
Summary
The effect of 19 redox-systems on the motility and the photo-phobotactic reactions of Phormidium uncinatum has been investigated. The E ′0 -values of these substances covered the range between-0.44 and +0.36 V. As a measure of the inhibitory effect the ED 50, i.e. the concentration in m that inhibits the movement and the phototactic reactions to an extent of 50% of the control, has been used.
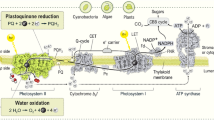
Substances with an E ′0 between ±0 and about +0.1 V strongly inhibit the phobotactic responses. The maximum inhibition, an ED 50 of 2×10-7 m, is shown by thionin, which has a redox-potential of +0.064 V. A second maximum of inhibition, smaller than the first, lies below-0.25 V, i.e. the range of viologenes and safranines. The most effective of these redox-systems is methyl-viologene (-0.44V) with an ED 50 of 3.6×10-6 m. Substances with redox-potentials between-0.25 and ±0 V and more than +0.1 V are less effective though not quite in-effective. Although every redox-system that is able to trap electrons from the electron transport chain of photosynthesis can influence the photo-phobotactic sensitivity, redox-systems with potentials of about-0.4 and between ±0 and +0.1 V are most effective. Obviously, the regions of electron transfer from the two pigment systems to the next electron acceptors, i.e. from the quencher to quinone and from Z to ferredoxin, are most sensitive to external redox-systems. Thus, we have to correct our concept of coupling between photosynthesis and phototaxis in as much as the linkage is not restricted to the second light reaction though the E ′0 range between quencher and quinone or cytochrome b 6 is the most sensitive point. Contrary to the hypothesis of Links, photo-phobotaxis is not coupled with photo-phosphorylation but with the electron flow along the electron transport chain of photosynthesis, and the phobic responses are due to sudden changes in the steady state of the electron flow caused by sudden changes in light intensity, and hence in the redox-state of the cell.
As every trapping of electrons from the transport chain of photosynthesis also decreases the efficiency of the phosphorylation apparatus, the redox-systems are expected to impair photokinesis, too. In fact, all substances investigated inhibit photokinesis to various degrees. There are also two maxima of inhibition. But contrary to the results of the phobotactic experiments, the redox-systems with highest negative E ′0 -values are most effective in inhibiting photokinesis, and the second smaller maximum lies between +0.06 and +0.08 V, i.e. the redox-potential of phenazine methosulphate, thionin and plastoquinone. Methylene blue (+0.011 V) and toluidine blue (+0.034 V) which strongly inhibit photo-phobotaxis (ED 50 at or below 10-6 m) have only a small effect on photokinesis (ED 50 at about 4×10-4 m). Thus, such redox-systems which are able to trap electrons from the cyclic electron transport, via ferredoxin, cytochrome b 6 or quinone, are most effective in inhibiting photokinesis. This finding is consistent with our concept that under normal conditions cyclic photophosphorylation is the main energy source of a photokinesis.
All the redox-systems evaluated inhibit, more or less, movement in the dark. Most effective are the safranines: safranine and phenosafranine, the thiazines: methylene blue, toluidine blue and thionin, and phenazine metho-sulphate. The ED 50-values of these substances are in the range between 10-6 and 10-5 m. Thus, the maxima of inhibition are shown by substances the redox-potentials of which are slightly more positive than the E ′0 -values of the native electron carriers of the respiratory chain, NAD, flavoprotein, and cytochrome b. Obviously, artificial electron acceptors may trap electrons from the respiratory chain, by-passing the oxydative phosphorylation. Consequently, no ATP is available, and the movement in the dark ceases.
Zusammenfassung
Es wurde der Einfluß von 19 Redox-Systemen, deren Normalpotentiale im Bereich zwischen-0,44 V und +0,36 V liegen, auf die Bewegungsaktivität im Licht und im Dunkeln sowie auf die photo-phobotaktische Reaktion von Phormidium uncinatum untersucht. Für jede Substanz wurde bei allen drei Reaktionstypen die Dosis-Effekt-Kurve bestimmt und die ED 50 ermittelt.
Die Photo-phobotaxis wird am stärksten durch Redox-systeme mit einem E ′0 zwischen ±0 und etwa +0,1 V gehemmt. Die maximale Wirksamkeit zeigt das Thionin mit einer ED 50 von 2·10-7 m. Ein zweites Hemmungsmaximum liegt im Bereich der Viologene. Von diesen hemmt das Methyl-viologen mit einer ED 50 von 3,6·10-6 m am stärksten. Substanzen mit Redox-Potentialen zwischen-0,25 und ±0 und mehr als +0,1 V sind schwächer wirksam, wenn auch keineswegs unwirksam. Offenbar wird die Photo-phobotaxis durch alle Redox-Systeme gehemmt, die aufgrund ihres Redox-Potentials Elektronen aus der Elektronentransportkette der Photosynthese abziehen können. Die stärkste Wirkung zeigen jedoch solche Substanzen, deren Redox-Potentiale die gleiche Größenordnung haben wie die der ersten Elektronen-Acceptoren der beiden Lichtreaktionen, d. h. Plastochinon und Ferredoxin.
Die Empfindlichkeit der Photokinese gegen die Viologene ist etwa von der gleichen Größenordnung wie im Falle der Photo-phobotaxis. Mit abnehmender Negativität des E ′0 der geprüften Redox-Systeme wird der Hemmungseffekt jedoch rasch geringer, um zwischen ±0 und +0,05 V ein Minimum zu erreichen. Ein zweites Maximum der Wirkung auf die Photokinese wird zwischen +0,06 und +0,08 V erreicht, d. h. im E ′0 -Bereich von Thionin, PMS und Plastochinon. Allerdings liegt es um etwa eine Zehnerpotenz niedriger als das Hauptmaximum im Viologenbereich. Die stärkste Wirksamkeit auf die Photokinese zeigen also solche Substanzen, die auf dem Wege über das Ferredoxin, das cytochrom b 6 oder das Plastochinon dem cyclischen Elektronentransport Elektronen entziehen und dadurch die cyclische Photophosphorylierung hemmen können.
Auch die Hemmung der Dunkelbewegung zeigt eine deutliche Abhängigkeit vom Redox-Potential. Am stärksten hemmen solche Redox-Systeme, deren Redox-Potential zwischen-0,3 und-0,2 V sowie zwischen ±0 und +0,1 V liegt. Offenbar konkurrieren sie mit den natürlichen Gliedern der Atmungskette, NAD, Flavoprotein und Cytochrom b um die Elektronen und schließen hierdurch die Atmungskette kurz, was zu einer Hemmung der oxydativen Phosphorylierung und damit der Bewegung im Dunkeln führt.
Similar content being viewed by others


Literatur
Arnon, D. I.: Cell-free photosynthesis and the energy conversion process. In: Light and life (William D. McElroy and Bentley Glass, eds.), pp. 489–569. Baltimore: The John Hopkins Press 1961.
—, M. B. Allen, and F. R. Whatley: Photosynthesis by isolated chloroplasts. IV. General concept and comparison of three photochemical reactions. Biochim. biophys. Acta (Amst.) 20, 449–461 (1956).
Clark, W. M.: Oxidation-reduction potentials of organic systems. Baltimore: Williams & Wilkins 1960.
Clayton, R. K.: Patterns of accumulation resulting from taxes and changes in motility of micro-organisms. Arch. Mikrobiol. 27, 311–319 (1957).
Geller, D. M., and F. Lipmann: Photophosphorylation in extracts of Rhodospirillum rubrum. J. biol. Chem. 235, 2478–2484 (1960).
Judah, J. D., and G. Williams-Ashman: The inhibition of oxidative phosphorylation. Biochem. J. 48, 33–42 (1951).
Kok, B.: Concentration and normal potential of primary photooxidants and reductants in photosynthesis. In: Currents in photosynthesis (J. B. Thomas and J. C. Goedheer, eds.), pp. 383–392. Rotterdam: A. D. Donker 1965.
Lehninger, A. L., and L. F. Remmert: An endogenous uncoupling and swelling agent in liver mitochondria and its enzymic formation. J. biol. Chem. 234, 2459–2471 (1959).
Links, J.: A hypothesis for the mechanism of (phobo) chemotaxis. Diss., Leiden 1955
Löw, H., P. Siekevitz, L. Ernster, and O. Lindberg: Studies on the relation of the adenosinetriphosphate-inorganic phosphate exchange of mitochondria to oxidative phosphorylation. Biochim. biophys. Acta (Amst.) 29, 392–405 (1958).
Losada, M., and D. I. Arnon: Selective inhibitors of photosynthesis. In: Metabolic inhibitors (R. M. Hochster and J. H. Quastel, eds.), vol. 2, pp. 559–593. New York and London: Academic Press 1963.
Nultsch, W.: Der Einfluß des Lichtes auf die Bewegung der Cyanophyceen. III. Mitteilung. Photophobotaxis von Phormidium uncinatum. Planta (Berl.) 58, 647–663 (1962).
—: Phototaktische Aktionsspektren von Cyanophyceen. Ber. dtsch. bot. Ges. 75, 443–453 (1963).
—: Light reaction systems in cyanophyceae. Photochem. Photobiol. 4, 613–619 (1965).
—: Über den Antagonismus von Atebrin und Flavinnucleotiden im Bewegungs-und Lichtreaktionsverhalten von Phormidium uncinatum. Arch. Mikrobiol. 55, 187–199 (1966).
—: Untersuchungen über den Einfluß von Entkopplern auf die Bewegungsaktivität und das phototaktische Reaktionsverhalten blaugrüner Algen. Z. Pflanzenphysiol. 56, 1–11 (1967).
—, u. Jeeji-Bai: Untersuchungen über den Einfluß von Photosynthese-Hemmstoffen auf das phototaktische und photokinetische Reaktionsverhalten blaugrüner Algen. Z. Pflanzenphysiol. 54, 84–98 (1966).
—, u. G. Richter: Aktionsspektren des photosynthetischen 14CO2-Einbaus von Phormidium uncinatum. Arch. Mikrobiol. 47, 207–213 (1963).
Racker, E.: Mechanisms of synthesis of adenosinetriphosphate. Advanc. Enzymol. 23, 323–399 (1961).
Rauen, H. M.: Biochemisches Taschenbuch, Teil 2, 2. Aufl. Berlin-Göttingen-Heidelberg-New York: Springer 1964.
Trebst, A.: Neuere Vorstellungen über den Mechanismus der Photosynthese. Ber. dtsch. bot. Ges. 77, 123–143 (1964).
Whatley, F. R., M. B. Allen, and D. I. Arnon: Photosynthesis by isolated chloroplasts. VII. Vitamin K and riboflavin phosphate as cofactors of cyclic photophosphorylation. Biochim. biophys. Acta (Amst.) 32, 32–46 (1959).
Witt, H. T., B. Rumberg, P. Schmidt-Mende, U. Sigel, B. Skerra, J. Vater u. J. Weikard: Über die Analyse der Photosynthese mit Blitzlicht. Angew. Chem. 77, 821–876 (1965).
—, B. Skerra, and J. Vater: Conditions for the isolation of reaction cycle. II. Rate of water splitting, location of the rate determining step of the overall reaction: the electron carrier Y. In: Currents in photosynthesis (J. B. Thomas and J. C. Goedheer, eds.), pp. 273–292. Rotterdam: A. D. Donker 1965.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Nultsch, W. Einfluß von Redox-Systemen auf die Bewegungsaktivität und das phototaktische Reaktionsverhalten von Phormidium uncinatum . Archiv. Mikrobiol. 63, 295–320 (1968). https://doi.org/10.1007/BF00412118
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF00412118