
Abstract
Chemical communication plays many key roles in mammalian reproduction, although attention has focused particularly on male scent signalling. Here, we review evidence that female chemical signals also play important roles in sexual attraction, in mediating reproductive competition and cooperation between females, and in maternal care, all central to female reproductive success. Female odours function not only to advertise sexual receptivity and location, they can also have important physiological priming effects on male development and sperm production. However, the extent to which female scents are used to assess the quality of females as potential mates has received little attention. Female investment in scent signalling is strongly influenced by the social structure and breeding system of the species. Although investment is typically male-biased, high competition between females can lead to a reversed pattern of female- biased investment. As among males, scent marking and counter-marking are often used to advertise territory defence and high social rank. Female odours have been implicated in the reproductive suppression of young or subordinate females across a range of social systems, with females of lower competitive ability potentially benefiting by delaying reproduction until conditions are more favourable. Further, the ability to recognise individuals, group members and kin through scent underpins group cohesion and cooperation in many social species, as well as playing an important role in mother-offspring recognition. However, despite the diversity of female scent signals, chemical communication in female mammals remains relatively understudied and poorly understood. We highlight several key areas of future research that are worthy of further investigation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A defining feature of all mammalian taxa is high female investment in reproduction through the process of lactation (Wade and Schneider 1992). Gestation, which occurs in all species except monotremes, further increases female investment, as does parental care, which is provided solely by females in around 90% of mammalian species (Royle et al. 2014; Wade and Schneider 1992; West and Capellini 2016). The high costs of gestation and lactation mean that female reproduction is often constrained by food availability (Gittleman and Thompson 1988; Wade and Schneider 1992; West and Capellini 2016), resulting in female competition for access to food sources as well as other limiting resources such as space or shelter (Clutton-Brock 2009; Stockley and Bro-Jorgensen 2011). In addition to securing enough resources to successfully raise their offspring, females also need to ensure they mate with high quality males (Andersson 1994). This can benefit females through access to male resources, such as parental care or territory, and / or by gaining genetic benefits for their offspring (Andersson 1994; Clutton-Brock 2016; Kokko et al. 2003; Møller and Thornhill 1998). The importance of male quality to female reproduction has led to a focus on male signals used to attract females or to mediate competition between rival males (Clutton-Brock and McAuliffe 2009; Maynard Smith and Harper 2003; Wong and Candolin 2005). However, there is growing understanding that females also use signals for sexual attraction, to mediate female competition and cooperation, and to facilitate maternal behaviours (Clutton-Brock and Huchard 2013; Nowak et al. 2000; Stockley and Bro-Jorgensen 2011; Stockley et al. 2013).
Female chemical signals provide information on species, sex and individual identity, as well as current reproductive state, social status and health (Blaustein 1981; Brown and Macdonald 1985; Thiessen and Rice 1976). Such signals can function to attract males, advertise territory ownership or social rank, and to facilitate mate, group and offspring recognition (Heymann 2006; Nowak et al. 2000; Stockley et al. 2013). However, few female chemical signals have been identified as yet (Table 1), perhaps because of the focus on male signalling (see Liberles 2014 for a review of mammalian chemical signalling). In some species, females may produce more specialised signals known as pheromones. These are chemical signals used to communicate between members of the same species that trigger a specific behavioural response or physiological process (Karlson and Luscher 1959; Wyatt 2010). Female pheromones are recognised to induce sexual behaviours in males (Briand et al. 2004a) or to facilitating suckling behaviour in offspring (Schaal et al. 2003). Female mammals also use scents in a competitive context (Stockley et al. 2013), displaying similar strategies in the deployment of scent to those previously described in males (Gosling and Roberts 2001; Hurst and Beynon 2004). A common form of female competitive signalling is through scent marking (specialized motor patterns used to deposit chemical secretions on environmental objects or conspecifics (Ralls 1971)), which females use to advertise territory ownership and dominance rank (Stockley et al. 2013). Female scent marks can also advertise the current or approaching receptivity of the owner, or a female’s quality to potential mates (Blaustein 1981; Johnson 1973; Stockley et al. 2013). Once deposited, scent marks can be used by other animals to gain information about specific individuals, social groups or the areas they occupy, influencing future interactions and decisions (Brown and Macdonald 1985; Halpin 1986).
Most studies investigating chemical communication in mammals have focused on male signalling (Apps 2013; Gosling and Roberts 2001; Hurst 2009; Johnson 1973), probably due to male-biased sexual dimorphism in both scent marking behaviour and glandular morphology found in many species (Blaustein 1981). However, female chemical signals are also important in mammalian reproduction, and in some species female investment equals or even exceeds that of males (Ferkin 1999; Heymann 1998; Sliwa and Richardson 1998). Male-biased dimorphism is often reduced in monogamous species while female-biased dimorphism occurs in species with high levels of female competition (Heymann 2006; Kleiman 1977). The investment in chemical signalling and deployment of scents by females is strongly influenced by the social structure and breeding system of the species, as well as the current physical and social environment of the signalling individuals. Here, we review current literature on chemical signals in female reproduction, with an emphasis on their role in sexual attraction, intrasexual competition and cooperation, and maternal behaviours.
Sexual Attraction
Olfaction plays several key roles in mediating sexual and mating behaviours in mammals. As well as helping animals to locate and ensure appropriate recognition of opposite sex conspecifics, scents are also used as indicators of mate quality and for individual mate assessment (Johansson and Jones 2007). Further, chemical signals can be used to coordinate mammalian reproduction by altering the behaviour and physiology of both sexes (reviewed in Petrulis 2013). While the majority of studies have focused on males signalling to females (see Apps 2013; Burger 2005; Gosling and Roberts 2001; Roberts et al. 2014 for reviews), there is growing evidence that chemical cues are also produced by females to attract and stimulate males.
Reproductive Advertisement
A widely studied function of female chemical signalling is to advertise sexual receptivity and fertility. Female scent marking often occurs at an increased frequency or exclusively during periods of sexual receptivity (rat: Birke 1978; meadow vole: Ferkin et al. 2004; coyote: Gese and Ruff 1997; domestic rabbit: Gonzalez-Mariscal et al. 1990; Hudson and Vodermayer 1992; general review: Johnson 1973; ringtailed lemur: Kappeler 1998; Canidae: Kleiman 1966; giant panda: Nie et al. 2012; klipspringer: Roberts and Dunbar 2000). Increased marking during these periods probably functions to attract mates, and females typically increase scent marking rates in the presence of male odours (meadow vole: Ferkin et al. 2004; tree shew: Holst and Eichmann 1998; domestic rabbit: Hudson and Vodermayer 1992; golden hamster: Johnston 1977). Advertisement of receptivity may be particularly important in solitary animals that need to attract mates from a distance and encounter conspecifics less frequently than more gregarious species (Waser and Jones 1983). In several solitary species, female marking rates peak just prior to oestrus, which may advertise their approaching receptivity and ensure a male is present during oestrus (golden hamster: Johnston 1977; tiger: Smith et al. 1989). Sexual advertisement can also be important in social or pair living species, where females deposit marks at territory edges to attract neighbouring males (aardwolf: Sliwa and Richardson 1998; yellow mongoose: Wenhold and Rasa 1994). Females may signal receptivity to increase competition between males, thereby increasing the likelihood of mating with males of high quality (Fischer and Brown 1993; Rasmussen et al. 1997). Males may gain a range of different information about a female’s reproductive state from various scent sources, e.g. saliva, urine, body glands and genital secretions, which combined may provide precise information about a female’s reproductive state (Lai et al. 1996).
The ability to distinguish between odours produced at different points in the female reproductive cycle is shared by many species, including domestic dog (Beach and Gilmore 1949), golden hamster (Huck et al. 1989), domestic sheep (Blissitt et al. 1990), cotton top tamarin (Ziegler et al. 1993), meadow vole (Ferkin and Johnston 1995), giant panda (Swaisgood et al. 2002) and ring-tailed lemur (Drea and Scordato 2008), with males often more attracted to female scents during periods of receptivity (e.g. domestic dog (Beach and Gilmore 1949), rat (Lydell and Doty 1972), golden hamster (Huck et al. 1989), giant panda (Swaisgood et al. 2002), meadow vole (Ferkin et al. 2004), cow (Sankar and Archunan 2004)). In strepsirrhine primates the volatile components of genital secretions vary between the breeding and non-breeding seasons (delBarco-Trillo et al. 2012; Drea and Scordato 2008; Greene and Drea 2014; Hayes et al. 2004; Morelli et al. 2013). Similar distinctions have been found in other species for urine (Barman et al. 2013; Vogt et al. 2016), faeces (Kimura 2001) and genital secretions (Harris et al. 2014). However, much of this work has been largely qualitative, with few studies identifying the compounds involved in discrimination or quantifying the level of change. Studies in several species have identified scent components that may be used to advertise female receptivity (mouse: Achiraman and Archunan 2006; Stopka et al. 2007; cow: Archunan and Kumar 2012; blackbuck: Archunan and Rajagopal 2013; elephant: Rasmussen et al. 1997). For example, in female Asian elephants (Elephas maximus) the urinary compound (Z)-7-dodencen-1-yl acetate elicits male flehmen responses, erections and premating behaviour (Rasmussen et al. 1996; Rasmussen et al. 1997). The concentration of (Z)-7-dodencen-1-yl acetate peaks just prior to oestrus, suggesting it functions to advertise the approach of female receptivity (Rasmussen 2001). Female receptivity may also be signalled by a combination of compounds. Studies in rats and bovine species found mixtures of compounds that change with female receptivity, with compound mixtures producing the strongest behavioural responses from males (Nielsen et al. 2011; Rajanarayanan and Archunan 2011; Sankar and Archunan 2008). A limitation, though, is that many of these studies focus on domesticated or laboratory animals and similar experiments on wild species are needed. While many of these compounds stimulate attraction and sexual behaviours in males (Achiraman and Archunan 2006; Achiraman et al. 2010; Archunan and Kumar 2012; Nielsen et al. 2011; Rajanarayanan and Archunan 2011; Rasmussen et al. 1997; Sankar and Archunan 2008), further tests are required to confirm their role in signalling female receptivity, including tests of response when female odours are specifically manipulated.
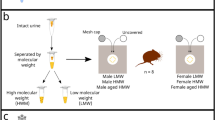
A particularly well studied example of female sexual advertisement concerns aphrodisin, a protein sex pheromone produced in the vaginal secretions of female golden hamsters (Mesocricetus auratus) that stimulates copulatory behaviour in males (Briand et al. 2004a; Singer and Macrides 1990). The expression of aphrodisin varies across the female reproductive cycle, reaching a maximum at oestrus (Briand et al. 2004a). Aphrodisin is an odorant binding protein that binds small hydrophobic molecules in vaginal secretions within its central cavity (Briand et al. 2000). The purified high molecular weight fraction of vaginal fluid containing aphrodisin stimulates mounting behaviour in male hamsters when detected through the vomeronasal system, even when painted onto anaesthetised males (Singer et al. 1986). This pheromonal effect may be due to low molecular weight ligands carried by aphrodisin, or to the complex of both protein and ligand, as recombinant aphrodisin alone was not effective in stimulating a copulatory response (Briand et al. 2004b).
Changes in female scent and scent marking behaviours are related to fluctuations in ovarian hormones across the female reproductive cycle (Takahashi 1990). Ovariectomy reduces or eliminates scent marking (golden hamster: Albers and Rowland 1989; domestic rabbit: Hudson and Vodermayer 1992; mouse: Kimura and Hagiwara 1985; Mongolian gerbil: Wallace et al. 1973), causes scent glands to regress (grasshopper mouse: Pinter 1985; Mongolian gerbil: Wallace et al. 1973) and reduces male attraction to female odours (meadow vole: Ferkin et al. 1991; domestic dog: Lisberg and Snowdon 2009). However, scent marking and glandular morphology can be restored through hormonal injections of estradiol (golden hamster: Albers and Rowland 1989; gray short-tailed opossum: Fadem 1990; mouse: Kimura and Hagiwara 1985) or estradiol and progesterone (rat: Birke 1984; Mongolian gerbil: Owen and Thiessen 1974). Hormone levels can also alter the chemical composition of female odours or the relative abundance of volatile chemicals in female scents (pine vole: Boyer et al. 1989; ringtailed lemur: Crawford et al. 2011; grey wolf: Raymer et al. 1986).
The best studied example of the hormonal control of female scent marking is vaginal marking in golden hamsters (reviewed in Been and Petrulis 2008). Vaginal marking peaks just prior to receptivity, but disappears during oestrus (Johnston 1977). This rise in vaginal marking is mediated by high levels of estradiol prior to receptivity (Lisk and Nachtigall 1988). The decline in marking during oestrus is probably due to falling levels of estradiol coupled with rising progesterone (Lisk and Nachtigall 1988). Implantations of estradiol and progesterone within the brain provided further evidence for the hormonal dependency of vaginal marking behaviour (Takahashi et al. 1985; Takahashi and Lisk 1985). However, vaginal marking rates also increase in the presence of male odours (Johnston 1977) and normal marking behaviour depends on an intact olfactory system (Johnston 1992; Petrulis et al. 1999), showing that vaginal marking is regulated by external chemosensory cues as well as internal hormonal cycles.
Have females evolved specific signals to inform males of their receptivity, or have males learned to detect changes in female cues that occur as a by-product of hormonally driven physiological changes? Although experimentally differentiating between signals and cues can be difficult, their evolutionary implications can be very different (Otte 1974). Signalling implies intentional information transfer which benefits the signaller, for example if females signal receptivity to attract a mate in solitary species or to gain benefits from mating with multiple males (Reynolds 1996; Steiger et al. 2010). However, when males detect changes in female cues that females are not actually signalling, this can have neutral or even detrimental effects on females (Otte 1974).
While advertisement of female receptivity has been widely investigated, the extent to which females signal outside of mating has received much less attention. Scent marking rates in female golden hamsters, a solitary species, are lowest during pregnancy and early lactation (Johnston 1979) and males are less attracted to odours from pregnant females (Johnston 1980). Reduced sexual advertisement during these periods could prevent males approaching pregnant or lactating females, reducing the risk of infanticide to their pups. Alternatively, the high energetic costs of gestation and lactation may limit female scent marking during this period (Clutton-Brock et al. 1989; Gubernick and Klopfer 1981; Wade and Schneider 1992). Male meadow voles (Microtus pennsylvanicus) are significantly less attracted to female odours immediately prior to parturition, although pre-parturition odours are still more attractive than male odours (Ferkin and Johnston 1995). Pregnant and lactating females are highly aggressive in many species (Svare 1981), so this decrease in male attraction may be due to females emitting a “stay away” signal. By advertising an increase in aggressiveness, females may prevent conspecifics from approaching them and their pups. Unfamiliar males and females pose a risk to newborn young in many species (Ebensperger 1998), so female “stay away” odours should result in avoidance by both sexes. However, the response of female voles to pre-parturition odours was not tested. By contrast, the odours of female voles in post-partum oestrus, a period of receptivity that occurs soon after parturition, are highly attractive to males (Ferkin and Johnston 1995). Females may advertise receptivity rather than aggression during postpartum oestrus, as the necessity to attract a mate may outweigh the desire to prevent conspecifics approaching vulnerable pups during this period. Alternatively, the attractiveness of female receptivity cues may cause males to ignore female signals of aggression.
Instead of advertising receptivity, some female mammals may try to conceal their reproductive state. In several socially monogamous species, where a single breeding male and single breeding female share a common range or territory and associate with each other for more than one breeding season (Lukas and Clutton-Brock 2013), female scent marking rates do not change across the reproductive cycle (pygmy marmoset: Converse et al. 1995; prairie vole: Wolff et al. 2002). Attracting males during periods of receptivity may be less important in monogamous species and concealed ovulation has been linked to the evolution of monogamy in primates (Alexander and Noonan 1979 but see Dixson 2012). However, lack of advertisement is not the same as concealment and socially monogamous males can still discriminate reproductive state from female odours (pygmy marmoset: Converse et al. 1995; cotton-top tamarin: Ziegler et al. 1993). Socially monogamous females can engage in extra-pair copulations so, may still benefit from advertising receptivity to neighbouring males (Birkhead and Moller 1992; Clutton-Brock and Isvaran 2006). Further, given the extent to which hormone levels can influence chemical secretions (see references above), and the sensitivity of vomeronasal receptors to sulphated derivatives of all major classes of steroid hormones excreted in urine (Nodari et al. 2008), it seems unlikely that females could ever completely conceal their reproductive state.
Effects of Female Chemosignals on Male Physiology
While the priming effects of male chemosignals on female reproductive physiology have been well studied (Koyama 2004; Koyama 2016), potential effects of female chemosignals on males have received less attention (Petrulis 2013). Male laboratory mice housed with an unrelated adult female reach sexual maturity faster than when housed alone or with an unrelated adult male (Vandenbergh 1971), while exposure to female scents can increase testis and seminal vesicle weights in juvenile male rodents (Babb and Terman 1982; Purvis and Haynes 1972; Terman 1984; Wayne and Rissman 1990). Puberty acceleration in males following exposure to adult females may help coordinate reproduction between the sexes. Further, males may increase their reproductive success by reaching puberty earlier when sexually mature females are available, particularly in short-lived rodents where male mortality is high (Berry and Bronson 1992; Promislow and Harvey 1990; Triggs 1991).
Exposure to an unfamiliar female or her odours causes an increase in plasma testosterone in male mice (Macrides et al. 1975), rats (Bonilla-Jaime et al. 2006), hamsters (Macrides et al. 1974), marmosets (Ziegler et al. 2005), macaques (Cerda-Molina et al. 2006) and humans (Miller and Maner 2010; but see Roney and Simmons 2012). This increase in testosterone occurs 30-60 minutes after initial exposure (Cerda-Molina et al. 2006; Richardson et al. 2004; Ziegler et al. 2005) and is often preceded by an increase in circulating luteinizing hormone (Cerda-Molina et al. 2006; Richardson et al. 2004). Although the adaptive functions of these hormonal changes are currently unknown, high testosterone levels are linked to many male-specific traits, as well as male mating success, aggression and dominance rank (Beehner et al. 2006; Mills et al. 2009; Muller 2017; Setchell et al. 2008; Wickings and Dixson 1992). Many male chemosignals are androgen dependent (mice: Achiraman and Archunan 2005; Harvey et al. 1989; Novotny et al. 1985; goat: Iwata et al. 2000; domestic pig: Loebel et al. 2001; rat: Ponmanickam et al. 2010), so increased levels of male hormones may increase production of male chemical signals that are attractive to females (Roberts et al. 2010; Zhang et al. 2008). However, hormonal increases in male laboratory mice were not dependent upon female reproductive state (Maruniak and Bronson 1976). The components of female scents that elicit surges in male hormones are yet to be identified, but in mice low molecular weight molecules bound by major urinary proteins (MUPs) may be responsible (Singer et al. 1988).
Testosterone is essential for spermatogenesis (Smith and Walker 2014; Walker 2011), so the elevation in testosterone in response to female chemical signals may function to increase sperm production. In agreement with this, increased sperm production in response to female chemical signals has been reported in laboratory rodents (Koyama and Kamimura 2000; Taylor et al. 1987). Dominant male laboratory mice, but not subordinates, show increased sperm density when housed with female bedding (Koyama and Kamimura 2000) and odour from group housed females produces a greater increase than odours from isolated females (Koyama 2004). However, a study in wild house mice (Mus musculus) found that although males showed plasticity in sperm production, this was caused by competitive cues from other males, rather than by female cues relating to mating opportunities (Ramm et al. 2015).
Male hormonal responses to females are context dependent, as male rodents quickly habituate to presentation of the same female, and the greatest elevation of luteinising hormone and testosterone follows exposure to a novel female (Coquelin and Bronson 1979; Shulman and Spritzer 2014). Further, hormone surges can be linked to neutral olfactory cues through learned association with receptive females (Graham and Desjardins 1980). Social conditions impact endocrine response in male common marmosets (Callithrix jacchus): males housed alone or with a female show elevated testosterone following exposure to a novel female odour but males housed in family groups do not (Ziegler et al. 2005). Male marmosets contribute to parental care but elevated levels of testosterone are linked to male aggression (Dixson 1980; Honess and Marin 2006; Rose et al. 1971). Those in family groups may inhibit the normal elevation of testosterone to minimise aggressive behaviours towards vulnerable infants. However, during control tests, males housed in families or pairs tended to have slightly higher testosterone levels than singly housed males, although the difference was not statistically significant (Ziegler et al. 2005). Alternatively, elevation of testosterone in response to female scents may function to increase male mating rates, for example through increasing spermatogenesis or production of attractive male chemosignals, which may be less important in socially monogamous male marmosets housed in a stable family group.
The renewal of sexual behaviour following mating when males are exposed to a novel female, called the Coolidge effect, occurs in many male mammals, including rats (Bermant et al. 1968; Brown 1974; Wilson et al. 1963), voles (Dewsbury 1973; Gray and Dewsbury 1975), cats (Whalen 1963), sheep (Pepelko and Clegg 1965) and hamsters (Bradford et al. 1977). Males often show more chemosensory investigation of novel females, suggesting that chemical cues may allow males to recognise familiar females (Johnston and Rasmussen 1984). Consistent with this, disruption of the main olfactory system abolishes the preference of sexually satiated male golden hamsters for a novel female (Johnston and Rasmussen 1984). Recent studies in laboratory rats have shown that sexually satiated males fail to ejaculate semen, as no spermatozoa or seminal plugs are found in the female genital tract following copulation. Further, sexually satiated male rats impregnate significantly fewer females than do rested males (Lucio et al. 2014). An alternative function of the Coolidge effect may be to reduce the probability of fertilization by rival males. In laboratory rats, sexually satiated males dislodge seminal plugs deposited by previous males, reducing sperm from rival males in the female’s genital tract (Lucio et al. 2014). Notably, no female rats that had recently mated gave birth after copulating with another male under the Coolidge effect (Lucio et al. 2014). The absence of any renewal of sexual behaviour in socially monogamous rodents on exposure to a novel female (Dewsbury 1971; Pierce et al. 1992) provides further evidence that the Coolidge effect may function to mediate post-copulatory competition between males in promiscuous species. Mating with multiple males can increase female reproductive fitness (Jennions and Petrie 2000), so females may not necessarily benefit from mating under the Coolidge effect.
Female Signals of Quality
While females often prefer males that deposit the greatest number of scent marks, and high rates of scent marking among males have been linked to their quality and competitive ability (Gosling and Roberts 2001; Rich and Hurst 1999), the extent to which female chemosignals are used by males to select a mate of high quality has received little attention. Further, the influence of female scent marking rate on male mate choice remains largely unknown. Females of several species, including bush dogs (Porton 1983), cotton top tamarins (French and Cleveland 1984) and moustached tamarins (Heymann 1998), display more frequent and diverse marking behaviour than males, and males typically spend more time investigating female scents (Heymann 1998). Female-biased marking in these species is probably due to high levels of male parental care, leading to increased female competition and male mate choice. Among callitrichid primates, scent marking rates are female-biased in species with male-biased parental care, but male-biased when parental care is equal or greater in females (Heymann 2006). The increased cost of marking to females may be offset by the assistance in offspring care from males.
Female preference between male scents can depend upon male status (Kruczek 1997; Mossman and Drickamer 1996; Zhang et al. 2001), health (Kavaliers and Colwell 1995; Willis and Poulin 2000) and genetic quality of the scent owner (Ilmonen et al. 2009; Thom et al. 2008). Few studies have investigated whether similar factors influence male preferences between female scents. Male ring-tailed lemurs (Lemur catta) are more attracted to scents from dominant females, but only if scents are from familiar females (Scordato and Drea 2007). The few studies that have investigated female quality signalling and male preference have focused on species in which females are dominant. Studies in species covering a range of other social systems are also needed.
Whether female chemical signals produce honest signals of quality has rarely been investigated, probably because female signalling is expected to be limited by the high costs of offspring production (Chenoweth et al. 2006; Fitzpatrick et al. 1995; Nordeide et al. 2013). Ferkin et al. (1997) found that male meadow voles were more attracted to odours from females fed on a high, compared to a low, protein diet. Female meadow voles occupy exclusive territories in areas where food patches often differ in quality (Bowers et al. 1996; Madison and McShea 1987). A protein-rich diet may provide a good indicator of a female’s ability to hold a high quality territory successfully, so may be a true indicator of female quality or ability to invest in offspring. In ring-tailed lemurs, individual heterozygosity correlates negatively with diversity of fatty acids and positively with the diversity of heavy fatty acid esters (Boulet et al. 2010). The authors argue that this may be an example of honest olfactory signalling as genetic heterozygosity correlates with health and survivorship in this captive population (Charpentier et al. 2008). However, subsequent behavioural tests revealed that males tended to spend more time near scent from less heterozygous females not less time, although this difference was not statistically significant (Charpentier et al. 2010). Chemical diversity correlates positively with genetic heterozygosity in female Antarctic fur seals (Arctocephalus gazella) (Stoffel et al. 2015). As heterozygosity increases early survivorship and breeding success in female fur seals (Stoffel et al. 2015), greater chemical diversity could be an indicator of female quality. However, behavioural tests are needed to test whether males prefer more heterozygous females and their scents.
Intrasexual Competition and Cooperation
Scent signals can play an integral role in mediating both competitive and cooperative interactions between females (Stockley et al. 2013). The high cost of lactation and gestation means that females frequently compete for access to resources (Stockley and Bro-Jorgensen 2011), using scent marks to signal ownership of particular resources or areas containing resources (Kruuk 1992; Miller et al. 2003; Ralls 1971). In group living species odours can be used to signal social rank (Heymann 2006; Ralls 1971), with high ranking females often benefiting from priority access to resources and high quality mates (Côté and Festa-Bianchet 2001; Pusey et al. 1997; van Noordwijk and van Schaik 1999). Females often counter-mark the scents of other females, by depositing scent marks on top of or adjacent to the original mark (Ewer 1968), to signal competitive ability (Gosling 1982; Rich and Hurst 1999). Odours also allow recognition of group members or cooperative partners, which may be particularly important in species that rear their young together, providing benefits through group defence as well as cooperative hunting, nesting and / or nursing (Gittleman 1989; Jennions and Macdonald 1994; Packer et al. 1990). Chemical signals have been linked to reproductive suppression in cooperatively breeding species, where reproduction is monopolised by the dominant pair within a group and subordinate females assist with offspring care (Clutton-Brock et al. 2001; Creel et al. 1997; Faulkes and Bennett 2001; French 1997). As discussed below, levels of female cooperation or competition depend on the degree of reproductive synchronisation, the social structure and mating system of the species, the relative ages of the competing females and current environmental conditions.
Territorial Marking
Female reproductive success is often constrained by the availability of resources such as food, water or shelter (Clutton-Brock 2009; Stockley and Bro-Jorgensen 2011). To maintain access to limiting resources, females often establish territories that they defend either alone or as part of a group (Ostfeld 1985; Stockley and Bro-Jorgensen 2011). Scent marking is used to advertise territory ownership in many species (e.g. common vole: Dobly 2005; general review: Gosling 1982; spotted hyena: Henschel and Skinner 1991; banded mongoose: Müller and Manser 2008; European rabbit: Mykytowycz 1965; aardwolf: Richardson 1991; klipspringer: Roberts and Dunbar 2000; Eurasian beaver: Rosell et al. 1998; Ethiopian wolf: Sillero-Zubiri and Macdonald 1998; tiger: Smith et al. 1989). The presence of foreign scent marks induces investigation from the territory owner (Palagi et al. 2005; Sliwa and Richardson 1998) followed by an increased rate of scent marking, particularly when intruders are the same sex (Dobly 2005; Hurst 1990; Johnston 1977; Sliwa and Richardson 1998). Territorial marking may be an honest form of signalling as only successful territory holders will have the most abundant and / or overall freshest scent marks, providing a continuous record of ownership (Gosling and Roberts 2001; Rich and Hurst 1998).
Territorial marking can also influence the spacing behaviour of mammals. In honey badgers (Mellivora capensis) token urination is performed almost exclusively by females and patterns of marking do not vary across seasons, suggesting it may function to maintain spatiotemporal separation between females (Begg et al. 2003). Scents may also effect spacing behaviour in golden hamsters as female hamsters avoid areas marked by other females (Fischer and McQuiston 1991). As resident animals are more likely to defend their home territory, the avoidance of areas scent marked by conspecifics may reduce costly aggressive encounters between females (Roberts 2012). Further, female golden hamsters are more likely to attack an intruder when their own odour is present in the local environment and the presence of a previously subordinate female’s scent can cause a reversal in dominance (Fischer and McQuiston 1991). While territorial marking does not directly prevent other individuals from entering a territory, it does allows intruders to make an informed decision about the relative costs or benefits before entering (Gosling 1982). Females may use the chemical composition of scent marks to assess the competitive ability of territory holders. Female house mice increase investment in MUPs when faced with competition from neighbouring females (Garratt et al. 2011). The frequency of aggressive behaviours towards unfamiliar females is also strongly related to the urinary protein output of the aggressor, with more aggressive females exhibiting higher protein investment (Stockley et al. 2013). Under high densities, female reproductive success is strongly influenced by the ability to successfully defend a territory (Hurst 1987), so investment in urinary protein in scent marks may deter intruding females by serving as an indicator of investment in territory defence by the resident female.
Territorial animals often differ in their behaviour towards familiar neighbours compared to unfamiliar animals. Many mammals can discriminate between odour from neighbouring individuals and strangers (giant kangaroo rat: Murdock and Randall 2001; European badger: Palphramand and White 2007; Eurasian beaver: Rosell and Bjørkøyli 2002; aardwolf: Sliwa and Richardson 1998) and typically spend longer investigating scents from unknown conspecifics (African lion: Gilfillan et al. 2017; European rabbit: Monclús et al. 2014; Colombian ground squirrel: Raynaud and Dobson 2011; Eurasian beaver: Rosell and Bjørkøyli 2002). When presented with translocated scent marks from neighbouring conspecifics, aardwolves (Proteles cristata) immediately visit and scent mark their shared border, suggesting that they are capable of recognising neighbouring individuals (Sliwa and Richardson 1998). Territory holders may reduce the energetic costs of territorial defence by decreasing aggression towards neighbouring conspecifics, the so called the “dear enemy phenomenon” (Fischer 1954; Temeles 1994; Ydenberg et al. 1988). Female meadow voles display less antagonistic behaviour towards females with a familiar scent they have previously encountered (Ferkin 1988). Additionally, female bank voles (Myodes glareolus) display higher rates of infanticide towards the offspring of unfamiliar females under semi-natural conditions (Ylonen et al. 1997). However, European rabbits (Oryctolagus cuniculus) exposed to repeated simulation of intrusions by neighbours increase their rates of counter marking, suggesting that animals adapt their behaviour to neighbours depending upon the perceived level of threat (Monclús et al. 2014).
The “nasty neighbour” hypothesis predicts that, rather than reducing agonistic behaviour, residents will display increased levels of aggression towards neighbouring conspecifics (Müller and Manser 2007). Banded mongoose groups (Mungos mungo) emit more worry calls and perform more inspection bouts in response to translocated scent marks from neighbouring groups compared to the scent marks of strangers (Müller and Manser 2007). Social animals often disperse alone or in small groups, so strangers may pose less threat to a group’s resources than neighbouring groups of similar size (Cant et al. 2001; Cant et al. 2002). Intolerance of neighbours may also increase with population density. Female mound building mice (Mus spicilegus) tend to be more aggressive towards their immediate neighbours, particularly when pregnant (Simeonovska-Nikolova 2012). Although the sex ratio is equal at the beginning of the breeding season, it becomes female biased during the late summer (Simeonovska-Nikolova 2012). The declining nutritional value of food resources coupled with the rising density of females during summer months increases competition between neighbours (Simeonovska-Nikolova 2012).
Dominance Signals
Scent signals are used to advertise and maintain social or reproductive dominance across a wide range of species (Barrette 1977; Ralls 1971). Dominant females often display higher rates of scent marking (Callitrichidae: Epple 1972; coyote: Gese and Ruff 1997; golden hamster: Johnston 1977; meerkat: Jordan 2007; African wild dog: Jordan et al. 2013) and counter-marking (banded mongoose: Müller and Manser 2008; ringtailed lemur: Palagi et al. 2004), and may have larger and more complex scent glands (golden hamster: Drickamer and Vandenbergh 1973). In ring-tailed lemurs, high-ranking females counter-mark the genital marks of other females more frequently than low-ranking females do (Palagi et al. 2004). Similarly, dominant female banded mongooses counter-mark the scents of other females at higher rates than subordinates (Müller and Manser 2008; but see Jordan et al. 2011). Rates of counter-marking by female mongooses increase during oestrus, suggesting that counter-marking may be involved in female competition for males during the breeding season. Further, in Eulemur, the group of Lemuridae known as brown lemurs, the chemical complexity of genital secretions is male-biased in subspecies that lack rank relations between the sexes (co-dominated social structure) but female-biased in female dominated species (delBarco-Trillo et al. 2012). However, as the chemical complexity of female secretions does not differ between co-dominated and female dominated species (delBarco-Trillo et al. 2012), this female-biased chemical complexity is probably due to a reduction in male signal complexity in female dominated species, rather than a greater complexity in females.
Different patterns of scent marking rates in relation to female social rank have also been reported. In common marmosets, subordinate females scent mark more frequently than dominant females during intergroup encounters (Lazaro-Perea et al. 1999). Similarly, subordinate female yellow mongooses (Cynictis penicillata) deposit more scent marks than both dominant and juvenile females and concentrate most scent marking at territory borders (Wenhold and Rasa 1994). Subordinate females often visit neighbouring territories during oestrus and are mated by neighbouring males (Wenhold and Rasa 1994), suggesting that their marking functions to advertise their presence to potential mates. Interestingly, a subsequent study carried out in a different population of yellow mongoose found no increased scent marking rates by subordinate females at territory borders (Le Roux et al. 2008). The difference between these two studies may be due to a difference in population density. Le Roux et al. (2008) studied a low-density population where females dispersed into new territories upon reaching sexual maturity. At the higher densities studied by Wenhold and Rasa (1994), females may have been unable to disperse so instead searched for mating opportunities within neighbouring groups.
Patterns of female scent marking also depend on whether females compete for reproductive opportunities within their group or between neighbouring groups. Alpha female golden lion tamarins (Leontopithecus rosalia) only display higher rates of scent marking than subordinates during intergroup encounters (Miller et al. 2003). Subordinate females are often daughters of the alpha pair, so pose little threat to the dominant female’s reproduction (Miller et al. 2003). Dispersing females can only enter a group as the alpha female (Baker and Dietz 1996), therefore alpha females may increase scent marking rates in the presence of female intruders to advertise their presence and deter other females from attempting to immigrate (Miller et al. 2003).
Reproductive Suppression and Synchronisation
Synchronisation of oestrus has been reported in a number of mammalian taxa, particularly among primates and rodents (French and Stribley 1987; Handelmann et al. 1980; McClintock 1971; McClintock 1978; Wallis 1985; Weller and Weller 1993; Weller and Weller 1997). Ovarian synchronisation may reduce monopolization of females by dominant males, allowing females to pursue copulations with other mates (Emlen and Oring 1977). Early studies in laboratory rats suggested this synchronisation was mediated by female chemical signals (McClintock 1978; McClintock and Adler 1978; McClintock 1984). Similarly, the timing of ovulation was suggested to be under female pheromonal control in humans (Preti et al. 1986; Russell et al. 1980; Stern and McClintock 1998). However, many of these original studies have been criticized for methodological and statistical errors (Arden and Dye 1998; Schank 2000; Schank 2001a; Strassmann 1999; Wilson 1987; Wilson 1992) and subsequent studies found no evidence for synchronisation (Erb et al. 1993; Fürtbauer et al. 2011; Monfort et al. 1996; Schank 2001b; Setchell et al. 2011; Strassmann 1997; Tobler et al. 2010; Trevathan et al. 1993; Yang and Schank 2006). Given the variability within and between female ovarian cycles, it is argued that true synchronisation of oestrus is highly unlikely because matching will not be achieved across multiple cycles (Schank 2000). Additionally, many of these studies investigated oestrus synchrony in captive animals, where breeding is artificially regulated, or in western human populations, where women often use contraceptives to control reproduction. Such studies may not be representative of natural populations where females spend significantly more time pregnant or lactating (Strassmann 1997).
As synchronisation of oestrus may lead to high levels of competition between females and to male mate choice, females may actually benefit by avoiding ovarian synchronisation (Emlen and Oring 1977; Schank 2004). Asynchronisation of oestrus has been reported in several species (golden hamster: Gattermann et al. 2002; chimpanzee: Matsumoto-Oda et al. 2007; ring-tailed lemur: Pereira 1991) and may function to reduce female competition for mates (Schank 2004). Alternatively, females may delay reproduction during periods of high competition. Oestrus cycles in group housed female laboratory mice are significantly longer than in mice housed alone or in small groups of 2-3, known as the Lee Boot effect (Champlin 1971; Whitten 1959). Replication of the Lee-Boot effect through exposure to urine from group housed anoestrus females indicates that this is mediated by olfactory cues (Drickamer 1974; McIntosh and Drickamer 1977). Subsequent tests revealed 2,5-dimethylpyrazine, produced by group-housed non-breeding females, to be the key component in lengthening the oestrus cycle (Ma et al. 1998; Novotny et al. 1986). The magnitude of the Lee-Boot effect correlates positively with both the density of non-breeding females and the length of time they are in groups (Coppola and Vandenbergh 1985). This suggests that it may function to prevent female reproduction at high population densities when pup survival is poor among crowded females (Christian and Lemunyan 1958; Southwick 1955a; Southwick 1955b). Delaying reproduction during periods of high competition may increase long term reproductive success in females of lower competitive ability (Wasser and Barash 1983).
Exposure to 2,5-dimethylpyrazine also causes puberty delay in both male and female mice (Jemiolo and Novotny 1993; Jemiolo and Novotny 1994). Urine from wild mice living at high density delays puberty in laboratory mice, providing a mechanism that can delay reproduction in natural populations under conditions of increased resource competition (Massey and Vandenbergh 1980). In support of the suppressive effects of 2,5-dimethylpyrazine, urine and soiled bedding from group housed females reduced population growth in mice living in two large outdoor enclosures due to a combination of fewer females attaining puberty and at a later age (Drickamer and Mikesic 1990). As reproductive success is often linked to body size and condition, young females particularly may benefit from delaying reproduction (McNamara and Houston 1996). Delayed sexual maturation has been reported in other female rodents (vole species: Batzli et al. 1977; Mongolian gerbil: Clark and Galef 2002; prairie vole: Getz et al. 1983; california mouse: Gubernick and Nordby 1992; deermouse: Haigh 1983; pine vole: Schadler 1990). However, the mechanisms of suppression in these species are yet to be determined, with some evidence for both behavioural (Brant et al. 1998; Gubernick and Nordby 1992) and chemical mechanisms (Batzli et al. 1977; Getz et al. 1983; Schadler 1990). Further, a study by Wolff et al. (2001) found no evidence that female meadow voles (M. pennsylvanicus) or prairie voles (Microtus ochrogaster) suppress reproduction in the presence of their mother. In some species, puberty delay may not be inhibited by female signals, but females may require the presence of an unfamiliar male to stimulate reproduction (Mongolian gerbil: Clark and Galef 2002; prarie vole: Hofmann and Getz 1988; McGuire and Getz 1991). Exposing young female Mongolian gerbils (Meriones unguiculatus) to an unfamiliar male accelerates development even in the presence of their reproductively active mother (Clark and Galef 2002). Stimulation by an unfamiliar male may reduce the risk of inbreeding among young females.
In many cooperatively breeding species, dominant females suppress subordinate reproduction (common marmoset: Abbott et al. 1988; Damaraland mole-rat: Bennett 1994; African wild dog: Creel et al. 1997; meerkat: O’Riain et al. 2000; general-review: Solomon and French 1997; Ethiopian wolf: van Kesteren et al. 2013). This benefits dominant females by reducing competition for resources, as well gaining assistance from suppressed subordinates in offspring care (Hodge 2009). Olfactory cues from dominant individuals have been implicated in the reproductive suppression of subordinate females in callitrichid primates (Barrett et al. 1990; Epple and Katz 1984; Heistermann et al. 1989; Savage et al. 1988). However, only odours from familiar, dominant females inhibit ovulation, suggesting that odours inhibit reproduction by signalling the presence of the familiar dominant rather than through a pheromonal cue produced by all dominant females (Abbott et al. 1997). Further, female common marmosets that remained in visual, but not in olfactory, contact with their dominant female were still reproductively suppressed, again indicating that suppression is not directly caused by a chemical cue (Barrett et al. 1993). Instead subordinate females may learn to associate cues from a familiar dominant female with the behavioural subordination that the female imposes.
If odour signals from dominant females function as a threat, reproductive suppression in subordinates may be self-imposed (Johnstone and Cant 1999; Saltzman et al. 2009). This may allow subordinates to avoid costly aggressive encounters including eviction from the group and infanticide (Clutton-Brock et al. 1998; Kutsukake and Clutton-Brock 2006; Saltzman et al. 2009; Young et al. 2006). Additionally, subordinate females may increase survival and long-term reproductive success by remaining on their natal territory until they can either replace the dominant female or disperse (Clutton-Brock et al. 1998; Clutton-Brock et al. 1999; Rood 1990). As subordinate females are often the offspring of the dominant pair, they may also gain indirect fitness benefits from assisting with raising siblings (Emlen 1995; Griffin and West 2003). In some species, self-imposed reproductive suppression may function to reduce inbreeding (Snowdon 1996). In Damaraland mole rats (Cryptomys damarensis) females continue to exhibit reproductive suppression 30 days after being removed from the dominant female or her cues (Clarke et al. 2001). Exposure to an unrelated male results in rapid onset of reproductive activation, even in the presence of the dominant female (Clarke et al. 2001; Cooney and Bennett 2000). Breeding by subordinate females following the introduction of an unrelated male has been reported in other cooperatively breeding species (common marmosets: Digby 1995; Saltzman et al. 2004; meerkats: O’Riain et al. 2000). However, subordinates still have significantly lower reproductive rates than dominant females (Digby 1995; O’Riain et al. 2000), suggesting that reproductive suppression may be caused by an interplay between rank-related breeding and inbreeding avoidance.
Choice of Social Partners
Olfactory cues can promote group cohesion and cooperation in gregarious species. Group living individuals benefit from reduced predation and increased success at locating or maintaining access to resources (Silk 2007). Group members often scent mark at communal marking sites, which may facilitate information transfer within the group and encourage group cohesion (Gittleman and Thompson 1988; Johnson 1973; Porton 1983; Sillero-Zubiri and Macdonald 1998). In spotted hyenas (Crocuta crocuta) anal gland secretions from high ranking females are preferentially overmarked by subordinates (Burgener et al. 2009). During overmarking, individuals “anoint” themselves with scent from the previous donor, a behaviour that may be involved in advertising continued clan membership (Burgener et al. 2008). Female marking rates decline with age, with young adult females displaying the highest marking rates, indicating a greater need for younger females to advertise group membership (East et al. 2013). Advertisement of group membership may be particularly important in spotted hyenas, which live in fission-fusion groups, where females may be absent from the clan territory for several days on long distant foraging trips (Hofer and East 1993; Kruuk 1972).
Members of the same group may have a distinctive shared odour (big brown bat: Bloss et al. 2002; spotted heyenas: Burgener et al. 2008; European badger: Gorman et al. 1984; meerkat: Leclaire et al. 2017; Bechstein’s bat: Safi and Kerth 2003). In some species of bats, females can identify roost-mates by scent (Bloss et al. 2002; Bouchard 2001; De Fanis and Jones 1995) and will attack foreign females that enter the colony (Kerth et al. 2002). Additionally, the volatile components of odours from different colonies are chemically distinct in Bechstein bats (Myotis bechsteinii, (Safi and Kerth 2003) and big brown bats (Eptesicus fuscus, (Bloss et al. 2002). In both species, females raise their offspring in maternal colonies which, despite disintegrating over winter, are stable over time as females return to the same colony each year (Bloss et al. 2002; Kerth et al. 2002; Kerth et al. 2011). Further, within some colonies females display fission-fusion societies, where females split into subgroups to occupy different day roosts (Kerth and Barbara 1999; Kerth et al. 2011), suggesting that colony specific odours may facilitate long term group stability. Group specific odours have also been reported in hyenas (Burgener et al. 2008), meerkats (Leclaire et al. 2017) and badgers (Gorman et al. 1984) and may be caused by differences in bacterial communities between groups (Leclaire et al. 2014; Leclaire et al. 2017; Theis et al. 2012; Theis et al. 2013). The fermentation hypothesis predicts that bacteria within scent glands metabolize glandular secretions, producing compounds that are used by the host to communicate with conspecifics (Albone and Gronnerberg 1977; Archie and Theis 2011; Gorman 1976). Members of the same social group harbour more similar odour-producing bacteria in their scent glands than members of different social groups (Archie and Theis 2011). Using GC-MS and deep sequencing techniques, Leclaire et al. (2017) found that the chemical composition of anal gland secretion and the composition of bacterial communities within the anal gland varied with group membership in wild meerkats, Suricata suricatta. These group-specific bacterial communities could arise through cross-infection from allomarking or rapid overmarking within groups (Buesching et al. 2003; Burgener et al. 2008; Theis et al. 2008).
Choice of social partners may be particularly important in species that care for offspring communally. Female house mice often raise their young in a communal nest (Manning et al. 1995; Wilkinson and Baker 1988), in which dams combine all their offspring and share maternal duties including nursing (König 1989a). Although prior familiarity between partners is a major factor influencing the success of communal nests (König 1993; König 1994), nesting with closely related females may also be beneficial (König 1994). Odour similarity correlates with genetic similarity (referred to as odour-gene covariance) across a range of species, providing a mechanism for assessing genetic relatedness among conspecifics through scent (Boulet et al. 2009; Todrank and Heth 2003; Tzur et al. 2009). However, the influence of many non-genetic factors on scents reduces the reliability of assessing degree of relatedness simply from overall chemical similarity (Hurst and Beynon 2010). While female house mice prefer unfamiliar nestmates that are related over equivalently unrelated females, their strongest preference is for nestmates that share the same polymorphic pattern of MUPs in their urine, which they can detect through urine scent (Green et al. 2015). As only females that are very closely related are likely to express the same inherited pattern of MUPs, this will be a highly reliable marker of close relatedness between females. Reliable assessment of close relatedness may be particularly important in the context of communal nursing when potential partners for cooperation are not highly familiar littermate sisters, allowing females to gain indirect fitness benefits from lactation investment in offspring that are not their own.
Maternal Behaviours
In mammals, the cost of reproduction continues after birth as neonates are entirely dependent upon their mother (Gubernick and Klopfer 1981). Postnatal investment represents a significant cost to females and reduces their opportunity to produce additional offspring, so there should be strong selection to ensure that own offspring are the beneficiaries (Clutton-Brock 1991). Chemical cues can be used for both offspring recognition and maternal recognition (Nowak et al. 2000) and odours facilitate suckling behaviour in a number of mammals (Arteaga et al. 2013; Schaal and Al Aïn 2014). As discussed below, patterns of parent-offspring interactions depend on both the developmental status of the neonate and the litter size, as well as the breeding system of the mother (Clutton-Brock 1991; Nowak et al. 2000).
Mother-Offspring Recognition
Discriminative care of offspring is common in mammals due to the high cost of lactation and other maternal behaviours (Clutton-Brock 1991; Gubernick and Klopfer 1981). Chemical cues are important in offspring recognition in many mammalian species including domestic ungulates (reviewed in Nowak et al. 2000; Poindron et al. 2007), rodents (Beach and Jaynes 1956; Ostermeyer and Elwood 1983; Porter et al. 1973), bats (Fanis and Jones 1996; Gustin and McCracken 1987) and pinnipeds (Pitcher et al. 2010a). One of the best studied examples is the domestic sheep, Ovis aries. Ewes can identify their own young shortly after parturition and prevent alien young from suckling (Keller et al. 2003; Poindron et al. 1993). Maternal behaviours and offspring recognition are disrupted following damage to the ewe’s olfactory system (Baldwin and Shillito 1974; Lévy et al. 1995; Morgan et al. 1975) or by restricting the ewe’s access to their lamb’s olfactory cues (Poindron and Neindre 1980; Poindron et al. 2007), indicating that chemical cues are important in identification. Offspring identification is based on a lamb’s individual olfactory signature, which is encoded by the genome (Porter et al. 1991; Romeyer et al. 1993). Although olfaction is clearly important in early offspring recognition, ewes can also recognise their lambs through auditory or visual cues (Alexander 1977; Ferreira et al. 2000; Terrazas et al. 1999). The use of multisensory signals to identify young also occurs in other species, including Mexican bats (Balcombe 1990; Gustin and McCracken 1987), laboratory mice (Cohen et al. 2011), Norway rats (Farrell and Alberts 2002) and Australian sea lions (Pitcher et al. 2010a; Pitcher et al. 2010b). Visual and auditory cues may allow recognition at a distance while olfactory cues provide confirmation at close quarters prior to nursing. However, exactly how such multisensory cues are integrated is unclear and is an important area of future study.
Identification of offspring may be particularly important in precocial species that breed in large herds or colonies, such that offspring from multiple females are present (see examples above). Here, females need to discriminate own from alien young to ensure that the benefits of parental care and lactation are received by own offspring. By contrast, in many altricial species, offspring are confined to a single den or nest until weaning (Gubernick and Klopfer 1981). As mothers are unlikely to encounter unrelated young within their nest, recognition of own young may be less important (Holmes and Sherman 1982). A recent study in domestic cats (Felis silvestris catus) found that despite being able to discriminate between the odours of own and alien young, females retrieved both equally (Banszegi et al. 2017). Solitary species, such as cats, may encounter unrelated young so infrequently that there is little need to discriminate them from own offspring. Further, the price of mistakenly rejecting own offspring, for example because they have picked up alien odours from the environment, may outweigh the cost of accepting unrelated young.
Recognition of young is expected to be particularly important for animals that raise their young in a communal nest. However, in many communally nesting species females nurse all young in the nest indiscriminately (degus: Ebensperger et al. 2006; mice: König 1989a; König 1989b; general review: Packer et al. 1992; evening bat: Watkins and Shump 1981), although some species may preferentially suckle own young during early lactation (Watkins and Shump 1981). To investigate the relative investment of communally nesting females, a study of degus (Octon degus) used a radionuclide to track milk transfer from mother to young in communal nests (Jesseau et al. 2009). Unrelated pairs of females preferentially nursed own offspring when sampled at 2 weeks old (though still gave some milk to their co-nesting partner’s offspring), whereas sister pairs nursed all young equally; there was no evidence of discrimination close to weaning at 4 weeks (Jesseau et al. 2009). Female degus can discriminate between body odours from own pups, sister’s pups and unrelated pups (Jesseau et al. 2008) and may use this discriminatory ability to preferentially nurse related offspring.
The ability to recognise own mothers by offspring may also be important in mammals as unfamiliar conspecifics often behave aggressively towards alien young. Although recognition of mothers has been demonstrated in some species, few studies have directly investigated the role of olfaction in recognition (Lickliter and Heron 1984; Poindron et al. 1993; Val-Laillet and Nowak 2008). A study in newborn lambs concluded that auditory and visual cues are more important than olfactory signals in maternal recognition (Nowak 1991). However, lambs were prevented from directly interacting with ewes during the experiment so it is not known whether odours might still facilitate maternal recognition in sheep at close contact.
Olfactory cues are also important in mother-offspring recognition in humans. Mothers can identify their own infant through odour alone (Kaitz et al. 1987; Porter et al. 1983; Schaal et al. 1980). These individually recognisable odours are produced by the infant rather than being deposited on the infant by the mother (Kaitz et al. 1987; Russell et al. 1983) and extensive postnatal interactions are not necessary for recognition to occur (Porter et al. 1983). Similarly, human babies can discriminate between odours emanating from their mother or an unrelated female (Cernoch and Porter 1985; Macfarlane 1975; Schaal et al. 1980). This discrimination occurred in breast and not bottle fed infants, suggesting odour recognition may be a learnt response (Cernoch and Porter 1985). Further support for olfactory learning in infants comes from a study by Schleidt and Genzel (1990), who showed that infants preferred their mother's perfume when lactating mothers perfumed their breasts.
Olfactory-Mediated Suckling Behaviour
Evidence from several mammalian species shows that females release highly specific chemical cues to guide their young to the nipples and elicit suckling behaviour (Arteaga et al. 2013; Schaal and Al Aïn 2014). The best studied example is the European rabbit which releases a mammary pheromone that causes suckling behaviour in their pups (Schaal et al. 2003). This may be a particularly important adaptation in rabbits, which only nurse their pups once a day for approximately 5 minutes (Zarrow et al. 1965). During this time the pups, which are born blind, have to orientate to the female’s abdomen, locate and attach to the nipples and suckle efficiently, in the context of severe sibling competition (Bautista et al. 2005; Drummond et al. 2000). A series of elegant experiments demonstrated that the volatile component of rabbit milk, 2-methylbut-2-enal (2MB2), was responsible for the stereotyped searching-grasping behaviour typically seen in new-born pups (Schaal et al. 2003). 2MB2 acts as a pheromone as it elicits a species specific response (Schaal et al. 2003) and the behavioural activity is independent of learning (Schaal et al. 2003). Further, the releasing effect of 2MB2 is concentration dependent (Coureaud et al. 2004). In rabbits, as with many mammals, the growth and survival of pups is highly dependent on their ability to effectively suckle during the first few days (Coureaud et al. 2000). By increasing neonatal suckling success, 2MB2 increases offspring survival and female reproductive fitness. As well as producing a stereotyped behavioural response, 2MB2 also induces neonatal odour learning (Coureaud et al. 2010). When new-born rabbits are exposed to a mixture of 2MB2 and a neutral odorant (or mixture of odorants, see (Coureaud et al. 2008)), they exhibit a strong search-grasping response towards the neutral odorant alone 24 hours later (Coureaud et al. 2006). This learned response is evident after a single conditioning session and persists for five days after the initial exposure. Pheromone induced learning of odorants may facilitate improved orientation to the dam and localization of the nipple (Coureaud et al. 2010). Further, learning of odours in the nest may allow social recognition of familiar conspecifics, such as the mother or sibling nestmates, upon weaning. Male rabbits tend to avoid conspecifics scented with an odorant they learnt through association with 2MB2 as a neonate, suggesting that one function of neonatal olfactory learning may be to avoid inbreeding later in life (Coureaud et al. 2010).
Similar nipple searching behaviour is seen in other altricial species, although the mechanisms have not been investigated as thoroughly as in rabbits (Schaal and Al Aïn 2014). Murine rodents are attracted to the scent of a lactating female but washing of the nipple removes the searching behaviour seen in pups (Logan et al. 2012; Teicher and Blass 1976). Application of maternal amniotic fluid, saliva or milk restored suckling behaviour in laboratory mice (Logan et al. 2012). However, the same study showed that only amniotic fluid initiated suckling behaviour in pups delivered by caesarean section, suggesting that pups require prior exposure to learn maternal cues (Logan et al. 2012). The authors suggest that suckling behaviour in mice may be initiated by a learned signature odour, similar to those underlying maternal recognition in sheep (Logan et al. 2012). However, under different experimental conditions, Al Aïn et al. (2014) found that attraction to milk and colostrum odours by newborn mice did not require prior exposure. These studies highlight the difficulty in identifying mammalian pheromones and the extent to which different experimental conditions can impact results.
Odours are also important in initiating suckling behaviour in humans. Odour emitted from the breast of lactating females are attractive to newborns (Makin and Porter 1989). Breast odours regulate arousal states (Doucet et al. 2007) and elicit head turning (Makin and Porter 1989; Marlier and Schaal 2005), eye opening (Doucet et al. 2007), oral repsonses (Marlier and Schaal 2005) and crawling (Varendi and Porter 2001) in human newborn babies. The compounds underpinning these behavioural responses in newborns have yet to be identified but are likely to be present in the colostrum, milk or in secretions from specialised Montgomery glands situated in the areola surrounding the nipple (Doucet et al. 2009; Marlier and Schaal 2005; Mizuno and Ueda 2004).
Conclusions
This review highlights the diversity of scent signals produced by female mammals, and the wide range of functions that these fulfil, including competitive signalling, sexual advertisement and facilitation of maternal behaviours. However, female chemical communication still remains poorly understood in the majority of mammals. We suggest several key areas for future study. First, although female scents can communicate a wealth of information about an individual, such as species, sex, reproductive state, health, dominance status and genotype (Brown and Macdonald 1985), the specific compounds or mixtures that signal such information have been identified in very few cases. Developments in molecular techniques have improved both the detection and identification of specific compounds in female scents. However, determination of the functional significance of such compounds, through a combination of behavioural testing and scent manipulation, is an essential requirement. Secondly, most research on chemical signalling in mammals has focused on domesticated and laboratory animals. While providing invaluable insight into both the functions and mechanisms of female chemical signalling, these animals may not necessarily be representative of most wild species. Thirdly, the majority of studies that have investigated scent communication in wild mammals have focused on rodent or primate species. Similarly, many of the functions discussed in this review have been investigated mainly in species with similar social structures; for example, studies investigating female signals of quality and male preference have focused mostly on species where males contribute to parental care. Broader investigation of female chemical signalling across a range of species with different social structures, breeding systems and ecological conditions are needed to explore the factors that influence the evolution of female chemical signals. Finally, understanding how chemical signals integrate with other sensory systems, such as auditory and visual signals, is a major challenge for future research.
References
Abbott D, Hodges J, George L (1988) Social status controls LH secretion and ovulation in female marmoset monkeys (Callithrix jacchus). J Endocrinol 117:329–339
Abbott DH, Saltzman W, Schultz-Darken NJ, Smith TE (1997) Specific neuroendocrine mechanisms not involving generalized stress mediate social regulation of female reproduction in cooperatively breeding marmoset monkeys. Ann N Y Acad Sci 807:219–238
Achiraman S, Archunan G (2005) 3-Ethyl-2,7-dimethyl octane, a testosterone dependent unique urinary sex pheromone in male mouse (Mus musculus). Anim Reprod Sci 87:151–161
Achiraman S, Archunan G (2006) 1-Iodo-2methylundecane, a putative estrus-specific urinary chemo-signal of female mouse (Mus musculus). Theriogenology 66:1913–1920
Achiraman S, Archunan G, Ponmanickam P, Rameshkumar K, Kannan S, John G (2010) 1-Iodo-2 methylundecane [1I2MU]: an estrogen-dependent urinary sex pheromone of female mice. Theriogenology 74:345–353
Al Aïn S, Mingioni M, Patris B, Schaal B (2014) The response of newly born mice to odors of murine colostrum and milk: unconditionally attractive, conditionally discriminated. Dev Psychobiol 56:1365–1376
Albers HE, Rowland CM (1989) Ovarian hormones influence odor stimulated flank marking behavior in the hamster (Mesocricetus auratus). Physiol Behav 45:113–117
Albone ES, Gronnerberg TO (1977) Lipids of the anal sac secretions of the red fox, Vulpes vulpes and of the lion, Panthera leo. J Lipid Res 18:474–479
Alexander G (1977) Role of auditory and visual cues in mutual recognition between ewes and lambs in Merino sheep. Appl Anim Ethol 3:65–81
Alexander RD, Noonan KM (1979) Concealment of ovulation, parental care, and human social evolution. In: Chagnon NA, Irons W (eds) Evolutionary Biology and Human Social Behaviour: An Anthropological Perspective. Duxbury Press, Massachusetts, pp 436–453
Andersson M (1994) Sexual Selection. Princeton University Press, Princeton
Apps PJ (2013) Are mammal olfactory signals hiding right under our noses? Naturwissenschaften 100:487–506
Archie EA, Theis KR (2011) Animal behaviour meets microbial ecology. Anim Behav 82:425–436
Archunan G, Kumar R (2012) 1-Iodoundecane, an estrus indicating urinary chemosignal in Bovine (Bos Taurus). J Vet Sci Technol 3
Archunan G, Rajagopal T (2013) Detection of estrus in Indian blackbuck: behavioural, hormonal and urinary volatiles evaluation. Gen Comp Endocrinol 181:156–166
Arden MA, Dye L (1998) The assessment of menstrual synchrony: comment on Weller and Weller (1997). J Comp Psychol 112:323–324
Arteaga L, Bautista A, González D, Hudson R (2013) Smell, suck, survive: chemical signals and suckling in the rabbit, cat, and dog. In: East ML, Dehnhard M (eds) Chemical Signals in Vertebrates 12. Springer New York, New York, pp 51–59
Babb TE, Terman CR (1982) The influence of social environment and urine exposure on sexual maturation of male prairie deermice (Peromyscus maniculatus bairdi). Res Popul Ecol 24:318–328
Baker AJ, Dietz JM (1996) Immigration in wild groups of golden lion tamarins (Leontopithecus rosalia). Am J Primatol 38:47–56
Balcombe JP (1990) Vocal recognition of pups by mother Mexican free-tailed bats, Tadarida brasiliensis mexicana. Anim Behav 39:960–966
Baldwin BA, Shillito EE (1974) The effects of ablation of the olfactory bulbs on parturition and maternal behaviour in soay sheep. Anim Behav 22:220–223
Banszegi O, Jacinto E, Urrutia A, Szenczi P, Hudson R (2017) Can but don't: olfactory discrimination between own and alien offspring in the domestic cat. Anim Cogn 20:795–804
Barman P, Yadav MC, Kumar H, Meur SK, Ghosh SK (2013) Gas chromatographic-mass spectrometric analysis of chemical volatiles in buffalo (Bubalus bubalis) urine. Theriogenology 80:654–658
Barrett J, Abbott DH, George LM (1990) Extension of reproductive suppression by pheromonal cues in subordinate female marmoset monkeys, Callithrix jacchus. J Reprod Fertil 90:411–418
Barrett J, Abbott DH, George LM (1993) Sensory cues and the suppression of reproduction in subordinate female marmoset monkeys, Callithrix jacchus. J Reprod Fertil 97:301–310
Barrette C (1977) Scent-marking in captive muntjacs, Muntiacus reevesi. Anim Behav 25:536–541
Batzli GO, Getz LL, Hurley SS (1977) Suppression of growth and reproduction of microtine rodents by social factors. J Mammal 58:583–591
Bautista A, Mendoza-Degante M, Coureaud G, Martínez-Gómez M, Hudson R (2005) Scramble competition in newborn domestic rabbits for an unusually restricted milk supply. Anim Behav 70:1011–1021
Beach FA, Gilmore RW (1949) Response of male dogs to urine from females in heat. J Mammal 30:391–392
Beach FA, Jaynes J (1956) Studies of maternal retrieving in rats I: recognition of young. J Mammal 37:177–180
Beehner JC, Bergman TJ, Cheney DL, Seyfarth RM, Whitten PL (2006) Testosterone predicts future dominance rank and mating activity among male chacma baboons. Behav Ecol Sociobiol 59:469–479
Been L, Petrulis A (2008) The neurobiology of sexual solicitation: vaginal marking in female syrian hamsters (Mesocricetus auratus). In: Hurst JL, Beynon RJ, Roberts SC, Wyatt TD (eds) Chemical Signals in Vertebrates 11. Springer New York, pp 231–239
Begg CM, Begg KS, Du Toit JT, Mills MGL (2003) Scent-marking behaviour of the honey badger, Mellivora capensis (Mustelidae), in the southern Kalahari. Anim Behav 66:917–929
Bennett NC (1994) Reproductive suppression in social Cryptomys damarensis colonies—a lifetime of socially-induced sterility in males and females (Rodentia: Bathyergidae). J Zool 234:25–39
Bermant G, Lott DF, Anderson L (1968) Temporal characteristics of the Coolidge effect in male rat copulatory behavior. J Comp Physiol Psychol 65:447–452
Berry RJ, Bronson FH (1992) Life history and bioeconomy of the house mouse. Biol Rev Camb Philos Soc 67:519–550
Birke LIA (1978) Scent-marking and the oestrous cycle of the female rat. Anim Behav 26:1165–1166
Birke LIA (1984) Effects of estradiol and progesterone on scent-marking behavior of female rats. Horm Behav 18:95–98
Birkhead TR, Moller AP (1992) Sperm Competition in Birds: Evolutionary Causes and Consequences. Academic Press, London
Blaustein AR (1981) Sexual selection and mammalian olfaction. Am Nat 117:1006–1010
Blissitt MJ, Bland KP, Cottrell DF (1990) Olfactory and vomeronasal chemoreception and the discrimination of oestrous and non-oestrous ewe urine odours by the ram. Appl Anim Behav Sci 27:325–335
Bloss J, Acree TE, Bloss JM, Hood WR, Kunz TH (2002) Potential use of chemical cues for colony-mate recognition in the big brown bat, Eptesicus fuscus. J Chem Ecol 28:819–834
Bonilla-Jaime H, Vázquez-Palacios G, Arteaga-Silva M, Retana-Márquez S (2006) Hormonal responses to different sexually related conditions in male rats. Horm Behav 49:376–382
Bouchard S (2001) Sex discrimination and roostmate recognition by olfactory cues in the African bats, Mops condylurus and Chaerephon pumilus (Chiroptera: Molossidae). J Zool 254:109–117
Boulet M, Charpentier MJ, Drea CM (2009) Decoding an olfactory mechanism of kin recognition and inbreeding avoidance in a primate. BMC Evol Biol 9:281
Boulet M, Crawford JC, Charpentier MJ, Drea CM (2010) Honest olfactory ornamentation in a female-dominant primate. J Evol Biol 23:1558–1563
Bowers MA, Gregario K, Brame CJ, Matter SF, Dooley JL, Jr. (1996) Use of space and habitats by meadow voles at the home range, patch and landscape scales. Oecologia 105:107-115
Boyer ML, Jemiolo B, Andreolini F, Wiesler D, Novotny M (1989) Urinary volatile profiles of pine vole, Microtus pinetorum, and their endocrine dependency. J Chem Ecol 15:649–662
Bradford NB, Bobby DB, Dewsbury DA (1977) Copulatory behavior of golden hamsters (Mesocricetus auratus). Behaviour 61:180–206
Brant CL, Schwab TM, Vandenbergh JG, Schaefer RL, Solomon NG (1998) Behavioural suppression of female pine voles after replacement of the breeding male. Anim Behav 55:615–627
Briand L, Huet J-C, Perez V, Lenoir G, Nespoulous C, Boucher Y, Trotier D, Pernollet J-C (2000) Odorant and pheromone binding by aphrodisin, a hamster aphrodisiac protein. FEBS Lett 476:179–185
Briand L, Trotier D, Pernollet J-C (2004a) Aphrodisin, an aphrodisiac lipocalin secreted in hamster vaginal secretions. Peptides 25:1545–1552
Briand L, Blon F, Trotier D, Pernollet J-C (2004b) Natural ligands of hamster aphrodisin. Chem Senses 29:425–430
Brown RE (1974) Sexual arousal, the Coolidge effect and dominance in the rat (rattus norvegicus). Anim Behav 22:634–637
Brown RE, Macdonald DW (1985) Social Odours in Mammals. Clarendon Press, Oxford
Buesching CD, Stopka P, MacDonald DW (2003) The social function of allomarking in the European badger (Meles meles). Behaviour 140:965–980
Burgener N, East ML, Hofer H, Dehnhard M (2008) Do spotted hyena scent marks code for clan membership? In: Hurst JL, Beynon RJ, Roberts SC, Wyatt TD (eds) Chemical Signals in Vertebrates 11. Springer, New York, pp 169–177
Burgener N, Dehnhard M, Hofer H, East ML (2009) Does anal gland scent signal identity in the spotted hyaena? Anim Behav 77:707–715
Burger BV (2005) Mammalian semiochemicals. In: Schulz S (ed) The Chemistry of Pheromones and Other Semiochemicals II. Springer, Berlin, Heidelberg, pp 231-278.
Cant MA, Otali E, Mwanguhya F (2001) Eviction and dispersal in co-operatively breeding banded mongooses (Mungos mungo). J Zool 254:155–162
Cant MA, Otali E, Mwanguhya F (2002) Fighting and mating between groups in a cooperatively breeding mammal, the banded mongoose. Ethology 108:541–555
Cerda-Molina AL, Hernández-López L, Chavira R, Cárdenas M, Paez-Ponce D, Cervantes-De la Luz H, Mondragón-Ceballos R (2006) Endocrine changes in male stumptailed macaques (Macaca arctoides) as a response to odor stimulation with vaginal secretions. Horm Behav 49:81–87
Cernoch JM, Porter RH (1985) Recognition of maternal axillary odors by infants. Child Dev 56:1593–1598
Champlin AK (1971) Suppression of oestrus in grouped mice: The effects of various densitied and the possible nature of the stimulus. J Reprod Fertil 27:233–241
Charpentier MJE, Williams CV, Drea CM (2008) Inbreeding depression in ring-tailed lemurs (Lemur catta): genetic diversity predicts parasitism, immunocompetence, and survivorship. Conserv Genet 9:1605–1615
Charpentier MJE, Crawford JC, Boulet M, Drea CM (2010) Message ‘scent’: lemurs detect the genetic relatedness and quality of conspecifics via olfactory cues. Anim Behav 80:101–108
Chenoweth SF, Doughty P, Kokko H (2006) Can non-directional male mating preferences facilitate honest female ornamentation? Ecol Lett 9:179–184
Christian JJ, Lemunyan CD (1958) Adverse effects of crowding on lactation and reproduction of mice and two generations of their progeny. Endocrinology 63:517–529
Clark MM, Galef BG (2002) Socially induced delayed reproduction in female Mongolian gerbils (Meriones unguiculatus): is there anything special about dominant females? J Comp Psychol 116:363–368
Clarke FM, Miethe GH, Bennett NC (2001) Reproductive suppression in female Damaraland mole–rats Cryptomys damarensis: dominant control or self–restraint? Proc R Soc B 268:899–909
Clutton-Brock TH (1991) The Evolution of Parental Care. Monographs in Behavior and Ecology. Princeton University Press, Princeton
Clutton-Brock T (2009) Sexual selection in females. Anim Behav 77:3–11
Clutton-Brock T (2016) Mammal Societies. John Wiley & Sons, London
Clutton-Brock TH, Huchard E (2013) Social competition and selection in males and females. Philos Trans R Soc B 368:20130074
Clutton-Brock TH, Isvaran K (2006) Paternity loss in contrasting mammalian societies. Biol Lett 2:513–516
Clutton-Brock T, McAuliffe K (2009) Female mate choice in mammals. Q Rev Biol 84:3–27
Clutton-Brock TH, Albon SD, Guinness FE (1989) Fitness costs of gestation and lactation in wild mammals. Nature 337:260
Clutton-Brock TH, P. N. M. B, Smith R, McIlrath GM, Kansky R, Gaynor D, O'Riain MJ, Skinner JD (1998) Infanticide and expulsion of females in a cooperative mammal. Proc R Soc B 265:2291-2295
Clutton-Brock TH, Gaynor D, McIlrath GM, Maccoll ADC, Kansky R, Chadwick P, Manser M, Skinner JD, Brotherton PNM (1999) Predation, group size and mortality in a cooperative mongoose, Suricata suricatta. J Anim Ecol 68:672–683
Clutton-Brock TH, Brotherton PNM, Russell AF, O'Riain MJ, Gaynor D, Kansky R, Griffin A, Manser M, Sharpe L, McIlrath GM, Small T, Moss A, Monfort S (2001) Cooperation, control, and concession in meerkat groups. Science 291:478–481
Cohen L, Rothschild G, Mizrahi A (2011) Multisensory integration of natural odors and sounds in the auditory cortex. Neuron 72:357–369
Converse LJ, Carlson AA, Ziegler TE, Snowdon CT (1995) Communication of ovulatory state to mates by female pygmy marmosets, Cebuella pygmaea. Anim Behav 49:615–621
Cooney R, Bennett NC (2000) Inbreeding avoidance and reproductive skew in a cooperative mammal. Proc R Soc B 267:801–806
Coppola DM, Vandenbergh JG (1985) Effect of density, duration of grouping and age of urine stimulus on the puberty delay pheromone in female mice. J Reprod Fertil 73:517–522
Coquelin A, Bronson F (1979) Release of luteinizing hormone in male mice during exposure to females: habituation of the response. Science 206:1099–1101
Côté SD, Festa-Bianchet M (2001) Reproductive success in female mountain goats: the influence of age and social rank. Anim Behav 62:173–181
Coureaud G, Schaal B, Coudert P, Rideaud P, Fortun-Lamothe L, Hudson R, Orgeur P (2000) Immediate postnatal sucking in the rabbit: its influence on pup survival and growth. Reprod Nutr Dev 40:19–32
Coureaud G, Langlois D, Sicard G, Schaal B (2004) Newborn rabbit responsiveness to the mammary pheromone is concentration-dependent. Chem Senses 29:341–350
Coureaud G, Moncomble AS, Montigny D, Dewas M, Perrier G, Schaal B (2006) A pheromone that rapidly promotes learning in the newborn. Curr Biol 16:1956–1961
Coureaud G, Thomas-Danguin T, Le Berre E, Schaal B (2008) Perception of odor blending mixtures in the newborn rabbit. Physiol Behav 95:194–199
Coureaud G, Charra R, Datiche F, Sinding C, Thomas-Danguin T, Languille S, Hars B, Schaal B (2010) A pheromone to behave, a pheromone to learn: the rabbit mammary pheromone. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 196:779–790
Crawford JC, Boulet M, Drea CM (2011) Smelling wrong: hormonal contraception in lemurs alters critical female odour cues. Proc R Soc B 278:122–130
Creel S, Creel NM, Mills MGL, Monfort SL (1997) Rank and reproduction in cooperatively breeding African wild dogs: behavioral and endocrine correlates. Behav Ecol 8:298–306
De Fanis E, Jones G (1995) The role of odour in the discrimination of conspecifics by pipistrelle bats. Anim Behav 49:835–839
delBarco-Trillo J, Sacha CR, Dubay GR, Drea CM (2012) Eulemur, me lemur: the evolution of scent-signal complexity in a primate clade. Philos Trans R Soc B 367:1909–1922
Dewsbury DA (1971) Copulatory behaviour of old-field mice (Peromyscus polionotus subgriseus). Anim Behav 19:192–204
Dewsbury DA (1973) Copulatory behavior of montane voles (Microtus montanus). Behaviour 44:186–202
Digby L (1995) Infant care, infanticide, and female reproductive strategies in polygynous groups of common marmosets (Callithrix jacchus). Behav Ecol Sociobiol 37:51–61
Dixson AF (1980) Androgens and aggressive behavior in primates: a review. Aggress Behav 6:37–67
Dixson, AF (2012) Primate Sexuality: Comparative Studies of the Prosimians, Monkeys, Apes, and Humans, 2nd edn. Oxford University Press, Oxford
Dobly A (2005) Scent marking by common voles Microtus arvalis in the presence of a same-sex neighbour. Acta Theriol 50:343–356
Doucet S, Soussignan R, Sagot P, Schaal B (2007) The “smellscape” of mother's breast: effects of odor masking and selective unmasking on neonatal arousal, oral, and visual responses. Dev Psychobiol 49:129–138
Doucet S, Soussignan R, Sagot P, Schaal B (2009) The secretion of areolar (Montgomery's) glands from lactating women elicits selective, unconditional responses in neonates. PLoS One 4:e7579
Drea CM, Scordato ES (2008) Olfactory communication in the ringtailed lemur (Lemur catta): form and function of multimodal signals. In: Hurst JL, Beynon RJ, Roberts SC, Wyatt TD (eds) Chemical Signals in Vertebrates 11. Springer, New York, pp 91–102
Drickamer LC (1974) Sexual maturation of female house mice: Social inhibition. Dev Psychobiol 7:257–265
Drickamer LC, Mikesic DG (1990) Urinary chemosignals, reproduction, and population size for house mice (Mus domesticus) living in field enclosures. J Chem Ecol 16:2955–2968
Drickamer LC, Vandenbergh JG (1973) Predictors of social dominance in the adult female golden hamster (Mesocricetus auratus). Anim Behav 21:564–570
Drummond H, Vázquez E, Sánchez-Colón S, Martinez-Gómez M, Hudson R (2000) Competition for milk in the domestic rabbit: survivors benefit from littermate deaths. Ethology 106:511–526
East ML, Gusset-Burgener N, Hofer H (2013) Sex differences in olfactory behaviours reflect the importance of scent marking for social integration in adult females and competition between reproductively active males in the spotted hyena. In: East ML, Dehnhard M (eds) Chemical Signals in Vertebrates 12. Springer, New York, pp 149–160
Ebensperger LA (1998) Strategies and counterstrategies to infanticide in mammals. Biol Rev 73:321–346
Ebensperger LA, Hurtado MJ, Valdivia I (2006) Lactating females do not discriminate between their own young and unrelated pups in the communally breeding rodent, Octodon degus. Ethology 112:921–929
Emlen ST (1995) An evolutionary theory of the family. Proc Natl Acad Sci U S A 92:8092–8099
Emlen S, Oring L (1977) Ecology, sexual selection, and the evolution of mating systems. Science 197:215–223
Epple G (1972) Social communication by olfactory signals in marmosets. Int Zoo Yearb 12:36–42
Epple G, Katz Y (1984) Social influences on estrogen excretion and ovarian cyclicity in saddle back tamarins (Saguinus fuscicollis). Am J Primatol 6:215–227
Erb GE, Edwards HE, Jenkins KL, Mucklow LC, Wynne-Edwards KE (1993) Induced components in the spontaneous ovulatory cycle of the Djungarian hamster (Phodopus campbelli). Physiol Behav 54:955–959
Ewer RF (1968) Ethology of Mammals. Logos Press, London
Fadem BH (1990) The effects of gonadal hormones on scent marking and related behavior and morphology in female gray short-tailed opossums (Monodelphis domestica). Horm Behav 24:459–469
Fanis E, Jones G (1996) Allomaternal care and recognition between mothers and young in pipistrelle bats (Pipistrellus pipistrellus). J Zool 240:781–787
Farrell WJ, Alberts JR (2002) Stimulus control of maternal responsiveness to Norway rat (Rattus norvegicus) pup ultrasonic vocalizations. J Comp Psychol 116:297–307
Faulkes CG, Bennett NC (2001) Family values: group dynamics and social control of reproduction in African mole-rats. Trends Ecol Evol 16:184–190
Ferkin MH (1988) The effect of familiarity on social interactions in meadow voles, Microtus pennsylvanicus: a laboratory and field study. Anim Behav 36:1816–1822
Ferkin MH (1999) Meadow voles (Microtus pennsylvanicus, Arvicolidae) over-mark and adjacent-mark the scent marks of same-sex conspecifics. Ethology 105:825–837
Ferkin MH, Johnston RE (1995) Effects of pregnancy, lactation and postpartum oestrus on odour signals and the attraction to odours in female meadow voles, Microtus pennsylvanicus. Anim Behav 49:1211–1217
Ferkin MH, Gorman MR, Zucker I (1991) Ovarian hormones influence odor cues emitted by female meadow voles, Microtus pennsylvanicus. Horm Behav 25:572–581
Ferkin MH, Sorokin ES, Johnston RE, Lee CJ (1997) Attractiveness of scents varies with protein content of the diet in meadow voles. Anim Behav 53:133–141
Ferkin MH, Lee DN, Leonard ST (2004) The reproductive state of female voles affects their scent marking behavior and the responses of male conspecifics to such marks. Ethology 110:257–272
Ferreira G, Terrazas A, Poindron P, Nowak R, Orgeur P, Lévy F (2000) Learning of olfactory cues is not necessary for early lamb recognition by the mother. Physiol Behav 69:405–412
Fischer JB (1954) Evolution and bird sociality. In: Huxley J, Hardy AC, Ford EB (eds) Evolution As A Process. Allen & Unwin, London, pp 71–83
Fischer RB, Brown PS (1993) Vaginal secretions increase the likelihood of intermale aggression in Syrian hamsters. Physiol Behav 54:213–214
Fischer RB, McQuiston J (1991) A possible role for Syrian hamster, Mesocricetus auratus, vaginal secretion in inter-female competition. Anim Behav 42:949–954
Fitzpatrick S, Berglund A, Rosenqvist G (1995) Ornaments or offspring: costs to reproductive success restrict sexual selection processes. Biol J Linn Soc 55:251–260
French JA (1997) Proximate regulation of singular breeding in callitrichid primates. In: Cooperative Breeding in Mammals. Cambridge University Press, New York, pp 34–75
French JA, Cleveland J (1984) Scent-marking in the tamarin, Saguinus oedipus: sex differences and ontogeny. Anim Behav 32:615–623
French JA, Stribley JA (1987) Synchronization of ovarian cycles within and between social groups in golden lion tamarins (Leontopithecus rosalia). Am J Primatol 12:469–478
Fürtbauer I, Mundry R, Heistermann M, Schülke O, Ostner J (2011) You mate, I mate: macaque females synchronize sex not cycles. PLoS One 6:e26144
Garratt M, Vasilaki A, Stockley P, McArdle F, Jackson M, Hurst JL (2011) Is oxidative stress a physiological cost of reproduction? An experimental test in house mice. Proc R Soc B 278:1098–1106
Gattermann R, Ulbrich K, Weinandy R (2002) Asynchrony in the estrous cycles of golden hamsters (Mesocricetus auratus). Horm Behav 42:70–77
Gese EM, Ruff RL (1997) Scent-marking by coyotes, Canis latrans: the influence of social and ecological factors. Anim Behav 54:1155–1166
Getz LL, Dluzen D, McDermott JL (1983) Suppression of reproductive maturation in male-stimulated virgin female microtus by a female urinary chemosignal. Behav Process 8:59–64
Gilfillan GD, Vitale JDT, McNutt JW, McComb K (2017) Spontaneous discrimination of urine odours in wild African lions, Panthera leo. Anim Behav 126:177–185
Gittleman JL (1989) Carnivore group living: comparative trends In: Gittleman JL (ed) Carnivore Behaviour, Ecology and Evolution. Cornell University Press USA,
Gittleman JL, Thompson SD (1988) Energy allocation in mammalian reproduction. Am Zool 28:863–875
Gonzalez-Mariscal G, Melo AI, Zavala A, Beyer C (1990) Variations in chin-marking behavior of New Zealand female rabbits throughout the whole reproductive cycle. Physiol Behav 48:361–365
Gorman ML (1976) A mechanism for individual recognition by odour in Herpestes auropunctatus (Carnivora: Viverridae). Anim Behav 24:141–145
Gorman ML, Kruuk H, Leitch A (1984) Social functions of the sub-caudal scent gland secretion of the European badger Metes metes (Carnivora: Mustelidae). J Zool 203:549–559
Gosling LM (1982) A reassessment of the function of scent marking in territories. Z Tierpsychol 60:89–118
Gosling LM, Roberts SC (2001) Scent-marking by male mammals: cheat-proof signals to competitors and mates. Adv Study Behav 30:169–217
Graham J, Desjardins C (1980) Classical conditioning: induction of luteinizing hormone and testosterone secretion in anticipation of sexual activity. Science 210:1039–1041
Gray GD, Dewsbury DA (1975) A quantitative description of the copulatory behaviour of meadow voles (Microtus pennsylvanicus). Anim Behav 23:261–267
Green JP, Holmes AM, Davidson AJ, Paterson S, Stockley P, Beynon RJ, Hurst JL (2015) The genetic basis of kin recognition in a cooperatively breeding mammal. Curr Biol 25:2631–2641
Greene LK, Drea CM (2014) Love is in the air: sociality and pair bondedness influence sifaka reproductive signalling. Anim Behav 88:147–156
Griffin AS, West SA (2003) Kin discrimination and the benefit of helping in cooperatively breeding vertebrates. Science 302:634–636
Gubernick DJ, Klopfer PH (1981) Parental Care in Mammals. Plenum Press, New York
Gubernick DJ, Nordby JC (1992) Parental influences on female puberty in the monogamous California mouse, Peromyscus californicus. Anim Behav 44:259–267
Gustin MK, McCracken GF (1987) Scent recognition between females and pups in the bat Tadarida brasiliensis mexicana. Anim Behav 35:13–19
Haigh GR (1983) Effects of inbreeding and social factors on the reproduction of young female Peromyscus maniculatus bairdii. J Mammal 64:48–54
Halpin ZT (1986) Individual odors among mammals: origins and functions. In: Rosenblatt JS, Beer C, Busnel M-C, Slater PJB (eds) Advances in the Study of Behavior, vol 16. Academic Press, London, pp 40–70
Handelmann G, Ravizza R, Ray WJ (1980) Social dominance determines estrous entrainment among female hamsters. Horm Behav 14:107–115
Harris RL, Holland BR, Cameron EZ, Davies NW, Nicol SC (2014) Chemical signals in the echidna: differences between seasons, sexes, individuals and gland types. J Zool 293:171–180
Harvey S, Jemiolo B, Novotny M (1989) Pattern of volatile compounds in dominant and subordinate male mouse urine. J Chem Ecol 15:2061–2072
Hayes RA, Morelli TL, Wright PC (2004) Anogenital gland secretions of Lemur catta and Propithecus verreauxi coquereli: a preliminary chemical examination. Am J Primatol 63:49–62
Heistermann M, Kleis E, Pröve E, Wolters H-J (1989) Fertility status, dominance, and scent marking behavior of family-housed female cotton-top tamarins (Saguinus oedipus) in absence of their mothers. Am J Primatol 18:177–189
Henschel JR, Skinner JD (1991) Territorial behaviour by a clan of spotted hyenas Crocuta crocuta. Ethology 88:223–235
Heymann EW (1998) Sex differences in olfactory communication in a primate, the moustached tamarin, Saguinus mystax (Callitrichinae). Behav Ecol Sociobiol 43:37–45
Heymann EW (2006) Scent marking strategies of new world primates. Am J Primatol 68:650–661
Hodge SJ (2009) Understanding variation in reproductive skew: directions for future empirical research. In: Jones CB, Hager R (eds) Reproductive Skew in Vertebrates: Proximate and Ultimate Causes. Cambridge University Press, Cambridge, pp 439–466
Hofer H, East ML (1993) The commuting system of Serengeti spotted hyaenas: how a predator copes with migratory prey. I. Social organization. Anim Behav 46:547–557
Hofmann JE, Getz LL (1988) Multiple exposures to adult males and reproductive activation of virgin female Microtus ochrogaster. Behav Process 17:57–61
Holmes WG, Sherman PW (1982) The ontogeny of kin recognition in two species of ground squirrels. Am Zool 22:491–517
Holst DV, Eichmann F (1998) Sex-specific regulation of marking behavior by sex hormones and conspecifics scent in tree shrews (Tupaia belangeri). Physiol Behav 63:157–164
Honess PE, Marin CM (2006) Behavioural and physiological aspects of stress and aggression in nonhuman primates. Neurosci Biobehav Rev 30:390–412
Hsu F, Nodari F, Kao L, Fu X, Holekamp TF, Turk J, Holy TE (2008) Strucutal characterization of sulfated steroids that activate mouse pheromone-sensing neurones. Biochemistry 47:14009–14019
Huck UW, Lisk RD, Kim S, Evans AB (1989) Olfactory discrimination of estrous condition by the male golden hamster (Mesocricetus auratus). Behav Neural Biol 51:1–10
Hudson R, Vodermayer T (1992) Spontaneous and odour-induced chin marking in domestic female rabbits. Anim Behav 43:329–336
Hurst JL (1987) Behavioral variation in wild house mice Mus domesticus Rutty - a quantitative assessment of female social organization. Anim Behav 35:1846–1857
Hurst JL (1990) Urine marking in populations of wild house mice Mus domesticus rutty. II. Communication between females. Anim Behav 40:223–232
Hurst JL (2009) Female recognition and assessment of males through scent. Behav Brain Res 200:295–303
Hurst JL, Beynon RJ (2004) Scent wars: the chemobiology of competitive signalling in mice. BioEssays 26:1288–1298
Hurst J, Beynon R (2010) Making progress in genetic kin recognition among vertebrates. J Biol 9:13
Hurst JL, Payne CE, Nevison CM, Marie AD, Humphries RE, Robertson DHL, Cavaggioni A, Beynon RJ (2001) Individual recognition in mice mediated by major urinary proteins. Nature 414:631–634
Ilmonen P, Stundner G, Thoß M, Penn DJ (2009) Females prefer the scent of outbred males: good-genes-as-heterozygosity? BMC Evol Biol 9:104
Iwata E, Wakabayashi Y, Kakuma Y, Kikusui T, Takeuchi Y, Mori Y (2000) Testosterone-dependent primer pheromone production in the sebaceous gland of male goat. Biol Reprod 62:806–810
Jemiolo B, Novotny M (1993) Long-term effect of a urinary chemosignal on reproductive fitness in female mice. Biol Reprod 48:926–929
Jemiolo B, Novotny M (1994) Inhibition of sexual maturation in juvenile female and male mice by a chemosignal of female origin. Physiol Behav 55:519–522
Jennions MD, Macdonald DW (1994) Cooperative breeding in mammals. Trends Ecol Evol 9:89–93
Jennions MD, Petrie M (2000) Why do females mate multiply? A review of the genetic benefits. Biol Rev Camb Philos Soc 75:21–64
Jesseau SA, Holmes WG, Lee TM (2008) Mother–offspring recognition in communally nesting degus, Octodon degus. Anim Behav 75:573–582
Jesseau SA, Holmes WG, Lee TM (2009) Communal nesting and discriminative nursing by captive degus, Octodon degus. Anim Behav 78:1183–1188
Johansson BG, Jones TM (2007) The role of chemical communication in mate choice. Biol Rev 82:265–289
Johnson RP (1973) Scent marking in mammals. Anim Behav 21:521–535
Johnston RE (1977) The causation of two scent-marking behaviour patterns in female hamsters (Mesocricetus auratus). Anim Behav 25:317–327
Johnston RE (1979) Olfactory preferences, scent marking, and "proceptivity" in female hamsters. Horm Behav 13:21–39
Johnston RE (1980) Responses of male hamsters to odors of females in different reproductive states. J Comp Physiol Psychol 94:894–904
Johnston RE (1992) Vomeronasal and/or olfactory mediation of ultrasonic calling and scent marking by female golden hamsters. Physiol Behav 51:437–448
Johnston RE, Rasmussen K (1984) Individual recognition of female hamsters by males: role of chemical cues and of the olfactory and vomeronasal systems. Physiol Behav 33:95–104
Johnstone RA, Cant MA (1999) Reproductive skew and the threat of eviction: a new perspective. Proc R Soc B 266:275–279
Jordan NR (2007) Scent-marking investment is determined by sex and breeding status in meerkats. Anim Behav 74:531–540
Jordan NR, Mwanguhya F, Kyabulima S, Rüedi P, Hodge SJ, Cant MA (2011) Scent marking in wild banded mongooses: 3. Intrasexual overmarking in females. Anim Behav 81:51–60
Jordan NR, Golabek KA, Apps PJ, Gilfillan GD, McNutt JW (2013) Scent-mark identification and scent-marking behaviour in African wild dogs (Lycaon pictus). Ethology 119:644–652
Kaitz M, Good A, Rokem AM, Eidelman AI (1987) Mothers' recognition of their newborns by olfactory cues. Dev Psychobiol 20:587–591
Kappeler PM (1998) To whom it may concern: the transmission and function of chemical signals in Lemur catta. Behav Ecol Sociobiol 42:411–421
Karlson P, Luscher M (1959) Pheromones': a new term for a class of biologically active substances. Nature 183:55–56
Kavaliers M, Colwell DD (1995) Discrimination by female mice between the odors of parasitized and non-parasitized males. Proc R Soc B 261:31–35
Keller M, Meurisse M, Poindron P, Nowak R, Ferreira G, Shayit M, Levy F (2003) Maternal experience influences the establishment of visual/auditory, but not olfactory recognition of the newborn lamb by ewes at parturition. Dev Psychobiol 43:167–176
Kerth G, Barbara K (1999) Fission, fusion and nonrandom associations in female Bechstein's bats (Myotis bechsteinii). Behaviour 136:1187–1202
Kerth G, Safi K, Barbara K (2002) Mean colony relatedness is a poor predictor of colony structure and female philopatry in the communally breeding Bechstein's bat (Myotis bechsteinii). Behav Ecol Sociobiol 52:203–210
Kerth G, Perony N, Schweitzer F (2011) Bats are able to maintain long-term social relationships despite the high fission–fusion dynamics of their groups. Proc R Soc B 278:2761–2767
Kimura R (2001) Volatile substances in feces, urine and urine-marked feces of feral horses. Can J Anim Sci 81:411–420
Kimura T, Hagiwara Y (1985) Regulation of urine marking in male and female mice: effects of sex steroids. Horm Behav 19:64–70
Kleiman DG (1966) Scent marking in the Canidae. In: Jewell PA, Loizos C (eds) Play, Exploration and Territory in Mammals, vol 18. Symposia of the Zoological Society of London. Academic Press, London, pp 167-177
Kleiman DG (1977) Monogamy in mammals. Q Rev Biol 52:39–69
Kokko H, Brooks R, Jennions MD, Morley J (2003) The evolution of mate choice and mating biases. Proc R Soc B 270:653–664
König B (1989a) Kin recognition and maternal care under restricted feeding in house mice (Mus domesticus). Ethology 82:328–343
König B (1989b) Behavioural ecology of kin recognition in house mice. Ethol Ecol Evol 1:99–110
König B (1993) Maternal investment of communally nursing female house mice (Mus musculus domesticus). Behav Process 30:61–73
König B (1994) Fitness effects of communal rearing in house mice: the role of relatedness versus familiarity. Anim Behav 48:1449–1457
Koyama S (2004) Primer effects by conspecific odors in house mice: a new perspective in the study of primer effects on reproductive activities. Horm Behav 46:303–310
Koyama S (2016) The primer effects. In: Koyama S (ed) Primer Effects by Murine Pheromone Signaling: Pheromonal Influences on Reproductive Conditions. Springer International Publishing, Cham, pp 23–37
Koyama S, Kamimura S (2000) Influence of social dominance and female odor on the sperm activity of male mice. Physiol Behav 71:415–422
Kruczek M (1997) Male rank and female choice in the bank vole, Clethrionomys glareolus. Behav Process 40:171–176
Kruuk H (1972) The Spotted Hyena: A Study of Predation and Social Behavior. University of Chicago Press, Chicago
Kruuk H (1992) Scent marking by otters (Lutra lutra): signaling the use of resources. Behav Ecol 3:133–140
Kutsukake N, Clutton-Brock TH (2006) Aggression and submission reflect reproductive conflict between females in cooperatively breeding meerkats Suricata suricatta. Behav Ecol Sociobiol 59:541–548
Lai SC, Vasilieva N, Johnston RE (1996) Odors providing sexual information in Djungarian hamsters: evidence for an across-odor code. Horm Behav 30:26–36
Lazaro-Perea C, Snowdon CT, de Fátima Arruda M (1999) Scent-marking behavior in wild groups of common marmosets (Callithrix jacchus). Behav Ecol Sociobiol 46:313–324
Le Roux A, Cherry MI, Manser MB (2008) The effects of population density and sociality on scent marking in the yellow mongoose. J Zool 275:33–40
Leclaire S, Nielsen JF, Drea CM (2014) Bacterial communities in meerkat anal scent secretions vary with host sex, age, and group membership. Behav Ecol 25:996–1004
Leclaire S, Jacob S, Greene LK, Dubay GR, Drea CM (2017) Social odours covary with bacterial community in the anal secretions of wild meerkats. Sci Rep 7:3240
Lévy F, Locatelli A, Piketty V, Tillet Y, Poindron P (1995) Involvement of the main but not the accessory olfactory system in maternal behavior of primiparous and multiparous ewes. Physiol Behav 57:97–104
Liberles SD (2014) Mammalian pheromones. Annu Rev Physiol 76:151–175
Lickliter RE, Heron JR (1984) Recognition of mother by newborn goats. Appl Anim Behav Sci 12:187–192
Lisberg AE, Snowdon CT (2009) The effects of sex, gonadectomy and status on investigation patterns of unfamiliar conspecific urine in domestic dogs, Canis familiaris. Anim Behav 77:1147–1154
Lisk RD, Nachtigall MJ (1988) Estrogen regulation of agonistic and proceptive responses in the golden hamster. Horm Behav 22:35–48
Loebel D, Scaloni A, Paolini S, Marchese S, Fini C, Ferrara L, Breer H, Pelosi P (2001) Chemical communication in the pig. In: Marchlewska-Koj A, Lepri JJ, Müller-Schwarze D (eds) Chemical Signals in Vertebrates 9. Springer, Boston, pp 177–181
Logan DW, Brunet LJ, Webb WR, Cutforth T, Ngai J, Stowers L (2012) Learned recognition of maternal signature odors mediates the first suckling episode in mice. Curr Biol 22:1998–2007
Lucio RA, Rodríguez-Piedracruz V, Tlachi-López JL, García-Lorenzana M, Fernández-Guasti A (2014) Copulation without seminal expulsion: the consequence of sexual satiation and the Coolidge effect. Andrology 2:450–457
Lukas D, Clutton-Brock TH (2013) The evolution of social monogamy in mammals. Science 341:526–530
Lydell K, Doty RL (1972) Male rat odor preferences for female urine as a function of sexual experience, urine age, and urine source. Horm Behav 3:205–212
Ma W, Miao Z, Novotny MV (1998) Role of the adrenal gland and adrenal-mediated chemosignals in suppression of estrus in the house mouse: the Lee-Boot effect revisited. Biol Reprod 59:1317–1320
Macfarlane A (1975) Olfaction in the development of social preferences in the human neonate. CIBA Found Symp:103–117
Macrides F, Bartke A, Fernandez F, D’Angelo W (1974) Effects of exposure to vaginal odor and receptive females on plasma testosterone in the male hamster. Neuroendocrinology 15:355–364
Macrides F, Bartke A, Dalterio S (1975) Strange females increase plasma testosterone levels in male mice. Science 189:1104–1106
Madison DM, McShea WJ (1987) Seasonal changes in reproductive tolerance, spacing, and social organization in meadow voles: a microtine model. Am Zool 27:899–908
Makin JW, Porter RH (1989) Attractiveness of lactating females' breast odors to neonates. Child Dev 60:803–810
Manning CJ, Dewsbury DA, Wakeland EK, Potts WK (1995) Communal nesting and communal nursing in house mice, Mus musculus domesticus. Anim Behav 50:741–751
Marlier L, Schaal B (2005) Human newborns prefer human milk: conspecific milk odor is attractive without postnatal exposure. Child Dev 76:155–168
Maruniak JA, Bronson FH (1976) Gonadotropic responses of male mice to female urine. Endocrinology 99:963–969
Massey A, Vandenbergh JG (1980) Puberty delay by a urinary cue from female house mice in feral populations. Science 209:821–822
Matsumoto-Oda A, Hamai M, Hayaki H, Hosaka K, Hunt KD, Kasuya E, Kawanaka K, Mitani JC, Takasaki H, Takahata Y (2007) Estrus cycle asynchrony in wild female chimpanzees, Pan troglodytes schweinfurthii. Behav Ecol Sociobiol 61:661–668
Maynard Smith J, Harper D (2003) Animal Signals. Oxford University Press. In: Oxford
McClintock MK (1971) Menstrual synchrony and suppression. Nature 229:244
McClintock MK (1978) Estrous synchrony and its mediation by airborne chemical communication (Rattus norvegicus). Horm Behav 10:264–275
McClintock MK (1984) Estrous synchrony: modulation of ovarian cycle length by female pheromones. Physiol Behav 32:701–705
McClintock MK, Adler NT (1978) Induction of persistent estrus by airborne chemical communication among female rats. Horm Behav 11:414–418
McGuire B, Getz L (1991) Response of young female prairie voles (Microtus ochrogaster) to nonresident males: implications for population regulation. Can J Zool 69:1348–1355
McIntosh TK, Drickamer LC (1977) Excreted urine, bladder urine, and the delay of sexual maturation in female house mice. Anim Behav 25:999–1004
McNamara JM, Houston AI (1996) State-dependent life histories. Nature 380:215
Miller SL, Maner JK (2010) Scent of a woman: men’s testosterone responses to olfactory ovulation cues. Psychol Sci 21:276–283
Miller KE, Laszlo K, Dietz JM (2003) The role of scent marking in the social communication of wild golden lion tamarins, Leontopithecus rosalia. Anim Behav 65:795–803
Mills SC, Grapputo A, Jokinen I, Koskela E, Mappes T, Oksanen TA, Poikonen T (2009) Testosterone-mediated effects on fitness-related phenotypic traits and fitness. Am Nat 173:475–487
Mizuno K, Ueda A (2004) Antenatal olfactory learning influences infant feeding. Early Hum Dev 76:83–90
Møller AP, Thornhill R (1998) Male parental care, differential parental investment by females and sexual selection. Anim Behav 55:1507–1515
Monclús R, Saavedra I, de Miguel J (2014) Context-dependent responses to neighbours and strangers in wild European rabbits (Oryctolagus cuniculus). Behav Process 106:17–21
Monfort SL, Bush M, Wildt DE (1996) Natural and induced ovarian synchrony in golden lion tamarins (Leontopithecus rosalia). Biol Reprod 55:875–882
Morelli TL, Hayes RA, Nahrung HF, Goodwin TE, Harelimana IH, MacDonald LJ, Wright PC (2013) Relatedness communicated in lemur scent. Naturwissenschaften 100:769–777
Morgan PD, Boundy CAP, Arnold GW, Lindsay DR (1975) The roles played by the senses of the ewe in the location and recognition of lambs. Appl Anim Ethol 1:139–150
Mossman CA, Drickamer LC (1996) Odor preferences of female house mice (Mus domesticus) in seminatural enclosures. J Comp Psychol 110:131–138
Muller MN (2017) Testosterone and reproductive effort in male primates. Horm Behav 91:36–51
Müller CA, Manser MB (2007) ‘Nasty neighbours’ rather than ‘dear enemies’ in a social carnivore. Proc R Soc B 274:959–965
Müller CA, Manser MB (2008) Scent-marking and intrasexual competition in a cooperative carnivore with low reproductive skew. Ethology 114:174–185
Murdock HG, Randall JA (2001) Olfactory communication and neighbor recognition in giant kangaroo rats. Ethology 107:149–160
Mykytowycz R (1965) Further observations on the territorial function and histology of the submandibular cutaneous (chin) glands in the rabbit, Oryctolagus cuniculus (L.). Anim Behav 13:400–412
Nie Y, Swaisgood RR, Zhang Z, Hu Y, Ma Y, Wei F (2012) Giant panda scent-marking strategies in the wild: role of season, sex and marking surface. Anim Behav 84:39–44
Nielsen BL, Jerôme N, Saint-Albin A, Thonat C, Briant C, Boué F, Rampin O, Maurin Y (2011) A mixture of odorant molecules potentially indicating oestrus in mammals elicits penile erections in male rats. Behav Brain Res 225:584–589
Nodari F, Hsu F, Fu X, Holekamp TF, Kao L, Turk J, Holy TE (2008) Sulfated steroids as natural ligands of mouse pheromone-sensing neurons. JNeurosci 28:6407–6418
Nordeide JT, Kekäläinen J, Janhunen M, Kortet R (2013) Female ornaments revisited – are they correlated with offspring quality? J Anim Ecol 82:26–38
Novotny M, Harvey S, Jemiolo B, Alberts J (1985) Synthetic pheromones that promote inter-male aggression in mice. Proc Natl Acad Sci U S A 82:2059–2061
Novotny M, Jemiolo B, Harvey S, Wiesler D, Marchlewska-Koj A (1986) Adrenal-mediated endogenous metabolites inhibit puberty in female mice. Science 231:722–725
Nowak R (1991) Senses involved in discrimination of merino ewes at close contact and from a distance by their newborn lambs. Anim Behav 42:357–366
Nowak R, Porter RH, Levy F, Orgeur P, Schaal B (2000) Role of mother-young interactions in the survival of offspring in domestic mammals. Rev Reprod 5:153–163
O’Riain MJ, Bennett NC, Brotherton PNM, McIlrath G, Clutton-Brock TH (2000) Reproductive suppression and inbreeding avoidance in wild populations of co-operatively breeding meerkats (Suricata suricatta). Behav Ecol Sociobiol 48:471–477
Ostermeyer M, Elwood R (1983) Pup recognition in Mus musculus: parental discrimination between their own and alien young. Dev Psychobiol 16:75–82
Ostfeld RS (1985) Limiting resources and territoriality in Microtine rodents. Am Nat 126:1–15
Otte D (1974) Effects and functions in the evolution of signaling systems. Annu Rev Ecol Syst 5:385–417
Owen K, Thiessen DD (1974) Estrogen and progesterone interaction in the regulation of scent marking in the female Mongolian gerbil (Meriones unguiculatus). Physiol Behav 12:351–355
Packer C, Scheel D, Pusey AE (1990) Why lions form groups: Food is not enough. Am Nat 136:1–19
Packer C, Lewis S, Pusey A (1992) A comparative analysis of non-offspring nursing. Anim Behav 43:265–281
Palagi E, Telara S, Tarli SMB (2004) Reproductive strategies in Lemur catta: balance among sending, receiving, and countermarking scent signals. Int J Primatol 25:1019–1031
Palagi E, Dapporto L, Borgognini Tarli S (2005) The neglected scent: on the marking function of urine in Lemur catta. Behav Ecol Sociobiol 58:437–445
Palphramand KL, White PCL (2007) Badgers, Meles meles, discriminate between neighbour, alien and self scent. Anim Behav 74:429–436
Pepelko WE, Clegg MT (1965) Studies of mating behaviour and some factors influencing the sexual response in the male sheep Ovis aries. Anim Behav 13:249–258
Pereira ME (1991) Asynchrony within estrous synchrony among ringtailed lemurs (Primates: Lemuridae). Physiol Behav 49:47–52
Petrulis A (2013) Chemosignals, hormones and mammalian reproduction. Horm Behav 63:723–741
Petrulis A, Peng M, Johnston RE (1999) Effects of vomeronasal organ removal on individual odor discrimination, sex-odor preference, and scent marking by female hamsters. Physiol Behav 66:73–83
Pierce JD, O’Brien KK, Dewsbury DA (1992) No effect of familiarity on the Coolidge effect in prairie voles (Microtus ochrogaster). Bull Psychon Soc 30:325–328
Pinter AJ (1985) Effects of hormones and gonadal status on the midventral gland of the grasshopper mouse Onychomys leucogaster. Anat Rec 211:318–322
Pitcher BJ, Harcourt RG, Schaal B, Charrier I (2010a) Social olfaction in marine mammals: wild female Australian sea lions can identify their pup's scent. Biol Lett 7:60–62
Pitcher BJ, Harcourt RG, Charrier I (2010b) Rapid onset of maternal vocal recognition in a colonially breeding mammal, the Australian sea lion. PLoS One 5:e12195
Poindron P, Neindre PL (1980) Endocrine and sensory regulation of maternal behavior in the ewe. In: Rosenblatt JS, Hinde RA, Beer C, Busnel M-C (eds) Advances in the Study of Behavior, vol 11. Academic Press, pp 75–119
Poindron P, Nowak R, Levy F, Porter RH, Schaal B (1993) Development of exclusive mother-young bonding in sheep and goats. Oxf Rev Reprod Biol 15:311–364
Poindron P, Lévy F, Keller M (2007) Maternal responsiveness and maternal selectivity in domestic sheep and goats: The two facets of maternal attachment. Dev Psychobiol 49:54–70
Ponmanickam P, Palanivelu K, Govindaraj S, Baburajendran R, Habara Y, Archunan G (2010) Identification of testosterone-dependent volatile compounds and proteins in the preputial gland of rat Rattus norvegicus. Gen Comp Endocrinol 167:35–43
Porter RH, Fullerton C, Berryman JC (1973) Guinea-pig maternal-young attachment behaviour. Z Tierpsychol 32:489–495
Porter RH, Cernoch JM, McLaughlin FJ (1983) Maternal recognition of neonates through olfactory cues. Physiol Behav 30:151–154
Porter RH, Levy F, Poindron P, Litterio M, Schaal B, Beyer C (1991) Individual olfactory signatures as major determinants of early maternal discrimination in sheep. Dev Psychobiol 24:151–158
Porton I (1983) Bush dog urine-marking: its role in pair formation and maintenance. Anim Behav 31:1061–1069
Preti G, Cutler WB, Garcia CR, Huggins GR, Lawley HJ (1986) Human axillary secretions influence women's menstrual cycles: the role of donor extract of females. Horm Behav 20:474–482
Promislow DEL, Harvey PH (1990) Living fast and dying young: a comparative analysis of life-history variation among mammals. J Zool 220:417–437
Purvis K, Haynes NB (1972) The effect of female rat proximity on the reproductive system of male rats. Physiol Behav 9:401–407
Pusey A, Williams J, Goodall J (1997) The influence of dominance rank on the reproductive success of female chimpanzees. Science 277:828–831
Rajanarayanan S, Archunan G (2011) Identification of urinary sex pheromones in female buffaloes and their influence on bull reproductive behaviour. Res Vet Sci 91:301–305
Ralls K (1971) Mammalian scent marking. Science 171:443–449
Ramm SA, Edward DA, Claydon AJ, Hammond DE, Brownridge P, Hurst JL, Beynon RJ, Stockley P (2015) Sperm competition risk drives plasticity in seminal fluid composition. BMC Biol 13:87
Rasmussen LEL (2001) Source and cyclic release pattern of (Z)-7-Dodecenyl acetate, the pre-ovulatory pheromone of the female Asian elephant. Chem Senses 26:611–623
Rasmussen LEL, Lee TD, Roelofs WL, Zhang A, Daves GD (1996) Insect pheromone in elephants. Nature 379:684–684
Rasmussen LE, Lee TD, Zhang A, Roelofs WL, Daves GD, Jr. (1997) Purification, identification, concentration and bioactivity of (Z)-7-dodecen-1-yl acetate: sex pheromone of the female Asian elephant, Elephas maximus. Chem Senses 22:417-437
Rasmussen LEL, Krishnamurthy V, Sukumar R (2005) Behavioural and chemical conformation of the prevoulatory pheromone, (Z)-dodecenyl acetate, in wild Asian elephants: Its relationship to musth. Behaviour 142:351–396
Raymer J, Wiesler D, Novotny M, Asa C, Seal US, Mech LD (1986) Chemical scent constituents in urine of wolf (Canis lupus) and their dependence on reproductive hormones. J Chem Ecol 12:297–314
Raynaud J, Dobson SF (2011) Scent communication by female Columbian ground squirrels, Urocitellus columbianus. Behav Ecol Sociobiol 65:351–358
Reynolds JD (1996) Animal breeding systems. Trends Ecol Evol 11:68–72
Rich TJ, Hurst JL (1998) Scent marks as reliable signals of the competitive ability of mates. Anim Behav 56:727–735
Rich TJ, Hurst JL (1999) The competing countermarks hypothesis: reliable assessment of competitive ability by potential mates. Anim Behav 58:1027–1037
Richardson PRK (1991) Territorial significance of scent marking during the non-mating season in the Aardwolf Proteles cristatus (Carnivora: Protelidae). Ethology 87:9–27
Richardson HN, Nelson AL, Ahmed EI, Parfitt DB, Romeo RD, Sisk CL (2004) Female pheromones stimulate release of luteinizing hormone and testosterone without altering GnRH mRNA in adult male Syrian hamsters (Mesocricetus auratus). Gen Comp Endocrinol 138:211–217
Roberts SC (2012) On the relationship between scent-marking and territoriality in Callitrichid primates. Int J Primatol 33:749–761
Roberts SC, Dunbar RIM (2000) Female territoriality and the function of scent-marking in a monogamous antelope (Oreotragus oreotragus). Behav Ecol Sociobiol 47:417–423
Roberts SA, Simpson DM, Armstrong SD, Davidson AJ, Robertson DH, McLean L, Beynon RJ, Hurst JL (2010) Darcin: a male pheromone that stimulates female memory and sexual attraction to an individual male's odour. BMC Biol 8:75
Roberts SA, Davidson AJ, Beynon RJ, Hurst JL (2014) Female attraction to male scent and associative learning: the house mouse as a mammalian model. Anim Behav 97:313–321
Romeyer A, Porter RH, Pascal P, Pierre O, Patrick C, Nati P (1993) Recognition of dizygotic and monozygotic twin lambs by ewes. Behaviour 127:119–139
Roney JR, Simmons ZL (2012) Men smelling women: Null effects of exposure to ovulatory sweat on men's testosterone. Evol Psychol 10:703–713
Rood JP (1990) Group size, survival, reproduction, and routes to breeding in dwarf mongooses. Anim Behav 39:566–572
Rose RM, Holaday JW, Bernstein IS (1971) Plasma testosterone, dominance rank and aggressive behaviour in male rhesus monkeys. Nature 231:366
Rosell F, Bjørkøyli T (2002) A test of the dear enemy phenomenon in the Eurasian beaver. Anim Behav 63:1073–1078
Rosell F, Bergan F, Parker H (1998) Scent-marking in the Eurasian beaver (Castor fiber) as a means of territory defense. J Chem Ecol 24:207–219
Royle NJ, Smiseth PT, Kölliker M (2014) The Evolution of Parental Care. Oxford University Press. In: Oxford
Russell MJ, Switz GM, Thompson K (1980) Olfactory influences on the human menstrual cycle. Pharmacol Biochem Behav 13:737–738
Russell MJ, Mendelson T, Peeke HVS (1983) Mother's identification of their infant's odors. Ethol Sociobiol 4:29–31
Safi K, Kerth G (2003) Secretions of the interaural gland contain information about individuality and colony membership in the Bechstein's bat. Anim Behav 65:363–369
Saltzman W, Pick RR, Salper OJ, Liedl KJ, Abbott DH (2004) Onset of plural cooperative breeding in common marmoset families following replacement of the breeding male. Anim Behav 68:59–73
Saltzman W, Digby LJ, Abbott DH (2009) Reproductive skew in female common marmosets: what can proximate mechanisms tell us about ultimate causes? Proc R Soc B 276:389–399
Sankar R, Archunan G (2004) Flehmen response in bull: role of vaginal mucus and other body fluids of bovine with special reference to estrus. Behav Process 67:81–86
Sankar R, Archunan G (2008) Identification of putative pheromones in bovine (Bos taurus) faeces in relation to estrus detection. Anim Reprod Sci 103:149–153
Savage A, Ziegler TE, Snowdon CT (1988) Sociosexual development, pair bond formation, and mechanisms of fertility suppression in female cotton-top tamarins (Saguinus oedipus oedipus). Am J Primatol 14:345–359. https://doi.org/10.1002/ajp.1350140404
Schaal B, Al Aïn S (2014) Chemical signals ‘selected for’ newborns in mammals. Anim Behav 97:289–299
Schaal B, Montagner H, Hertling E, Bolzoni D, Moyse A, Quichon R (1980) Les stimulations olfactives dans les relations entre l'enfant et la mère. Reprod Nutr Dévelop 20:843–858
Schaal B, Coureaud G, Langlois D, Ginies C, Semon E, Perrier G (2003) Chemical and behavioural characterization of the rabbit mammary pheromone. Nature 424:68–72
Schadler MH (1990) Social organization and population control in the pine vole, Microtus pinetorum. In: Tamarin RH, Ostfeld RS, Pugh SR, Bujalska G (eds) Social Systems and Population Cycles in Voles. Birkhäuser, Basel, pp 121–130
Schank JC (2000) Menstrual-cycle variability and measurement: further cause for doubt. Psychoneuroendocrinology 25:837–847
Schank JC (2001a) Measurement and cycle variability: reexamining the case for ovarian-cycle synchrony in primates. Behav Process 56:131–146
Schank JC (2001b) Do Norway rats (Rattus norvegicus) synchronize their estrous cycles? Physiol Behav 72:129–139
Schank JC (2004) Avoiding synchrony as a strategy of female mate choice. Nonlinear Dyn Psychol Life Sci 8:147–176
Schleidt M, Genzel C (1990) The significance of mother's perfume for infants in the first weeks of their life. Ethol Sociobiol 11:145–154
Scordato ES, Drea CM (2007) Scents and sensibility: information content of olfactory signals in the ringtailed lemur, Lemur catta. Anim Behav 73:301–314
Setchell JM, Smith T, Wickings EJ, Knapp LA (2008) Social correlates of testosterone and ornamentation in male mandrills. Horm Behav 54:365–372
Setchell JM, Kendal J, Tyniec P (2011) Do non-human primates synchronise their menstrual cycles? A test in mandrills. Psychoneuroendocrinology 36:51–59
Shulman LM, Spritzer MD (2014) Changes in the sexual behavior and testosterone levels of male rats in response to daily interactions with estrus females. Physiol Behav 133:8–13
Silk JB (2007) The adaptive value of sociality in mammalian groups. Philos Trans R Soc B 362:539–559
Sillero-Zubiri C, Macdonald DW (1998) Scent-marking and territorial behaviour of Ethiopian wolves Canis simensis. J Zool 245:351–361
Simeonovska-Nikolova D (2012) Neighbuor relationships and spacing behaviour of mound-building mouse, Mus spicilegus (Mammalia: Rodentia) in summer. Acta Zool Bulg 64:135–143
Singer AG, Macrides F (1990) Aphrodisin: pheromone or transducer? Chem Senses 15:199–203
Singer AG, Macrides F, Clancy AN, Agosta WC (1986) Purification and analysis of a proteinaceous aphrodisiac pheromone from hamster vaginal discharge. J Biol Chem 261:13323–13326
Singer AG, Clancy AN, Macrides F, Agosta WC, Bronson FH (1988) Chemical properties of a female mouse pheromone that stimulates gonadotropin secretion in males. Biol Reprod 38:193–199
Sliwa A, Richardson PRK (1998) Responses of aardwolves, Proteles cristatus, Sparrman 1783, to translocated scent marks. Anim Behav 56:137–146
Smith LB, Walker WH (2014) The regulation of spermatogenesis by androgens. Semin Cell Dev Biol 30:2–13
Smith JLD, McDougal C, Miquelle D (1989) Scent marking in free-ranging tigers, Panthera tigris. Anim Behav 37:1–10
Snowdon CT (1996) Infant care in cooperatively breeding species. In: Rosenblatt JS, Snowdon CT (eds) Advances in the Study of Behavior, vol 25. Academic Press, pp 643–689
Solomon NG, French JA (1997) Cooperative Breeding in Mammals. Cambridge University Press, Cambridge
Southwick CH (1955a) Regulatory mechanisms of house mouse populations: social behavior affecting litter survival. Ecology 36:627–634
Southwick CH (1955b) The population dynamics of confined house mice supplied with unlimited food. Ecology 36:212–225
Steiger S, Schmitt T, Schaefer HM (2010) The origin and dynamic evolution of chemical information transfer. Proc R Soc B 278:970–979
Stern K, McClintock MK (1998) Regulation of ovulation by human pheromones. Nature 392:177
Stockley P, Bro-Jorgensen J (2011) Female competition and its evolutionary consequences in mammals. Biol Rev Camb Philos Soc 86:341–366
Stockley P, Bottell L, Hurst JL (2013) Wake up and smell the conflict: odour signals in female competition. Philos Trans R Soc B 368
Stoffel MA, Caspers BA, Forcada J, Giannakara A, Baier M, Eberhart-Phillips L, Müller C, Hoffman JI (2015) Chemical fingerprints encode mother–offspring similarity, colony membership, relatedness, and genetic quality in fur seals. Proc Natl Acad Sci U S A 112:E5005–E5012
Stopka P, Janotova K, Heyrovsky D (2007) The advertisement role of major urinary proteins in mice. Physiol Behav 91:667–670
Strassmann BI (1997) The biology of menstruation in Homo sapiens: total lifetime menses, fecundity, and nonsynchrony in a natural-fertility population. Curr Anthropol 38:123–129
Strassmann BI (1999) Menstrual synchrony pheromones: cause for doubt. Hum Reprod 14:579–580
Svare BB (1981) Maternal aggression in mammals. In: Gubernick DJ, Klopfer PH (eds) Parental Care in Mammals. Plenum Press, New York, pp 179–210
Swaisgood RR, Lindburg DG, Zhang H (2002) Discrimination of oestrous status in giant pandas (Ailuropoda melanoleuca) via chemical cues in urine. J Zool 257:381–386
Takahashi LK (1990) Hormonal regulation of sociosexual behavior in female mammals. Neurosci Biobehav Rev 14:403–413
Takahashi LK, Lisk RD (1985) Estrogen action in anterior and ventromedial hypothalamus and the modulation of heterosexual behavior in female golden hamsters. Physiol Behav 34:233–239
Takahashi LK, Lisk RD, Burnett AL (1985) Dual estradiol action in diencephalon and the regulation of sociosexual behavior in female golden hamsters. Brain Res 359:194–207
Taylor GT, Weiss J, Rupich R (1987) Male rat behavior, endocrinology and reproductive physiology in a mixed-sex, socially stressful colony. Physiol Behav 39:429–433
Teicher M, Blass E (1976) Suckling in newborn rats: eliminated by nipple lavage, reinstated by pup saliva. Science 193:422–425
Temeles EJ (1994) The role of neighbours in territorial systems: when are they 'dear enemies'? Anim Behav 47:339–350
Terman CR (1984) Sexual maturation of male and female white-footed mice (Peromyscus leucopus noveboracensis): influence of physical or urine contact with adults. J Mammal 65:97–102
Terrazas A, Ferreira G, Lévy F, Nowak R, Serafin N, Orgeur P, Soto R, Poindron P (1999) Do ewes recognize their lambs within the first day postpartum without the help of olfactory cues? Behav Process 47:19–29
Theis KR, Heckla AL, Verge JR, Holekamp KE (2008) The ontogeny of pasting behavior in free-living spotted hyenas, Crocuta crocuta. In: Chemical Signals in Vertebrates 11. Springer, New York, pp 179–187
Theis KR, Schmidt TM, Holekamp KE (2012) Evidence for a bacterial mechanism for group-specific social odors among hyenas. Sci Rep 2:615
Theis KR, Venkataraman A, Dycus JA, Koonter KD, Schmitt-Matzen EN, Wagner AP, Holekamp KE, Schmidt TM (2013) Symbiotic bacteria appear to mediate hyena social odors. Proc Natl Acad Sci U S A 110:19832–19837
Thiessen D, Rice M (1976) Mammalian scent gland marking and social behavior. Psychol Bull 83:505–539
Thom MD, Stockley P, Jury F, Ollier WER, Beynon RJ, Hurst JL (2008) The direct assessment of genetic heterozygosity through scent in the mouse. Curr Biol 18:619–623
Tobler R, Pledger S, Linklater W (2010) No evidence for ovarian synchrony or asynchrony in hamadryas baboons. Anim Behav 80:829–837
Todrank J, Heth G (2003) Odor–genes covariance and genetic relatedness assessments: rethinking odor-based “recognition” mechanisms in rodents. In: Slater PJB, Rosenblatt JS, Snowdon CT, Roper TJ (eds) Advances in the Study of Behavior, vol 32. Academic Press, pp 77–130
Trevathan WR, Burleson MH, Gregory WL (1993) No evidence for menstrual synchrony in lesbian couples. Psychoneuroendocrinology 18:425–435
Triggs GS (1991) The population ecology of house mice (Mus domesticus) on the Isle of May, Scotland. J Zool 225:449–468
Tzur S, Todrank J, Juergens A, Nevo E, Heth G (2009) Odour–genes covariance within a natural population of subterranean Spalax galili blind mole rats. Biol J Linn Soc 96:483–490
Val-Laillet D, Nowak R (2008) Early discrimination of the mother by rabbit pups. Appl Anim Behav Sci 111:173–182
van Kesteren F, Paris M, Macdonald DW, Millar R, Argaw K, Johnson PJ, Farstad W, Sillero-Zubiri C (2013) The physiology of cooperative breeding in a rare social canid; sex, suppression and pseudopregnancy in female Ethiopian wolves. Physiol Behav 122:39–45
van Noordwijk MA, van Schaik CP (1999) The effects of dominance rank and group size on female lifetime reproductive success in wild long-tailed macaques, Macaca fascicularis. Primates 40:105–130
Vandenbergh JG (1971) The influence of the social environment on sexual maturation in male mice. J Reprod Fertil 24:383–390
Varendi H, Porter RH (2001) Breast odour as the only maternal stimulus elicits crawling towards the odour source. Acta Paediatr 90:372–375
Vogt K, Boos S, Breitenmoser U, Kölliker M (2016) Chemical composition of Eurasian lynx urine conveys information on reproductive state, individual identity, and urine age. Chemoecology 26:205–217
Wade GN, Schneider JE (1992) Metabolic fuels and reproduction in female mammals. Neurosci Biobehav Rev 16:235–272
Walker WH (2011) Testosterone signaling and the regulation of spermatogenesis. Spermatogenesis 1:116–120
Wallace P, Owen K, Thiessen DD (1973) The control and function of maternal scent marking the Mongolian gerbil. Physiol Behav 10:463–466
Wallis J (1985) Synchrony of estrous swelling in captive group-living chimpanzees (Pan troglodytes). Int J Primatol 6:335–350
Waser PM, Jones WT (1983) Natal philopatry among solitary mammals. Q Rev Biol 58:355–390
Wasser SK, Barash DP (1983) Reproductive suppression among female mammals: implications for biomedicine and sexual selection theory. Q Rev Biol 58:513–538
Watkins LC, Shump KA (1981) Behavior of the evening bat Nycticeius humeralis at a nursery roost. Am Midl Nat 105:258–268
Wayne NL, Rissman EF (1990) Effects of photoperiod and social variables on reproduction and growth in the male musk shrew (Suncus murinus). J Reprod Fertil 89:707–715
Weller L, Weller A (1993) Human menstrual synchrony: a critical assessment. Neurosci Biobehav Rev 17:427–439
Weller A, Weller L (1997) Menstrual synchrony under optimal conditions: Bedouin families. J Comp Psychol 111:143–151
Wenhold BA, Rasa OAE (1994) Territorial marking in the yellow mongoose Cynictis penicillata: sexual advertisement for subordinates. Z Säugetierkd 59:129–138
West HER, Capellini I (2016) Male care and life history traits in mammals. NatCommun 7:11854
Whalen RE (1963) Sexual behavior of cats. Behaviour 20:321–342
Whitten WK (1959) Occurrence of anoestrus in mice caged in groups. J Endocrinol 18:102–107
Wickings EJ, Dixson AF (1992) Testicular function, secondary sexual development, and social status in male mandrills (Mandrillus sphinx). Physiol Behav 52:909–916
Wilkinson GS, Baker AEM (1988) Communal nesting among genetically similar house mice. Ethology 77:103–114
Willis C, Poulin R (2000) Preference of female rats for the odours of non-parasitised males: the smell of good genes? Folia Parasitol 47:6–10
Wilson HC (1987) Female axillary secretions influence women's menstrual cycles: a critique. Horm Behav 21:536–550
Wilson HC (1992) A critical review of menstrual synchrony research. Psychoneuroendocrinology 17:565–591
Wilson JR, Kuehn RE, Beach FA (1963) Modification in the sexual behavior of male rats produced by changing the stimulus female. J Comp Physiol Psychol 56:636–644
Wolff JO, Dunlap AS, Ritchhart E (2001) Adult female prairie voles and meadow voles do not suppress reproduction in their daughters. Behav Process 55:157–162
Wolff JO, Mech SG, Thomas SA (2002) Scent marking in female prairie voles: a test of alternative hypotheses. Ethology 108:483–494
Wong BBM, Candolin U (2005) How is female mate choice affected by male competition? Biol Rev 80:559–571
Wyatt TD (2010) Pheromones and signature mixtures: defining species-wide signals and variable cues for identity in both invertebrates and vertebrates. J Comp Physiol A 196:685–700
Yang Z, Schank JC (2006) Women do not synchronize their menstrual cycles. Hum Nat 17:433–447
Ydenberg RC, Giraldeau LA, Falls JB (1988) Neighbours, strangers, and the asymmetric war of attrition. Anim Behav 36:343–347
Ylonen H, Koskela E, Mappes T (1997) Infanticide in the bank vole (Clethrionomys glareolus): occurrence and the effect of familiarity on female infanticide. Ann Zool Fenn 34:259–266
Young AJ, Carlson AA, Monfort SL, Russell AF, Bennett NC, Clutton-Brock T (2006) Stress and the suppression of subordinate reproduction in cooperatively breeding meerkats. Proc Natl Acad Sci U S A 103:12005–12010
Zarrow MX, Denenberg VH, Anderson CO (1965) Rabbit: frequency of suckling in the pup. Science 150:1835–1836
Zhang J-X, Zhang Z-B, Wang Z-W (2001) Scent, social status, and reproductive condition in rat-like hamsters (Cricetulus triton). Physiol Behav 74:415–420
Zhang J-X, Sun L, Zhang J-H, Feng Z-Y (2008) Sex- and gonad-affecting scent compounds and 3 male pheromones in the rat. Chem Senses 33:611–621
Ziegler TE, Epple G, Snowdon CT, Porter TA, Belcher AM, Küderling I (1993) Detection of the chemical signals of ovulation in the cotton-top tamarin, Saguinus oedipus. Anim Behav 45:313–322
Ziegler TE, Schultz-Darken NJ, Scott JJ, Snowdon CT, Ferris CF (2005) Neuroendocrine response to female ovulatory odors depends upon social condition in male common marmosets, Callithrix jacchus. Horm Behav 47:56–64
Acknowledgements
HC is supported by a Natural Environment Research Council (NERC) studentship through the Adapting to the Challenges of a Changing Environment Doctoral Training Partnership. PS and JLH acknowledge support from NERC grant NE/M002977/1.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Coombes, H.A., Stockley, P. & Hurst, J.L. Female Chemical Signalling Underlying Reproduction in Mammals. J Chem Ecol 44, 851–873 (2018). https://doi.org/10.1007/s10886-018-0981-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10886-018-0981-x