
Summary
Polytene chromosomes in cells of salivary gland, Malpighian tubules and intestine of Rhynchosciara angelae are very favorable for study. The polytene chromosomes of the salivary gland are among the largest available for cytogenetics work. The ones in Malpighian tubules and in some parts of the intestine are as large and as favorable for cytological studies as the salivary chromosomes of many species of Drosophila.
Two additional characteristics of Rhynchosciara make these flies excellent material for studies on the development of polytene chromosomes. 1.It is possible to observe the banding pattern of the polytene chromosomes at many stages of the larval life for at least 30 days before pupation, and 2. since the gregarious larvae develop simultaneously, one can sample the group at any stage desired. Sampling the group every day, it is possible to follow the development of the chromosomes as though one studied a single individual by observing it every day.
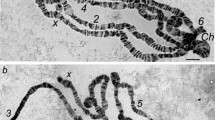
We have followed in detail the behavior of the bands in two sections of chromosome B and in one section of chromosome C, at different stages of larval development. Some regions of the chromosomes which are represented by typical euchromatic bands at one stage of the larval development may develop in enormous bulbs, and later on may return to the banded stage again.
The formation of the bulbs is not uniform in different sections of the same or of different chromosomes. In section 2 of chromosome B a certain locus swells enormously and then develops an enormous bulb, and later returns to the banded stage. At the point where the bulb was formed there is an accumulation of DNA, in amounts probably several times greater than before the bulb formation. In section 3 of chromosome B and section 3 of chromosome C the extra accumulation of DNA preceeds the formation of the bulb and is maintained during and after it. In the bulb formed in section 3 of chromosome C a single band seems to be responsible for the process.
As shown by several authors, experimental evidence suggests that a gene is located within a band. The bulb formation in polytene chromosomes may then be morphological evidence of gene activities. This type of bulb formations and of return to the banded stage is a property of many chromosomes bands, during larval development. This type of behavior of many bands in polytene chromosomes is related to the process of nucleolus formation. However, this behavior may be found in almost all (if not in all) bands of the polytene chromosomes. If so, the behavior of the nucleolus organizer region is only a special case of this general process.
The accumulation of DNA in different parts of the chromosome in cells of the same or of different tissues may be an argument against the theory of the constancy of the amount of DNA in all cells of a species. The bulb formations is not peculiar to R. angelae but occurs in several other Diptera.
Similar content being viewed by others


Bibliography
Alfert, M., and H. Swift: Nuclear DNA constancy: A critical evaluation of some exceptions reported by Lison and Pasteels, Exper. Cell Res. 5, 455–460 (1953).
Bauer, H: Die Chromosomen im Soma der Metazoen. Verh. Dtsch. Zool. Ges. Freiburg 1952.
- Die Istanbuler Hypothese von Entwicklung und Bau der Riesenchromosomen. Verh. Dtsch. Zool. Ges. Freiburg 1952.
Bauer, H., u. W. Beermann: Die Polytänie der Riesenchromosomen. Chromosoma 4, 630 bis 648 (1952).
Beermann, W.: Chromosomenstruktur und Zelldifferenzierung in der Speicheldrüse von Trichocladius vitripennis. Z. Naturforsch. 7b, 237–242 (1952).
— Chromomerenkonstanz und spezifische Modifikationen der Chromosomenstruktur in der Entwicklung und Organdifferenzierung von Chironomus tentans. Chromosoma 5, 139–198 (1952).
Boivin, A., R. Vendrely et C. Vendrely: L'Acide désozyribonucléique du noyau cellulaire, dépositaire des caractères héréditaires; arguments d'ordre analytique. C. r. Acad. Sci. Paris 226, 1061–1063 (1948).
Breuer, M. E., e C. Pavan: Gens na differenciação. Ciência e Cultura 4, 141 (1952).
- Salivary chromosomes and differentiation. Proc. IX. Internat. Congr. Genetics 1953.
Bridges, C. B.: Salivary chromosome maps. J. Hered. 26, 60–64 (1935).
Bridges, C. B.: A revised map of the salivary gland X chromosome of Drosophila melanogaster. J. Hered. 29, 11–13 (1938).
Demerec, M.: Cytology, Genetics and Evolution. Philadelphia: Univ. Pennsylvania Press 1941.
Dobzhansky, Th.: Distribution of Heterochromatin in the chromosomes of Drosophila pallidipennis. Amer. Naturalist 78, 193–213 (1944).
Dreyfus, A., E. Nonato, M. E. Breuer e C. Pavan: Cromosomas politenicos em vários orgãos de „Rhynchosciara angelae“. Rev. brasil. Biol. 11, 439–450 (1951).
Heitz, E., u. H. Bauer: Beweise für die Chromosomennatur der Kernschleifen in den Knäuelkernen von Bibio nortulanus L. Cytologische Untersuchungen an Dipteren. I. Z. Zellforsch. 17, 67–82 (1933).
Kosswig, C.: Demonstration of preparations of salivary chromosomes of different tissues of Chironomus. Proc. 8. Internat. Congr. Genetics 1948, 609.
Kosswig, C., and A. Şengun: Intraindividual variability of chromosome IV of Chironomus. J. Hered. 38, 235–239 (1947).
Lewis, E. B.: The relation of repeats to position effect in Drosophila melanogaster. Genetics 30, 137–166 (1945).
Mechelke, R.: Reversible Strukturmodifikationen der Speicheldrüsenchromosomen von Acricotopus lucidus. Chromosoma 5, 511–543 (1953).
Metz, C. W.: Structure of salivary gland chromosomes. Symposia Quant. Biol. 9, 23–39 (1941).
Painter, T. S.: A new method for the study of chromosome rearrangements and the plotting of chromosome maps. Science 78, 585–586 (1933).
—: A new method for the study of chromosome aberrations and the plotting of chromosome maps in Drosophila melanogaster. Genetics 19, 175–188 (1934).
—: An experimental study of salivary chromosomes. Symposia Quant. Biol. 9, 47–54 (1941).
Pavan, C., and M. Breuer: Polytene chromosomes in different tissues of Rhynchosciara. J. Hered. 43, 150–157 (1952).
Poulson, D. F., and C. W. Metz: Studies on the structure of nucleolus forming regions and related structures in the giant salivary gland chromosomes of Diptera. J. of Morph. 63, 363–395 (1938).
Ris, H., and A. E. Mirsky: Quantitative cytochemioal determination of desoxyribosenuoleic acid with the Feulgen nuoleal reaction. J. Gener. Physiol. 33, 125 to 145 (1949).
Şengün, A.: Difference in structure between the same giant chromosomes from the same larvae of Drosophila repleta. Nature (Lond.) 163, 1002 (1949).
—: Vergleichend-ontogenetische Untersuchungen über die Riesenchromosomen verschiedener Gewebearten der Chironomiden. Istanbul Üniv. fen Fak. Mecmuasi, Ser. B, Cilt XVI Sayi 1, 1–44 (1951).
—: Meiotische und somatische Chromosomen von Chironomus-Larven. Istanbul Üniv. fen Fak. Mecmuasi, Ser. B, Cilt XVI Sayi 4, 345–350 (1951).
Şengün, A., u. C. Kosswig: Weiteres über den Bau der Riesenchromosomen in verschiedenen Geweben von Chironomus-Larven. Chromosoma 3, 195–207 (1948).
Slizynsky, B. M.: Chironomus versus Drosophila. J. Genet. 50, 77–78 (1950).
Stern, C.: Two or three bristles. Amer. Sci. 42, 213–247 (1954).
Swift, H. H.: The desoxyribose nucleic acid content of animal nuclei. Physiol. Zool. 22, 169–198 (1950).
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Breuer, M.E., Pavan, C. Behavior of polytene chromosomes of rhynchosciara angelae at different stages of larval development. Chromosoma 7, 371–386 (1955). https://doi.org/10.1007/BF00329733
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF00329733