
Summary
-
1.
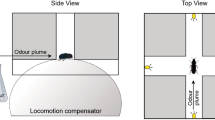
The tenebrionid beetles Pimelia grosso, P. tenuicornis, Tenebrio molitor, the dung beetles Geotrupes silvaticus and G. stercorarius can, under specific conditions, use pure anemomenotactic orientation (Figs. 1–8). They prefer running-angles which have relatively narrow angular deviations, left and right, to the basic directions (Figs. 3, 4, 26).
-
2.
The preferences for particular angle sizes do not constantly alter with respect to the times of day, temperature (between 18 and 28° C) or feeding conditions (Figs. 8–11).
-
3.
The lowest wind velocities which will evoke menotactic orientation lie at about 0.15 m/sec (Geotrupes) and 0.4 m/sec (Pimelia). The upper limit for anemomenotactic orientation are velocities higher than those against which the animal can move.
-
4.
Only stimulus direction, not strength is evaluated for menotaxis (Figs. 13–15).
-
5.
The continuity of the air stream is not a hard and fast requirement for anemomenotactic orientation: beetles can also orient themselves to short puffs of wind (Figs. 17, 19, 21). During still air, between puffs, the beetles characteristically deviate from their adopted course (Fig. 18). However the more puffs and intervals the less the deviation becomes (Fig. 20).
-
6.
The anemomenotactic orientation is not greatly disturbed in partly turbulent air (for example over rough ground)(Fig. 22).
-
7.
The sense organ which allows the intact beetle to detect wind direction is situated in the pedicellus-flagellum joint. In Geotrupes the loss of the clubs does not have a detrimental effect on either choice of direction or angular preference, neither does it result in significantly larger standard deviations. 2 segments of the flagellum allow the beetle to anemomenotactically orientate to wind directions when the air stream is moving at, or above, 1 m/sec (Table 3).
-
8.
3 dung beetles were able to orient themselves to wind directions 4 weeks respectively 4 months after antennal amputation (Fig. 23, Table 1).
-
9.
Beetles are able to transpose their running-angles intramodally (Figs. 28, 29). This ability is most obvious in experimentally naive animals whose locomotion is flight motivated. Of the four possible directions they choose the one nearest to that of their long axis after they have been placed on the substrate (Figs. 25, 26).
-
10.
The existence and working direction of a course order, and the participation of both basic orientations in anemomenotaxis are proven (Fig. 32). The ability to provide a sustained course order size (that can never deviate over the 90° limit of the basic directions) with different signs (of the turning direction) and the possibility to reverse taxis explains the orientation physiology of the four intramodal transpositions.
-
11.
A compensation mechanism is suggested as the basis for anemomenotaxis of beetles by the experimental evidence in this account. Two main objections against such a mechanism are debated in the discussion.
-
12.
The possible biological significances of anemomenotaxis are discussed. It is proposed that this taxis has its most important significance in appetetive behavior of beetles that are in readiness for an olfactory sign-stimulus. It is proposed that anemomenotaxis can also fullfill the basic rôle of space orientation (for example, compass true alignment during flight).
Zusammenfassung
-
1.
Die Feistkäfer Pimelia grossa, P. tenuicornis, der Mehlkäfer Tenebrio molitor, die Mistkäfer Geotrupes silvaticus und G. stercorarius können sich unter entsprechenden Bedingungen rein anemomenotaktisch orientieren (Abb. 1–8). Sie bevorzugen Laufwinkel, die in relativ enge Winkelbereiche rechts und links der beiden Grundrichtungen führen (Abb. 3, 4, 26).
-
2.
Die Bevorzugung bestimmter Winkelgrößen ändert sich bei Geotrupes nicht gesetzmäßig mit der Tageszeit, der Temperatur (im Bereich 18–28° C) oder dem Fütterungszustand (Abb. 8–11).
-
3.
Die untere Grenze der Windstärke, die eine menotaktische Einstellung ermöglicht, liegt für die Mistkäfer bei etwa 0,15 m/sec, für die Feistkäfer bei etwa 0,4 m/sec. Die obere Grenze befindet sich bei Geschwindigkeiten, die den Käfern ein Vorwärtskommen unmöglich machen.
-
4.
Bei der menotaktischen Einstellung wird nur die Beizrichtung nicht aber die Reizstärke bewertet (Abb. 13–15).
-
5.
Die Kontinuität des Luftstroms ist keine Voraussetzung für die anemomenotaktische Orientierung: Die Käfer orientieren sich auch nach kurzen Windstößen (Abb. 17, 19, 21). Während der Windstille kommt es zu regelhaften Abweichungen von dem bei Wind eingehaltenen Kurs (Abb. 18). Das Ausmaß dieser Abweichungen wird nach häufigen Windunterbrechungen stark verringert (Abb. 20).
-
6.
Gegen Turbulenzen des Luftstroms, wie sie über unebenem Untergrund entstehen, ist die Anemomenotaxis der Käfer nicht sehr anfällig (Abb. 22).
-
7.
Die Sinnesorgane, die dem intakten Käfer die Windrichtungsbestimmung ermöglichen, sprechen auf Bewegungen im Pedicellus-Flagellumgelenk an. Ein Verlust der Endkolben hat beim Mistkäfer keinen Einfluß auf die Richtungs- und Winkelgrößenwahl, auch die Streuung wird nicht signifikant größer. 2 Flagellenglieder pro Antenne ermöglichen bei Windgeschwindigkeiten um oder über 1 m/sec noch eine anemomenotaktische Orientierung (Tabelle 3).
-
8.
Bei 3 Mistkäfern, deren Fühler 4 Wochen bzw. 4 Monate vor dem Versuch entfernt worden waren, konnte wieder eine Orientierung nach der Windrichtung nachgewiesen werden (Abb. 23, Tabelle 1).
-
9.
Die Käfer können Laufwinkel intramodal vierdeutig transponieren (z.B. Abb. 28, 29). Am deutlichsten tritt diese Fähigkeit bei Versuchsneulingen zutage, deren Läufe rein fluchtmotiviert sind: Sie wählen normalerweise denjenigen der 4 möglichen Laufwinkel, der der Aufsetzrichtung am nächsten liegt (vgl. Abb. 25, 26).
-
10.
Die Existenz und die Wirkungsrichtung eines Drehkommandos, sowie die Beteiligung beider Grundorientierungen an der Anemomenotaxis werden nachgewiesen (Abb. 31). Die Fähigkeit, eine gleichbleibende Drehkommandogröße (die nie zu einer stärkeren Abweichung als 90° von einer Grundrichtung führen kann) mit verschiedenem Vorzeichen der Drehrichtung versehen zu können und die Möglichkeit zur Taxisumkehr (Abb. 32) erklären die orientierungsphysiologische Seite des vierdeutigen intramodalen Transponierens.
-
11.
Versuchsergebnisse, die Aussagen über den physiologischen Mechanismus der Anemomenotaxis der Käfer zulassen, sprechen für einen Kompensationsmechanismus. Die gegen die Kompensationstheorie der Menotaxis (Jander, 1957) vorgebrachten Argumente werden im Rahmen der bisherigen Resultate kurz diskutiert.
-
12.
Die möglichen biologischen Bedeutungen der Anemomenotaxis werden besprochen. Es wird angenommen, daß sie beim Appetenzverhalten des nach geruchlichen Schlüsselreizen „suchenden“ Käfers ihre biologisch wichtigste Aufgabe erfüllt. Sie kann auch die basalen Aufgaben einer Raumorientierung übernehmen und so z. B. kompaßtreue Fluchtkurse steuern.
Similar content being viewed by others


Literatur
Bässler, U.: Versuche zur Orientierung der Stechmücken: Die Schwarmbildung und die Bedeutung des Johnstonschen Organs. Z. vergl. Physiol. 41, 300–330 (1958).
—: Zum Einfluß von Schwerkraft und Licht auf die Ruhestellung der Stabheuschrecke (Carausius morosus). Z. Naturforsoh. 17b, 477–480 (1962).
Batschelet, E.: Statistical methods for the analysis of problems in animal orientation and certain biological rhythms. American Institute of Biological Sciences (AIBS Monograph), Washington 1965.
Birukow, G.: Photo-Geomenotaxis bei Geotrupes silvaticus Panz. und ihre zentralnervöse Koordination. Z. vergl. Physiol. 36, 176–211 (1954).
—: Zur Punktion der Antennen beim Mistkäfer (Geotrupes silvaticus). Z. Tierpsychol. 15, 265–276 (1958).
Beaemer, W.: Verhaltensphysiologische Untersuchungen am optischen Apparat bei Fischen. Z. vergl. Physiol. 39, 374–398 (1957).
Buddenbrock, W. v.: Untersuchungen über den Schattenreflex. Z. vergl. Physiol. 13, 164–213 (1931).
Burkhardt, D., u. G. Schneider: Die Antennen von Calliphora als Anzeiger der Fluggeschwindigkeit. Z. Naturforsch. 12b, 139–143 (1957).
Creutzberg, F.: The role of tidal streams in the navigation of migrating elvers (Anguilla vulgaris Turt.). Ergebn. Biol. 26, 118–127 (1963).
Czeloth, H.: Untersuchungen über die Raumorientierung von Triton. Z. vergl. Physiol. 13, 74–163 (1931).
Danzer, A.: Die Duftorientierung von Geotrupes silvaticus im natürlichen Biotop. Z. vergl. Physiol. 39, 76–83 (1956).
Daykin, P. N.: Orientation of Aedes aegypti in vertical air currents. Canad. Entomol. 99, 303–308 (1967).
Doflein, I.: Chemotaxis und Rheotaxis bei Plannarien. Ein Beitrag zur Reizphysiologie und Biologie der Süßwassertrikladen. Z. vergl. Physiol. 3, 62–112 (1926).
Eggers, F.: Zur Kenntnis der antennalen stiftführenden Sinnesorgane der Insekten. Z. Morph. Ökol. Tiere 2, 259–349 (1924).
Flügge, Ch.: Geruchliche Raumorientierung von Drosophila melanogaster. Z. vergl. Physiol. 20, 463–501 (1934).
Fraenkel, G. S., and D. L. Gunn: The orientation of animals. New York: Dover 1961.
Geisler, M.: Untersuchungen zur Tagesperiodik des Mistkäfers Geotrupes silvaticus Panz. Z. Tierpsvchol. 18, 389–420 (1961).
Gewecke, M.: Die Wirkung von Luftströmung auf die Antennen und das Flugverhalten der Blauen Schmeißfliege (Calliphora erythrocephala). Z. vergl. Physiol. 54, 121–164 (1967).
—: Der Bewegungsapparat der Antennen von Calliphora erythrocephala. Z. Morph. Ökol. Tiere 59, 95–133 (1967).
Haaften, J. L. van, and J. Verwey: The role of water currents in the orientation of marine animals. Arch. Néerl. Zool. 13, 493–499 (1960).
Hangartner, W.: Spezifität und Inaktivierung des Spurhormons von Lasius fuliginosus Latr. und Orientierung der Arbeiterinnen im Duftfeld. Z. vergl. Physiol. 57, 103–136 (1967).
Hartung, E.: Untersuchungen über die Geruchsorientierung von Calliphora erythrocephala. Z. vergl. Physiol. 22, 119–144 (1935).
Hassenstein, B.: Kybernetik und biologische Forschung. Handbuch der Biologie, Bd. I, S. 629–719. Frankfurt a. M.: Akad. Verl.-Ges. Athenaion 1966.
Heran, H.: Wahrnehmung und Regelung der Flugeigengeschwindigkeit bei Apis mellifica L. Z. vergl. Physiol. 42, 103–163 (1959).
—: Anemotaxis und Fluchtorientierung des Bachläufers Velia caprai Tam. Z. vergl. Physiol. 46, 129–149 (1962).
Holst, E. von: Die Arbeitsweise des Statolithenapparates bei Fischen. Z. vergl. Physiol. 32, 60–120 (1950).
Jander, R.: Die optische Richtungsorientierung der roten Waldameise (Formica rufa). Z. vergl. Physiol. 40, 162–238 (1957).
—: Grundleistungen der Licht- und Schwereorientierung von Insekten. Z. vergl. Physiol. 47, 381–430 (1963).
—, u. Ch. Voss: Die Bedeutung von Streifenmustern für das Formensehen der Roten Waldameise (Formica rufa L.). Z. Tierpsychol. 20, 1–9 (1963).
Koehler, O.: Beiträge zur Sinnesphysiologie der Süßwasserplanarien. Z. vergl. Physiol. 16, 606–756 (1932).
Lindauer, M., u. H. Martin: Über die Orientierung der Biene im Duftfeld. Naturwissenschaften 50, 509–514 (1963).
Linsenmair, K. E.: Anemomenotaktische Orientierung bei Skorpionen (Chelicerata, Scorpiones). Z. vergl. Physiol. 60, 445–449 (1968).
Linsenmair-Ziegler, Ch.: Vergleichende Untersuchungen zum photogeotaktischen Winkeltransponieren pterygoter Insekten. Diss. Frankfurt a. M. 1969.
Markl, H.: Geomenotaktische Fehlorientierung bei Formica polyctena Förster. Z. vergl. Physiol. 48, 552–586 (1964).
—: Schwerkraftdressuren an Honigbienen. I. Die geomenotaktische Fehlorientierung. Z. vergl. Physiol. 53, 353–371 (1966).
Martin, H.: Zur Nahorientierung der Biene im Duftfeld. Zugleich ein Nachweis für die Osmostropotaxis bei Insekten. Z. vergl. Physiol. 48, 481–533 (1964).
Mittelstaedt, H.: Probleme der Kursregelung bei freibeweglichen Tieren. Aufnahme und Verarbeitung von Nachrichten durch Organismen, S. 138–148. Stuttgart: Hirzel 1961.
—: Control Systems of orientation in insects. Ann. Rev. Entomol. 7, 177–198 (1962).
—: Bikomponententheorie der Orientierung. Ergebn. Biol. 26, 253–258 (1963).
Murr-Danielczick, L.: Über den Geruchssinn der Mehlmottenschlupfwespe Habrobracon juglandis Ashmead. Z. vergl. Physiol. 11, 210–270 (1930).
Otto, E.: Untersuchungen über die Frage der geruchlichen Orientierung bei Insekten. Zool. Jb., Abt. allg. Zool. u. Physiol. 62, 65–92 (1951).
Schanz, M.: Der Geruchssinn des Kartoffelkäfers (Leptinotarsa decemlineata Say). Z. vergl. Physiol. 35, 353–379 (1953).
Schlegel, P.: Einzelableitungen von einem Stellungsreceptor im Pedicellus-Funiculus-Gelenk des Blauen Brummers (Calliphora vicina Rob. Desv., erythrocephala auct.). Z. vergl. Physiol. 55, 278–285 (1967).
Schneider, D.: Elektrophysiologische Untersuchungen von Chemo- und Mechanoreceptoren der Antenne des Seidenspinners Bombyx mori L. Z. vergl. Physiol. 40, 8–41 (1957).
Schneider, P.: Vergleichende Untersuchungen zur Steuerung der Fluggeschwindigkeit bei Calliphora vicina Rob.-Desvoidy (Diptera). Z. wiss. Zool. 173, 114–173 (1965).
Schöne, H.: Die Lichtorientierung der Larven von Acilius sulcatus L. und Dytiscus marginalis L. Z. vergl. Physiol. 33, 63–98 (1951).
—: Die Lageorientierung mit Statolithenorganen und Augen. Ergebn. Biol. 21, 161–209 (1959).
—: Optisch gesteuerte Lageänderungen (Versuche an Dytiscidenlarven zur Vertikalorientierung). Z. vergl. Physiol. 45, 590–604 (1962).
Schwinck, I.: Experimentelle Untersuchungen über Geruchssinn und Strömungswahrnehmung in der Orientierung bei Nachtschmetterlingen. Z. vergl. Physiol. 37, 19–57 (1954).
—: Weitere Untersuchungen zur Frage der Geruchsorientierung der Nachtschmetterlinge: Partielle Fühleramputationen bei Spinnermännchen, insbesondere am Seidenspinner Bombyx mori L. Z. vergl. Physiol. 37, 439–458 (1955).
Steiner, G.: Zur Duftorientierung fliegender Insekten. Naturwissenschaften 40, 514–515 (1953).
—: Über die Geruchsfernorientierung von Drosophila melanogaster in ruhender Luft. Naturwissenschaften 41, 287 (1954).
Tenckhoff-Eikmanns, I.: Licht- und Erdschwereorientierung beim Mehlkäfer Tenebrio molitor L. und bei einigen anderen Insekten. Zool. Beiträge, N.F. 4, 307–341 (1959).
Traynier, R. M. M.: Sex attraction in the mediterranean flour moth, Anagasta küniella: location of the female by the male. Canad. Entomol. 100, 5–10 (1968).
Vowles, D. M.: The orientation of ants. I. The substitution of stimuli. J. exp. Biol. 81, 341–355 (1954).
—: The orientation of ants. II. Orientation to light, gravity and polarized light. J. exp. Biol. 31, 356–375 (1954).
Warnke, G.: Experimentelle Untersuchungen über den Geruchssinn von Geotrupes silvaticus Panz. und Geotrupes vernalis L. Z. vergl. Physiol. 14, 121–199 (1931).
Wisby, W. J., and A. D. Hasler: Effect of olfactory occlusion on migrating silver salmon (O. kisutch). J. Fish. Res. Board Canada 11, 472–478 (1954).
Author information
Authors and Affiliations
Additional information
Mit dankenswerter Unterstützung durch die Deutsche Forschungsgemeinschaft. Für die kritische Durchsicht des Manuskriptes danke ich Herrn Dr. R. Jander.
Rights and permissions
About this article
Cite this article
Eduard Linsenmair, K. Anemomenotaktische Orientierung bei Tenebrioniden und Mistkäfern (Insecta, Coleopterd). Z. Vergl. Physiol. 64, 154–211 (1969). https://doi.org/10.1007/BF00297944
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF00297944