carried to the liver where they arrest and undergo gliding motility on the surface of the sinusoids that form the reticuloendothelial system. This is proposed to be mediated through a multivalent interaction of the parasite’s surface-expressed circumsporozoite protein (CSP) and thrombospondin-related adhesive protein (TRAP) with sulfated heparin sulfate proteoglycans (HSPGs), secreted by liver stellate cells, which protrude through the endothelial fenestrae into the sinusoidal lumen. This interaction and arrest in the liver sinusoid explains why circulating sporozoites are virtually undetectable within minutes of infection, discovered by intravenously infecting rats with P. berghei sporozoites [177]. Traversal of the liver endothelium in rats does not appear to be through the liver endothelial cells, rather it involves a membrane invagination process (with formation of a parasitophorous vacuole) of the Kupffer cells present in the endothelial lining by P. berghei and P. yoelii [178]. Functional studies to block the activity of these phagocytic cells, either by heat killing or by specific inhibitors, suggested their involvement in sporozoite traversal, with a smaller percentage of traversal explained by a biologically active process [179]. Stronger evidence is from mouse experiments in which macrophage-colony-stimulating factor 1 (CSF-1) was knocked out, leading to greatly reduced numbers of Kupffer cells. These mice were significantly less susceptible to P. yoelii infection, indicating the necessary role of Kupffer in access to the liver parenchyma [180]. Once in the parenchyma of the liver, sporozoites traverse a number of hepatocytes in a non-invasive, membrane-wounding manner before finally invading a host hepatocyte through invagination and formation of a parasitophorous vacuole.
Plasmodium invasion of the erythrocyte
The merozoites released into the bloodstream from the liver exoerythrocytic cycle are small (~1 μm) and somewhat pear-shaped. At their apical end is located a specialized structure known as the apical complex in which reside specialized secretory organelles called rhoptries and micronemes that contain proteins necessary for invasion (Fig. 3). The limited tropism of merozoites is due to specific ligand-receptor interactions between the parasites and the erythrocytes. Initial interactions occur through proteins covering the surface of the merozoite, followed by a reorientation and formation of a tight junction between the erythrocyte and the apical end of the parasite. One of the best described proteins important in this initial interaction is the GPI-anchored protein, merozoite surface protein-1 (MSP-1), which has been suggested to interact with two nonglycosylated extracellular regions of Band 3 protein on the surface of the erythrocyte [181]. Orientation of the parasite to its apical end approximating the erythrocyte membrane then occurs followed by tight junction formation through high-affinity interactions between parasite ligands and erythrocyte receptors. Two families of proteins, the erythrocyte-binding-like (EBLs) protein and the reticulocyte-binding-like (RBLs or P. falciparum Rhs) proteins, have been well recognized as major parasite ligands responsible for binding and invasion and have been shown to play a cooperative role in invasion, as functional loss of the former leads to the increased transcription of the latter [182]. Both are localized to apical organelles and are released onto the parasite surface during invasion. EBLs and RBLs both appear to have specific receptors on the surface of erythrocytes, such as EBL-175 binding predominantly to glycophorin A [183]. In P. falciparum five EBLs and six RBLs have been described. With the exception of PfRh1, all the PfRhs bind in a sialic acid-independent manner and only PfRh4 has an identified receptor, complement receptor 1. Both families are present throughout Plasmodium spp. It is notable that P. falciparum appears to use multiple and alternative ligand-receptor interactions and is capable of switching invasion mechanisms between sialic acid dependent and sialic acid independent pathways, demonstrated using neuraminidase and trypsin treatments on erythrocytes [184, 185]. In different regions of India and Gambia, field isolates were tested and it was found that the majority of parasites invade by the EBL-175/glycophorin A pathway [186, 187]. However, in Brazil only a minority of field isolates used this pathway [188]. This is evidence that there is variation of invasion mechanisms in filed isolates from different regions of the world.

Mechanisms of host-cell invasion by apicomplexan parasites
Another well known binding interaction necessary for invasion occurs between the Duffy antigen and the Duffy Binding Protein (DBP) of P. vivax. The Duffy antigen, expressed on human erythrocytes, is the binding receptor for a cysteine rich domain of DBP [189]. Duffy negative [Fy(a-b-)] individuals, most commonly found in Western Africa, are naturally resistant to P. vivax infection [190]. The Duffy antigen has also been shown vital for P. knowlesi binding in vitro [191, 192]. The cysteine rich domain in P. knowlesi DBP is found to bind to Duffy antigen but with different specificity than that of P. vivax [189].
Toxoplasma gondii Life cycle
T. gondii invades a wide variety of cell types in a life cycle that can be described in four steps: (1) Tachyzoites are the fast-growing form that causing acute infection in humans and other animals. These infect nucleated host cells, replicating into a large number of progeny and egress by lysing the host cell; (2) Under immune pressure from the host, the parasite can differentiate into a slow-growing form, termed bradyzoites. This occurs in long-living tissue types such as muscle or neuronal cells; (3) Ingestion of animal tissue bradyzoites, by cats or humans releases parasites in the intestinal tract. In cats this results in sexual development and subsequent shedding of oocysts into the environment and in humans it can lead to disseminated infection; (4) Shed oocysts from cats develop into haploid sporozoites in the environment. The oral ingestion of these sporozoites leads to differentiation into tachyzoites and acute infection by humans or other mammals [193].
Toxoplasma gondii invasion of host cells
T. gondii tachyzoites use gliding motility, a mechanism not expanded upon in this review, to facilitate movement and interaction with host cells. Like a Plasmodium merozoite, surface antigens aid in the interaction between the tachyzoite and the host cell. One antigen known to have membrane-binding activity is Perforin-like Protein 1 (PLP-1), forming pores in the host-cell membrane after binding and as well as playing a role in egress [194]. The parasite then re-orients its position such that the apical end has contact with the host cell. T. gondii secretes proteins needed for invasion from micronemes, rhoptries and dense granules. The contents of the micronemes are secreted first during the initial contact with the host cell, including the full-length cellular form of microneme protein 2 (MIC2), which binds specifically to the host cell [195]. MIC2, along with MIC1, MIC4, MIC6 and MIC8, is involved the interaction between the parasite and the host cell [196, 197]. As the parasite invades the target cell these microneme proteins are proteolytically cleaved from the surface of the parasite. The rhoptries then secrete rhoptry neck proteins (RON). RON2 is inserted into the host-cell membrane of the host cell being invaded, while RON4, 5, 8, 9, and 10 form a complex at membrane surface of the membrane [198–201]. Tachyzoites then actively invade similar to Plasmodium merozoites, with the formation of the moving junction and a motor complex described below.
Process of apicomplexan invasion
The process of active invasion is so similar in Plasmodium and T. gondii that this section summarizes this invasion process used by both species of parasites with differences or unique mechanisms specifically noted.
Following tight junction formation between the parasite and the host-cell surface, parasites forcibly enter through invagination of the host-cell membrane, eventually forming and residing within parasitophorous vacuoles. The process involves a tight junction “motor” termed a moving junction (MJ) (Fig. 3). This junction can be thought of a “ring of contact” between the invading merozoite and the erythrocyte and is used by the parasite to “pull” itself into the cell. Once the parasite is completely inside the host cell, the MJ disappears as the parasitophorous vacuole resolves. No host proteins have been identified in the MJ although studies suggest host cytoskeleton components are involved [202, 203]. The most widely studied parasite protein involved in MJ formation is the apical membrane antigen-1 (AMA1), which is highly conserved across Plasmodium sp. [204–207] and T. gondii [200, 208]. AMA1 is secreted by micronemes and interacts with RON2, RON4 and RON5 [198, 209] as well as aldolase, an F-actin binding protein. In T. gondii, MIC6 also interacts directly with aldolase [196]. The RON proteins are part of a parasite derived protein complex termed the RON complex, which is secreted into the host cell upon MJ formation. The interaction between AMA1 and the RON complex is essential for parasite invasion [209]. In fact, antibodies to AMA1 can block invasion [210] and a 20-residue AMA1 binding peptide also specifically blocks invasion [211, 212], providing evidence that AMA-1 could be a potential vaccine target. As the MJ progresses around the parasite during invasion many proteins are shed from the surface of the parasite [213–215], including AMA-1 [216] and PTRAMP [217], a process required for invasion [214, 218, 219]. Recent work in Plasmodium has identified a subtilisin-like serine protease, PfSUB2, as the “sheddase” responsible for this indispensable event [220]. In addition to the role of SUB2 as a sheddase, a number of intramembrane proteases called rhomboids (ROM) are also implicated [215, 221–223]. In T. gondii, ROM4 is implicated in cleaving surface proteins to produce a gradient of adhesins, these including MIC2, AMA1, and MIC3 [224].
Therapeutic considerations for malaria
There are many drugs that treat malaria, the most recently developed and effective being multi-drug artemisinin therapies. However, to date all current antimalarials have encountered resistant parasites [225], highlighting the great need for continued drug development. Theoretically, a drug or vaccine that targets invasion of either hepatocytes or erythrocytes can be a useful preventative measure or treatment for malaria. Recent vaccine strategies are focusing on antigens crucial to invasion of hepatocytes, such as CSP, or of erythrocytes, such as EBLs and RBLs. Due to functional redundancy and antigen switching, multi-antigen vaccine strategies are more likely to act synergistically, reducing the chance of resistance and showing a more effective immune response.
Targeting hepatocytes invasion could be a great tool for prophylaxis drugs, vaccines or treatments to prevent recurrent malaria, as in the case of a P. vivax infection. The vaccine RTS,S, which is based on CSP from P. vivax, was recently reported in phase 3 clinical trials in Africa to be effective at decreasing the malaria rate in young children by nearly 50 % [226, 227]. Serum from rabbits treated with this P. vivax CSP-based vaccine has shown cross-species recognition of P. falciparum and P. berghei sporozoites, which is promising for the use of RTS,S against malaria caused by the various species of Plasmodium or even mixed infections [228].
Targeting erythrocytic invasion proteins could also serve as a useful vaccine strategy. Studies of polymorphisms of invasion-related ligands PfEBL and PfRh in field isolates from Colombia, Peru, and Brazil highlight the importance of understanding the variety of invasion pathways being utilized by parasites in different geographical regions [229]. This needs to be taken into account by developing multi-targeted vaccines, which might overcome “vaccine resistance” and be effective against parasites from different regions, different species of parasites, or mixed infections. As an additional example of this, antibodies against PfRh5 used in combination with seven other merozoite antigens, most notably PfRh4, have been shown to act synergistically in inhibiting growth of P. falciparum in culture [230].
Drug-targets include the proteases involved in invasion due to the essentiality of these proteases in egress and re-invasion of erythrocytes. Two peptidyl alpha-ketoamides based on the P. falciparum subtilisin-like protease 1 (SUB1) inhibited orthologues in P. berghei, P. vivax and P. knowlesi, suggesting the possibility of “pan-reactive” drugs based on this protease [231]. P. falciparum signal peptide peptidase, an intramembrane aspartyl protease associated with micronemes, is shown to interact with a transmembrane receptor on host erythrocytes and antibodies against it block invasion by P. falciparum in vitro [232]. Rhomboid proteases may also provide an effective drug target as they are essential to invasion [233, 234]. And as mentioned previously, the AMA1 and RON complex interaction is also of interest in vaccine development as antibodies and binding peptides to AMA-1 block invasion [210, 211, 220].
Therapeutic considerations for toxoplasmosis
Toxoplasmosis is also a treatable Apicomplexan disease, although careful consideration is needed for pregnant women and immune-compromised patients like those infected with HIV. Most medications target only the tachyzoite, leaving the bradyzoite unharmed. Current medications include pyrimethamine, which is also a malaria medication, and sulfadiazine. Vaccine studies using these invasion proteins have been relatively successful. A recent study has shown that a ROP8 DNA-based vaccine, encoding one of the rhoptry proteins secreted and essential for invasion, has induced a protective immune response in mice [235]. A DNA vaccine using a fusion protein of PLP-1 and MIC6 induced protective immunity in mice [236], as did MIC1 and MIC4 [237], indicating that these are promising candidates for vaccines. It is likely that other targets of invasion, as mentioned above for malaria, could also make good drug targets for toxoplasmosis.
Overall, invasion and traversal are crucial mechanisms for Apicomplexan parasite survival and infectivity. With multiple invasion events in multiple cell types, the molecular components of invasion are attractive targets for multi-component drugs and/or vaccines, which have the potential to both increase protective immune responses in humans and decrease chance of parasite resistance.
References
Kane MM, Mosser DM (2000) Leishmania parasites and their ploys to disrupt macrophage activation. Curr Opin Hematol 7:26–31
Rogers ME, Chance ML, Bates PA (2002) The role of promastigote secretory gel in the origin and transmission of the infective stage of Leishmania mexicana by the sandfly Lutzomyia longipalpis. Parasitology 124:495–507
Gossage SM, Rogers ME, Bates PA (2003) Two separate growth phases during the development of Leishmania in sand flies: implications for understanding the life cycle. Int J Parasitol 33:1027–1034
Kamhawi S (2006) Phlebotomine sand flies and Leishmania parasites: friends or foes? Trends Parasitol 22:439–445. doi:10.1016/j.pt.2006.06.012
Sacks DL, Perkins PV (1985) Development of infective stage Leishmania promastigotes within phlebotomine sand flies. Am J Trop Med Hyg 34:456–459
Kimblin N, Peters N, Debrabant A et al (2008) Quantification of the infectious dose of Leishmania major transmitted to the skin by single sand flies. Proc Natl Acad Sci USA 105:10125–10130. doi:10.1073/pnas.0802331105
le Fichoux YY, Quaranta JFJ, Aufeuvre JPJ et al (1999) Occurrence of Leishmania infantum parasitemia in asymptomatic blood donors living in an area of endemicity in southern France. J Clin Microbiol 37:1953–1957
Scarlata F, Vitale F, Saporito L et al (2008) Asymptomatic Leishmania infantum/chagasi infection in blood donors of western Sicily. Trans R Soc Trop Med Hyg 102:3. doi:10.1016/j.trstmh.2008.01.011
Da Silva RP, Hall BF, Joiner KA, Sacks DL (1989) CR1, the C3b receptor, mediates binding of infective Leishmania major metacyclic promastigotes to human macrophages. J Immunol 143:617–622
Mosser DM, Edelson PJ (1987) The third component of complement (C3) is responsible for the intracellular survival of Leishmania major. Nature 327:329–331. doi:10.1038/327329b0
Puentes SM, Da Silva RP, Sacks DL et al (1990) Serum resistance of metacyclic stage Leishmania major promastigotes is due to release of C5b-9. J Immunol 145:4311–4316
Hermoso T, Fishelson Z, Becker SI et al (1991) Leishmanial protein kinases phosphorylate components of the complement system. EMBO J 10:4061–4067
Brittingham A, Morrison CJ, McMaster WR et al (1995) Role of the Leishmania surface protease gp63 in complement fixation, cell adhesion, and resistance to complement-mediated lysis. J Immunol 155:3102–3111
Locksley RM, Heinzel FP, Fankhauser JE et al (1988) Cutaneous host defense in leishmaniasis: interaction of isolated dermal macrophages and epidermal Langerhans cells with the insect-stage promastigote. Infect Immun 56:336–342
Bogdan C, Donhauser N, Döring R et al (2000) Fibroblasts as host cells in latent leishmaniosis. J Exp Med 191:2121–2130
Titus RG, Ribeiro JM (1988) Salivary gland lysates from the sand fly Lutzomyia longipalpis enhance Leishmania infectivity. Science 239:1306–1308
Lerner EAE, Ribeiro JMJ, Nelson RJR, Lerner MRM (1991) Isolation of maxadilan, a potent vasodilatory peptide from the salivary glands of the sand fly Lutzomyia longipalpis. J Biol Chem 266:11234–11236
Lerner EA, Shoemaker CB (1992) Maxadilan. Cloning and functional expression of the gene encoding this potent vasodilator peptide. J Biol Chem 267:1062–1066
Anjili COC, Mbati PAP, Mwangi RWR et al (1995) The chemotactic effect of Phlebotomus duboscqi (Diptera: Psychodidae) salivary gland lysates to murine monocytes. Acta Trop 60:97–100
Costa DJD, Favali CC, Clarêncio JJ et al (2004) Lutzomyia longipalpis salivary gland homogenate impairs cytokine production and costimulatory molecule expression on human monocytes and dendritic cells. Infect Immun 72:1298–1305. doi:10.1128/IAI.72.3.1298- 1305.2004
Qureshi AAA, Asahina AA, Ohnuma MM et al (1996) Immunomodulatory properties of maxadilan, the vasodilator peptide from sand fly salivary gland extracts. Am J Trop Med Hyg 54:665–671
Rohousová II, Volf PP, Lipoldová MM (2005) Modulation of murine cellular immune response and cytokine production by salivary gland lysate of three sand fly species. Parasit Immunol 27:469–473. doi:10.1111/j.1365-3024.2005.00787.x
Mbow MLM, Bleyenberg JAJ, Hall LRL, Titus RGR (1998) Phlebotomus papatasi sand fly salivary gland lysate down-regulates a Th1, but up-regulates a Th2, response in mice infected with Leishmania major. J Immunol 161:5571–5577
Waitumbi JJ, Warburg AA (1998) Phlebotomus papatasi saliva inhibits protein phosphatase activity and nitric oxide production by murine macrophages. Infect Immun 66:1534–1537
Kamhawi S, Belkaid Y, Modi G et al (2000) Protection against cutaneous leishmaniasis resulting from bites of uninfected sand flies. Science 290:1351–1354
Belkaid YY, Kamhawi SS, Modi GG et al (1998) Development of a natural model of cutaneous leishmaniasis: powerful effects of vector saliva and saliva preexposure on the long-term outcome of Leishmania major infection in the mouse ear dermis. J Exp Med 188:1941–1953
Valenzuela JG, Belkaid Y, Garfield MK et al (2001) Toward a defined anti-Leishmania vaccine targeting vector antigens: characterization of a protective salivary protein. J Exp Med 194:331–342
Oliveira F, Lawyer PG, Kamhawi S, Valenzuela JG (2008) Immunity to distinct sand fly salivary proteins primes the anti-Leishmania immune response towards protection or exacerbation of disease. PLoS Negl Trop Dis 2:e226–e226. doi:10.1371/journal.pntd.0000226
Gomes RR, Teixeira CC, Teixeira MJM et al (2008) Immunity to a salivary protein of a sand fly vector protects against the fatal outcome of visceral leishmaniasis in a hamster model. Proc Natl Acad Sci USA 105:7845–7850. doi:10.1073/pnas.0712153105
Gomes RR, Oliveira FF, Teixeira CC et al (2012) Immunity to sand fly salivary protein LJM11 modulates host response to vector-transmitted leishmania conferring ulcer-free protection. 132:2735–2743. doi:10.1038/jid.2012.205
Morris RVR, Shoemaker CBC, David JRJ et al (2001) Sandfly maxadilan exacerbates infection with Leishmania major and vaccinating against it protects against L. major infection. J Immunol 167:5226–5230
Oliveira FF, Traoré BB, Gomes RR et al (2013) Delayed-type hypersensitivity to sand fly saliva in humans from a leishmaniasis-endemic area of Mali is t(h)1-mediated and persists to midlife. J Invest Dermatol 133:452–459. doi:10.1038/jid.2012.315
Ehrchen JM, Roebrock K, Foell D et al (2010) Keratinocytes determine Th1 immunity during early experimental leishmaniasis. PLoS Pathog 6:e1000871. doi:10.1371/journal.ppat.1000871
Ng LG, Hsu A, Mandell MA et al (2008) Migratory dermal dendritic cells act as rapid sensors of protozoan parasites. PLoS Pathog 4:e1000222. doi:10.1371/journal.ppat.1000222
Moll H, Fuchs H, Blank C, Röllinghoff M (1993) Langerhans cells transport Leishmania major from the infected skin to the draining lymph node for presentation to antigen-specific T cells. Eur J Immunol 23:1595–1601. doi:10.1002/eji.1830230730
Kautz-Neu K, Noordegraaf M, Dinges S et al (2011) Langerhans cells are negative regulators of the anti-Leishmania response. J Exp Med 208:885–891. doi:10.1084/jem.20102318
Peters NC, Egen JG, Secundino N et al (2008) In vivo imaging reveals an essential role for neutrophils in leishmaniasis transmitted by sand flies. Science 321:970–974. doi:10.1126/science.1159194
Bianchi ME (2007) DAMPs, PAMPs and alarmins: all we need to know about danger. J Leukoc Biol 81:1–5. doi:10.1189/jlb.0306164
Kaye P, Scott P (2011) Leishmaniasis: complexity at the host-pathogen interface. Nat Rev Microbiol 9:604–615. doi:10.1038/nrmicro2608
Pearson RD, Steigbigel RT (1981) Phagocytosis and killing of the protozoan Leishmania donovani by human polymorphonuclear leukocytes. J Immunol 127:1438–1443
Chang KP (1981) Leishmanicidal mechanisms of human polymorphonuclear phagocytes. Am J Trop Med Hyg 30:322–333
Guimarães-Costa AB, Nascimento MTC, Froment GS et al (2009) Leishmania amazonensis promastigotes induce and are killed by neutrophil extracellular traps. Proc Natl Acad Sci USA 106:6748–6753. doi:10.1073/pnas.0900226106
Wenzel UA, Bank E, Florian C et al (2012) Leishmania major parasite stage-dependent host cell invasion and immune evasion. FASEB J 26:29–39. doi:10.1096/fj.11-184895
Remaley AT, Kuhns DB, Basford RE et al (1984) Leishmanial phosphatase blocks neutrophil O-2 production. J Biol Chem 259:11173–11175
Gueirard P, Laplante A, Rondeau C et al (2008) Trafficking of Leishmania donovani promastigotes in non-lytic compartments in neutrophils enables the subsequent transfer of parasites to macrophages. Cell Microbiol 10:100–111. doi:10.1111/j.1462-5822.2007.01018.x
Tuwaijri al AS, Mofleh al IA, Mahmoud AA (1990) Effect of Leishmania major on human polymorphonuclear leucocyte function in vitro. J Med Microbiol 32:189–193
van Zandbergen GG, Hermann NN, Laufs HH et al (2002) Leishmania promastigotes release a granulocyte chemotactic factor and induce interleukin-8 release but inhibit gamma interferon-inducible protein 10 production by neutrophil granulocytes. Infect Immun 70:4177–4184. doi:10.1128/IAI.70.8.4177- 4184.2002
Wenzel A, van Zandbergen G (2009) Lipoxin A4 receptor dependent leishmania infection. Autoimmunity 42:331–333. doi:10.1080/08916930902828239
Faria MS, Reis FCG, Azevedo-Pereira RL et al (2011) Leishmania inhibitor of serine peptidase 2 prevents TLR4 activation by neutrophil elastase promoting parasite survival in murine macrophages. J Immunol 186:411–422. doi:10.4049/jimmunol.1002175
Aga E, Katschinski DM, van Zandbergen G et al (2002) Inhibition of the spontaneous apoptosis of neutrophil granulocytes by the intracellular parasite Leishmania major. J Immunol 169:898–905
van Zandbergen G, Klinger M, Mueller A et al (2004) Cutting edge: neutrophil granulocyte serves as a vector for Leishmania entry into macrophages. J Immunol 173:6521–6525
Charmoy M, Brunner-Agten S, Aebischer D et al (2010) Neutrophil-derived CCL3 is essential for the rapid recruitment of dendritic cells to the site of Leishmania major inoculation in resistant mice. PLoS Pathog 6:e1000755. doi:10.1371/journal.ppat.1000755
Ribeiro-Gomes FL, Peters NC, Debrabant A, Sacks DL (2012) Efficient capture of infected neutrophils by dendritic cells in the skin inhibits the early anti-leishmania response. PLoS Pathog 8:e1002536. doi:10.1371/journal.ppat.1002536
Blackwell JM, Ezekowitz RA, Roberts MB et al (1985) Macrophage complement and lectin-like receptors bind Leishmania in the absence of serum. J Exp Med 162:324–331
Brittingham A, Chen G, McGwire BS et al (1999) Interaction of Leishmania gp63 with cellular receptors for fibronectin. Infect Immun 67:4477–4484
Soteriadou KP, Remoundos MS, Katsikas MC et al (1992) The Ser-Arg-Tyr-Asp region of the major surface glycoprotein of Leishmania mimics the Arg-Gly-Asp-Ser cell attachment region of fibronectin. J Biol Chem 267:13980–13985
Ueno N, Wilson ME (2012) Receptor-mediated phagocytosis of Leishmania: implications for intracellular survival. Trends Parasitol 28:335–344. doi:10.1016/j.pt.2012.05.002
Chakraborty P, Ghosh D, Basu MK (2001) Modulation of macrophage mannose receptor affects the uptake of virulent and avirulent Leishmania donovani promastigotes. J Parasitol 87:1023–1027. doi:10.1645/0022-3395(2001)087[1023:MOMMRA]2.0
Russell DG, Wright SD (1988) Complement receptor type 3 (CR3) binds to an Arg-Gly-Asp-containing region of the major surface glycoprotein, gp63, of Leishmania promastigotes. J Exp Med 168:279–292
Vinet AF, Fukuda M, Turco SJ, Descoteaux A (2009) The Leishmania donovani lipophosphoglycan excludes the vesicular proton-ATPase from phagosomes by impairing the recruitment of synaptotagmin V. PLoS Pathog 5:e1000628. doi:10.1371/journal.ppat.1000628
Olivier M, Gregory DJ, Forget G (2005) Subversion mechanisms by which Leishmania parasites can escape the host immune response: a signaling point of view. Clin Microbiol Rev 18:293–305. doi:10.1128/CMR.18.2.293-305.2005
Scianimanico S, Desrosiers M, Dermine JF et al (1999) Impaired recruitment of the small GTPase rab7 correlates with the inhibition of phagosome maturation by Leishmania donovani promastigotes. Cell Microbiol 1:19–32
Holm A, Tejle K, Magnusson KE et al (2001) Leishmania donovani lipophosphoglycan causes periphagosomal actin accumulation: correlation with impaired translocation of PKCalpha and defective phagosome maturation. Cell Microbiol 3:439–447
Lodge R, Descoteaux A (2005) Leishmania donovani promastigotes induce periphagosomal F-actin accumulation through retention of the GTPase Cdc42. Cell Microbiol 7:1647–1658. doi:10.1111/j.1462-5822.2005.00582.x
Lodge R, Diallo TO, Descoteaux A (2006) Leishmania donovani lipophosphoglycan blocks NADPH oxidase assembly at the phagosome membrane. Cell Microbiol 8:1922–1931. doi:10.1111/j.1462-5822.2006.00758.x
Antoine JC, Prina E, Jouanne C, Bongrand P (1990) Parasitophorous vacuoles of Leishmania amazonensis-infected macrophages maintain an acidic pH. Infect Immun 58:779–787
Antoine JC, Prina E, Lang T, Courret N (1998) The biogenesis and properties of the parasitophorous vacuoles that harbour Leishmania in murine macrophages. Trends Microbiol 6:392–401
Lodge R, Descoteaux A (2006) Phagocytosis of Leishmania donovani amastigotes is Rac1 dependent and occurs in the absence of NADPH oxidase activation. Eur J Immunol 36:2735–2744. doi:10.1002/eji.200636089
Olivier M, Brownsey RW, Reiner NE (1992) Defective stimulus-response coupling in human monocytes infected with Leishmania donovani is associated with altered activation and translocation of protein kinase C. Proc Natl Acad Sci USA 89:7481–7485
Bhattacharyya S, Ghosh S, Jhonson PL et al (2001) Immunomodulatory role of interleukin-10 in visceral leishmaniasis: defective activation of protein kinase C-mediated signal transduction events. Infect Immun 69:1499–1507. doi:10.1128/IAI.69.3.1499- 1507.2001
Nandan D, Reiner NE (1995) Attenuation of gamma interferon-induced tyrosine phosphorylation in mononuclear phagocytes infected with Leishmania donovani: selective inhibition of signaling through Janus kinases and Stat1. Infect Immun 63:4495–4500
Sen S, Roy K, Mukherjee S et al (2011) Restoration of IFNγR subunit assembly, IFNγ signaling and parasite clearance in Leishmania donovani infected macrophages: role of membrane cholesterol. PLoS Pathog 7:e1002229. doi:10.1371/journal.ppat.1002229
Ghosh S, Bhattacharyya S, Das S et al (2001) Generation of ceramide in murine macrophages infected with Leishmania donovani alters macrophage signaling events and aids intracellular parasitic survival. Mol Cell Biochem 223:47–60
Nandan D, Lo R, Reiner NE (1999) Activation of phosphotyrosine phosphatase activity attenuates mitogen-activated protein kinase signaling and inhibits c-FOS and nitric oxide synthase expression in macrophages infected with Leishmania donovani. Infect Immun 67:4055–4063
Ghosh S, Bhattacharyya S, Sirkar M et al (2002) Leishmania donovani suppresses activated protein 1 and NF-kappaB activation in host macrophages via ceramide generation: involvement of extracellular signal-regulated kinase. Infect Immun 70:6828–6838
Cameron P, McGachy A, Anderson M et al (2004) Inhibition of lipopolysaccharide-induced macrophage IL-12 production by Leishmania mexicana amastigotes: the role of cysteine peptidases and the NF-kappaB signaling pathway. J Immunol 173:3297–3304
Yang Z, Mosser DM, Zhang X (2007) Activation of the MAPK, ERK, following Leishmania amazonensis infection of macrophages. J Immunol 178:1077–1085
Mathur RK, Awasthi A, Wadhone P et al (2004) Reciprocal CD40 signals through p38MAPK and ERK-1/2 induce counteracting immune responses. Nat Med 10:540–544. doi:10.1038/nm1045
Srivastava N, Sudan R, Saha B (2011) CD40-modulated dual-specificity phosphatases MAPK phosphatase (MKP)-1 and MKP-3 reciprocally regulate Leishmania major infection. J Immunol 186:5863–5872. doi:10.4049/jimmunol.1003957
Jaramillo M, Gomez MA, Larsson O et al (2011) Leishmania repression of host translation through mTOR cleavage is required for parasite survival and infection. Cell Host Microbe 9:331–341. doi:10.1016/j.chom.2011.03.008
Wilson J, Huynh C, Kennedy KA et al (2008) Control of parasitophorous vacuole expansion by LYST/Beige restricts the intracellular growth of Leishmania amazonensis. PLoS Pathog 4:e1000179. doi:10.1371/journal.ppat.1000179
Cummings HE, Barbi J, Reville P et al (2012) Critical role for phosphoinositide 3-kinase gamma in parasite invasion and disease progression of cutaneous leishmaniasis. Proc Natl Acad Sci USA 109:1251–1256. doi:10.1073/pnas.1110339109
Kar S, Ukil A, Das PK (2011) Cystatin cures visceral leishmaniasis by NF-κB-mediated proinflammatory response through co-ordination of TLR/MyD88 signaling with p105-Tpl2-ERK pathway. Eur J Immunol 41:116–127. doi:10.1002/eji.201040533
Das L, Datta N, Bandyopadhyay S, Das PK (2001) Successful therapy of lethal murine visceral leishmaniasis with cystatin involves up-regulation of nitric oxide and a favorable T cell response. J Immunol 166:4020–4028
Banerjee S, Ghosh J, Sen S et al (2009) Designing therapies against experimental visceral leishmaniasis by modulating the membrane fluidity of antigen-presenting cells. Infect Immun 77:2330–2342. doi:10.1128/IAI.00057-09
Ghosh J, Das S, Guha R et al (2012) Hyperlipidemia offers protection against Leishmania donovani infection: role of membrane cholesterol. J Lipid Res 53:2560–2572. doi:10.1194/jlr.M026914
Vilhardt F, van Deurs B (2004) The phagocyte NADPH oxidase depends on cholesterol-enriched membrane microdomains for assembly. EMBO J 23:739–748. doi:10.1038/sj.emboj.7600066
Chakraborty D, Banerjee S, Sen A et al (2005) Leishmania donovani affects antigen presentation of macrophage by disrupting lipid rafts. J Immunol 175:3214–3224
Delaporte F (2012) Chagas disease: history of a continent’s scourge. Fordham University Press
Van Overtvelt L, Vanderheyde N, Verhasselt V et al (1999) Trypanosoma cruzi infects human dendritic cells and prevents their maturation: inhibition of cytokines, HLA-DR, and costimulatory molecules. Infect Immun 67:4033–4040
Coura JR (2006) Transmission of chagasic infection by oral route in the natural history of Chagas disease. Rev Soc Bras Med Trop 39(Suppl 3):113–117
Pinto AYDN, Valente SA, Valente VDC et al (2008) Acute phase of Chagas disease in the Brazilian Amazon region: study of 233 cases from Pará, Amapá and Maranhão observed between 1988 and 2005. Rev Soc Bras Med Trop 41:602–614
Marcili A, Valente VC, Valente SA et al (2009) Trypanosoma cruzi in Brazilian Amazonia: Lineages TCI and TCIIa in wild primates, Rhodnius spp. and in humans with Chagas disease associated with oral transmission. Int J Parasitol 39:615–623. doi:10.1016/j.ijpara.2008.09.015
Nóbrega AA, Garcia MH, Tatto E et al (2009) Oral transmission of Chagas disease by consumption of açaí palm fruit, Brazil. Emerging Infect Dis 15:653–655. doi:10.3201/eid1504.081450
Alarcón de Noya B, Díaz-Bello Z, Colmenares C et al (2010) Large urban outbreak of orally acquired acute Chagas disease at a school in Caracas, Venezuela. J Infect Dis 201:1308–1315. doi:10.1086/651608
Fitzpatrick MA, Caicedo JC, Stosor V, Ison MG (2010) Expanded infectious diseases screening program for Hispanic transplant candidates. Transpl Infect Dis 12:336–341. doi:10.1111/j.1399-3062.2010.00517.x
Bern CC, Montgomery SPS, Katz LL et al (2008) Chagas disease and the US blood supply. Curr Opin Infect Dis 21:476–482. doi:10.1097/QCO.0b013e32830ef5b6
Maeda FY, Cortez C, Yoshida N (2012) Cell signaling during Trypanosoma cruzi invasion. Front Immunol 3:361. doi:10.3389/fimmu.2012.00361
Nagajyothi F, Machado FS, Burleigh BA et al (2012) Mechanisms of Trypanosoma cruzi persistence in Chagas disease. Cell Microbiol 14:634–643. doi:10.1111/j.1462-5822.2012.01764.x
Fernandes MC, Andrews NW (2012) Host cell invasion by Trypanosoma cruzi: a unique strategy that promotes persistence. FEMS Microbiol Rev 36:734–747. doi:10.1111/j.1574-6976.2012.00333.x
Burleigh BA, Woolsey AM (2002) Cell signalling and Trypanosoma cruzi invasion. Cell Microbiol 4:701–711
Weston D, Patel B, Van Voorhis WC (1999) Virulence in Trypanosoma cruzi infection correlates with the expression of a distinct family of sialidase superfamily genes. Mol Biochem Parasitol 98:105–116
Egima CM, Briones MR, Freitas Júnior LH et al (1996) Organization of trans-sialidase genes in Trypanosoma cruzi. Mol Biochem Parasitol 77:115–125
Manque PM, Eichinger D, Juliano MA et al (2000) Characterization of the cell adhesion site of Trypanosoma cruzi metacyclic stage surface glycoprotein gp82. Infect Immun 68:478–484
Ramirez MI, Boscardin SB, Han SW et al (1999) Heterologous expression of a Trypanosoma cruzi surface glycoprotein (gp82) in mammalian cells indicates the existence of different signal sequence requirements and processing. J Eukaryot Microbiol 46:557–565
Cortez M, Neira I, Ferreira D et al (2003) Infection by Trypanosoma cruzi metacyclic forms deficient in gp82 but expressing a related surface molecule, gp30. Infect Immun 71:6184–6191
Ruiz RC, Favoreto S, Dorta ML et al (1998) Infectivity of Trypanosoma cruzi strains is associated with differential expression of surface glycoproteins with differential Ca2+ signalling activity. Biochem J 330(Pt 1):505–511
Málaga S, Yoshida N (2001) Targeted reduction in expression of Trypanosoma cruzi surface glycoprotein gp90 increases parasite infectivity. Infect Immun 69:353–359. doi:10.1128/IAI.69.1.353-359.2001
Yoshida N, Cortez M (2008) Trypanosoma cruzi: parasite and host cell signaling during the invasion process. Subcell Biochem 47:82–91
Andrade D, Serra R, Svensjö E et al (2012) Trypanosoma cruzi invades host cells through the activation of endothelin and bradykinin receptors: a converging pathway leading to chagasic vasculopathy. Br J Pharmacol 165:1333–1347. doi:10.1111/j.1476-5381.2011.01609.x
Scharfstein J, Schmitz V, Morandi V et al (2000) Host cell invasion by Trypanosoma cruzi is potentiated by activation of bradykinin B(2) receptors. J Exp Med 192:1289–1300
Caler EV, Morty RE, Burleigh BA, Andrews NW (2000) Dual role of signaling pathways leading to Ca(2+) and cyclic AMP elevation in host cell invasion by Trypanosoma cruzi. Infect Immun 68:6602–6610
Caler EV, Vaena de Avalos S, Haynes PA et al (1998) Oligopeptidase B-dependent signaling mediates host cell invasion by Trypanosoma cruzi. EMBO J 17:4975–4986. doi:10.1093/emboj/17.17.4975
Burleigh BA, Caler EV, Webster P, Andrews NW (1997) A cytosolic serine endopeptidase from Trypanosoma cruzi is required for the generation of Ca2+ signaling in mammalian cells. J Cell Biol 136:609–620
Schenkman S, Kurosaki T, Ravetch JV, Nussenzweig V (1992) Evidence for the participation of the Ssp-3 antigen in the invasion of nonphagocytic mammalian cells by Trypanosoma cruzi. J Exp Med 175:1635–1641
Schenkman S, Jiang MS, Hart GW, Nussenzweig V (1991) A novel cell surface trans-sialidase of Trypanosoma cruzi generates a stage-specific epitope required for invasion of mammalian cells. Cell 65:1117–1125
Andrews NW, Hong KS, Robbins ES, Nussenzweig V (1987) Stage-specific surface antigens expressed during the morphogenesis of vertebrate forms of Trypanosoma cruzi. Exp Parasitol 64:474–484
Chuenkova MV, PereiraPerrin M (2009) Trypanosoma cruzi targets Akt in host cells as an intracellular antiapoptotic strategy. Sci Signal 2:ra74. doi:10.1126/scisignal.2000374
Chuenkova MV, PereiraPerrin M (2004) Chagas’ disease parasite promotes neuron survival and differentiation through TrkA nerve growth factor receptor. J Neurochem 91:385–394. doi:10.1111/j.1471-4159.2004.02724.x
Chuenkova MV, Furnari FB, Cavenee WK, Pereira MA (2001) Trypanosoma cruzi trans-sialidase: a potent and specific survival factor for human Schwann cells by means of phosphatidylinositol 3-kinase/Akt signaling. Proc Natl Acad Sci USA 98:9936–9941. doi:10.1073/pnas.161298398
Santana JM, Grellier P, Schrével J, Teixeira AR (1997) A Trypanosoma cruzi-secreted 80 kDa proteinase with specificity for human collagen types I and IV. Biochem J 325(Pt 1):129–137
Moody TN, Ochieng J, Villalta F (2000) Novel mechanism that Trypanosoma cruzi uses to adhere to the extracellular matrix mediated by human galectin-3. FEBS Lett 470:305–308
Alves MJM, Colli W (2008) Role of the gp85/trans-sialidase superfamily of glycoproteins in the interaction of Trypanosoma cruzi with host structures. Subcell Biochem 47:58–69
Nde PN, Lima MF, Johnson CA et al (2012) Regulation and use of the extracellular matrix by Trypanosoma cruzi during early infection. Front Immunol 3:337. doi:10.3389/fimmu.2012.00337
Ortega-Barria E, Pereira ME (1992) Entry of Trypanosoma cruzi into eukaryotic cells. Infect Agents Dis 1:136–145
Ortega-Barria E, Pereira ME (1991) A novel T. cruzi heparin-binding protein promotes fibroblast adhesion and penetration of engineered bacteria and trypanosomes into mammalian cells. Cell 67:411–421
Kulkarni MM, Olson CL, Engman DM, McGwire BS (2009) Trypanosoma cruzi GP63 proteins undergo stage-specific differential posttranslational modification and are important for host cell infection. Infect Immun 77:2193–2200. doi:10.1128/IAI.01542-08
Cuevas IC, Cazzulo JJ, Sánchez DO (2003) gp63 homologues in Trypanosoma cruzi: surface antigens with metalloprotease activity and a possible role in host cell infection. Infect Immun 71:5739–5749
Andrade LO, Andrews NW (2005) The Trypanosoma cruzi-host-cell interplay: location, invasion, retention. Nat Rev Microbiol 3:819–823. doi:10.1038/nrmicro1249
Fernandes MC, Cortez M, Flannery AR et al (2011) Trypanosoma cruzi subverts the sphingomyelinase-mediated plasma membrane repair pathway for cell invasion. J Exp Med 208:909–921. doi:10.1084/jem.20102518
Chakrabarti S, Andrade LO, Andrews NW (2005) Trypanosoma cruzi invades synaptotagmin VII-deficient cells by a PI-3 kinase independent pathway. Mol Biochem Parasitol 141:125–128. doi:10.1016/j.molbiopara.2005.01.016
Caler EV, Chakrabarti S, Fowler KT et al (2001) The Exocytosis-regulatory protein synaptotagmin VII mediates cell invasion by Trypanosoma cruzi. J Exp Med 193:1097–1104
Woolsey AM, Burleigh BA (2004) Host cell actin polymerization is required for cellular retention of Trypanosoma cruzi and early association with endosomal/lysosomal compartments. Cell Microbiol 6:829–838. doi:10.1111/j.1462-5822.2004.00405.x
Woolsey AM, Sunwoo L, Petersen CA et al (2003) Novel PI 3-kinase-dependent mechanisms of trypanosome invasion and vacuole maturation. J Cell Sci 116:3611–3622. doi:10.1242/jcs.00666
Barrias ES, Reignault LC, De Souza W, Carvalho TMU (2010) Dynasore, a dynamin inhibitor, inhibits Trypanosoma cruzi entry into peritoneal macrophages. PLoS One 5:e7764. doi:10.1371/journal.pone.0007764
Wilkowsky SE, Barbieri MA, Stahl PD, Isola ELD (2002) Regulation of Trypanosoma cruzi invasion of nonphagocytic cells by the endocytically active GTPases dynamin, Rab5, and Rab7. Biochem Biophys Res Commun 291:516–521. doi:10.1006/bbrc 2002.6474
Andrade LO, Andrews NW (2004) Lysosomal fusion is essential for the retention of Trypanosoma cruzi inside host cells. J Exp Med 200:1135–1143. doi:10.1084/jem.20041408
Romano PS, Arboit MA, Vázquez CL, Colombo MI (2009) The autophagic pathway is a key component in the lysosomal dependent entry of Trypanosoma cruzi into the host cell. Autophagy 5:6–18
Alvarez VE, Kosec G, Sant Anna C et al (2008) Blocking autophagy to prevent parasite differentiation: a possible new strategy for fighting parasitic infections? Autophagy 4:361–363
Alvarez VE, Kosec G, Sant’Anna C et al (2008) Autophagy is involved in nutritional stress response and differentiation in Trypanosoma cruzi. J Biol Chem 283:3454–3464. doi:10.1074/jbc.M708474200
Barclay JJ, Morosi LG, Vanrell MC et al (2011) Trypanosoma cruzi coexpressing ornithine decarboxylase and green fluorescence proteins as a tool to study the role of polyamines in Chagas disease pathology. Enzyme Res 2011:657460. doi:10.4061/2011/657460
Mortara RA, Andreoli WK, Fernandes MCDC et al (2008) Host cell actin remodeling in response to Trypanosoma cruzi: trypomastigote versus amastigote entry. Subcell Biochem 47:101–109
Mortara RA, Andreoli WK, Taniwaki NN et al (2005) Mammalian cell invasion and intracellular trafficking by Trypanosoma cruzi infective forms. An Acad Bras Cienc 77:77–94
Fernandes MC, Flannery AR, Andrews N, Mortara RA (2012) Extracellular amastigotes of Trypanosoma cruzi are potent inducers of phagocytosis in mammalian cells. Cell Microbiol. doi:10.1111/cmi.12090
Yoshida N (2008) Trypanosoma cruzi infection by oral route: how the interplay between parasite and host components modulates infectivity. Parasitol Int 57:105–109. doi:10.1016/j.parint.2007.12.008
Neira I, Silva FA, Cortez M, Yoshida N (2003) Involvement of Trypanosoma cruzi metacyclic trypomastigote surface molecule gp82 in adhesion to gastric mucin and invasion of epithelial cells. Infect Immun 71:557–561
Yoshida N (2009) Molecular mechanisms of Trypanosoma cruzi infection by oral route. Mem Inst Oswaldo Cruz 104(Suppl 1):101–107
Covarrubias C, Cortez M, Ferreira D, Yoshida N (2007) Interaction with host factors exacerbates Trypanosoma cruzi cell invasion capacity upon oral infection. Int J Parasitol 37:1609–1616. doi:10.1016/j.ijpara.2007.05.013
Alvarez MN, Peluffo G, Piacenza L, Radi R (2011) Intraphagosomal peroxynitrite as a macrophage-derived cytotoxin against internalized Trypanosoma cruzi: consequences for oxidative killing and role of microbial peroxiredoxins in infectivity. J Biol Chem 286:6627–6640. doi:10.1074/jbc.M110.167247
Thomson L, Gadelha FR, Peluffo G et al (1999) Peroxynitrite affects Ca2+ transport in Trypanosoma cruzi. Mol Biochem Parasitol 98:81–91
Rubbo H, Denicola A, Radi R (1994) Peroxynitrite inactivates thiol-containing enzymes of Trypanosoma cruzi energetic metabolism and inhibits cell respiration. Arch Biochem Biophys 308:96–102. doi:10.1006/abbi.1994.1014
Hissa B, Duarte JG, Kelles LF et al (2012) Membrane cholesterol regulates lysosome-plasma membrane fusion events and modulates Trypanosoma cruzi invasion of host cells. PLoS Negl Trop Dis 6:e1583. doi:10.1371/journal.pntd.0001583
Barrias ES, Dutra JMF, de Souza W, Carvalho TMU (2007) Participation of macrophage membrane rafts in Trypanosoma cruzi invasion process. Biochem Biophys Res Commun 363:828–834. doi:10.1016/j.bbrc.2007.09.068
Fernandes MC, Cortez M, Geraldo Yoneyama KA et al (2007) Novel strategy in Trypanosoma cruzi cell invasion: implication of cholesterol and host cell microdomains. Int J Parasitol 37:1431–1441. doi:10.1016/j.ijpara.2007.04.025
Kedderis GL, Miwa GT (1988) The metabolic activation of nitroheterocyclic therapeutic agents. Drug Metab Rev 19:33–62. doi:10.3109/03602538809049618
Viodé C, Bettache N, Cenas N et al (1999) Enzymatic reduction studies of nitroheterocycles. Biochem Pharmacol 57:549–557
Maya JD, Bollo S, Nuñez-Vergara LJ et al (2003) Trypanosoma cruzi: effect and mode of action of nitroimidazole and nitrofuran derivatives. Biochem Pharmacol 65:999–1006
Docampo R, Moreno SN (1986) Free radical metabolism of antiparasitic agents. Fed Proc 45:2471–2476
Gonzalez NS, Cazzulo JJ (1989) Effects of trypanocidal drugs on protein biosynthesis in vitro and in vivo by Trypanosoma cruzi. Biochem Pharmacol 38:2873–2877
Matta Guedes PM, Gutierrez FRS, Nascimento MSL et al (2012) Antiparasitical chemotherapy in Chagas’ disease cardiomyopathy: current evidence. Trop Med Int Health 17:1057–1065. doi:10.1111/j.1365-3156.2012.03025.x
Marin-Neto JA, Rassi A, Avezum A et al (2009) The BENEFIT trial: testing the hypothesis that trypanocidal therapy is beneficial for patients with chronic Chagas heart disease. Mem Inst Oswaldo Cruz 104(Suppl 1):319–324
de Andrade AL, Zicker F, de Oliveira RM et al (1996) Randomised trial of efficacy of benznidazole in treatment of early Trypanosoma cruzi infection. Lancet 348:1407–1413
Sajid M, Robertson SA, Brinen LS, McKerrow JH (2011) Cruzain : the path from target validation to the clinic. Adv Exp Med Biol 712:100–115. doi:10.1007/978-1-4419-8414-2_7
Doyle PS, Zhou YM, Engel JC, McKerrow JH (2007) A cysteine protease inhibitor cures Chagas’ disease in an immunodeficient-mouse model of infection. Antimicrob Agents Chemother 51:3932–3939. doi:10.1128/AAC.00436-07
Barr SC, Warner KL, Kornreic BG et al (2005) A cysteine protease inhibitor protects dogs from cardiac damage during infection by Trypanosoma cruzi. Antimicrob Agents Chemother 49:5160–5161. doi:10.1128/AAC.49.12.5160-5161.2005
Jose Cazzulo J, Stoka V, Turk V (2001) The major cysteine proteinase of Trypanosoma cruzi: a valid target for chemotherapy of Chagas disease. Curr Pharm Des 7:1143–1156
World Health Organization (WHO) (2010) Guidelines for the treatment of malaria. WHO, Geneva, Switzerland, p 194
Centers for disease control and prevention (CDC) (2013) Parasites—toxoplasmosis. Georgia, USA, CD. http://www.cdc.gov/toxoplasmosis/
Baum J, Gilberger T-W, Frischknecht F, Meissner M (2008) Host-cell invasion by malaria parasites: insights from Plasmodium and Toxoplasma. Trends Parasitol 24:557–563. doi:10.1016/j.pt.2008.08.006
Ménard R (2000) The journey of the malaria sporozoite through its hosts: two parasite proteins lead the way. Microbes Infect 2:633–642
Ghosh A, Edwards MJ, Jacobs-Lorena M (2000) The journey of the malaria parasite in the mosquito: hopes for the new century. Parasitol Today (Regul Ed) 16:196–201
Tyler JS, Treeck M, Boothroyd JC (2011) Focus on the ringleader: the role of AMA1 in apicomplexan invasion and replication. Trends Parasitol 27:410–420. doi:10.1016/j.pt.2011.04.002
Kappe SHI, Kaiser K, Matuschewski K (2003) The Plasmodium sporozoite journey: a rite of passage. Trends Parasitol 19:135–143
Shen B, Sibley LD (2012) The moving junction, a key portal to host cell invasion by apicomplexan parasites. Curr Opin Microbiol 15:449–455. doi:10.1016/j.mib.2012.02.007
Sibley LD (2011) Invasion and intracellular survival by protozoan parasites. Immunol Rev 240:72–91. doi:10.1111/j.1600-065X.2010.00990.x
Garcia CR, de Azevedo MF, Wunderlich G et al (2008) Plasmodium in the postgenomic era: new insights into the molecular cell biology of malaria parasites. Int Rev Cell Mol Biol 266:85–156. doi:10.1016/S1937-6448(07)66003-1
Shin SC, Vanderberg JP, Terzakis JA (1982) Direct infection of hepatocytes by sporozoites of Plasmodium berghei. J Protozool 29:448–454
Pradel G, Frevert U (2001) Malaria sporozoites actively enter and pass through rat Kupffer cells prior to hepatocyte invasion. Hepatology 33:1154–1165. doi:10.1053/jhep.2001.24237
Sihden RE, Smith JE (1982) The role of the Kupffer cell in the infection of rodents by sporozoites of Plasmodium: uptake of sporozoites by perfused liver and the establishment of infection in vivo. Acta Trop 39:11–27
Baer K, Roosevelt M, Clarkson AB et al (2007) Kupffer cells are obligatory for Plasmodium yoelii sporozoite infection of the liver. Cell Microbiol 9:397–412. doi:10.1111/j.1462-5822.2006.00798.x
Goel VK, Li X, Chen H et al (2003) Band 3 is a host receptor binding merozoite surface protein 1 during the Plasmodium falciparum invasion of erythrocytes. Proc Natl Acad Sci USA 100:5164–5169. doi:10.1073/pnas.0834959100
Lopaticki S, Maier AG, Thompson J et al (2011) Reticulocyte and erythrocyte binding-like proteins function cooperatively in invasion of human erythrocytes by malaria parasites. Infect Immun 79:1107–1117. doi:10.1128/IAI.01021-10
Dolan SA, Proctor JL, Alling DW et al (1994) Glycophorin B as an EBA-175 independent Plasmodium falciparum receptor of human erythrocytes. Mol Biochem Parasitol 64:55–63
Dolan SA, Miller LH, Wellems TE (1990) Evidence for a switching mechanism in the invasion of erythrocytes by Plasmodium falciparum. J Clin Invest 86:618–624. doi:10.1172/JCI114753
Mitchell GH, Hadley TJ, McGinniss MH et al (1986) Invasion of erythrocytes by Plasmodium falciparum malaria parasites: evidence for receptor heterogeneity and two receptors. Blood 67:1519–1521
Okoyeh JN, Pillai CR, Chitnis CE (1999) Plasmodium falciparum field isolates commonly use erythrocyte invasion pathways that are independent of sialic acid residues of glycophorin A. Infect Immun 67:5784–5791
Baum J, Pinder M, Conway DJ (2003) Erythrocyte invasion phenotypes of Plasmodium falciparum in The Gambia. Infect Immun 71:1856–1863
Lobo C-A, de Frazao K, Rodriguez M et al (2004) Invasion profiles of Brazilian field isolates of Plasmodium falciparum: phenotypic and genotypic analyses. Infect Immun 72:5886–5891. doi:10.1128/IAI.72.10.5886-5891.2004
Chitnis CE, Miller LH (1994) Identification of the erythrocyte binding domains of Plasmodium vivax and Plasmodium knowlesi proteins involved in erythrocyte invasion. J Exp Med 180:497–506
Gelpi AP, King MC (1976) Association of Duffy blood groups with the sickle cell trait. Hum Genet 32:65–68
Mason SJ, Miller LH, Shiroishi T et al (1977) The Duffy blood group determinants: their role in the susceptibility of human and animal erythrocytes to Plasmodium knowlesi malaria. Br J Haematol 36:327–335
Miller LH, Mason SJ, Dvorak JA et al (1975) Erythrocyte receptors for (Plasmodium knowlesi) malaria: Duffy blood group determinants. Science 189:561–563
Dubey JP, Lindsay DS, Speer CA (1998) Structures of Toxoplasma gondii tachyzoites, bradyzoites, and sporozoites and biology and development of tissue cysts. Clin Microbiol Rev 11:267–299
Roiko MS, Carruthers VB (2013) Functional dissection of Toxoplasma gondii perforin-like protein 1 reveals a dual domain mode of membrane binding for cytolysis and parasite egress. J Biol Chem 288:8712–8725. doi:10.1074/jbc.M113.450932
Carruthers VB, Giddings OK, Sibley LD (1999) Secretion of micronemal proteins is associated with toxoplasma invasion of host cells. Cell Microbiol 1:225–235
Zheng B, He A, Gan M et al (2009) MIC6 associates with aldolase in host cell invasion by Toxoplasma gondii. Parasitol Res 105:441–445. doi:10.1007/s00436-009-1401-5
Kessler H, Herm-Götz A, Hegge S et al (2008) Microneme protein 8–a new essential invasion factor in Toxoplasma gondii. J Cell Sci 121:947–956. doi:10.1242/jcs.022350
Alexander DL, Mital J, Ward GE et al (2005) Identification of the moving junction complex of Toxoplasma gondii: a collaboration between distinct secretory organelles. PLoS Pathog 1:e17. doi:10.1371/journal.ppat.0010017
Straub KW, Cheng SJ, Sohn CS, Bradley PJ (2009) Novel components of the Apicomplexan moving junction reveal conserved and coccidia-restricted elements. Cell Microbiol 11:590–603. doi:10.1111/j.1462-5822.2008.01276.x
Besteiro S, Michelin A, Poncet J et al (2009) Export of a Toxoplasma gondii rhoptry neck protein complex at the host cell membrane to form the moving junction during invasion. PLoS Pathog 5:e1000309. doi:10.1371/journal.ppat.1000309
Lamarque MH, Papoin J, Finizio A-L et al (2012) Identification of a new rhoptry neck complex RON9/RON10 in the Apicomplexa parasite Toxoplasma gondii. PLoS One 7:e32457. doi:10.1371/journal.pone.0032457
Gonzalez V, Combe A, David V et al (2009) Host cell entry by apicomplexa parasites requires actin polymerization in the host cell. Cell Host Microbe 5:259–272. doi:10.1016/j.chom.2009.01.011
Sweeney KR, Morrissette NS, LaChapelle S, Blader IJ (2010) Host cell invasion by Toxoplasma gondii is temporally regulated by the host microtubule cytoskeleton. Eukaryot Cell 9:1680–1689. doi:10.1128/EC.00079-10
Cowman AF, Crabb BS (2006) Invasion of red blood cells by malaria parasites. Cell 124:755–766. doi:10.1016/j.cell.2006.02.006
Riglar DT, Richard D, Wilson DW et al (2011) Super-resolution dissection of coordinated events during malaria parasite invasion of the human erythrocyte. Cell Host Microbe 9:9–20. doi:10.1016/j.chom.2010.12.003
Waters AP, Thomas AW, Deans JA et al (1990) A merozoite receptor protein from Plasmodium knowlesi is highly conserved and distributed throughout Plasmodium. J Biol Chem 265:17974–17979
Gaffar FR, Yatsuda AP, Franssen FFJ, de Vries E (2004) Erythrocyte invasion by Babesia bovis merozoites is inhibited by polyclonal antisera directed against peptides derived from a homologue of Plasmodium falciparum apical membrane antigen 1. Infect Immun 72:2947–2955
Hehl AB, Lekutis C, Grigg ME et al (2000) Toxoplasma gondii homologue of Plasmodium apical membrane antigen 1 is involved in invasion of host cells. Infect Immun 68:7078–7086
Richard D, MacRaild CA, Riglar DT et al (2010) Interaction between Plasmodium falciparum apical membrane antigen 1 and the rhoptry neck protein complex defines a key step in the erythrocyte invasion process of malaria parasites. J Biol Chem 285:14815–14822. doi:10.1074/jbc.M109.080770
Hodder AN, Crewther PE, Anders RF (2001) Specificity of the protective antibody response to apical membrane antigen 1. Infect Immun 69:3286–3294. doi:10.1128/IAI.69.5.3286- 3294.2001
Harris KS, Casey JL, Coley AM et al (2009) Rapid optimization of a peptide inhibitor of malaria parasite invasion by comprehensive N-methyl scanning. J Biol Chem 284:9361–9371. doi:10.1074/jbc.M808762200
Harris KS, Casey JL, Coley AM et al (2005) Binding hot spot for invasion inhibitory molecules on Plasmodium falciparum apical membrane antigen 1. Infect Immun 73:6981–6989
Blackman MJ, Heidrich HG, Donachie S et al (1990) A single fragment of a malaria merozoite surface protein remains on the parasite during red cell invasion and is the target of invasion-inhibiting antibodies. J Exp Med 172:379–382
Blackman MJ (2000) Proteases involved in erythrocyte invasion by the malaria parasite: function and potential as chemotherapeutic targets. Curr Drug Targets 1:59–83
Howell SA, Hackett F, Jongco AM et al (2005) Distinct mechanisms govern proteolytic shedding of a key invasion protein in apicomplexan pathogens. Mol Microbiol 57:1342–1356. doi:10.1111/j.1365-2958.2005.04772.x
Howell SA, Withers-Martinez C, Kocken CH et al (2001) Proteolytic processing and primary structure of Plasmodium falciparum apical membrane antigen-1. J Biol Chem 276:31311–31320. doi:10.1074/jbc.M103076200
Green JL, Hinds L, Grainger M et al (2006) Plasmodium thrombospondin related apical merozoite protein (PTRAMP) is shed from the surface of merozoites by PfSUB2 upon invasion of erythrocytes. Mol Biochem Parasitol 150:114–117
Fleck SL, Birdsall B, Babon J et al (2003) Suramin and suramin analogues inhibit merozoite surface protein-1 secondary processing and erythrocyte invasion by the malaria parasite Plasmodium falciparum. J Biol Chem 278:47670–47677. doi:10.1074/jbc.M306603200
Dutta S, Haynes JD, Barbosa A et al (2005) Mode of action of invasion-inhibitory antibodies directed against apical membrane antigen 1 of Plasmodium falciparum. Infect Immun 73:2116–2122. doi:10.1128/IAI.73.4.2116-2122.2005
Harris PK, Yeoh S, Dluzewski AR et al (2005) Molecular identification of a malaria merozoite surface sheddase. PLoS Pathog 1:241–251. doi:10.1371/journal.ppat.0010029
Howell SA, Well I, Fleck SL et al (2003) A single malaria merozoite serine protease mediates shedding of multiple surface proteins by juxtamembrane cleavage. J Biol Chem 278:23890–23898. doi:10.1074/jbc.M302160200
O’Donnell RA, Hackett F, Howell SA et al (2006) Intramembrane proteolysis mediates shedding of a key adhesin during erythrocyte invasion by the malaria parasite. J Cell Biol 174:1023–1033. doi:10.1083/jcb.200604136
Baker RP, Wijetilaka R, Urban S (2006) Two Plasmodium rhomboid proteases preferentially cleave different adhesins implicated in all invasive stages of malaria. PLoS Pathog 2:e113. doi:10.1371/journal.ppat.0020113
Buguliskis JS, Brossier F, Shuman J, Sibley LD (2010) Rhomboid 4 (ROM4) affects the processing of surface adhesins and facilitates host cell invasion by Toxoplasma gondii. PLoS Pathog 6:e1000858. doi:10.1371/journal.ppat.1000858
Cowman AF (1995) Mechanisms of drug resistance in malaria. Aust N Z J Med 25:837–844
Agnandji ST, Lell B, Soulanoudjingar SS et al (2011) First results of phase 3 trial of RTS, S/AS01 malaria vaccine in African children. N Engl J Med 365:1863–1875. doi:10.1056/NEJMoa1102287
RTS,S Clinical Trials Partnership, Agnandji ST, Lell B et al (2012) A phase 3 trial of RTS,S/AS01 malaria vaccine in African infants. N Engl J Med 367:2284–2295. doi:10.1056/NEJMoa1208394
Yadava A, Sattabongkot J, Washington MA et al (2007) A novel chimeric Plasmodium vivax circumsporozoite protein induces biologically functional antibodies that recognize both VK210 and VK247 sporozoites. Infect Immun 75:1177–1185
Lopez-Perez M, Villasis E, Machado RLD et al (2012) Plasmodium falciparum field isolates from South America use an atypical red blood cell invasion pathway associated with invasion ligand polymorphisms. PLoS One 7:e47913
Williams AR, Douglas AD, Miura K et al (2012) Enhancing blockade of Plasmodium falciparum erythrocyte invasion: assessing combinations of antibodies against PfRH5 and other merozoite antigens. PLoS Pathog 8:e1002991. doi:10.1371/journal.ppat.1002991
Withers-Martinez C, Suarez C, Fulle S et al (2012) Plasmodium subtilisin-like protease 1 (SUB1): insights into the active-site structure, specificity and function of a pan-malaria drug target. Int J Parasitol 42:597–612
Li X, Chen H, Oh SS, Chishti AH (2008) A Presenilin-like protease associated with Plasmodium falciparum micronemes is involved in erythrocyte invasion. Mol Biochem Parasitol 158:22–31. doi:10.1016/j.molbiopara.2007.11.007
Ejigiri I, Ragheb DRT, Pino P et al (2012) Shedding of TRAP by a rhomboid protease from the malaria sporozoite surface is essential for gliding motility and sporozoite infectivity. PLoS Pathog 8:e1002725. doi:10.1371/journal.ppat.1002725
Brossier F, Starnes GL, Beatty WL, Sibley LD (2008) Microneme rhomboid protease TgROM1 is required for efficient intracellular growth of Toxoplasma gondii. Eukaryot Cell 7:664–674. doi:10.1128/EC.00331-07
Parthasarathy S, Fong MY, Ramaswamy K, Lau YL (2013) Protective Immune Response in BALB/c Mice Induced by DNA Vaccine of the ROP8 gene of Toxoplasma gondii. Am J Trop Med Hyg. doi:10.4269/ajtmh.12-0727
Yan H-K, Yuan Z-G, Song H-Q et al (2012) Vaccination with a DNA vaccine coding for perforin-like protein 1 and MIC6 induces significant protective immunity against Toxoplasma gondii. Clin Vaccine Immunol 19:684–689. doi:10.1128/CVI.05578-11
Lourenço EV, Bernardes ES, Silva NM et al (2006) Immunization with MIC1 and MIC4 induces protective immunity against Toxoplasma gondii. Microbes Infect 8:1244–1251. doi:10.1016/j.micinf.2005.11.013
Acknowledgments
Research in Dr. Satoskar’s lab is supported by National Institutes of Health grants R03AI090231, RC4AI092624, R34AI100789, R21AT004160 and R03CA164399.
Author information
Authors and Affiliations
Corresponding author
Additional information
B. S. McGwire, M. E. Drew and A. R. Satoskar share equal contribution in authorship.
Rights and permissions
About this article
Cite this article
Walker, D.M., Oghumu, S., Gupta, G. et al. Mechanisms of cellular invasion by intracellular parasites. Cell. Mol. Life Sci. 71, 1245–1263 (2014). https://doi.org/10.1007/s00018-013-1491-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00018-013-1491-1