
Abstract
The vast majority of eukaryotic transcription occurs in bursts during discrete periods of promoter activity, separated by periods of deep repression and inactivity. Elucidating the factors responsible for triggering transitions between these two states has been extremely challenging, partly due to the difficulties in measuring transcriptional bursting genome-wide, but also due to the vast array of candidate transcriptional and epigenetic factors and their complex and dynamic interactions. Additionally, this long-held view of transcriptional bursting as a two-state process has become increasingly challenged, and a resulting lack in consensus on terminology of the involved events has further complicated our understanding of the molecular mechanisms involved. Here, we review the impact of epigenetics on dynamic gene expression, with a focus on transcription bursting. We summarise current understanding of the epigenetic regulation of transcription bursting and propose new terminology for the interpretation of future results measuring transcription dynamics.
Similar content being viewed by others


Background
Transcription noise: an updated view of transcription bursting
Epigenetic gene regulation plays a major role in establishing and maintaining cell lineage-specific transcription programs, while being able to undergo changes to environmental cues [1]. The epigenetic composition marked by chemical modifications to deoxyribonucleic acid (DNA) and associated histone proteins, is associated with folding of the genome into defined permissive and restrictive chromatin structures and nuclear arrangements. Derangements in epigenetic regulation largely contribute to the initiation and maintenance of a diseased state [2]. The importance of epigenetic gene regulation has become clear with whole-genome mapping, less is known about dynamics and variability of interactions at individual cell level [3]. Epigenetic gene regulation was originally discovered in the context of developmental biology to explain how cells with identical DNA are capable of differentiating into the complex array of cell types. Nowadays, we realise that epigenetic gene regulation is of vital importance for every stage of single and multicellular eukaryotic life. This is exemplified by the fact that a considerably higher percentage of the human genome is devoted to epigenetic regulation of transcription than to the coding of functional proteins. Only 1% of the 3.2 billion base pairs of the human genome codes for protein; however, 80.4% of the genome participates in non-protein-coding regulation of gene expression in at least one cell type of our body [4]. This includes the production of non-coding ribonucleic acid (RNA) and the regulation of chromatin-chromatin interactions.
Gene expression regulation is not solely concerned with an increase or decrease of gene expression, but also with the control of cell-to-cell variability in gene expression, often referred to as transcriptional noise [5]. This variance in messenger RNA (mRNA) content across a population of cells is frequently measured by the coefficient of variation, a mean normalised standard deviation [6]. This noise in gene expression can be divided into two categories, extrinsic and intrinsic noise, depending on the underlying source [7]. Intrinsic and extrinsic respectively refer to sources originating from within the cell or externally. Extrinsic noise generally is a result of (subtle) differences in the cellular microenvironment, affecting processes such as growth rate and cellular volume. Accordingly, these sources of noise will most likely affect all gene expression within each cell uniformly. Intrinsic noise differs in that it results from the randomness of biochemical reactions that involve low molecule numbers within cells, such as transcription factor binding to cognate motifs, and as such, will in principle affect every gene in the same cell uniquely. There are likely many sources of extrinsic and intrinsic noise and multiple mechanisms to buffer them, (e.g. regulation of mRNA decay or protein translation both could serve as mechanisms to buffer intrinsic noise). The Elowitz lab performed a classical biological experiment (in bacteria) to distinguish intrinsic noise from extrinsic noise by measuring the dynamics in gene activity of two reporter genes expressing yellow and cyano fluorescence (YFP and CFP) under control of the same promoter [8]. Without intrinsic noise within a single cell (i.e. identical YFP and CFP expression), the dynamics in fluorescence intensity of the combined signal was noted to be identical in a population of cells. Fluctuations gene expression dynamics of the YFP, CFP combined signal was found to be derived from extrinsic noise. The contribution of intrinsic noise was measured by quantifying the difference in the fluorescence intensity dynamics of YFP and CFP within a single cell. It was concluded that intrinsic noise in gene activity is caused by random fluctuations in biochemical reactions whereas extrinsic noise relates to a fluctuation in shared gene regulation resources or upstream signalling pathways. Undeniably, the greatest source of intrinsic noise stems from the stochastic nature of gene promoters to switch between active and inactive states, with gene expression only occurring in the active, permissive state [9, 10].
The ultimate result on gene expression is that mRNA is produced in short discrete bursts of activity, interspersed by longer periods of relative inactivity [11]. This process is called transcription bursting [12]. Transcription bursting can be defined as discontinuous gene expression over time, in ‘bursts’ or ‘pulses’ of mRNA production resulting from a transcriptionally active ‘ON’ state alternated with an inactive ‘OFF’ state. Cell-to-cell transcription variability (noise) results from the number of RNA molecules produced during the ON state (burst size), the frequency of transitions to the ON state (burst frequency), and the degradation rate of the resulting transcript. Transcription bursting occurs at multiple timescales ranging from minutes to tens of minutes up to hours.
It has recently been demonstrated that fluctuations in gene expression, potentially resulting from transcription bursting, are capable of persisting for several cell divisions [13] in line with the view that transcription bursting and associated gene expression noise may drive phenotypic divergence of otherwise identical cells [14, 15]. On a shorter timescale, however, it provides a way for cells to rapidly respond to a variety of stimuli, and therefore is a key regulatory process of dynamic gene expression. Understanding the molecular mechanisms behind transcription burst regulation is of great importance for a variety of fields, including cancer drug resistance, where heterogeneity in gene expression is associated with poor prognosis [16].
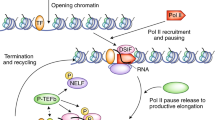
Much of the difficulty in identifying the molecular mechanisms of transcription burst dynamics stems from a lack of consensus on the appropriate model description for transcription bursting. The ‘two-state’ model of transcription is frequently used as a framework for interpreting the results from transcriptional experiments; however, evidence is now mounting for the presence of more than two promoter states [12, 17,18,19,20]. We propose, in agreement with others [19, 20], that transcription bursting can be categorised into distinct OFF and ON promoter states, but that during the ON state, multiple small bursts may occur (microbursts) corresponding to RNA polymerase II (RNAPII) convoys, with short and shallow inactive OFF states in between (Fig. 1A). Accumulation of these microbursts within each ON state produces a larger macroburst. In support of this, cycling of promoters on two separate timescales has been noted [19], namely at a timescale of hours and of minutes. Shallow OFF states were noted to occur for approximately 1.5 min, whereas the deep OFF states were found to extend for much longer time periods, i.e. 9–34 min. It can be considered that multiple RNAPII convoys are released (microbursts) during an ON state before the promoter transitions to the deep OFF state becoming briefly refractory to restimulation.

Transcription bursting and the contribution of epigenetic chromatin composition. A Cartoon representation of transcription bursting in time. On the Y-axis the state of transcriptional access is marked in colour. Green represents the ON state in which transcription takes place, orange the OFF permissive state in which chromatin is accessible for transcription to occur but does not yet take place, red the OFF non-permissive state in which chromatin is inaccessible for transcription to occur. On the X-axis the time illustrating moments of a microburst and macroburst are shown. The large horizontal arrow represents a macroburst whereas the small arrow represents one of the microbursts within a macroburst. The numbers 1–3 represent different moments of transcription dynamics. B Cartoon (simplified) representations of a suggested relationship between the epigenetic composition of the chromatin and the transcriptional state allowing transcription micro- and/or macrobursting with RNAPII progression to occur or resist. (I) DNA methylation of the promoter region will not allow transcription (RNAPII) to access. The transcriptional state will remain in state 1. (II) Turnover and displacement of H3 into H3.3 variant histone is related to transcriptional activity allowing micro- or macrobursts at state 3. (III) Nucleosome decondensation and histone acetylation induce decompaction of the chromatin allowing transcription to occur and produce micro- or macrobursts, going from the OFF permissive state (2) to the ON state (3). (IV) Histone H3K79 methylation relates active transcription. An increase in transcription burst frequency in state 3 is known to be related to the presence of H3K79me2. (V) H3K4 demethylation by histone demethylase KDM5B creates variability in transcription, i.e. going from a state 3 with micro, macrobursts to state 2, being either ON or OFF. (VI) Chromatin looping induces a connection between the enhancer with the promoter thereby allowing transcription into micro- or macrobursts at state 3
A three-state model may help to explain disparities in the interpretation of comparative studies, based purely on the resolution of the method used to determine transcription kinetics [21,22,23,24,25]. For example, studies inferring burst kinetics from single cell transcriptomics generally operate at the macroscale since they lack time resolution and frequently involve fitting of pre-determined models to distribution data [26]. On the other hand, short imaging time courses of transcription in live cells using MS2 or PP7 bacteriophage coat protein binding to specific stem-loop structure (MS2 and PP7, respectively) as reporter systems would generally operate at the micro-scale because they allow very high temporal resolution [11]. Genomically integrated MS2 or PP7 reporter systems enable transcript tagging and microscopical visualisation, making use of the natural interaction of the MS2 or PP7 bacteriophage coat protein with a MS2 or PP7 stem-loop structure from the phage genome [27].
Additionally, for studies based on microscopy and imaging, the interpretation of the results and resolution will be influenced by the length of the investigated gene (assuming a constant RNAPII elongation speed) [28]. For example, MS2/PP7 integration and live cell imaging of transcription of a long gene will lead to the simultaneous progression of multiple RNAPII convoys along the gene length and as such, the macroburst size and transcription frequency will be measured. MS2/PP7 integration within a short gene will result in just one or a few RNAPII convoys elongating at a given time, allowing quantification of the microburst size and frequency. The same applies for burst size quantification based on the analysis of the size and intensity of transcription sites in RNA FISH data of fixed cells [19].
The distinction of transcription burst dynamics within this framework of micro and macrobursting is extremely important for interpreting transcription dynamics, as it will help to compare studies attempting to elucidate the underlying contributions of specific epigenetic factors to transcription bursting.
Main text
Epigenetic regulation of transcription dynamics
By definition, epigenetic modifications impact the process of transcription; however, for many epigenetic modifications, the underlying molecular mechanisms affecting transcription bursting remain poorly understood. This is especially true when considering the array of combinatorial interactions possible between epigenetic factors, where specific combinations may dictate unique mechanisms of transcriptional regulation. Untangling these epigenetic networks may require a more complex systems approach; however, given the recent advances in our understanding of transcription, we here summarise some of the key roles of epigenetics in regulating dynamic gene expression (that is affecting changes within one cell cycle), framed within the exciting context of transcription bursting (Table 1, Fig. 1B).
DNA methylation as a trigger of ON to OFF state transitions driving ‘burstiness’
DNA methylation is the driving force of transgenerational inheritance and genomic imprinting. Even within these contexts, DNA methylation is a more dynamic process than generally appreciated [29]. Active DNA demethylation can be noted during both germline establishment and early embryogenesis [30]. DNA methylation is also capable of responding rapidly to extracellular stimuli, as seen for instance (i) by changes in immune cell DNA methylation following bacterial infection primarily at distal enhancers [31] and (ii) in response to neuronal stimulation (murine post-mitotic neurons), where 1.4% of assessed CpG sites either were noted to rapidly undergo demethylation or de novo methylation [32], for instance following fear conditioning where de novo DNA methyltransferases (DNMTs) were found to be upregulated in the adult rat hippocampus [33]. Our understanding of the cause-consequence relationship between CpG methylation and gene activity is far from complete. Of interest, 87% of the differentially methylated cytosine-phosphate-guanine (CpG) sites were found to induce a decrease in DNA methylation after an associated increase in transcription factor binding and hence increased gene expression [34], suggesting that changes in DNA methylation are rather a consequence of differential gene expression [34,35,36]. In fact, multiple transcription factors have been shown to recruit DNMT3A or DNMT3B and trigger gene promoter hypermethylation [37,38,39]. In a recent study, the effect of CpG methylation on the binding of 542 transcription factors was determined illustrating both decreased and increased transcription factor binding depending on the CpG methylation of associated DNA-binding motifs [40, 41]. As another example, the binding of the architectural protein CTCF in mouse embryonic stem cells (mESCs) has been shown to induce regions of low methylation [42]. There are several well-defined examples of CpG methylation-mediated recruitment of transcription factors, such as with methyl-CpG-binding domain (MBD) containing proteins, e.g. MBD2, MBD3, MBD4 and Methyl-CpG-binding protein 2 (MeCP2) that bind DNA in a sequence-independent fashion, based only on the presence of single 5-methylcytosine bases. Typically, these regulatory proteins can effectively facilitate transcription repression through the recruitment of histone deacetylases or co-repressors, such as histone deacetylase I (HDAC1) and Switch-independent 3A (Sin3A), respectively [43, 44]. More recently, the role of MeCP2 specifically has been updated, i.e. MeCP2 was noted to also induce gene upregulation and recruitment of CAMP responsive element-binding protein 1 (CREB1) to the promoter as noted in ~ 85% of the differentially expressed genes in a mouse model of Rett synrome [45]. In support of this MeCP2 binding was shown to elicit extensive chromatin unfolding, suggesting a role in facilitating gene activity and affecting transcription dynamics and transcription noise [46]. Clearly, the longstanding view of DNA methylation as a non-dynamic and epigenetic mark instructing stable gene expression is challenged and evidence is accumulating that DNA methylation could represent a consequence of transcription factor binding [34,35,36].
For years, the consensus was that cytosine methylation within CpG islands (CGIs) consist of a mechanism to override the pro-transcriptional chromatin state by impairing the binding efficacy of transcriptional activators [47]. CGIs are short (1000 bp) DNA stretches that promote a reduced nucleosome occupancy and a more open chromatin state around gene promoters and transcription sites, facilitating mRNA production through the binding of transcription factors [48]. In addition to CGI-based DNA methylation around promoter sites which is mostly viewed as a repressive player, DNA methylation within gene bodies is in general associated with transcriptional activation [49, 50]. DNA methylation within gene bodies is suggested to either lead to recruitment of elongation factors and/or splicing factors, or to repress transcription from cryptic intragenic promoters [50].
The impact of DNA methylation on transcriptional noise evidently depends on the genomic location of methylated cytosines. Promoter methylation has been implicated with higher levels of transcriptional noise, and gene body methylation with reduced levels of transcriptional noise [51]. The latter finding (replicated in Arabidopsis thaliana and the sea anemone Exaiptasia pallida [52, 53]) supports the hypothesis that gene body methylation acts to suppress intragenic transcription from cryptic promoters resulting in tighter regulation of gene expression. Intragenic DNA methylation could regulate gene expression noise by inhibiting deposition of a histone H2A variant (H2A.Z), which is itself associated with increased transcription noise [54, 55]. DNA methylation at the promoter has been suggested to act as a trigger, forcing ON to OFF state transitions, thereby increasing the ‘burstiness’ of genes, while those promoters that lack DNA methylation are permitted to remain in an active state, reducing transcriptional noise [51]. Indeed in mESCs, DNA methylation was identified as a key regulator of stochastic phenotypic state switching [56]. Conversely, the assessment of DNA methylation at active genes targeted by Polycomb repressive proteins failed to note any impact of DNA methylation on gene expression heterogeneity [10].
Histone (variant) turnover and nucleosome density relate to promoter transition timing
Each nucleosome comprises two copies of each of the canonical histones H3, H4, H2A and H2B, with 147 bp of DNA wrapped around and held in place by the linker histone H1. To facilitate certain biochemical processes, each of these histone proteins have multiple variants, which can be replaced within a cell cycle independently of DNA replication at any site where there is a high nucleosome turnover, for example in response to transcription, DNA damage repair or recombination [57, 58]. This replacement can either drastically alter the nucleosome stability and the array of proteins capable of interacting with it, or the effects can be very subtle. For example, the exchange of H3 for centromeric H3 variant (CENPA) is fundamental for the formation of centromeric chromatin [57], whereas replacement with the H3.3 histone variant appears less consequential [57] and results in a nucleosome almost identical in structure to H3.1 nucleosomes [59]. Of interest, it has been noted that H3.3 turnover time of approximately 2 h is similar to the duration between transcription bursts for (hormone responsive) genes [11, 20, 60] suggesting H3.3 occupancy could act as a form of transcriptional memory. Indeed, H3.3 (specifically the fourth lysine residue) is essential for retention of memory of gene activity states following endoderm nuclei transplantation to enucleated eggs [61].
At both gene promoters and enhancers, the histone chaperone protein histone cell cycle regulation defective homolog A (HIRA), was shown to form complexes with the single stranded DNA-binding replication protein A (RPA) and mediate deposition of H3.3 [62]. This mechanism is restricted to chromatin that is in a decondensed configuration and is dependent on transcription. A second chaperone, death domain-associated protein (DAXX) is capable of H3.3 deposition within condensed inactive chromatin [63] facilitating maintenance of gene repression through recruitment of histone deacetylase II (HDACII) [64]. These opposing genomic contexts for H3.3 make it unlikely that H3.3 functions independently to regulate gene expression, but rather depends on other colocalising features, such as chaperone recruitment [65].
H2A is another histone subunit which has been shown to have multiple variants and functions. One of these histone variants, H2A.Z is implicated in both gene activation and gene silencing, possibly through the regulation of transcription factor recruitment, such as OCT4 [65, 66], and may be key in regulating expression noise and phenotypic variance [67]. Promoters which are poised for expression and those that are actively transcribing are enriched for H2A.Z; however, only active genes show high levels of acetylated H2A.Z indicating that the presence of the H2A.Z variant and its acetylation are crucial for determining regulation [68]. Similarly, a second H2A variant, macroH2A (mH2A) has been implicated in both gene expression and repression. Here, recruitment of Nuclear Respiratory factor 1 (NRF-1) by mH2A nucleosomes reduced transcriptional noise possibly by ensuring consistent interactions with the transcriptional machinery (and maintaining the ON state) while mH2A NRF-1 free nucleosomes could promote stochastic switching of promoters between ON and OFF states, exacerbating expression noise [69].
Finally, the density of nucleosomes around the promoter region, irrespective of the histone variant composition, has been linked to burst frequency and gene expression noise [70]. Here, based on both deoxyribonuclease I (DNase I) sensitivity and analysis of RNA with green fluorescent protein (GFP) expression distributions of various clones following lentiviral reporter integration, it was found that a higher density of promoter-proximal nucleosomes induced a drop in burst frequency and an increase in expression noise, i.e. decreasing the rate of promoter transitions from the OFF to ON state.
Histone post-translational modifications influence the pre/absence of transcriptional consistency
The most widely studied histone post-translational modification (PTM) with respect to gene expression dynamics, and particularly transcriptional noise is acetylation. Generally speaking, histone acetylation demarcates active genes, promoting chromatin permissiveness and gene expression. Histone H4 lysine 20 acetylation (H4K20ac), enriched at transcription start sites of lowly expressed genes appears to be an exception [71]. This consistency in effect made histone acetylation an attractive first candidate for studying the epigenetic regulation of active transcription, especially when considering that most drugs modulating histone acetylation are non-specific, acting on a range of proteins. For example, the commonly used Food and Drug Administration (FDA)-approved histone acetylation inhibiting cancer drug Suberoyl anilide hydroxamic acid (SAHA) (tradename Vorinostat) which inhibits all class I, II, and IV histone deacetylases (HDACs) leading to a universal increase in histone and protein acetylation. Initial studies with HDAC inhibitors, in this case trichostatin (TSA), revealed responses with large context dependency, where for some promoters an increase in transcription burst size was noted while the transcriptional dynamics of other promoters did not change [25]. Although some studies with HDAC inhibitors also show modulation of transcription burst frequency [72] evidence is accumulating to support a main role of histone acetylation in burst size regulation. For example, both TSA and SAHA were found to increase the burst size of latent Human immunodeficiency virus (HIV) production in response to tumour necrosis factor (TNF) via an increase in initiated or paused RNAPII [73] and in a PP7-transcript tagged cell line, live cell imaging of transcription following treatment with the HDAC inhibitor sodium butyrate revealed the transcription burst size to be predominantly affected [60]. The recent rise of the clustered regularly interspaced short palindromic repeats (CRISPR) with CRISPR-associated Cas protein (CRISPR/Cas) platform using mutated inactive, dead Cas (dCas)-tagged fusion proteins allows for precise epigenetic rewriting at defined genes and therefore determines the cause-consequence relationship between the presence of histone modifications and transcription bursting. So far, it has been shown that targeted site-specific acetylation predominantly influences transcription burst frequency, for instance of the circadian brain and muscle Arnt-like protein-1 (Bmal1) promoter [74]. Conversely, site-specific enhancer acetylation has been shown to increase transcription burst duration (size) of the Fos gene family gene (FOS) target gene in neurons, whereas deacetylation of the same site resulted in a decreased burst frequency [75]. Reader protein Bromodomain containing 4 (BRD4) was suggested to be the mediator of this effect, recruited by enhancer histone acetylation and regulating the release and elongation of paused RNAPII.
There are far fewer examples of histone methylation-mediated effects on transcription dynamics with respect to transcription bursting. One modification which has received considerable attention is Histone H3 lysine 4 methylation (H3K4me), having been repeatedly associated with transcriptional memory. As mentioned above, H3.3 was found to mediate memory following nuclei transplantations, but this ability was dependent on the ability to methylate lysine 4 on the histone tail [61]. H3K4 di- and tri-methylation (H3K4me3 and H3K4me2, respectively) are also important histone marks for transcriptional memory in yeast, persisting along the gene body for over 60 min after transcription has ceased [76]. The presence of these epigenetic marks may act to regulate the refractory period between transcription bursts, controlling transcription burst frequency. Indeed, H3K4me was necessary to transmit burst frequency between mother and daughter Dictyostelium cells, maintaining both high and low burst frequencies [77]. More recently, the H3K4-specific demethylase Jumonji histone demethylase (KDM5B) was shown to promote transcriptomic heterogeneity in breast cancer cells, resulting in a reduced sensitivity to anti-oestrogen therapy and higher risk of drug resistance [16]. Here, broad peaks of H3K4me3 were found to be associated with more consistent homogenous expression, while narrow peaks were associated with heterogenous expression. This provides clear evidence of a link between H3K4me and transcriptomic heterogeneity; however, the underlying mechanism remains unclear.
Genomic context dependency of histone marks, particularly as a result of using HDAC inhibitors is not surprising given the number of histone residues capable of undergoing modifications and their diverse mechanisms of action. Regarding histone H3 lysine acetylation (H3Kac) for example, H3K14ac, H3K122ac and H3K64ac all facilitate gene expression by promoting disassembly or eviction of nucleosomes [78,79,80], whereas H3K9ac and H3K27ac are more associated with recruitment of the super elongation complex (SEC) promoting RNAPII pause release [81, 82]. An interesting hypothesis is that localisation of histone modifications within the gene mediates their effect on transcription, where promoter-associated marks could regulate both transcription burst size and burst frequency, while marks located along the gene body mediate burst frequency only [83]. In support of this, knocking out disruptor of telomeric silencing-1-like histone methyltransferase (DOT1L), the epigenetic writer of histone H3 lysine 79 di-methylation (H3K79me2) (a gene body associated mark) resulted in a reduced transcription burst frequency [83], which is consistent with the broad H3K4me3 peaks (extending into gene bodies) being associated with lower levels of gene expression variability.
Chromatin architecture constrains gene activity
Transcription regulation is mostly considered from the viewpoint that cells are molecular units with dedicated transcriptional reactions at a locus-by-locus basis. But in fact gene activity occurs in a highly complex, interrelated dense nanoenvironment, involving the physical organisation of the genome, which massively affects involved chemical reactions such as accessibility, diffusion and free energy of chemical reactions [84]. The physical chromatin architecture occurs at a wide range of scales, from the folding of DNA around histone proteins (< 10 nm) into nucleosomes, up to micron scale hetero- and euchromatin domains within chromosomal territories consisting of defined chromatin compartments and sub-Mega base pair (Mbp)-sized domains. Increased heterogeneity of the physical chromatin structure correlates with increased heterogeneity of gene networks [85].
Nucleosomes enable packaging of DNA at the smallest scale, but these nucleosomes are themselves subjected to macro-level organisation producing larger-scale structures that constitute chromatin. With the advent of DNA fluorescence in situ hybridisation (FISH), this macro-level of genomic architecture was revealed, initially showing that individual chromosomes tend to self-associate within nuclei [86]. Locally confined chromatin interactions can be explained by confined diffusion, with a range approx. 0.5 μm in human fibrosarcoma (HT1080) cells [87]; however, this explanation is not compatible with the many apparent long range interactions that far exceed this radius. How these interactions are formed remains to be elucidated, but it could involve temporal elevation in chromatin diffusion coefficients, or active targeting [88]. Chromatin localisation at the nuclear periphery is often linked with gene inactivity [89] and artificially induced tethering of chromosomes to the nuclear periphery was found to impinge upon gene expression in a histone deacetylase dependent manner [90]. While this implies a link between the macro-level of chromatin arrangements, histone acetylation and gene regulation, the causalities of the relationships and how they relate to transcription bursting have yet to be determined.
Chromatin Capture technologies noted that chromosomes are organised in two spatial compartments, referred to as compartment A and B, depicting distinct properties. Compartment A tends to be gene rich, has a high guanine-cytosine (GC) content, consists of histone marks related to gene activity and is frequently positioned in the interior of the nucleus. Compartment B is rather gene poor, contains compact chromatin, is mostly enriched with histone marks related to gene inactivity and is localised at the periphery of the nucleus. Within A and B compartments smaller topologically associated domains (TADs) can be identified. These domains are of sub-Mega base pair (Mbp) size (median of approximately 800 kb in mice [91]), the boundaries of which are demarcated by convergent-binding sites of CCCTC-binding factor (CTCF) supposedly acting as insulator elements. There is a higher density of transcription start sites at the boundaries of TADs and a strong enrichment of housekeeping genes. It is suggested that cell identity-related gene expression and TAD-scale genomic architecture are in some way connected [92]. This is exemplified by the finding in mice that disruption of TAD organisation can lead to aberrant regulation of Ephrin type-A-receptor 4 gene (Epha4) that is related to development in the neural system, with subsequent changes in gene expression and limb malformation [93]. However, the ubiquity of this relationship between TAD chromatin organisation and gene expression was challenged when Hi-C and matched RNA sequencing data from genomically unstable D. melanogaster revealed that topological rearrangements rarely affected gene expression [94]. This could indicate a reversed causality, i.e. perhaps gene expression is responsible for the maintenance of TAD formation. Contrary to this, dCas9-mediated gene activation was found to be insufficient for inducing TAD boundary formation during mouse neural differentiation [95]. It has been suggested that TADs act as the primary unit of inheritance [96], being evolutionarily conserved across species and cell types [92]. As such, while there is evidence that TADs play a role in constraining gene expression (or vice versa), they are unlikely to contribute to the acute temporal transcriptional changes, i.e. transcription bursting.
Taken together, the evidence presented suggests that macro-level chromatin architecture such as chromatin compartments and TADs tend to be associated with differentiation and stable gene expression patterns being less likely to directly contribute to dynamic changes in gene expression within a cell cycle. Still the physical chromatin organisation may represent an independent ‘folding code’ regulating the global expression dynamics of genes [97]. In comparison with the relatively well-characterised role of histone chemical modifications acting at the level of individual genes, the folding code might act at a global gene expression level as a common denominator independent of the involved molecular pathway.
Cis-regulatory enhancer interactions adjust transcription bursting dynamics
Enhancers are short, distally located regulatory DNA sequences that can increase transcriptional output via interactions with target gene promoters, dictating spatial and temporal gene expression patterns that drive cell lineage specification and greatly contribute to the complexity of eukaryotic organisms [98]. Active enhancers can be identified by co-occurrence of histone H3 lysine 4 mono-methylation (H3K4me1) and histone H3 lysine 27 acetylation (H3K27ac), in addition to recruitment and binding of specific chromatin remodelling enzymes such as E1A-associated protein histone acetyltransferase p300 [99]. Enhancer-promoter loops often span as much as 3 Mbp [100]; however, the causality of these contacts has yet to be elucidated, with most studies simply observing simultaneous looping and gene activation. Absence of these interactions as measured with chromatin capture analysis, i.e. in high-throughput chromosome conformation capture (Hi-C) contact maps, suggests that enhancer-promoter coupling is far more dynamic than TAD boundary formation. This is because Hi-C measurements typically average across the whole population of cells, so dynamics interactions present in subsets of cells at any given time are lost. One fundamental consequence of TADs is that they limit unnecessary contacts between sections of chromatin in adjacent TADs, while increasing the frequency of intra-TAD contacts. Consequently, cis-regulatory contacts occur primarily within TADs with only one third of the interactions occurring across TAD boundaries [101].
TAD boundaries are partially defined by the convergent binding of CTCF; however, not all CTCF binding is associated with TAD formation, not surprising given that there are 40,000 to 80,000 CTCF binding sites in the mammalian genome [102]. A second role of CTCF is to mediate enhancer promoter loop formation. In a recent study, specific CTCF sites were deleted from mouse T Helper 2 (Th2) cells and inducing a markedly increased gene expression noise as a consequence of reduced enhancer-promoter loop stability indicating that CTCF-mediated cis interactions could facilitate consistent gene expression, i.e. an increased transcription frequency or reduced burst size [103]. Another study attempting to do the opposite by forcing enhancer-promoter loop formation found that increasing the contact frequency between an enhancer and promoter induced an increase in the frequency of transcription bursts but not their size [104]. Similarly, in a synthetic reporter gene system in Drosophila, integration of a stronger enhancer was linked to an increased transcription burst frequency, while not affecting the amplitude or duration of transcription bursts [105]. On a larger scale, it was recently reported that enhancers regulate transcription burst dynamics primarily through burst frequency modulation genome-wide [106]. Additionally, the mediator complex responsible for linking enhancers to cognate promoters has been found to drive the rapid succession of initiation events that ultimately results in a burst of RNAPII activity, while not affecting the frequency or duration of transcriptional ‘OFF’ periods [19].
Precisely how enhancers could regulate transcription burst dynamics has yet to be determined. On the one hand, statistical assessment of burst dynamics from two alleles competing for the same enhancer indicates a model of rapid, short-lived enhancer-promoter contacts [104], whereas contradictory findings in a dual reporter system in Drosophila showed that one enhancer can induce temporally simultaneous responses [105]. Of interest, RNAPII recruitment rate was found to not vary, so it seems likely that enhancer-promoter looping could increase the transcription initiation rate, or reduce the duration of promoter-proximal pausing [107]. Given the finding that enhancer-promoter contacts can persist throughout transcription elongation [108], a second model has been proposed whereby the enhancer travels with RNAPII [109]. The overall mechanism of enhancer-promoter regulated transcription bursting is far from clear.
Conclusions
Epigenetics plays a crucial role in mediating the transcriptional responses required by cells to differentiate during development and to survive in changing environments. In the former case epigenetic marks tend to accumulate as the cells rapidly divide, forcing lineage specification; however, in the latter case, epigenetic marks may be deposited or removed independently of the cell cycle. Interrogating the contribution of these more dynamic epigenetic changes to the equally dynamic transcription bursting is something which has only been possible in recent years due to advances in modern biochemical and imaging techniques. Understanding these contributions is somewhat clouded by a lack in consensus over the correct conceptual model of transcription bursting. Here we have reviewed the contribution of epigenetic features to dynamic transcription bursting and we summarise our considerations based on a three-state model of macro- and microbursting.
Promoter-proximal DNA methylation is associated with lower levels of transcriptional noise and is a well-defined repressive epigenetic mark. It is therefore likely that it acts to limit the frequency of promoter transitions from the OFF to ON state, reducing macroburst frequency. The role of DNA methylation within the gene body is less well defined, but it may prevent aberrant binding of the transcriptional machinery and transcription from cryptic promoters. Indeed, for most experimental techniques, this cryptic transcript production would be indistinguishable from the intended canonical RNA production; thus, intragenic DNA methylation would reduce the burst size. Whether this is the true function of DNA methylation for transcription bursting remains to be seen, alternatively DNA methylation may act together with other epigenetic marks, e.g. repressing deposition of histone variants or PTMs.
Histone variant deposition is transcriptionally dependent and therefore unlikely to contribute to the frequency of transcriptional macrobursts but once in the ON state, variant deposition could facilitate microburst frequency/size. Proof for this comes from the finding that the mH2A variant has been shown to maintain the promoter ON state and thus the macroburst size. It has however been suggested that it is the decay of transcriptionally deposited histone variants that acts as a clock, dictating the duration that a certain promoter state is maintained. The link between histone variants such as H3.3 and transcriptional memory is in support of this clock-based decay of histone variants. For promoter-proximal regions, lower density of nucleosomes appears to drive deep OFF to ON state transitions increasing the frequency of macrobursts.
Regarding histone PTMs each of the chemical moiety versus histone residue combinations may affect transcription dynamics in a unique way and this may also depend on other colocalising histone marks. Nevertheless, some progress has been made in deciphering the role of each histone PTM on transcriptional bursting. At the broader scale, much evidence now links changes in histone acetylation levels with burst size regulation but whether this is through macro- or microburst size regulation is unclear. Conversely, site-specific epigenetic interference at the promoter appeared to result in changes in transcription burst frequency at a macroscale. This discrepancy could stem from the non-specific nature of epigenetic acetylation modifying drugs which modulate acetylation levels globally, resulting in secondary effects and obscuring the more subtle site-specific effects. Evidence for this comes from the finding that modulating acetylation at promoter and enhancer sites has conflicting effects on burst dynamics.
Many of the effects of histone methylation appear to mediate the transcriptional memory of cells, such as with H3K4me3 transmitting transcriptional status to daughter cells following cell division and to enucleated cells after nuclear transplantations. Findings in Dictyostelium that H3K4me3 can transmit both high and low frequency of transcription bursts suggests that the enrichment of H3K4me3 only potentiates a pre-existing state, and does not directly regulate burst dynamics. When assayed in breast cancer cells, broad H3K4me3 peaks were associated with transcriptional consistency which may result from increased macroburst frequency or decreased macroburst size. Unlike promoter-associated H3K4me3, H3K36me3 is found along gene bodies of actively transcribing genes and is suggested to trigger repeated rounds of RNAPII initiation increasing frequency of micro- or macrobursts.
Enhancer promoter contacts appear to dictate microburst frequency over burst size, based on high-resolution live cell imaging in Drosophila [105] and RNA FISH transcription site intensity analysis (of a short gene [104]). This suggests that enhancer-promoter loops are very dynamic, possibly triggering the release of paused RNAPII convoys (microbursts), in agreement with the mediator complex findings. Genome-wide inference of transcription burst size and burst frequency revealed that enhancers primarily regulate transcription burst frequency but this undoubtedly takes place at the macroscale.
While much progress has been made investigating the effects of epigenetics on transcription dynamics based on bulk cell techniques or targeted imaging approaches, the overall picture is far from clear. Much of the evidence is swayed by local genomic or epigenomic context. Eliminating this context dependent variation will probably require high throughput imaging experiments to monitor transcription at thousands of loci following epigenetic perturbations. With these kinds of techniques on the horizon, and the recent explosion in single cell transcriptomics, it will not be long before the true genome-wide effects of epigenetics on transcription bursting are identified.
Availability of data and materials
This manuscript is based on published literature.
Abbreviations
- Bmal1:
-
Brain and muscle Arnt-like protein-1
- BRD4:
-
Bromodomain containing 4
- Cas:
-
CRISPR associated protein
- CENPA:
-
Centromeric H3 variant
- CGIs:
-
CpG islands
- CpG:
-
Cytosine-phosphate-guanine
- CREB1:
-
CAMP responsive element binding protein 1
- CRISPR:
-
Clustered regularly interspaced short palindromic repeats
- CTCF:
-
CCCTC-binding factor
- DAXX:
-
Death domain-associated protein
- CFP:
-
Cyanofluorescent protein
- dCas:
-
Dead (mutated) Cas
- dCas9:
-
Dead (mutated) Cas9
- DNA:
-
Deoxyribonucleic acid
- DNase I:
-
Deoxyribonuclease I
- DNMT3:
-
DNA methyltransferase 3A
- DNMT3A:
-
DNA methyltransferase 3A
- DNMT3B:
-
DNA methyltransferase 3B
- DOT1L:
-
Disruptor of telomeric silencing-1-like (histone methyltransferase)
- Epha4:
-
Ephrin type-A-receptor 4 gene
- FDA:
-
Food and Drug Administration
- FISH:
-
Fluorescence in situ hybridisation
- FOS:
-
Fos gene family gene
- GFP:
-
Green fluorescent protein
- GC:
-
Guanine-cytosine content
- H1:
-
Histone H1
- H2A:
-
Histone H2A
- H2A.Z:
-
Histone H2A.Z variant
- H2B:
-
Histone H2B
- H3:
-
Histone H3
- H3.1:
-
Histone H3.1 variant
- H3.3:
-
Histone H3.3 variant
- H3Kac:
-
Histone H3 lysine acetylation
- H3K122ac:
-
Histone H3 lysine 12 acetylation
- H3K14ac:
-
Histone H3 lysine 14 acetylation
- H3K27ac:
-
Histone H3 lysine 27 acetylation
- H3K36me3:
-
Histone lysine 36 tri-methylation
- H3K4me:
-
Histone H3 lysine 4 methylation
- H3K4me1:
-
Histone H3 lysine 4 mono-methylation
- H3K4me2:
-
Histone H3 lysine 4 di-methylation
- H3K4me3:
-
Histone H3 lysine 4 tri-methylation
- H3K64ac:
-
Histone H3 lysine 64 acetylation
- H3K79me2:
-
Histone lysine 79 di-methylation
- H3K9ac:
-
Histone H3 lysine 9 acetylation
- H4:
-
Histone H4
- H4K20ac:
-
Histone H4 lysine 20 acetylation
- HDAC:
-
Histone deacetylase
- HDACI:
-
Histone deacetylase 1
- HDACII:
-
Histone deacetylase II
- Hi-C:
-
High-throughput chromosome conformation capture
- HIRA:
-
Histone cell cycle regulation defective homolog A (histone chaperone protein)
- HIV:
-
Human immunodeficiency virus
- HT1080 cells:
-
Human fibrosarcoma cells
- KDM5B:
-
Jumonji histone demethylase subfamily member
- mH2A:
-
Histone variant macroH2A
- MBD:
-
Methyl-CpG-binding domain protein
- MBD2:
-
Methyl-CpG-binding domain protein 2
- MBD3:
-
Methyl-CpG-binding domain protein 3
- MBD4:
-
Methyl-CpG-binding domain protein 4
- Mbp:
-
Mega base pairs
- MeCP2:
-
Methyl-CpG-binding protein 2
- mESCs:
-
Mouse embryonic stem cells
- mRNA:
-
Messenger ribonucleic acid
- MS2:
-
Bacteriophage coat protein binding to specific stem-loop structure
- MYC:
-
Myelocytomatosis oncogene
- NF-kB:
-
Nuclear factor kappa-light-chain-enhancer of activated B cells
- NRF-1:
-
Nuclear respiratory factor 1 (transcription factor)
- OCT4:
-
Octamer-binding transcription factor 4
- p300:
-
E1A-associated protein (histone acetyltransferase)
- PP7:
-
Bacteriophage coat protein binding to specific stem-loop structure
- PTM:
-
Post-translational modification
- RNA:
-
Ribonucleic acid
- RNAPII:
-
RNA polymerase II
- RPA:
-
Replication protein A
- SAHA:
-
Suberoyl anilide hydroxamic acid (histone deacetylase inhibitor, trade name Vorinostat)
- SEC:
-
Super elongation complex
- SETD2:
-
SET domain containing 2 histone lysine methyltransferase
- Sin3A:
-
Switch-independent 3A
- smFISH:
-
Single molecule fluorescence in situ hybridisation
- TAD:
-
Topologically associated domain
- Th2:
-
T Helper 2 cells
- TNF:
-
Tumour necrosis factor
- TSA:
-
Trichostatin (histone deacetylase inhibitor)
- YFP:
-
Yellow fluorescent protein
References
Allis CD, Jenuwein T. The molecular hallmarks of epigenetic control. Nat Rev Genet. 2016;178(17):487–500.
Flavahan WA, Gaskell E, Bernstein BE. Epigenetic plasticity and the hallmarks of cancer. Science. 2017;357.
Shema E, Bernstein BE, Buenrostro JD. Single-cell and single-molecule epigenomics to uncover genome regulation at unprecedented resolution. Nat Genet. 2019;51:19–25.
Dunham I, et al. An integrated encyclopedia of DNA elements in the human genome. Nature. 2012;489:57–74.
Chalancon G, et al. Interplay between gene expression noise and regulatory network architecture. Trends Genet. 2012;28:221–32.
Eling, N., Morgan, M. D. & Marioni, J. C. Challenges in measuring and understanding biological noise. Nat Rev Genet 2019 209 20, 536–548 (2019).
Raj A, van den Bogaard P, Rifkin SA, van Oudenaarden A, Tyagi S. Imaging individual mRNA molecules using multiple singly labeled probes. Nat Methods. 2008;5:877–9.
Elowitz MB, Levine AJ, Siggia ED, Swain PS. Stochastic gene expression in a single cell. Science (80-). 2002;297:1183–6.
Sanchez, A., Choubey, S. & Kondev, J. Regulation of Noise in Gene Expression. 2013;42:469–91.
Kar G, et al. Flipping between Polycomb repressed and active transcriptional states introduces noise in gene expression. Nat Commun. 2017;8:1–14.
Rodriguez J, Larson DR. Transcription in Living Cells: Molecular Mechanisms of Bursting. Annu Rev Biochem. 2020;89:189–212.
Tunnacliffe E, Chubb JR. What is a transcriptional burst? Trends Genet. 2020;36:288–97.
Shaffer SM, et al. Memory sequencing reveals heritable single-cell gene expression programs associated with distinct cellular behaviors. Cell. 2020;182:947–959.e17.
Raj A, Rifkin SA, Andersen E, Van Oudenaarden A. Variability in gene expression underlies incomplete penetrance. Nature. 2010;463:913–8.
Wernet MF, et al. Stochastic spineless expression creates the retinal mosaic for colour vision. Nature. 2006;440:174–80.
Hinohara K, et al. KDM5 histone demethylase activity links cellular transcriptomic heterogeneity to therapeutic resistance. Cancer Cell. 2018;34:939–953.e9.
Corrigan AM, Tunnacliffe E, Cannon D, Chubb JR. A continuum model of transcriptional bursting. Elife. 2016;5.
Rybakova KN, et al. Multiplex eukaryotic transcription (In)activation: timing, bursting and cycling of a ratchet clock mechanism. PLoS Comput Biol. 2015;11.
Tantale K, et al. A single-molecule view of transcription reveals convoys of RNA polymerases and multi-scale bursting. Nat Commun. 2016;7.
Rodriguez J, et al. Intrinsic dynamics of a human gene reveal the basis of expression heterogeneity MS2 labeling of endogenous loci transcription site imaging Integrated model of gene regulation Heterogeneity. Cell. 2019;176:213–226.e18.
Senecal A, et al. Transcription factors modulate c-Fos transcriptional bursts. Cell Rep. 2014;8:75.
Harper CV, et al. Dynamic analysis of stochastic transcription cycles. PLoS Biol. 2011;9:e1000607.
Cesbron F, Oehler M, Ha N, Sancar G, Brunner M. Transcriptional refractoriness is dependent on core promoter architecture. Nat Commun. 2015;6:1–11.
Suter DM, Molina N, Naef F, Schibler U. Origins and consequences of transcriptional discontinuity. Curr Opin Cell Biol. 2011;23:657–62.
Suter DM, et al. Mammalian genes are transcribed with widely different bursting kinetics. Science (80-. ). 2011;332:472–4.
Rodriguez J, et al. Intrinsic dynamics of a human gene reveal the basis of expression heterogeneity. Cell. 2019;176:213–226.e18.
Ferguson ML, Larson DR. Measuring transcription dynamics in living cells using fluctuation analysis. Methods Mol. Biol. 2013;1042:47–60.
Pichon X, Lagha M, Mueller F, Bertrand E. A growing toolbox to image gene expression in single cells: sensitive approaches for demanding challenges. Molecular Cell. 2018;71:468–80.
Heard E, Martienssen RA. Transgenerational epigenetic inheritance: myths and mechanisms. Cell. 2014;157:95–109.
Messerschmidt DM, Knowles BB, Solter D. DNA methylation dynamics during epigenetic reprogramming in the germline and preimplantation embryos. Genes and Development. 2014;28:812–28.
Pacis A, et al. Bacterial infection remodels the DNA methylation landscape of human dendritic cells. Genome Res. 2015;25:1801–11.
Guo JU, et al. Neuronal activity modifies the DNA methylation landscape in the adult brain. Nat. Neurosci. 2011;14:1345–51.
Miller CA, Sweatt JD. Covalent Modification of DNA Regulates Memory Formation. Neuron. 2007;53:857–69.
Pacis A, et al. Gene activation precedes DNA demethylation in response to infection in human dendritic cells. Proc. Natl. Acad. Sci. U. S. A. 2019;116:6938–43.
Héberlé É, Bardet AF. Sensitivity of transcription factors to DNA methylation. Essays in Biochemistry. 2019;63:727–41.
Zhu H, Wang G, Qian J. Transcription factors as readers and effectors of DNA methylation. Nat. Rev. Genet. 2016;17:551–65.
Velasco G, et al. Dnmt3b recruitment through E2F6 transcriptional repressor mediates germ-line gene silencing in murine somatic tissues. Proc Natl Acad Sci U. S. A. 2010;107:9281–6.
Di Croce L, et al. Methyltransferase recruitment and DNA hypermethylation of target promoters by an oncogenic transcription factor. Science (80-). 2002;295:1079–82.
Li J, et al. SETD2: An epigenetic modifier with tumor suppressor functionality. Oncotarget. 2016;7:50719–34.
Yin Y, et al. Impact of cytosine methylation on DNA binding specificities of human transcription factors. Science (80-). 2017;356.
Luo C, et al. Global DNA methylation remodeling during direct reprogramming of fibroblasts to neurons. Elife. 2019;8.
Stadler MB, et al. DNA-binding factors shape the mouse methylome at distal regulatory regions. Nature. 2011;480:490–5.
Klose R, Bird A. MeCP2 Repression Goes Nonglobal. Science (80-). 2003;302:793–5.
Marano D, et al. ATRX Contributes to MeCP2-Mediated Pericentric Heterochromatin Organization during Neural Differentiation. Int J Mol Sci. 2019;20:5371.
Chahrour M, et al. MeCP2, a key contributor to neurological disease, activates and represses transcription. Science (80-). 2008;320:1224–9.
Brink MC, et al. A Role for MeCP2 in switching gene activity via chromatin unfolding and HP1γ displacement. PLoS One. 2013;8.
Tate PH, Bird AP. Effects of DNA methylation on DNA-binding proteins and gene expression. Curr Opin Genet Dev. 1993;3:226–31.
Tazi J, Bird A. Alternative chromatin structure at CpG islands. Cell. 1990;60:909–20.
Anastasiadi D, Esteve-Codina A, Piferrer F. Consistent inverse correlation between DNA methylation of the first intron and gene expression across tissues and species. Epigenetics and Chromatin. 2018;11:37.
Greenberg MVC, Bourc’his, D. The diverse roles of DNA methylation in mammalian development and disease. Nat Rev Molecular Cell Biol. 2019;20:590–607.
Huh I, Zeng J, Park T, Yi SV. DNA methylation and transcriptional noise. Epigenetics and Chromatin. 2013;6:9.
Horvath R, Laenen B, Takuno S, Slotte T. Single-cell expression noise and gene-body methylation in Arabidopsis thaliana. Heredity (Edinb). 2019;123:81–91.
Li Y, et al. DNA methylation regulates transcriptional homeostasis of algal endosymbiosis in the coral model Aiptasia. Sci Adv. 2018;4:eaat2142.
Ecker S, Pancaldi V, Valencia A, Beck S, Paul DS. Epigenetic and transcriptional variability shape phenotypic plasticity. BioEssays. 2018;40:1700148.
Coleman-Derr D, Zilberman D. Deposition of histone variant H2A.Z within gene bodies regulates responsive genes. PLoS Genet. 2012;8.
Singer ZS, et al. Dynamic heterogeneity and DNA methylation in embryonic stem cells. Mol. Cell. 2014;55:319–31.
Talbert PB, Henikoff S. Histone variants on the move: substrates for chromatin dynamics. Nat Rev Molecular Cell Biol. 2017;18:115–26.
Pardal AJ, Fernandes-Duarte F, Bowman AJ. The histone chaperoning pathway: From ribosome to nucleosome. Essays in Biochemistry. 2019;63:29–43.
Tachiwana H, et al. Structures of human nucleosomes containing major histone H3 variants. Acta Crystallogr Sect D Biol Crystallogr. 2011;67:578–83.
Fritzsch C, et al. Estrogen-dependent control and cell-to-cell variability of transcriptional bursting. Mol Syst Biol. 2018;14:e7678.
Ng., R. K. & Gurdon, J. B. Epigenetic memory of an active gene state depends on histone H3.3 incorporation into chromatin in the absence of transcription. Nat Cell Biol. 2008;10:102–9.
Zhang H, et al. RPA interacts with HIRA and regulates H3.3 deposition at gene regulatory elements in mammalian cells. Mol Cell. 2017;65:272–84.
Drané P, Ouararhni K, Depaux A, Shuaib M, Hamiche A. The death-associated protein DAXX is a novel histone chaperone involved in the replication-independent deposition of H3.3. Genes Dev. 2010;24:1253–65.
Hollenbach AD, McPherson CJ, Mientjes EJ, Iyengar R, Grosveld G. Daxx and histone deacetylase II associate with chromatin through an interaction with core histones and the chromatin-associated protein Dek. J Cell Sci. 2002;115:3319–30.
Martire S, Banaszynski LA. The roles of histone variants in fine-tuning chromatin organization and function. Nat Rev Molecular Cell Biol. 2020;21:522–41.
Hu G, et al. H2A.Z facilitates access of active and repressive complexes to chromatin in embryonic stem cell self-renewal and differentiation. Cell Stem Cell. 2013;12:180–92.
Richard M, Yvert G. How does evolution tune biological noise? Front. Genet. 2014;5:374.
Giaimo BD, Ferrante F, Herchenröther A, Hake SB, Borggrefe T. The histone variant H2A.Z in gene regulation. Epigenetics and Chromatin. 2019;12:1–22.
Lavigne, M. D., Agelopoulos, M. & Thanos, D. Composite macroH2A/NRF-1 nucleosomes suppress noise and generate robustness in gene expression accession numbers GSE53133 GSE53103 Lavigne et al. CellReports 11, 1090–1101 (2015).
Dey SS, Foley JE, Limsirichai P, Schaffer DV, Arkin AP. Orthogonal control of expression mean and variance by epigenetic features at different genomic loci. Mol Syst Biol. 2015;11:806.
Kaimori JY, et al. Histone H4 lysine 20 acetylation is associated with gene repression in human cells. Sci Rep. 2016;6:1–10.
Viñuelas J, et al. Quantifying the contribution of chromatin dynamics to stochastic gene expression reveals long, locus-dependent periods between transcriptional bursts. BMC Biol. 2013;11:15.
Wong VC, et al. NF-κB-chromatin interactions drive diverse phenotypes by modulating transcriptional noise. Cell Rep. 2018;22:585–99.
Nicolas D, Zoller B, Suter DM, Naef F. Modulation of transcriptional burst frequency by histone acetylation. Proc. Natl. Acad. Sci U. S. A. 2018;115:7153–8.
Chen LF, et al. Enhancer histone acetylation modulates transcriptional bursting dynamics of neuronal activity-inducible genes. Cell Rep. 2019;26:1174–1188.e5.
Ng HH, Robert F, Young RA, Struhl K. Targeted recruitment of Set1 histone methylase by elongating Pol II provides a localized mark and memory of recent transcriptional activity. Mol Cell. 2003;11:709–19.
Muramoto T, Müller I, Thomas G, Melvin A, Chubb JR. Methylation of H3K4 Is required for inheritance of active transcriptional states. Curr Biol. 2010;20:397–406.
Luebben WR, Sharma N, Nyborg JK. Nucleosome eviction and activated transcription require p300 acetylation of histone H3 lysine 14. Proc Natl Acad Sci U. S. A. 2010;107:19254–9.
Di Cerbo V, et al. Acetylation of histone H3 at lysine 64 regulates nucleosome dynamics and facilitates transcription. Elife. 2014;2014.
Tropberger P, et al. Regulation of transcription through acetylation of H3K122 on the lateral surface of the histone octamer. Cell. 2013;152:859–72.
Gates LA, et al. Acetylation on histone H3 lysine 9 mediates a switch from transcription initiation to elongation. J Biol Chem. 2017;292:14456–72.
Gao Y, et al. Acetylation of histone H3K27 signals the transcriptional elongation for estrogen receptor alpha. Commun Biol. 2020;3:1–10.
Wu S, et al. Independent regulation of gene expression level and noise by histone modifications. PLoS Comput Biol. 2017;13:e1005585.
Bancaud A, et al. Molecular crowding affects diffusion and binding of nuclear proteins in heterochromatin and reveals the fractal organization of chromatin. EMBO J. 2009;28:3785–98.
Gibcus JH, Dekker J. The hierarchy of the 3D genome. Mol Cell. 2013;49:773–82.
Lichter P, Cremer T, Borden J, Manuelidis L, Ward DC. Delineation of individual human chromosomes in metaphase and interphase cells by in situ suppression hybridization using recombinant DNA libraries. Hum Genet. 1988;80.
Chubb JR, Boyle S, Perry P, Bickmore WA. Chromatin motion is constrained by association with nuclear compartments in human cells. Curr Biol. 2002;12:439–45.
Chuang CH, Belmont AS. Moving chromatin within the interphase nucleus-controlled transitions? Semin Cell Dev Biol. 2007;18:698–706.
van Steensel B, Belmont AS. Lamina-associated domains: links with chromosome architecture, heterochromatin, and gene repression. Cell. 2017;169:780–91.
Finlan LE, et al. Recruitment to the nuclear periphery can alter expression of genes in human cells. PLoS Genet. 2008;4:e1000039.
Szabo Q, Bantignies F, Cavalli G. Principles of genome folding into topologically associating domains. Science. Advances. 2019;5 eaaw1668.
Dixon JR, et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature. 2012;485:376–80.
Lupiáñez DG, et al. Disruptions of topological chromatin domains cause pathogenic rewiring of gene-enhancer interactions. Cell. 2015;161:1012–25.
Ghavi-Helm Y, et al. Highly rearranged chromosomes reveal uncoupling between genome topology and gene expression. Nat Genet. 2019;51:1272–82.
Bonev B, et al. Multiscale 3D Genome Rewiring during Mouse Neural Development. Cell. 2017;171:557–572.e24.
Krefting J, Andrade-Navarro MA, Ibn-Salem J. Evolutionary stability of topologically associating domains is associated with conserved gene regulation. BMC Biol. 2018;16:87.
Almassalha LM, et al. The Global Relationship between Chromatin Physical Topology, Fractal Structure, and Gene Expression. Sci Rep. 2017;7:41061.
Bulger M, Groudine M. Functional and mechanistic diversity of distal transcription enhancers. Cell. 2011;144:327–39.
Sartorelli V, Lauberth SM. Enhancer RNAs are an important regulatory layer of the epigenome. Nat Struct Mol Biol. 2020;27:521–8.
Krivega I, Dean A. Enhancer and promoter interactions-long distance calls. Curr Opin Genet Dev. 2012;22:79–85.
Javierre BM, et al. Lineage-specific genome architecture links enhancers and non-coding disease variants to target gene promoters. Cell. 2016;167:1369–1384.e19.
Arzate-Mejía RG, Recillas-Targa F, Corces VG. Developing in 3D: the role of CTCF in cell differentiation, vol. 145. Development (Cambridge, England); 2018.
Ren G, et al. CTCF-mediated enhancer-promoter interaction is a critical regulator of cell-to-cell variation of gene expression. Mol Cell. 2017;67:1049–1058.e6.
Bartman CR, Hsu SC, Hsiung CCS, Raj A, Blobel GA. Enhancer regulation of transcriptional bursting parameters revealed by forced chromatin looping. Mol Cell. 2016;62:237–47.
Fukaya T, Lim B, Correspondence ML, Levine M. Enhancer control of transcriptional bursting article enhancer control of transcriptional bursting. Cell. 2016;166:358–68.
Larsson AJM, et al. Genomic encoding of transcriptional burst kinetics. Nature. 2019;565:251–4.
Bartman CR, et al. Transcriptional burst initiation and polymerase pause release are key control points of transcriptional regulation. Mol Cell. 2019;73:519–32.
Lee K, Hsiung CCS, Huang P, Raj A, Blobel GA. Dynamic enhancer–gene body contacts during transcription elongation. Genes Dev. 2015;29:1992–7.
Schoenfelder S, Fraser P. Long-range enhancer–promoter contacts in gene expression control. Nat Rev Genetics. 2019;20:437–55.
Nicolas D, Zoller B, Suter DM, Naef F. Modulation of transcriptional burst frequency by histone acetylation. Proc Natl Acad Sci. 2018;115:7153–8.
lien Bancaud, A., et al. Molecular crowding affects diffusion and binding of nuclear proteins in heterochromatin and reveals the fractal organization of chromatin. EMBO J. 2009;28:3785–98.
Larsson AJMM, et al. Genomic encoding of transcriptional burst kinetics. Nature. 2019;565:251–4.
Acknowledgements
We acknowledge all group members of the Verschure group especially Diewertje Piebes, Anchal Nigam and Misa Koncz for their helpful suggestions. We thank the EpiPredict consortium and INC COST Action CA18127 for interesting discussions.
Funding
This work was supported by the EpiPredict programme coordinated by PJV which received funding from the European Union’s Horizon 2020 research and innovation programme under Marie Sklodowska-Curie grant agreement No. 642691. MALJ received funding from COFUND Bio4Med, from the European Union’s Horizon 2020 research and innovation programme under Marie Sklodowska-Curie grant agreement No. 665735.
Author information
Authors and Affiliations
Contributions
WFB and PJV conceived the study. WFB performed the literature screen and analysis and drafted the manuscript. MALJ created the figures and tables and final editing of the manuscript. PJV supervised the project and helped draft the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Beckman, W.F., Jiménez, M.Á.L. & Verschure, P.J. Transcription bursting and epigenetic plasticity: an updated view. Epigenetics Commun. 1, 6 (2021). https://doi.org/10.1186/s43682-021-00007-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43682-021-00007-1