
Abstract
Background
The bark of Casuarina equisetifolia contains several active phytoconstituents that are suitable for the biosynthesis of gold nanoparticles (Au-NPs). These nanoparticles were subsequently evaluated for their effectiveness in reducing the toxicity induced by Chlorpyrifos (CPF) in rats.
Results
Various hematological and biochemical measurements were conducted in this study. In addition, markers of oxidative stress and inflammatory reactions quantified in liver and brain tissues were evaluated. Histopathological examinations were performed on both liver and brain tissues. Furthermore, the native electrophoretic protein and isoenzyme patterns were analyzed, and the relative expression levels of apoptotic genes in these tissues were determined. The hematological and biochemical parameters were found to be severely altered in the group injected with CPF. However, the administration of Au-C. equisetifolia nano-extract normalized these levels in all treated groups. The antioxidant system markers showed a significant decrease (P ≤ 0.05) in conjunction with elevated levels of inflammatory and fibrotic markers in both liver and brain tissues of the CPF-injected group. In comparison, the pre-treated group exhibited a reduction in these markers when treated with the nano-extract, as opposed to the CPF-injected group. Additionally, the nano-extract mitigated the severity of histopathological lesions induced by CPF in both liver and brain tissues, with a higher ameliorative effect observed in the pre-treated group. Electrophoretic assays conducted on liver and brain tissues revealed that the nano-extract prevented the qualitative changes induced by CPF in the pre-treated group. Furthermore, the molecular assay demonstrated a significant increase in the relative expression of apoptotic genes in the CPF-injected rats. Although the nano-extract ameliorated the relative expression of these genes compared to the CPF-injected group, it was unable to restore their values to normal levels.
Conclusion
Our results demonstrated that the nano-extract effectively reduced the toxicity induced by CPF in rats at hematological, biochemical, histopathological, physiological, and molecular levels, in the group pre-treated with the nano-extract.
Similar content being viewed by others
Background
Several hundred pesticides have been synthesized to be used widely for reducing unwanted (harmful) insects and disease carriers to increase food production [1]. It was reported that Europe is thought to be the region that uses pesticides the most, followed by China then the USA [2]. Misuse of the pesticides (uncensored) leads to serious contamination that is harmful to human health due to their affinity to accumulate in the environment [3]. Consumption of vegetables and fruits grown in pesticide-contaminated soil affected the body organs and causes chronic diseases such as neurotoxicity, cardiac disease, diabetes, cancer, and asthma in addition to reproductive disorder [4]. The quaternary nitrogen compounds are associated with neurodegenerative diseases. Additionally, the cancer problem is caused by these compounds, but breast cancer is the most common among all cancer types and is associated with organophosphorus (OP) compounds that affect cellular growth and proliferation [5]. The OP compounds are part of a vast family of pesticides that have become widespread pollutants in the environment [6]. These compounds exert their deleterious biological effects through stimulating the oxidative stress and electrophilic attack of cellular constituents by overproduction of reactive oxygen species (ROS), which results in loss of cellular homeostasis due to membrane damage. They cause lipid peroxidation products to accumulate in several targeting tissues [7, 8]. The subchronic and chronic exposure to OP pesticides causes different histological alterations in the targeting tissues including liver, kidney, brain, lung, and testes [9].
Chlorpyrifos (O,O-diethyl-O-(3,5,6-trichloro-2-pyridyl) phosphorothionate, CPF) is a commonly used OP compound utilized under different registered trademarks around the world [10]. It caused a number of cellular alterations, including decreased synaptogenesis, problems with cell differentiation and genetic transcription, changes in DNA synthesis, and breakdown of the aqueous layer, which exacerbated DNA degradation [11].
Acetylcholinesterase (AChE) exhibits its biological activity in the nervous system by regulating the neuronal communication through hydrolyzing acetylcholine (the omnipresent neurotransmitter) in the synaptic cleft [12]. It is well known that OP pesticides impede AChE action, causing acetylcholine to accumulate in the synaptic cleft, where it then remains charged and prevents further nerve impulse transmission via the cleft and hence prevents the smooth transmission of nerve functions [13]. As a result, AChE activity is regarded as a biomarker of toxicity caused by OP compounds [14].
The public and medical experts are increasingly accepting herbal treatment [15]. Many previous studies were designed to appraise the efficacy of the plant extracts against the OP pesticide–induced oxidative injuries. It was documented that root of green tea [16], black tea [17], curcumin [18], Paronychia argentea [19], Ginkgo biloba extracts, and different plant oils [20] are rich in various active biomolecules exhibiting effectiveness in reducing the peroxidation reaction and rebuilding the antioxidant system after being exposed to OP pesticides, which caused oxidative stress [21].
Casuarina equisetifolia bark contains a variety of active phytoconstituents that contribute to the pharmacological actions. These constituents include carbohydrates, alkaloids, proteins, glycosides, saponins, phenolics, flavonoids, steroids, reducing sugars, and triterpenoids [22, 23] in addition to the condensed tannins, which mostly consisted of procyanidin mixed with prodelphinidin and propelargonidin and had polymer chain lengths ranging from trimers to tridecamers of epicatechin (the main extension unit) [24]. It is well known by its antihyperlipidemic [25], hepatoprotective [26], nephroprotective [27], gastroprotective [28], antidiabetic and scavenging activities [24] in addition to the antimicrobial activities [29].
Bioavailability and efficiency of the most active phytoconstituents decreased due to their large molecular weights which affect their absorption through cellular membranes. Nanotechnology is used to solve such problems, where it increases their efficiency, bioavailability, stability, and solubility; decreases their toxicities; and achieves their release to the appropriate site of action [30]. The development of nano-extracts incorporated with these particles is considered as the most promising solution to the inherent stability issue of metal nanoparticles (M-NPs). Compared to native plant extracts alone, the plant extracts combined with the M-NPs had better antioxidant activity at lower doses because the total phenolic components were increased [31, 32]. In 2022, Aboulthana et al. [33] reported that total methanolic extract of C. equisetifolia bark was the most effective (suitable) extract for biosynthesis of gold nanoparticles (Au-NPs) and it was found that the extract demonstrated more in vitro biological activity after incorporating Au-NPs than the native extract alone. Therefore, the current study was designed to explore the efficiency of C. equisetifolia bark extract incorporated with Au-NPs against the adverse effect caused by OP compounds in rats.
Methods
Administration of Au-C. equisetifolia nano-extract
Based on our previous findings [33], the in vitro phytochemical and biological measurements of total methanolic extract and its successive (polar and non-polar) fractions showed that total methanolic extract possessed the highest biological activity. Therefore, it was chosen to be used for biosynthesis of Au-NPs during preparation of Au-C. equisetifolia nano-extract. Furthermore, it was observed that value of the median lethal dose (LD50) of the nano-extract was about 9333.33 mg/kg and hence the most suitable dose used for in vivo study was about 933.33 mg/kg b.w. (1/10 of LD50). The nano-extract was administered orally by a stomach tube.
Induction of toxicity by pesticides
Prior to treatment, animals were fasted for 18 h. The CPF solution (dissolved in dimethyl sulfoxide (DMSO)) was administered orally to rats through a stomach tube. Volume of the dose depends on weight of the animals, and the dose recommended to induce toxicity was about 1/10 of LD50 of the pesticide used [6]. It was found that the LD50 of CPF was about 229 mg/kg b.w. for rats [34]. Therefore, the dose administered orally was about 22.9 mg/kg b.w.
Experimental design
Fifty mature male Wistar rats (weighing 120–150 g) were obtained and maintained in the Animal House, National Research Centre, Giza, Egypt under controlled environmental conditions (at 25 ± 2 °C) then provided with water ad libitum and standard food. They were randomly divided into five cages (ten per cage) as the following:
-
Control group: Rats were fed with normal diet and received distilled water in parallel with DMSO for 28 days.
-
Au-C. equisetifolia bark nano-extract-treated group: Rats were treated orally with Au-C. equisetifolia bark nano-extract orally for 21 days.
-
CPF-treated (toxic) group: Rats were treated orally with CPF for 28 days.
-
Toxic group pre-treated with gold nano-extract: Rats were treated with nano-extract for 21 days followed by treating with CPF for another 28 days.
-
Toxic group post-treated with gold nano-extract: Rats were treated with CPF for 28 days followed by treating with nano-extract for another 21 days.
Collection of blood samples and tissues
The last treatment dose was administered after fasting for 18 h. The animals were then anesthetized using ketamine (100 mg/ml) and subsequently sacrificed by cervical dislocation. Blood samples were withdrawn from the retro-orbital plexus into heparinized tubes for hematological measurements and to determine the activity of the AChE enzyme. Additional blood samples were allowed to clot and then centrifuged at 3000 rpm for 15 min. The separated sera were stored at − 20 °C until biochemical assays could be conducted. Liver and brain tissues were removed from the animals and washed in ice-cold saline. A few autopsied pieces of these tissues were preserved in a neutral-buffered formalin solution (10%) for histopathological investigation. The remaining portion of the tissues was homogenized in potassium phosphate buffer (pH 7.4) and then centrifuged at 3000 rpm for 10 min. The clear supernatants were stored at − 80 °C for use in biochemical assays. For molecular analysis, the last part of the tissues was rapidly frozen using liquid nitrogen.
Body weight gain and relative organ weight assays
Body weight and body weight gain assay
Body weights of all treated groups were measured using a sensitive balance throughout the treatment period for 30 consecutive days, once at the beginning of the experiment, once after 15 days of injection, and once on the day of sacrifice (i.e., after 30 days). The biological value of Au-C. equisetifolia nano-extract was assessed by determining the effect of the nano-extract on body weight gain using the formula proposed by Al-Attar [35] at the end of each experimental period.
Relative organ weight assay
After exsanguinations, all animals were euthanized. After removing liver and brain tissues, the tissues were rinsed with saline, dried, and then weighed (absolute organ weight). As suggested by Al-Attar and Al-Rethea [36], specific formula was used for calculating the relative organ weight to body weight of each animal.
Hematological measurements
The hematological parameters, including indices of red blood cells (hemoglobin (HB), red blood cells (RBCs), hematocrit (HCT), mean corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), mean corpuscular hemoglobin concentration (MCHC), red blood cell distribution width (RDW), mean platelet volume (MPV), and platelet (PLT) count), white blood cells (WBCs), and differential blood cells (lymphocytes, monocytes, and granulocytes), were quantified in heparinized blood samples using an automatic blood analyzer (ABX Micros 60 manufactured by HORIBA ABX SAS). Moreover, activity of AChE enzyme was determined in plasma and RBC specimens using the Ellman method which was documented by Ellman et al. [37] and modified by Gorun et al. [38].
Biochemical measurements
All the conventional biochemical parameters (liver [alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), and gamma-glutamyl aminotransferases (GGT)], heart enzymes [creatine kinase (CK) and lactate dehydrogenase (LDH)], kidney functions [urea, creatinine, and uric acid], protein [total protein (TP) and albumin (Alb)], and lipid profiles [total cholesterol (TC), triglycerides (TGs), and high-density lipoprotein cholesterol (HDL-c)]) were spectrophotometrically evaluated in sera specimens using the colorimetric kits (Spectrum Diagnostics Egyptian Company for Biotechnology, Cairo, Egypt). Furthermore, the formula that was suggested by Schumann and Klauke [39] used for calculating low-density lipoprotein cholesterol (LDL-c).
Biochemical assays in supernatants of tissue homogenates
Markers of the oxidative stress
The total antioxidant capacity (TAC) [40] and reduced glutathione (GSH) [41] were assayed in liver and brain tissue homogenates. Activities of superoxide dismutase (SOD) [42], catalase (CAT) [43], and glutathione peroxidase (GPx) [44] were determined as unit per gram of tissue. Concentrations of lipid peroxidation product (LPO) [45] and total protein carbonyl (TPC) content [46] were also measured.
Inflammatory and fibrotic markers
The quantitative sandwich enzyme immunoassay technique was used for quantifying levels of tumor necrosis factor-α (TNF-α) [47] and interleukin-6 (IL-6) [48] that were expressed as pg/g in supernatants of tissue homogenates. AChE activity was determined in tissue homogenates by the Ellman method. Level of the hydroxyproline that was used as a fibrotic biomarker was assessed in liver tissue homogenates using the method suggested by Reddy and Enwemeka [49] as kg/mg tissue. In brain tissue homogenates, the β-amyloid (Aβ) content was determined using ELISA kits (USCN Life Science, Inc.).
Histopathological examination
The preserved liver and brain specimens were used for preparing paraffin sections at the thickness of 5 µm then stained with hematoxylin and eosin (H&E) as proposed by Suvarna et al. [50]. A light microscope (BX43, Olympus) was used for inspecting the stained sections that were photographed by the Olympus DP27 camera provided with cellSens dimension software (Olympus). Histopathological abnormalities in the liver were assessed and scored from 0 to 3 through determining the percentage of the lesions in five microscopically examined fields per animal (n = 5). No changes were indicated by the score of 0. The mild, moderate, and severe changes were indicated by the scores of 1, 2, and 3, respectively. The percentages were calculated according to the method suggested by Fouad and Ahmed [51] for determining the grade (mild changes (< 30%), moderate changes (< 30–50%), and severe changes (> 50%)). Neuropathologic damage was graded from 0 to 4. No changes were indicated by the grade of 0 while the grades of 1, 2, 3, and 4 indicated the percentage of affected area as follows: < 10%, 20–30%, 40–60%, and > 60%, respectively [52].
Electrophoretic assays
Native electrophoretic patterns
Known weights (0.2 g) of the tissue (liver and brain) were homogenized in extraction buffer (1 ml) then centrifuged. From each group, equal amounts of the individual (clear) supernatants were pooled together and used as a single sample and the total protein concentration was quantified in all mixed samples using the method documented by Bradford [53]. During the electrophoretic assays, all the samples were diluted with loading dye for making the protein concentrations equal in all wells.
The polyacrylamide gel electrophoresis (PAGE) was used for separating the native proteins that were stained with Coomassie brilliant blue (CBB) [54] for assaying the protein bands that appeared as blue bands, stained with Sudan Black B (SBB) [55] for assaying the lipid moiety of native protein that appeared as black bands, and stained with Alizarin Red S [56] for assaying the calcium moiety of native protein that appeared as yellow bands.
The native gel was incubated with hydrogen peroxide (H2O2) that was used as a substrate then stained with potassium iodide (KI) [57] for detecting the CAT types electrophoretically that appeared as yellow bands and stained with benzidine [58] for detecting the peroxidase (POX) types that appeared as brown bands. The electrophoretic α-amylase (α-Amy) pattern was processed by incubating the native gel with a substrate consisting of soluble starch solution then stained with iodine solution [59] for detecting the α-Amy types that appeared as yellow bands. The electrophoretic esterase (EST) patterns were assayed by incubating the native gel in conditioning buffer for optimizing the enzyme activity then stained with a reaction mixture containing Fast Blue RR (as a dye coupler) along with substrates consisting of an α- and β-naphthyl acetate solution for detecting the α- and β-EST types that appeared as brown and pink bands, respectively [60].
Data analysis
After photographing the PAGE plates, the Quantity One software (version 4.6.2) was utilized for analyzing the relative mobility (Rf), band intensity (Int.), and band percent (B%) of the electrophoretically separated bands. The equation proposed by Nei and Li [61] was used for calculating percentages of the similarity index (SI%).
Molecular assay
The relative expression of the apoptotic genes (Bcl2-associated X protein (BAX), B-cell leukemia/lymphoma (Bcl2), tumor suppressor protein 53 (p53), and caspase-3) was assayed in both liver and brain tissues. The total RNA was isolated by homogenizing the tissue samples in TRIzol® Reagent (Invitrogen, USA). Integrity of the isolated RNA was assured using ethidium bromide stain analysis of 28S and 18S bands. The DNA residues were digested by treating total RNA with RQ1 RNase-free DNase (1 U) then re-suspended in DEPC-treated water. The RevertAid™ First Strand cDNA Synthesis Kit (MBI Fermentas, Germany) used during the reverse transcription process for converting complete poly(A)+ RNA (isolated from tissue samples) into cDNA. StepOne™ Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA, USA) was used for determining the number of the copies in the tissue samples. The reaction program was allocated to three steps. The first step was at 95.0 °C for 3 min. The second step consisted of 40 cycles in which each cycle was divided into three steps: (a) at 95.0 °C for 15 s, (b) at 55.0 °C for 30 s, and (c) at 72.0 °C for 30 s. The third step consisted of 71 cycles which started at 60.0 °C and then increased at about 0.5 °C every 10 s up to 95.0 °C. The primers with the sequences suitable to the targeted genes are compiled in Supplementary Table 1. At the end of each qPCR, quality of the specific primers was checked at 95.0 °C using a melting curve analysis. Levels of the targeted genes were quantified in triplicate using the 2−ΔΔCT method as described by Wang et al. [62].
Statistical analysis
Data were statistically evaluated using one-way analysis of variance (ANOVA) and are presented in Tables 1 and 2 and Figs. 1 and 2 as mean ± standard error (SE). The difference was considered significant when the P value was less than 0.05.

Effect of C. equisetifolia extract incorporated with gold nanoparticles (Au-NPs) against toxicity induced by chlorpyrifos (CPF) on the activity of acetylcholinesterase (AChE) in the blood of rats. Data were calculated from five replicates and expressed as mean ± SE, a significant versus control group and b significant versus toxic (CPF) group at P ≤ 0.05

Effect of C. equisetifolia extract incorporated with gold nanoparticles (Au-NPs) against toxicity induced by chlorpyrifos (CPF) on a) lipid peroxidation product and b) total protein carbonyl content in liver and brain tissues of rats. Data were calculated from five replicates and expressed as mean ± SE, a significant versus control group and b significant versus toxic (CPF) group at P ≤ 0.05
Results
There were various adverse effects occurred at the hematological, biochemical, and molecular levels following CPF injection.
Body weight gain and relative organ weight assays
During the experimentation period, toxic symptoms were noticed in any of the animals. There were no changes in the nature of the treated rats’ stool, urine, and eye color. Prior to the beginning of the experiment (on day 0), the animals were weighed, and then body weight gains were calculated after 15 days and at the end of the experiment (after 30 days) of recovery period. As illustrated in Supplementary Fig. 1a, CPF injection decreased body weight gain significantly (P ≤ 0.05) throughout the duration of the study. Compared to the CPF-injected group, the body weight gain increased in the nano-extract pre- and post-treated groups significantly (P ≤ 0.05) but it could not be returned to the normal weights as control rats. Although the Au-C. equisetifolia nano-extract alone decreased the body weight, it increased gain of the body weight when administered in combination with CPF especially in the pre-treated group. As shown in Supplementary Fig. 1b, the absolute and relative weights of liver and brain tissues increased in the CPF-injected group significantly (P ≤ 0.05) compared to the control group. The nano-extract reduced the relative weights of these tissues significantly (P ≤ 0.05) in the nano-extract pre- and post-treated groups compared to the CPF-injected group. Compared to the CPF-injected group, administration of nano-extract decreased the relative weights of liver and brain tissues significantly (P ≤ 0.05) and it normalized their values to the nearest relative weights in control rats at the end of the experiment.
Hematological measurements
During the present study, it was found that the hematological and biochemical measurements are considered as the preliminary mirror to the toxicity induced by these OP compounds. As shown in Supplementary Table 2, the most hematological parameters including indices of red blood cells (RBCs, HB, HCT, MCV, RDW, MPV, and PLT) and WBCs and their differential parameters decreased in the CPF-intoxicated group significantly (P ≤ 0.05) compared to the control group. No significant changes were noticed in the MCH and MCHC levels. Compared to the CPF-injected group, the gold nano-extract normalized the levels of these measurements in all treated groups.
Biochemical measurements
As depicted in Supplementary Table 3, it was shown that activities of liver enzymes (ALT, AST, ALP, and GGT) elevated significantly (P ≤ 0.05) in sera of CPF-injected group compared to those of control group. In regard to renal functions, CPF injection caused significant (P ≤ 0.05) elevation in levels of urea, creatinine, and uric acid with lowering levels of T. protein and albumin. Moreover, CPF affected heart functions by increasing activities of heart enzymes (CK and LDH) significantly (P ≤ 0.05) with elevating levels of lipid measurements (TC, TGs, and LDL-c) compared to the control group. The Au-C. equisetifolia nano-extract restored levels of the biochemical parameters to normalcy in all nano-extract-treated groups. As illustrated in Fig. 1, activity of AChE enzyme decreased in plasma and RBCs significantly (P ≤ 0.05) in the CPF-intoxicated group compared to the control group. Compared to the CPF-injected group, the nano-extract restored the AChE activity to normal value in both the pre- and post-treated groups.
Markers of oxidative stress
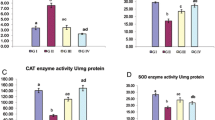
It was noticed that levels of non-enzymatic (TAC and GSH) and enzymatic (SOD, CAT, and GPx) antioxidants decreased as depicted in Table 1 with increasing concentrations of LPO and TPC content significantly (P ≤ 0.05) in both liver and brain tissues of CPF-injected group compared to those of the control group (Fig. 2). Administration of Au-C. equisetifolia nano-extract enhanced the measurements of the antioxidant system significantly (P ≤ 0.05) with declining concentrations of LPO and TPC content in both the pre- and post-treated groups compared to the CPF-injected group. It normalized levels of these measurements in the nano-extract pre-treated group.
Inflammatory and fibrotic markers
Markers of the inflammatory (TNF-α and IL-6) and fibrotic (hydroxyproline) reactions in addition to AChE activity and level of Aβ are considered as indicators for integrity of liver and brain tissues. Data presented in Table 2 showed that levels of these markers increased in liver tissues of CPF-injected group significantly (P ≤ 0.05) with reducing AChE activity. The Au-C. equisetifolia nano-extract lowered the levels of these biomarkers in both the pre- and post-treated groups significantly (P ≤ 0.05) compared to the CPF-injected group, but it could not restore their levels to normalcy. In regard to AChE activity, Au-C. equisetifolia nano-extract caused no ameliorative effect in both the pre- and post-treated groups compared to the CPF-injected group. In brain tissue, the intoxication with CPF increased levels of inflammatory markers (TNF-α and IL-6) and Aβ with reducing AChE activity significantly (P ≤ 0.05) compared to the control group. The gold nano-extract reduced the levels of inflammatory markers and Aβ content with elevating AChE activity significantly (P ≤ 0.05) compared to the CPF-injected group. It showed a higher ameliorative effect in the pre-treated group.
Histopathological examination
In liver sections of control rats, light microscopic examination exhibited normal histological architecture of hepatic parenchyma (Fig. 3A). Likewise, no histopathological alterations were seen in the liver of rats treated with Au-C. equisetifolia nano-extract (Fig. 3B). On contrary, liver of rats intoxicated with CPF showed prominent histopathological damage characterized by infiltration of inflammatory cells and focal hepatocellular necrosis associated with hepatocellular vacuolar degeneration (Fig. 3C), proliferation of Kupffer cells, apoptosis of sporadic hepatocytes (Fig. 3D), and vesicular hepatocytic nuclei. Meanwhile, the hepatic histopathological alterations were ameliorated markedly in the liver of nano-extract pre-treated rats. In addition, the examined sections revealed proliferation of Kupffer cells (Fig. 3E). On the other hand, proliferation of Kupffer cells, necrosis of sparsely hepatocytes, and edema in the portal triad were observed in the liver of nano-extract post-treated rats (Fig. 3F). Data presented in Supplementary Table 4 summarizes scores of the histopathological lesions in livers of different experimental groups, and it was noticed that the highest deleterious effect exists in the liver of CPF-intoxicated rats. The highest ameliorative efficacy exists in the nano-extract pre-treated group.

Representative photomicrographs of H&E-stained liver sections of rats (scale bar = 50 µm). A Normal control group showing the normal histoarchitecture of hepatic tissue. B Au-C. equisetifolia nano-extract-treated group showing no histopathological alterations. C and D Toxic (CPF-intoxicated) group showing hepatocellular vacuolar degeneration (black arrow), focal hepatocellular necrosis associated with infiltration of inflammatory cells (black circle), proliferation of Kupffer cells (blue arrow), and apoptosis of sporadic hepatocytes (red arrow). E Au-C. equisetifolia nano-extract pre-treated group showing proliferation of Kupffer cells (blue arrow). F Au-C. equisetifolia nano-extract post-treated group showing edema in the portal triad (asterisk)
Concerning brain tissue, the cerebral cortex of control rats as well as nano-extract-treated rats exists with normal histological architecture with intact neurons (Fig. 4A, B). Contrariwise, the remarkable neuropathic lesions investigated in CPF-intoxicated rats were characterized by marked necrosis of neurons, neuronophagia (Fig. 4C), perivascular cuffing with glial cells, and focal cerebral necrosis associated with proliferation of glial cells (Fig. 4D). Otherwise, the cerebral cortex of rats in the nano-extract pre-treated group revealed marked amelioration with regressed histopathological lesions. The examined sections showed necrosis of neuronophagia and sporadic neurons (Fig. 4E). Furthermore, the cerebral cortex of rats in the nano-extract post-treated group showed necrosis of some neurons (Fig. 4F), neuronophagia, and mild proliferation of glial cells. Data depicted in Supplementary Table 5 summarizes scores of the histopathological lesions in brain tissue in rats of different experimental groups, and it was observed that intoxication of CPF caused a statistically significant (P ≤ 0.05) adverse effect. The highest ameliorative effect of nano-extract exists in the pre-treated group.

Representative photomicrographs of H&E-stained cerebral cortex of rats (scale bar = 50 µm). A Control and B Au-C. equisetifolia nano-extract-treated groups showing the normal histological architecture with intact neurons. C and D Toxic (CPF-intoxicated) group showing dark necrotic neurons (black arrow), neuronophagia (red arrow), and focal cerebral necrosis associated with proliferation of glial cells (black circle). E Au-C. equisetifolia nano-extract pre-treated group showing necrosis of sporadic neurons (black arrow) and neuronophagia (red arrow). F Au-C. equisetifolia nano-extract post-treated group showing necrosis of some neurons (black arrow)
Electrophoretic assays
Electrophoretic protein pattern
In the liver of control rats (Fig. 5a), the native protein pattern was electrophoretically represented by nine bands (Rf values 0.08, 0.22, 0.32, 0.42, 0.54, 0.62, 0.66, 0.80, and 0.88; Int. 150.18, 151.31, 170.80, 154.86, 187.91, 189.35, 154.95, 171.49, and 173.10; B% 9.99, 10.06, 11.36, 10.30, 12.49, 12.59, 10.30, 11.40, and 11.51, respectively). Four common bands (at Rf values 0.08, 0.54, 0.80, and 0.88) were identified. In the CPF-injected group, the physiological alterations were represented by hiding four normal protein bands with the appearance of two abnormal (characteristic) ones identified at Rf values 0.50 and 0.59 (Int. 174.82 and 158.12; B% 14.55 and 13.16, respectively). Compared to the control group, SI values (SI = 62.50%) decreased in the liver of CPF-injected group. The protein pattern was improved by the pre-treatment with nano-extract through restoring four normal ones (Rf values 0.22, 0.41, 0.62, and 0.66; Int. 149.40, 157.49, 189.30, and 160.04; B% 9.75, 10.28, 12.36, and 10.45, respectively) with hiding the abnormal (characteristic) bands. The post-treatment with nano-extract showed a slight ameliorative effect by restoring four normal bands identified at Rf values 0.23, 0.54, 0.63, and 0.67 (Int. 149.02, 198.87, 189.52, and 171.96; B% 11.11, 14.93, 14.23, and 12.91, respectively) associated with hiding the characteristic bands in addition to hiding another normal one. Therefore, the SI values increased in the nano-extract pre- (SI = 100.00%) and post-treated (SI = 94.12%) groups.

Native electrophoretic protein patterns showing an effect of C. equisetifolia extract incorporated with gold nanoparticles (Au-NPs) against the physiological alterations induced by chlorpyrifos (CPF) in a) liver and b) brain tissues of rats
In the brain of control rats (Fig. 5b), it was noticed that native protein pattern was represented by seven bands (Rf values 0.08, 0.20, 0.33, 0.56, 0.66, 0.80, and 0.88; Int. 142.86, 149.92, 140.17, 174.08, 124.96, 151.88, and 116.54; B% 14.28, 14.99, 14.01, 17.40, 12.49, 15.18, and 11.65, respectively). Five common bands (at Rf values 0.08, 0.20, 0.56, 0.80, and 0.88) were identified. No characteristic bands were detected. Injection of CPF caused physiological alterations represented by hiding two normal protein bands with existence of one abnormal band (Rf 0.45; Int. 158.24; B% 18.25). Compared to the control group, SI values (SI = 76.92%) decreased in the brain of CPF-injected group. The pre-treatment with nano-extract ameliorated the protein pattern by restoring two normal bands (Rf values 0.33 and 0.65; Int. 138.75 and 142.91; B% 13.57 and 13.98, respectively) with hiding the abnormal one. The post-treatment with gold nano-extract showed a lower ameliorative effect than the pre-treated group by restoring only one normal band (Rf 0.65; Int. 137.73; B% 12.82) without hiding the abnormal one that exists at Rf 0.44 (Int. 149.51; B% 13.92). Compared to the control group, the SI values increased in the nano-extract pre- (SI = 100.00%) and post-treated (SI = 85.71%) groups.
Electrophoretic lipid moiety of native protein pattern
In the liver of control rats (Fig. 5a), the lipid moiety of native protein pattern was represented by seven bands (Rf values 0.12, 0.27, 0.40, 0.47, 0.62, 0.76, and 0.93; Int. 195.42, 204.73, 221.62, 186.95, 188.37, 167.73, and 163.19; B% 14.72, 15.42, 16.69, 14.08, 14.18, 12.63, and 12.29, respectively). Three common bands (at Rf values 0.40, 0.62, and 0.93) were identified. In the CPF-injected group, the physiological alterations were represented by hiding four normal protein bands with existence of one abnormal (characteristic) band (Rf 0.84; Int. 173.35; B% 23.66). Compared to control, the SI values (SI = 54.55%) decreased in the liver of CPF-injected group. The pre-treatment with nano-extract improved this pattern by restoring four normal bands (Rf values 0.11, 0.26, 0.47, and 0.76; Int. 191.07, 203.21, 185.15, and 173.59; B% 14.40, 15.314, 13.95, and 13.08, respectively) with hiding the abnormal (characteristic) one. The post-treatment with gold nano-extract showed a slight ameliorative effect by restoring only three normal bands (Rf values 0.40, 0.61, and 0.92; Int. 208.56, 194.40, and 174.16; B% 18.36, 17.11, and 15.33, respectively) with hiding the characteristic one. The SI values increased in the nano-extract pre- (SI = 100.00%) and post-treated (SI = 92.31%) groups compared to the control group.
In the brain of control rats (Fig. 5b), it was observed that the lipid moiety of native protein pattern was represented by four bands (Rf values 0.26, 0.50, 0.66, and 0.86; Int. 207.38, 185.82, 162.99, and 156.48; B% 29.10, 26.07, 22.87, and 21.96, respectively). Three common bands (at Rf values 0.26, 0.50, and 0.86) were identified. The CPF caused alterations in this native pattern represented by hiding one normal band with existence of one abnormal (characteristic) band (Rf 0.75; Int. 157.04; B% 22.06). Therefore, SI values (SI = 75.00%) decreased in the liver of CPF-injected group compared to the control group. The nano-extract showed the same ameliorative effect at all therapeutic modes by hiding the abnormal band with restoring the normal band in the nano-extract pre- (Rf 0.65; Int. 167.63; B% 22.86) and post-treated (Rf 0.65; Int. 161.12; B% 22.05) groups. The nano-extract pre- and post-treated groups became physiologically similar to the control group (SI = 100.00%).
Electrophoretic calcium moiety of native protein pattern
In the liver of control rats (Fig. 5a), the calcium moiety of native protein pattern was represented by four bands (Rf values 0.18, 0.53, 0.68, and 0.85; Int. 138.68, 168.60, 138.76, and 177.67; B% 22.23, 27.03, 22.25, and 28.49, respectively). Two common bands (at Rf values 0.53 and 0.85) were identified. No characteristic bands were noticed. In the CPF-injected group, the physiological abnormalities in this native pattern were represented only by hiding two normal bands. Compared to the control, the SI values (SI = 66.67%) decreased in the liver of CPF-injected group. The gold nano-extract showed the same ameliorative effect at all therapeutic modes by restoring the normal bands in the pre- (Rf values 0.18 and 0.69; Int. 145.21 and 138.89; B% 24.11 and 23.06, respectively) and post-treated (Rf values 0.18 and 0.68; Int. 139.79 and 139.37; B% 22.57 and 22.50, respectively) groups. Therefore, the nano-extract pre- and post-treated groups became physiologically identical to the control group (SI = 100.00%).
In the brain of control rats (Fig. 5b), it was noticed that the calcium moiety of native protein pattern was represented by four bands (Rf values 0.20, 0.54, 0.73, and 0.91; Int. 157.22, 154.39, 134.61, and 133.50; B% 27.12, 26.63, 23.22, and 23.03, respectively). There at two common bands (Rf values 0.20 and 0.73). The CPF caused abnormalities in this native pattern represented by hiding two normal bands with existence of one abnormal (characteristic) band (Rf 0.36; Int. 127.28; B% 31.54). Therefore, SI values (SI = 57.14%) decreased in the CPF-injected group compared to the control group. Administration of gold nano-extract showed the same ameliorative effect by hiding the characteristic band with restoring the normal bands in the pre- (Rf values 0.73 and 0.91; Int. 149.96 and 141.15; B% 26.55 and 24.99, respectively) and post-treated (Rf values 0.73 and 0.90; Int. 146.91 and 129.15; B% 25.72 and 22.61, respectively) groups. Therefore, the nano-extract pre- and post-treated groups became physiologically qualitatively identical to the control group (SI = 100.00%).
Electrophoretic CAT pattern
As illustrated in Fig. 6a, the CAT isoenzyme pattern in the liver of control rats was represented by four types (Rf values 0.18, 0.54, 0.74, and 0.90; Int. 146.68, 199.27, 122.55, and 96.58; B% 25.96, 35.26, 21.69, and 17.09, respectively). All CAT types are considered as common bands. In the CPF-injected group, the qualitative abnormalities in the CAT isoenzyme pattern were represented by appearance of one abnormal (characteristic) band (Rf 0.32; Int. 135.44; B% 20.09) without hiding normal types. Compared to the control group, the lowest SI value (SI = 88.89%) was recorded in the CPF-injected group. The nano-extract showed the same ameliorative effect at all therapeutic modes by hiding the characteristic band. The CAT isoenzyme pattern in both nano-extract pre- and post-treated groups was completely similar to that in the control group (SI = 100.00%).

Native electrophoretic isoenzyme patterns showing an effect of C. equisetifolia extract incorporated with gold nanoparticles (Au-NPs) against the physiological alterations induced in catalase (CAT) and peroxidase (POX) by chlorpyrifos (CPF) in a) liver and b) brain tissues of rats
In the brain of control rats (Fig. 6b), it was found that the CAT isoenzyme pattern was represented by three types (Rf values 0.20, 0.56, and 0.88; Int. 134.79, 133.57, and 121.64; B% 34.56, 34.25, and 31.19, respectively). The three CAT types are considered as common bands. The CPF caused physiological abnormalities in the CAT isoenzyme pattern represented by appearance of one abnormal band (Rf 0.36; Int. 131.63; B% 26.34) without hiding one or more of normal types. Therefore, SI values (SI = 85.71%) decreased in the CPF-injected group compared to the control group. The pre-treatment with nano-extract showed a higher ameliorative effect detected qualitatively in the CAT isoenzyme pattern by hiding the abnormal band completely. Therefore, this isoenzyme pattern in the nano-extract pre-treated group became physiologically identical to that in the control group (SI = 100.00%). In the nano-extract post-treated group, the nano-extract could not prevent the qualitative alterations completely but it decreased the intensity of the abnormal band (Rf 0.37 and Int. 112.06) compared to that in the CPF-injected group (Rf 0.36 and Int. 131.63). The SI value in the nano-extract post-treated group was equal to that in the CPF-injected group (SI = 85.71%) compared to the control group.
Electrophoretic POX pattern
As shown in Fig. 6a, the POX isoenzyme pattern in the liver of control rats was represented by six types (Rf values 0.11, 0.25, 0.42, 0.55, 0.75, and 0.91; Int. 179.03, 194.01, 162.55, 181.09, 210.09, and 200.59; B% 15.88, 17.21, 14.42, 16.06, 18.64, and 17.79, respectively). Four POX types (POX2, POX4, POX5, and POX6) were regarded as common bands. The CPF caused qualitative abnormalities represented by hiding the POX1 type with existence of one abnormal (characteristic) band (Rf 0.07; Int. 183.95; B% 18.90). Compared to the control group, the CPF-injected group exists with the lowest SI value (SI = 72.73%). The nano-extract showed the same ameliorative effect at all therapeutic modes by hiding the characteristic band. In the nano-extract pre- and post-treated groups, the POX isoenzyme pattern was physiologically identical to that in the control group (SI = 100.00%).
In the brain of control rats (Fig. 6b), it was found that the POX isoenzyme pattern was represented by six types (Rf values 0.12, 0.27, 0.42, 0.53, 0.73, and 0.89; Int. 192.96, 184.91, 158.05, 195.21, 186.79, and 175.60; B% 17.65, 16.91, 14.45, 17.85, 17.08, and 16.09, respectively). The four POX types (POX2, POX4, POX5, and POX6) that were identified at Rf values 0.27, 0.53, 0.73, and 0.89 are considered as common bands. The CPF caused physiological abnormalities in the POX isoenzyme pattern represented by hiding the POX1 and POX3 types with existence of one abnormal (characteristic) band (Rf 0.06; Int. 164.58; B% 17.74). Compared to the control group, the CPF-injected group exists with the lowest SI value (SI = 72.73%). The nano-extract showed a higher ameliorative effect in the pre-treated group by hiding the abnormal band with restoring the two normal POX types (POX1 and POX3 types) identified at Rf values 0.12 and 0.42 (Int. 193.25 and 158.51; B% 18.09 and 14.83, respectively). The POX isoenzyme pattern in the nano-extract pre-treated group was physiologically identical to that in the control group (SI = 100.00%). In the post-treated group, the nano-extract showed a slight ameliorative effect by hiding the abnormal band with restoring one normal type (POX1) (Rf 0.11; Int. 187.29; B% 21.05). Therefore, the POX isoenzyme pattern in the nano-extract post-treated group was identical to that in the control group by 90.91%.
Electrophoretic α-Amy pattern
In the liver of control rats (Fig. 7a), the electrophoretic α-Amy isoenzyme pattern was represented by two types (Rf values 0.24 and 0.70; Int. 95.47 and 105.18; B% 47.58 and 52.42, respectively). The second α-Amy type (α-Amy2) identified at Rf 0.70 is considered as a common band. The qualitative abnormalities in the α-Amy isoenzyme pattern were represented in the CPF-injected group by hiding the α-Amy1 type with appearance of two abnormal bands (Rf values 0.10 and 0.45; Int. 98.19 and 122.22; B% 30.28 and 37.69, respectively). The first abnormal band (Rf 0.10) is considered as a characteristic band. Compared to the control group, the CPF-injected group exists with the lowest SI value (SI = 40.00%). The nano-extract ameliorated the α-Amy isoenzyme pattern in the pre-treated group by hiding the abnormal bands with restoring the normal α-Amy type (α-Amy1) (Rf 0.26; Int. 98.97; B% 46.59). Therefore, the α-Amy isoenzyme pattern in this group was physiologically identical to that in the control group (SI = 100.00%). In the post-treated group, the nano-extract showed a slight ameliorative effect by restoring the normal α-Amy type (α-Amy1) identified at Rf 0.25 (Int. 86.01 and B% 29.00) with hiding only one abnormal band. Therefore, the α-Amy isoenzyme pattern in this group was identical to that in the control group by 80.00%.

Native electrophoretic isoenzymes patterns showing an effect of C. equisetifolia extract incorporated with gold nanoparticles (Au-NPs) against the physiological alterations induced in α-amylase (α-Amy), α-esterase (α-EST), and β-esterase (β-EST) by chlorpyrifos (CPF) in a) liver and b) brain tissues of rats
In the brain of control rats (Fig. 7b), the electrophoretic α-Amy isoenzyme pattern was represented by two types (Rf values 0.29 and 0.69; Int. 97.80 and 101.18; B% 49.15 and 50.85, respectively). Both of the two α-Amy types are considered as common bands. Injection of the CPF caused no qualitative or quantitative abnormalities in the α-Amy isoenzyme pattern. Administration of the nano-extract alone or in combination with CPF in the pre- and post-treated groups caused no differences from the control group in the α-Amy isoenzyme pattern. Therefore, the α-Amy isoenzyme pattern in all treated groups was physiologically identical to that in the control group (SI = 100.00%).
Electrophoretic EST pattern
In the liver of control rats (Fig. 7a), the electrophoretic α-EST isoenzyme pattern was represented by three types (Rf values 0.11, 0.34, and 0.64; Int. 159.05, 113.20, and 184.45; B% 34.83, 24.79, and 40.39, respectively). The two α-EST types (α-EST1 and α-EST2) identified at Rf values 0.34 and 0.64 are considered as common bands. The CPF caused qualitative abnormalities in the α-EST isoenzyme pattern represented by hiding the first α-EST (α-EST1) type without existence of abnormal bands. The CPF-injected group exists with the lowest SI value (SI = 80.00%) compared to the control group. Administration of the nano-extract showed a higher ameliorative effect in the pre-treated group by restoring the normal α-EST (α-EST1) type (Rf 0.09; Int. 154.57; B% 37.69). Therefore, the α-EST isoenzyme pattern in the nano-extract pre-treated group was physiologically identical to that in the control group (SI = 100.00%). In the post-treated group, the nano-extract could not prevent the physiological abnormalities. Therefore, the SI value in this group was equal to that in the CPF-injected group (SI = 80.00%) when compared to the control group.
In the brain of control rats (Fig. 7b), the α-EST isoenzyme pattern was represented by two types (Rf values 0.36 and 0.74; Int. 115.95 and 111.25; B% 51.03 and 48.97, respectively). The first α-EST type (α-EST1) identified at Rf 0.36 is considered as a common band. The CPF caused qualitative abnormalities in this isoenzyme pattern represented by hiding the α-EST2 type with existing two abnormal bands (Rf values 0.16 and 0.57; Int. 95.15 and 101.04; B% 30.59 and 32.48, respectively). The second abnormal band (Rf 0.57) is considered as a characteristic (unique) band. Compared to the control group, the CPF-injected group exists with the lowest SI value (SI = 40.00%). Administration of the nano-extract showed a higher ameliorative effect in the pre-treated group by restoring the normal α-EST (α-EST2) type identified at Rf 0.74 (Int. 98.91 and B% 47.46) with hiding the abnormal band. Therefore, the α-EST isoenzyme pattern in this group was qualitatively identical to that in the control group (SI = 100.00%). Regarding the post-treated group, the nano-extract showed a slight ameliorative effect by restoring α-EST2 type identified at Rf 0.73 (Int. 97.92 and B% 31.30) without hiding the abnormal band (Rf 0.16; Int. 94.36; B% 30.16). Therefore, the α-EST isoenzyme pattern in this group was qualitatively similar to that in the control group by 80.00%.
In the liver of control rats (Fig. 7a), the electrophoretic β-EST isoenzyme pattern was represented by four types (Rf values 0.17, 0.42, 0.74, and 0.89; Int. 189.18, 229.45, 149.51, and 96.66; B% 28.46, 34.51, 22.49, and 14.54, respectively). Both of the β-EST types are considered as common bands. Injection of the CPF caused no qualitative or quantitative abnormalities in the β-EST isoenzyme pattern. Administration of the nano-extract solely or in combination with CPF in the pre- and post-treated groups caused no differences from the control group in this isoenzyme pattern. Therefore, the β-EST isoenzyme pattern in all treated groups was physiologically identical to that in the control group (SI = 100.00%).
In the brain of control rats (Fig. 7b), the β-EST isoenzyme pattern was represented by three types (Rf values 0.20, 0.45, and 0.68; Int. 131.85, 110.46, and 103.21; B% 38.16, 31.97, and 29.87, respectively). The two β-EST types (β-EST2 and β-EST3) identified at Rf values 0.45 and 0.68 are considered as common bands. In all treated groups, no characteristic bands were identified. In the CPF-injected group, the qualitative alterations were represented by hiding the first β-EST (β-EST1) type with existing one abnormal band (Rf 0.82; Int. 108.56; B% 33.93). Compared to the control group, the CPF-injected group exists with the lowest SI value (SI = 66.67%). Administration of the nano-extract showed a higher ameliorative effect in the pre-treated group by restoring the normal β-EST (β-EST1) type (Rf 0.19; Int. 123.61; B% 34.92) with hiding the abnormal band. Therefore, the β-EST isoenzyme pattern in this group was qualitatively identical to that in the control group (SI = 100.00%). Regarding the post-treated group, the nano-extract ameliorated this pattern slightly by restoring the normal β-EST (β-EST1) type identified at Rf 0.19 (Int. 125.50 and B% 26.43) without hiding the abnormal band (Rf 0.83; Int. 104.26; B% 21.96). Therefore, the β-EST isoenzyme pattern in this group was qualitatively similar to that in the control group by 85.71%.
Molecular assay
It was carried out by detecting the expression of the apoptotic genes (BAX, Bcl2, p53, and caspase-3). As shown in Fig. 8, it was noticed that the relative expression of BAX, p53, and caspase-3 genes elevated significantly (P ≤ 0.05) with decreasing expression of Bcl2 gene in both liver and brain tissues of CPF-injected group compared to that of the control group. The nano-extract at all therapeutic modes resulted in decreasing the relative expression of BAX, p53, and caspase-3 genes significantly (P ≤ 0.05), as well as increasing the relative expression of Bcl2 gene in both liver and brain tissues compared to the CPF-injected group. Although nano-extract showed a higher ameliorative effect in the pre-treated group, it could not restore the relative expression levels of these genes to normal values.

Effect of C. equisetifolia extract incorporated with gold nanoparticles (Au-NPs) against the alterations in the relative expression of apoptotic genes, a) BAX, b) Bcl2, and c) p53, and d caspase-3 genes, induced by chlorpyrifos (CPF) in liver and brain tissues of rats
Discussion
The toxicity may occur mainly due to eating the meat of animals immersed in solutions of the OP compounds which are used widely in animal husbandry and agriculture [63]. Herbal products can be used as an effective strategy for enhancing the biological processes. Therefore, there is a considerable inclination to consume these natural remedies to diminish the toxicity brought on by the exposure to these toxicants [64]. The medicinal plants have gained prominence in nanobiotechnology due to availability of secondary metabolites that used during Au-NP biosynthesis as reducing agents [65]. The present work was designed to appraise the efficiency of the phytochemically synthesized Au-NPs against the toxicity induced by CPF. Hematological, biochemical, and molecular parameters as well as the histopathological examination were assessed to support the proposed hypothesis.
Body weight gain and relative organ weight assays
Relative organ weight analysis is a crucial criterion for identifying substances that affect human health negatively [66]. It was demonstrated that the body weight gain decreased after CPF injection throughout the duration of the study and there is a decrease in the rate of the weight recovery. This was in accordance with Mossa and Abbassy [67] who postulated that the decline in body weight gains of CPF-intoxicated group might be related to decreasing consumption of the food that corresponded to lowering water and food intake compared to control rats and/or due to the direct effect of CPF on increasing degradation of lipids and proteins [68]. Also, it might refer to the ability of CPF to cause physical or reflex development [69].
The relative organ weights were used as the most sensitive indicator for revealing the deleterious effects of toxins [70]. The present study showed that the absolute and relative weights of liver and brain tissues increased significantly in rats injected with CPF, and this was supported with Pollakova et al. [71] who supposed that the relative liver weight increased in the CPF-injected group due to decreasing cell proliferation and damage of cell membrane in liver tissues. Regarding the brain tissue, the CPF caused an increase in the relative brain weight and this agreed with Akhtar et al. [72] who emphasized that the changes in brain weights might be related to reducing the body weight gain in addition to severity of neurotoxicity that occurs with histopathological changes. The Au-C. equisetifolia nano-extract alone caused a decrease in the body weight gains due to the presence of the Au-NPs that have the ability to accumulate rapidly in the abdominal fat tissue showing significant reduction in abdominal fat mass by improving lipid metabolic profile compared to control [73]. Treatment of the CPF-injected rats with nano-extract increased the body weight gain, and this agreed with Fereidounni and Dhawan [74] who suggested that the biosynthesized Au-NPs achieved a remarkable improvement in body weight due to increasing the phenolic groups in plant extract which prevented oxidative injury by increasing antioxidant action.
Hematological measurements
It is well known that CPF affected spleen and bone marrow cells by accelerating apoptosis and inducing the DNA damage [75]. The current study showed that indices of red blood cells (RBCs, HB, HCT, MCV, RDW, MPV, and PLT) declined in the CPF-intoxicated group and this was in agreement with Deabes et al. [76] who demonstrated that the hematological measurements were altered in the CPF-injected group due to elevating products of the peroxidation reactions as a consequence of stimulating the oxidative stress. Moreover, the differential WBCs decreased in the CPF-injected group compared to the control group. This was coincided with Hashem and El-Sharkawy [77] who reported that CPF caused lymphopenic leucopenia due to the oxidative damage to the leukocytes. Treatment of the CPF-injected rats with Au-C. equisetifolia nano-extract normalized levels of these measurements in all groups treated with nano-extract, and this might be attributed to the existence of various polyphenolic compounds with high antioxidant activity and high ability to scavenge the ROS generated due to the exposure to CPF [76]. The nano-extract is rich in the active phytoconstituents which are able to improve absorption of iron by facilitating conversion of the iron from the ferrous (oxidized) state to the ferric (reduced) state in addition to ameliorating the integrity of the RBCs and WBCs compromised by the oxidative stress induced by CPF [78]. Moreover, they improved platelet count due to their ability to protect the platelet from oxidative damage, thus reducing the production of lipid peroxides within platelet membranes [79].
The AChE is considered as a standard biomarker for neurotoxic effects induced by OP compounds [80]. It is a membrane-bound enzyme that catalyzes the transformation of the acetylcholine (neurotransmitter) at the myoneural junction into choline and acetate [81]. The present study showed that CPF decreased the activity of the AChE enzyme in the plasma, and this was in accordance with Wielgomas and Krechniak [82] who suggested that decreasing the activity of this enzyme might be attributed to the binding of CPF to AChE, causing its inhibition irreversible at neuromuscular junctions and cholinergic synapses due to the oxidative damage to cholinergic cells by CPF. This consequently leads to accumulating acetylcholine in the synapses resulting in cholinergic toxicity due to inducing the hyperactivity in cholinergic pathways. As a result of blocking acetylcholine release from presynaptic membranes, the accumulated acetylcholine prevents conduction of the nerve impulse eventually [83]. Treatment of CPF-injected rats with nano-extract enhanced the activity of AChE enzyme in plasma and RBCs compared to that of the CPF-injected group due to the presence of the active (phenolic) ingredients that possess antioxidant properties [84]. Recently, Bekheit et al. [85] supported that efficiency of these phenolic compounds increased after incorporating Au-NPs.
Biochemical measurements
Profiles of the blood chemistry were measured to assess the physiological and pathology state of vital organs in animals [86]. Data of our study showed a progressive increment in levels of the liver enzymes in sera of CPF-injected group, and this was supported by Heikal et al. [87], who proposed that the increase in activities of these enzymes is a result of functional damage in the liver (the final target site of CPF). Elevated levels of these hepatic enzymes could indicate dysfunction of the hepatocytes and disturbance in the biosynthesis of these enzymes, as well as changes in hepatocyte membrane permeability [88, 89]. The current study revealed that levels of urea, creatinine, and uric acid elevated in CPF-injected rats and this was in accordance with Kasteel et al. [90] who have proven that increasing these measurements might refer to degradation of both purines and pyrimidines and/or accelerating rate of protein catabolism rather than decreased urinary excretion of urea in addition to impairment of kidney function and reduction of the glomerular filtration rate that consequently lead to retention of urea in the blood. Also, concentrations of total protein and albumin decreased in CPF-injected rats due to increasing protein catabolism [91]. Intoxication of CPF increased levels of the heart enzymes (CK and LDH) and measurements of lipid profile. This agreed with Saoudi et al. [92] who documented that perturbations of the heart functions were attributable to an oxidative damage in CPF-injected rats. Moreover, the increase in levels of these measurements might be related to increasing permeability of cellular membranes [93]. The Au-C. equisetifolia nano-extract alleviated the hepatic, renal, and heart toxicity induced by CPF especially in the pre-treated group. The protective potential against liver damages might refer to the role of the phenolic compounds (the major phytoconstituents) in plant extracts that were responsible for an inhibitory effect on activities of transaminases [94]. Also, the biosynthesized Au-NPs showed a prominent decrease in activities of the liver enzymes due to stabilizing membrane of hepatocytes and preventing liberation of these enzymes [95]. Levels of the renal measurements restored to normalcy due to the presence of the bioactive components that exhibit various biological activities [96]. The presence of the Au-NPs enhanced the regenerative ability of the renal tubules (the primary site for Au-NP accumulation) and showed a pronounced effect on protein content due to its ability to normalize measurements of profile [97]. Levels of heart enzymes and lipid measurements were restored by nano-extract to normal values due to the hypolipidemic properties of the phytoconstituents which decrease levels of TC, TGs, and LDL [98]. Level of HDL increased by the nano-extract because of stimulating lecithin cholesterol acyltransferase involved in regulating serum lipids [99].
Markers of oxidative stress
The changes in activities of antioxidant enzymes (SOD, CAT, and GPx) have been reported to be an indicator of oxidative stress induced by the exposure to OP compounds [100]. The current study showed that intoxication of CPF produced a significant decrease in markers of the antioxidant system in hepatic and brain tissues, and these findings are consistent with Ibrahim and Khwanes [101] who showed that CPF decreased activities of these enzymes due to overproduction of the ROS which could be linked to hepatocyte and brain lesions, as well as insufficient detoxification capacity of the CPF. The present study emphasized that the level of LPO elevated in hepatic and brain tissues in the CPF-injected group and this agreed with Kasteel et al. [90] who suggested that the LPO level increased due to disequilibrium in the biological system between pro-oxidants and antioxidants, resulting in overproduction of the ROS that consequently leads to damage of cellular biomacromolecules or cell death. Also, level of TPC content increased in those tissues after injecting CPF and this agreed with Ojha and Gupta [102] who suggested that the TPC content level increased in liver and brain (especially cerebrum and cerebellum regions) following the exposure to CPF due to the influx of ROS, which take protons from amino acids (methylene groups) to produce carbonyls; nucleophilic centers are often damaged; and sulfhydryl groups are oxidized, leading to the loss of protein activities. Furthermore, lowering levels of enzymatic (SOD, CAT, and GPx) and non-enzymatic (TAC and GSH) antioxidants might be associated with increasing peroxidation reaction products (LPO and TPC content) [103]. The Au-C. equisetifolia nano-extract alleviated the negative effects induced by CPF in the pre-treated group by restoring measurements of the antioxidant system in liver and brain tissues due to enhancing the phenolic compounds which are responsible for the antioxidant activity in nano-extract [104]. The phycocyanin, carotenoids, vitamins, lipids, proteins, and carbohydrates belong to the major phytoconstituents that exist in the C. equisetifolia extract and exhibit antioxidant and scavenging activities as proposed by Dib et al. [105] and supported consequently by Aboulthana et al. [106]. These components decreased the levels of LPO and TPC content due to scavenging the reactive species that induce lipid peroxidation or by inhibiting xanthine oxidase, a recognized enzyme that generates free radicals [107]. In addition, the presence of Au-NPs in the nano-extract enhanced the antioxidant properties and demonstrated higher protective role against ROS [33].
Inflammatory and fibrotic markers
Cytokines (small molecular weight proteins) are considered as biomarkers for inflammation, immune reactivity, tissue injury, and repair [108]. In the current study, the pro-inflammatory cytokines (TNF-α and IL-6) elevated in liver and brain tissues of CPF-injected group significantly and this was coincided with the findings obtained by Abolaji et al. [109] who suggested that the inflammasome and subsequent innate immune response were stimulated by the mitochondrial oxidative stress mediated by CPF. Therefore, the ROS scavengers might be used to repair the OP compound–induced lesions [110]. Level of hydroxyproline (fibrotic marker) increased in the CPF-injected group, and this might refer to inducing damage to intestinal barrier and increasing the intestinal permeability [111] and/or due to translocation of gut bacteria to liver, leading to observed inflammatory reaction [112].
The CPF injection reduced the activity of AChE enzyme in both liver and brain tissues, and this was in accordance with Mohamed et al. [89] who proposed that the decrease in AChE activity is strongly related to excessive generation of the ROS that affect the enzyme activity due to a possible oxidative injury to cholinergic cells. Moreover, AChE activity might be decreased as a result of the microsomal oxidation in the liver and formation of the active metabolite CPF-oxone forms (a potent inhibitor of AChE in the peripheral tissues) that were generated after hepatic metabolism of CPF by cytochrome P450 2B6 enzyme and this metabolite possesses inhibiting potency higher than that of the parent compounds [113]. By inhibiting AChE activity, accumulation of acetylcholine is increased by CPF, stimulating its receptors more and downregulating them as a result [114]. In brains of rats, CPF caused significant inhibition in the AChE and this was in accordance with Imam et al. [115] who emphasized that AChE inhibition is categorized as the primary mechanism of the CPF action affecting redox processes and the lipid peroxidation that are greatly implicated in the chronic outcomes following the CPF exposure. Pre-administration of the nano-extract ameliorated the inflammation induced as a result of intoxication of CPF. This was in agreement with Rahmouni et al. [116] who demonstrated that the nano-extract is rich in flavonoids and other constituents with polyphenolic structures enable them to possess anti-inflammatory properties by decreasing levels of the pro-inflammatory biomarkers. The presence of the Au-NPs in the plant extract interferes with transmission of inflammatory signaling, and this is mostly attributed to the extracellular interactions with IL-6 which aggregates around Au-NPs, thus inhibiting IL-6 binding to cellular receptors [117]. The Au-NPs decreased the progression of inflammation by decreasing mRNA expression specific to TNF-α and IL-6 in addition to inducible nitric oxide synthase [118].
Histopathological examination
The biochemical and histopathological studies carried out by Almazroo et al. [119] showed that the liver and brain are the most target organs susceptible for CPF accumulation. During the current study, the microscopic examination of the liver in the CPF-injected group revealed various histopathological changes represented by severe alterations in the architecture of the liver with increasing degeneration of the hepatic cells. These changes are consistent with the biochemical measurements assessed during our investigation. This agreed with Tanvir et al. [88] who revealed that the CPF can easily cross the cell membrane due to its lipophilic properties causing damage inside the liver (an organ responsible for detoxification). The histopathological changes in the CPF-injected group might be related to the oxidative stress that causes severe lesions in the hepatocytes due to decreasing measurements of the antioxidant system in liver tissues, leading to overproduction of ROS and hence injury then death of healthy cells [106]. In regard to brain tissues, severe histopathological alterations were detected in the CPF-injected group and this agreed with Altun et al. [120] who proposed that the hemorrhages in brain tissues were caused by the acute CPF intoxication. Latuszynska et al. [121] added that the CPF caused numerous histopathological changes, including formation of eosinophilic plagues, hemorrhage in the covering meninges, and pyknosis in the cerebellum area with edema and pyknosis in the cerebral cortex accompanied with accumulation of the cytoplasm in neurocytes. The oxygen, hydroxyl, and hydroperoxyl radicals belong to ROS that were produced mostly in mitochondria during electron transport chain to form peroxynitrite (disrupting the redox status) after the interaction with NO [122]. Treatment of the CPF-injected rats with Au-C. equisetifolia nano-extract might protect the liver and brain tissues against oxidative lesions induced by CPF. It caused less histopathological changes in liver tissues than in the CPF-injected group, indicating its protective effects and stimulated restoration of the total number and mass of hepatocytes. This might be attributed to existence of the flavonoids that exhibit ability to donate electrons or hydrogen molecules [123]. These constituents are well known by their ability to act as free radical scavengers to prevent oxidative damage and to stabilize membranes [124]. Furthermore, the nano-extract decreased scores of the histopathological damage due to existence of the Au-NPs that exhibit antioxidant and the anti-inflammatory effects. In agreement with the previous findings, other investigators reported that Au-NPs reduced leukocyte migration and also macrophage infiltration [125].
Electrophoretic patterns
Electrophoresis is used for detecting the mutagenic differences that may occur by hiding normal bands and/or existing abnormal ones in the different protein and isoenzyme patterns [126]. Percent of the SI is related the physiological state of tissue, and it is inversely proportional to the qualitative alterations [127].
The present study showed that CPF caused qualitative abnormalities in the native electrophoretic protein patterns in liver and brain tissues. Changing composition of the native protein might be related to altering function and organization of the DNA and hence affect expression of different proteins [128]. Also, the disturbances in the protein pattern might refer to the role of CPF in inhibiting expression of some genes or activating the others to produce specific mRNAs which may be translated subsequently into specific proteins and/or due to altering metabolism (synthesis/degradation) of various proteins [129]. Furthermore, appearance of new protein bands in the liver and brain of CPF-injected rats might refer to changing the cytoplasmic protein due to inhibiting the protein anabolic actions and/or due to altering the functional conformations of the structural proteins, resulting in denaturation of the high molecular weight proteins [130].
Lipids possess an important impact in maintaining the integrity of the cell membrane [131]. The present study demonstrated that CPF caused physiological variations in lipid moiety of native protein pattern in liver tissue and this agreed with Kalender et al. [8] who showed that CPF affected the permeability of the hepatocyte membranes and might be attributable to the blockage of the bile duct which reduces the secretion of cholesterol into the duodenum. Also, this might refer to increasing the hepatic synthesis and/or diminishing hepatic degradation of lipids due to the reducing activity of the lipoprotein lipase [132]. In the brain of CPF-injected group, the alterations in the protein pattern might refer to its adverse effect on the cellular lipids, causing abnormal synthesis or defective degradation of lipids [133]. Moreover, the peroxidation products that accumulated due to depletion of the antioxidant defenses affect the lipid portion by inducing the formation of the ROS that leads consequently to modifications of the lipid moieties oxidatively and hence their denaturation [134].
Calcium-binding proteins are acidic proteins with low molecular weights, exhibiting a crucial role in resistance of the tissues against the deleterious effects induced by the toxic substances. They altered the mineralization process by inhibiting the formation of the hydroxyapatite [135]. The current study showed that CPF injection caused changes in calcium moiety of the native protein and this agreed with Abulyazid et al. [136] who reported that the abnormal tissue mineralization is responsible for altering this native protein pattern. El-Sayed et al. [134] demonstrated that the changes in quality and quantity of this native protein pattern might refer to overproduction of the ROS that convert an active hydrogen atom from these biologically macromolecules. Moreover, these alterations might be related to an increasing concentration of calcium which stimulates oxidative stress in the tissue, leading to variations in this protein pattern [137].
The treatment of the CPF-injected group with gold nano-extract had an ameliorative effect against the changes induced by the attack of free radical in the different electrophoretic protein patterns. This could allude to the existence of the active constituents (polyphenolic compounds) that elevated as a result of incorporation of M-NPs [138]. The nano-extract ameliorated the lipid moiety of native protein pattern due to presence of the phytoconstituents that exhibit lipolysis-stimulating agent in adipocytes. This lipolytic action was mainly mediated by phosphorylation of hormone-sensitive lipase which is regulated post-transcriptionally by reversible phosphorylation by protein kinase A [139].
Activities of the antioxidant enzymes vary from tissue to tissue. Therefore, they are considered as a tissue-dependent tool. The changes in their activities in the CPF-injected group might be related to degenerating the protein contents or altering the metabolic pathways due to the attack of the ROS targeting protein contents [140]. As shown in the current study, CPF altered the electrophoretic CAT pattern, which may be explained by the fact that when CPF binds to native macromolecules, it causes modifications in the secondary structure of the enzyme [141]. The abnormalities in the electrophoretic POX pattern might be related to the peroxidation reactions stimulated by the oxidative stress [142]. Additionally, ROS may have an impact on these enzymes directly or on the protein component of these enzymes, which would explain the decline in their activity. Hence, these ROS have the ability to alter the physico-chemical and immunologic characteristics of endogenous CAT and POX enzymes.
The α-amylase belongs to the digestive enzymes that catalyze hydrolysis of the α-(1,4)-d-glycosidic bonds of starch and other glucose polymers, which leads to the conversion of dietary carbohydrates to oligosaccharides and disaccharides [143]. The present study demonstrated that CPF altered the electrophoretic α-Amy isoenzyme pattern and this might refer to changing the fractional activity that is correlated with altering the rate of protein expression secondary to DNA damage caused oxidatively by ROS [127]. Abulyazid et al. [136] added that the oxidative stress induced structural changes in protein portion of native enzymes. The alterations in expression of the different proteins are responsible for changing the enzymatic activities. Moreover, the alterations in the α-Amy isoenzyme pattern might be related to the hormonal and metabolic alterations [144]. Esterases are a wide family of polymorphic and tissue-specific lysosomal lipolytic enzymes that are distinguished by their capacity to catalyze hydrolyzing ester linkages in neutral lipids into the corresponding carboxylic acids [145]. The brain is rich in EST enzymes that exist in membrane structures and exhibit their role in the neurotransmission and communication of messages by stimulating the breakdown of acetylcholine liberated during nervous stimulation [146]. The present study showed that the CPF injection caused mutagenicity in the electrophoretic α- and β-EST patterns due to changing fluidity of the membrane, reflecting alterations in the physical state of the membrane lipids. Therefore, these membrane-bound enzymes are sensitive to these deleterious effects [147]. Also, these variations might be directly related to excessive generation of the ROS which stimulate the fragmentation of polypeptide chain through sulfhydryl-mediated cross-linking of labile amino acids affecting the integrity of protein molecule [148]. The gold nano-extract ameliorated the electrophoretic isoenzyme patterns due to existence of the active phytoconstituents that stimulate the antioxidant status of the tissue to resist the attack of the ROS targeting these biomacromolecules [149]. Incorporation of the Au-NPs increased the polyphenolic compounds and hence enhanced the ameliorative effect of the extract in maintaining the integrity of these biomacromolecules against the adverse effect induced by oxidative reactions [85].
Molecular assay
The current study showed that the CPF injection increased levels of the relative expression levels of caspase-3, p53, and Bax genes associated with decreasing expression of the Bcl2 gene in liver tissues. This might be attributable to the role of CPF in upregulating the pro-apoptotic mediators in this tissue [89] in addition to the role of the excessive ROS in inducing cell apoptosis in the CPF-injected group [73]. In brain tissues, levels of the relative expression of the apoptotic genes increased in the CPF-injected group due to inducing transcription of the critical genes required for apoptosis [150]. The relative expression of the Bcl2 protein decreased with increasing the relative Bax expression contrarily in both cerebrum and cerebellum. The relative expression of these proteins changed due to initiation of apoptotic machinery by ROS generation and releasing the stress signal in response to CPF intoxication [151]. Treatment of CPF-injected rats with Au-C. equisetifolia nano-extract regulated the expression of the pro- and anti-apoptosis mediators and produced a synergistic effect on the expression of apoptosis-related proteins by decreasing the expression of p53, Bax, and caspase-3 with increasing the Bcl2 expression. This agreed with El Gamal et al. [152] who demonstrated that the presence of flavonoids and polyphenolic chemicals, which have potent antioxidant, radical scavenging, and anti-inflammatory properties, may be responsible for upregulation of anti-apoptotic proteins and downregulation of pro-apoptotic proteins. Also, the presence of the Au-NPs demonstrated strong antioxidant activity and was capable of simulating the activity of antioxidant enzymes by interacting with hydroxyl radicals and superoxide anions directly to produce less-reactive side products [85, 153]. Zhou et al. [154] demonstrated that Au-NPs directly bind and neutralize reactive species, and this effect depends on the size and surface of the Au-NPs.
Conclusions
Our results proved that the alterations induced by CPF were diminished by Au-C. equisetifolia nano-extract which restore levels of the hematological and biochemical parameters in all nano-extract-treated groups to normalcy. It normalized measurements of the antioxidant system and restored levels of the inflammatory and fibrotic markers in both liver and brain tissues in the nano-extract pre-treated group. Moreover, severity of the histopathological lesions induced in these tissues by the CPF was decreased by the nano-extract which showed the highest ameliorative efficiency in the pre-treated group. The different electrophoretic protein and isoenzyme patterns altered in the CPF-intoxicated group were ameliorated by the nano-extract especially in the pre-treated group.
Availability of data and materials
This paper has data included as electronic supplementary material.
Abbreviations
- AChE:
-
Acetylcholinesterase
- ALP:
-
Alkaline phosphatase
- ALT:
-
Alanine aminotransferase
- ANOVA:
-
Analysis of variance
- AST:
-
Aspartate aminotransferase
- Au-NPs:
-
Gold nanoparticles
- Aβ:
-
β-Amyloid
- B%:
-
Band percent
- BAX:
-
Bcl2-associated X protein
- Bcl2:
-
B-cell leukemia/lymphoma
- CAT:
-
Catalase
- CBB:
-
Coomassie brilliant blue
- CK:
-
Creatine kinase
- CPF:
-
Chlorpyrifos
- DMSO:
-
Dimethyl sulfoxide
- EST:
-
Esterase
- GGT:
-
Gamma-glutamyl aminotransferases
- GPx:
-
Glutathione peroxidase
- GSH:
-
Reduced glutathione
- HB:
-
Hemoglobin
- HCT:
-
Hematocrit
- HDL-c:
-
High-density lipoprotein cholesterol
- IL-6:
-
Interleukin-6
- Int.:
-
Band intensity
- LD50 :
-
Median lethal dose
- LDH:
-
Lactate dehydrogenase
- LDL-c:
-
Low-density lipoprotein cholesterol
- LPO:
-
Lipid peroxidation product
- MCH:
-
Mean corpuscular hemoglobin
- MCHC:
-
Mean corpuscular hemoglobin concentration
- MCV:
-
Mean corpuscular volume
- M-NPs:
-
Metal nanoparticles
- MPV:
-
Mean platelet volume
- OP:
-
Organophosphorus
- p53 :
-
Tumor suppressor protein 53
- PAGE:
-
Polyacrylamide gel electrophoresis
- PLT:
-
Platelet
- RBCs:
-
Red blood cells
- RDW:
-
Red blood cell distribution width
- Rf:
-
Relative mobility
- ROS:
-
Reactive oxygen species
- SBB:
-
Sudan Black B
- SE:
-
Standard error
- SI%:
-
Percentage of similarity index
- SOD:
-
Superoxide dismutase
- TAC:
-
Total antioxidant capacity
- TC:
-
Total cholesterol
- TGs:
-
Triglycerides
- TNF-α:
-
Tumor necrosis factor-α
- TPC:
-
Total protein carbonyl
- WBCs:
-
White blood cells
- α-Amy:
-
α-Amylase
References
Abhilash PC, Singh N (2009) Pesticide use and application: an Indian scenario. J Hazard Mater 165:1–12
Bernhardt ES, Rosi EJ, Gessner MO (2017) Synthetic chemicals as agents of global change. Front Ecol Environ 15:84–90
Udeigwe TK, Teboh JM, Eze PN, Hashem Stietiya M, Kumar V, Hendrix J, Kandakji T (2015) Implications of leading crop production practices on environmental quality and human health. J Environ Manage 151:267–279
Kalyabina VP, Esimbekova EN, Kopylova KV, Kratasyuk VA (2021) Pesticides: formulants, distribution pathways and effects on human health – a review. Toxicol Rep 8:1179–1192
Calaf GM (2021) Role of organophosphorous pesticides and acetylcholine in breast carcinogenesis. Semin Can Biol 76:206–217
Raj J, Chandra M, Dogra TD, Pahuja M, Raina A (2013) Determination of median lethal dose of combination of endosulfan and cypermethrin in Wistar rat. Toxicol Int 20(1):1–5
Saulsbury MD, Heyliger SO, Wang K, Johnson DJ (2009) Chlorpyrifos induces oxidative stress in oligodendrocyte progenitor cells. Toxicol 259:1–9
Kalender S, Uzun FG, Durak D, Demir F, Kalender Y (2010) Malathion-induced hepatotoxicity in rats: the effects of vitamins C and E. Food Chem Toxicol 48(2):633–638
Kopjar N, Žunec S, Mendaš G, Micek V, Kašuba V, Anja Mikolić A, Lovaković BT, Milić M, Pavičić I, Čermak AMM, Pizent A, Vrdoljak AL, Želježić D (2018) Evaluation of chlorpyrifos toxicity through a 28-day study: cholinesterase activity, oxidative stress responses, parent compound/metabolite levels, and primary DNA damage in blood and brain tissue of adult male Wistar rats. Chem Biol Interact 279:51–63
Toan PV, Sebesvari Z, Blasing M, Rosendahl I, Renaud FG (2013) Pesticide management and their residues in sediments and surface and drinking water in the Mekong Delta. Vietnam Sci Total Environ 452:28–39
Braquenier JB, Quertemont E, Tirelli E, Plumier JC (2010) Anxiety in adult female mice following perinatal exposure to chlorpyrifos. Neurotoxicol Teratol 32:234–239
Silman I, Sussman JL (2005) Acetylcholinesterase: ‘classical’ and ‘non-classical’ functions and pharmacology. Curr Opin Pharmacol 5:293–302
Kaur R, Sandhu HS (2008) In vivo changes in antioxidant system and protective role of selenium in chlorpyrifos-induced subchronic toxicity in Bubalus bubalis. Environ Toxicol Pharmacol 26:45–48
Tam NT, Berg H, Hang NTT, Cong NV (2015) Effects of chlorpyrifos ethyl on acetylcholinesterase activity in climbing perch cultured in rice fields in the Mekong Delta. Vietnam Ecotoxicol Environ Saf 117:34–40
Panda VS, Naik SR (2009) Evaluation of cardioprotective activity of Ginkgo biloba and Ocimum sanctum in rodents. Altern Med Rev 14:161–171
Mehri N, Felehgari H, Harchegani AL, Behrooj H, Kheiripour N, Ghasemi H, Mirhoseini M, Ranjbar A (2016) Hepatoprotective effect of the root extract of green tea against malathion-induced oxidative stress in rats. J HerbMed Pharmacol 5(3):116–119
Khan SM, Sobti RC, Kataria L (2005) Pesticide-induced alteration in mice hepato-oxidative status and protective effects of black tea extract. Clin Chim Acta 358(1–2):131–138
Canales-Aguirre AA, Gomez-Pinedo UA, Luquin S, Ramírez-Herrera MA, Mendoza-Magaña ML, Feria-Velasco A (2012) Curcumin protects against the oxidative damage induced by the pesticide parathion in the hippocampus of the rat brain. Nutr Neurosci 15(2):62–69
Zama D, Meraihi Z, Tebibel S, Benayssa W, Benayache F, Benayache S, Vlietinck AJ (2007) Chlorpyrifos-induced oxidative stress and tissue damage in the liver, kidney, brain and fetus in pregnant rats: the protective role of the butanolic extract of Paronychia argentea L. Indian J Pharmacol 39(3):145–150
Al-Attar AM, Elnaggar MHR, Almalki EA (2017) Protective effect of some plant oils on diazinon induced hepatorenal toxicity in male rats. Saudi J Biol Sci 24(6):1162–1171
Hajirezaee S, Rafieepour A, Shafiei S, Rahimi R (2019) Immunostimulating effects of Ginkgo biloba extract against toxicity induced by organophosphate pesticide, diazinon in rainbow trout, Oncorhynchus mykiss: innate immunity components and immune-related genes. Environ Sci Pollut Res Int 26(9):8798–8807
Kishore DV, Rumana R (2014) Spasmolytic activity of Casuarina equisetifolia bark extract. Int J Pharm Sci Res 3(5):1452–1456
Al-Snafi AE (2015) The pharmacological importance of Casuarinaequisetifolia - an overview. IJPSM 5(1):4–9
Gumgumjee NM, Hajar AS (2012) Antimicrobial efficacy of Casuarinaequisetifolia extracts against some pathogenic microorganisms. J Med Plant Res 6(47):5819–5825
Kantheti USK, Kumar DY, Ganinna B, Nath PK (2014) Casuarinaequisetifolia effect as antidiabetic and antihyperlipidemic on streptozocin induced rats with diabetes. IJCTPR 2(3):432–436
Sriram N (2011) Antidiabetic and antihyperlipidemic activity of bark of Casuarinaequisetifolia on streptozocin induced diabetic rats. Int J Pharm Rev Res 1(1):4–8
El-Tantawy WH, Mohamed SA, Abd Al Haleem EN (2013) Evaluation of biochemical effects of Casuarina equisetifolia extract on gentamicin-induced nephrotoxicity and oxidative stress in rats. Phytochemical analysis. J Clin Biochem Nutr 53(3):158–165
Shalini S, Kumar AS (2011) Study on phytochemical profile and anti-ulcerogenic effect of Casuarina equisetifolia (L.). Asian J Pharm Sci Tech 1(1):12–17
Moazzem Hossen SM, Islam J, Shakhawat Hossain SM, Mofizur Rahman M, Ahmed F (2014) Phytochemical and biological evaluation of MeOH extract of Casuarina equisetifolia (Linn.) leaves. European J Med Plants 4(8):927–936
Mamillapalli V, Atmakuri AM, Khantamneni P (2016) Nanoparticles for herbal extracts. Asian J Pharm 10(2):S54–S60
Abdel-Aziz MS, Shaheen MS, El-Nekeety AA, Abdel-Wahhab MA (2014) Antioxidant and antibacterial activity of silver nanoparticles biosynthesized using Chenopodiummurale leaf extract. J Saudi Chem Soc 18:356–363
Johnson AS, Obot IB, Ukpong US (2014) Green synthesis of silver nanoparticles using Artemisiaannua and Sidaacuta leaves extract and their antimicrobial, antioxidant and corrosion inhibition potentials. J Mater Environ Sci 5(3):899–906
Aboulthana WMK, Refaat E, Khaled SE, Ibrahim NE, Youssef AM (2022) Metabolite profiling and biological activity assessment of Casuarinaequisetifolia bark after incorporating gold nanoparticles. Asian Pac J Cancer Prev 23(10):3457–3471
Gosselin RE, Smith RP, Hodge HC, Jeannet Braddock J (1984) Clinical toxicology of commercial products, 5th edn. Williams and Wilkins, Baltimore, MD
Al-Attar AM (2010) Physiological effects of some plant oils supplementation on streptozotocin-induced diabetic rats. Merit Res J Med Med Sci 5:55–71
Al-Attar AM, Al-Rethea HA (2017) Chemoprotective effect of omega-3 fatty acids on thioacetamide induced hepatic fibrosis in male rats. Saudi J Biol Sci 24:956–965
Ellman GL, Courtney KD, Andres V Jr, Featherstone RM (1961) A new and rapid colorimetric determination of acetyl-cholinesterase activity. Biochem Pharmacol 7(2):88–95
Gorun V, Proinov L, Baltescu V, Balaban G, Barzu O (1978) Modified Ellman procedure for assay of cholinesterase in crude enzymatic preparations. Anal Biochem 86(1):324–326
Schumann G, Klauke R (2003) New IFCC reference procedures for the determination of catalytic activity concentrations of five enzymes in serum: preliminary upper reference limits obtained in hospitalized subjects. Clinica Chim Acta 327(1–2):69–79
Koracevic D, Koracevic G, Djordjevic V, Andrejevic S, Cosic V (2001) Method for the measurement of antioxidant activity in human fluids. J Clin Pathol 54(5):356–361
Beutler E, Duron O, Kelly BM (1963) Improved method for the determination of blood glutathione. J Lab Clin Med 61:882–890
Sun Y, Oberley LW, Li Y (1988) A simple method for clinical assay of superoxide dismutase. Clin Chem 34:479–500
Aebi H (1984) Catalase in vitro. Methods Enzymol 105:121–126
Paglia DE, Valentine WN (1967) Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med 70(1):158–163
Ohkawa H, Ohishi N, Yagi K (1979) Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem 95:351–358
Levine RL, Williams JA, Stadtman ER, Shacter E (1994) Carbonyl assays for determination of oxidatively modified proteins. Methods Enzymol 233:346–357
Engelmann H, Novick D, Wallach D (1990) Two tumor necrosis factor-binding proteins purified from human urine. Evidence for immunological cross-reactivity with cell surface tumor necrosis factor receptors. J Biol Chem 265:1531–1563
March CJ, Mosley B, Larsen A, Cerretti DP, Braedt G, Price V, Gillis S, Henney CS, Kronheim SR, Grabstein K (1985) Cloning, sequence and expression of two distinct human interleukin-1 complementary DNAs. Nature 315:641–647
Reddy GK, Enwemeka CS (1996) A simplified method for the analysis of hydroxyproline in biological tissues. Clin Biochem 29:225–229
Suvarna SK, Layton C, Bancroft JD (2019) Bancroft’s theory and practice of histological techniques. Churchill Livingstone Elsevier, Oxford
Fouad GI, Ahmed KA (2022) Curcumin ameliorates doxorubicin-induced cardiotoxicity and hepatotoxicity via suppressing oxidative stress and modulating iNOS, NF-κB, and TNF-α in rats. Cardiovasc Toxicol 22(2):152–166
Farag OM, Abd-Elsalam RM, Ogaly HA, Ali SE, El Badawy SA, Alsherbiny MA, Li CG, Ahmed KA (2021) Metabolomic profiling and neuroprotective effects of purslane seeds extract against acrylamide toxicity in rat’s brain. Neurochem Res 46(4):819–842
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72(1–2):248–254
Darwesh OM, Moawad H, Barakat OS, Abd El-Rahim WM (2015) Bioremediation of textile reactive blue azo dye residues using nanobiotechnology approaches. RJPBCS 6(1):1202–1211
Subramaniam HN, Chaubal KA (1990) Evaluation of intracellular lipids by standardized staining with a Sudan Black B fraction. J Biochem Biophys Methods 21(1):9–16
Abd Elhalim SA, Sharada HM, Abulyazid I, Aboulthana WM, Abd Elhalim ST (2017) Ameliorative effect of carob pods extract (Ceratonia siliqua L.) against cyclophosphamide induced alterations in bone marrow and spleen of rats. J App Pharma Sci. 7(10):168–181
Siciliano MJ, Shaw CR (1976) Separation and visualization of enzymes on gels. In Chromatographic and electrophoretic techniques. In: Smith I (ed) Chromatographic and electrophoretic techniques, vol 2. Zone electrophoresis, pp 185–209
Rescigno A, Sanjust E, Montanari L, Sollai F, Soddu G, Rinaldi AC, Oliva S, Rinaldi A (1997) Detection of laccase, peroxidase, and polyphenol oxidase on a single polyacrylamide gel electrophoresis. Anal Lett 30(12):2211–2220
Rammesmayer G, Praznik W (1992) Fast and sensitive simultaneous staining method of Q-enzyme, α-amylase, R-enzyme, phosphorylase and soluble starch synthase separated by starch: polyacrylamide gel electrophoresis. J Chromatogr 623(2):399–402
Ahmad A, Maheshwari V, Ahmad A, Saleem R, Ahmad R (2012) Observation of esterase-like-albumin activity during N’-nitrosodimethyl amine induced hepatic fibrosis in a mammalian model. Maced J Med Sci 5(1):55–61
Nei M, Li WS (1979) Mathematical model for studying genetic variation in terms of restriction endonuclease. Proc Natl Acad Sci USA 76:5269–5273
Wang J, Deng X, Zhang F, Chen D, Ding W (2014) ZnO nanoparticle-induced oxidative stress triggers apoptosis by activating JNK signaling pathway in cultured primary astrocytes. Nanoscale Res Lett 9(1):1–12
Ghasemi Pirbalouti A, Shah Vali A, Saghaee F, Shahrzad A, Hamedi B, Shahgholian L (2010) Effect of Cichorium intybus L. extracts and Kelussia oderatassima Mozaff. essential oil on toxic of organophosphouros insecticides in RAT. J Herb Drugs 1(2):31–36
Valimehr S, Sanjarian F, Sohi HH, Sharafi A, Sabouni F (2014) A reliable and efficient protocol for inducing genetically transformed roots in medicinal plant Nepeta pogonosperma. Physiol Mol Biol Plants 20(3):351–356
Sastry M, Ahmed A, Khan MI, Kumar R (2003) Biosynthesis of metal nanoparticles using fungi and actinomycete. Curr Sci 85(2):162–170
Sellers RS, Morton D, Michael B, Roome N, Johnson JK, Yano BL, Perry R, Schafer K (2007) Society of Toxicologic Pathology position paper: organ weight recommendations for toxicology studies. Toxicol Pathol 35(5):751–755
Mossa ATH, Abbassy MA (2012) Adverse haematological and biochemical effects of certain formulated insecticides in male rats. Res J Environ Toxicol 6(4):160–168
Bozkurt A, Yardan T, Ciftcioglu E, Baydin A, Hakligor A, Bitigic M, Bilge S (2010) Time course of serum S100B protein and neuron-specific enolase levels of a single dose of chlorpyrifos in rats. Basic Clin Pharmacol Toxicol 107(5):893–898
Jaiswal C, Verma Y (2012) Effect of alpha-tocopherol on haematological alteration induced by chlorpyrifos in albino rats. Haryana Vet 51:108–110
Michael B, Yano BL, Sellers RS, Perry R, Morton D, Roome N, Johnson JK, Schafer K (2007) Evaluation of organ weights for rodent and non-rodent toxicity studies: a review of regulatory guidelines and a survey of current practices. Toxicol Pathol 35(5):742–750
Pollakova J, Pistl J, Kovalkovicova N, Csank T, Kocisova A, Legath J (2012) Use of cultured cells of mammal and insect origin to assess cytotoxic effects of the pesticide chlorpyrifos. Pol J Environ Stud 21(4):1001–1006
Akhtar N, Srivastava MK, Raizada RB (2009) Assessment of chlorpyrifos toxicity on certain organs in rat, Rattus norvegicus. J Environ Biol 30(6):1047–1053
Chen R, Cui Y, Zhang X, Zhang Y, Chen M, Zhou T, Lan X, Dong W, Pan C (2018) Chlorpyrifos induction of testicular-cell apoptosis through generation of reactive oxygen species and phosphorylation of AMPK. J Agric Food Chem 66:12455–12470
Fereidounni S, Dhawan DK (2018) Understanding the role of quercetin during neurotoxicity induced by Chlorpyrifos. J Phytopharmacol 7(1):33–39
Li D, Huang Q, Lu M, Zhang L, Yang Z, Zong M, Tao L (2015) The organophosphate insecticide chlorpyrifos confers its genotoxic effects by inducing DNA damage and cell apoptosis. Chemosphere 135:387–393
Deabes MM, Aboulthana WM, Ahmed KA, Naguib KM (2021) Assessment of the ameliorative effect of Bacillussubtilis against the toxicity induced by aflatoxin B1 in rats. Egypt J Chem 64(4):2141–2164
Hashem MA, El-Sharkawy NI (2009) Hemato-biochemical and immunotoxicological effects of low electromagnetic field and its interaction with lead acetate in mice. Iraqi J Vet Sci 23(3):105–114
Ambali SF, Onukak C, Idris SB, Yaqub LS, Shittu MU, Aliyu H, Kawu MU (2010) Vitamin C attenuates short-term hematological and biochemical alterations induced by acute chlorpyrifos exposure in Wistar rats. J Med Med Sci 1(10):465–477
Ambali SF, Angani M, Shittu M, Kawu MU (2011) Hematological changes induced by subchronic co-administration of chlorpyrifos and lead in Wistar rats: alleviating effect of vitamin C. Der Pharmacia Sinica 2(2):276–284
El-Deeb AEA, Abd El-Aleem IM, Ghaleb SS (2007) Harmful effect of some insecticides on vital parameters of albino rats. J Egypt Soc Toxicol 36:53–60
Jope RS, Walter-Ryan WG, Alarcon RD, Lally KM (1985) Cholinergic processes in blood samples from patients with major psychiatric disorders. Biol Psychiat 20(12):1258–1266
Wielgomas B, Krechniak J (2007) Effect of α-cypermethrin and chlorpyrifos in a 28-day study on free radical parameters and cholinesterase activity in Wistar rats. Pol J Environ Stud 16(1):91–95
Ricceri L, Venerosi A, Capone F, Cometa MF, Lorenzini P, Fortuna S, Calamandrei G (2006) Developmental neurotoxicity of organophosphorous pesticides: fetal and neonatal exposure to chlorpyrifos alters sex-specific behaviors at adulthood in mice. Toxicol Sci 93:105–113
Yu F, Wang Z, Ju B, Wang Y, Wang J, Bai D (2008) Apoptotic effect of organophosphorus insecticide chlorpyrifos on mouse retina in vivo via oxidative stress and protection of combination of vitamins C and E. Exp Toxicol Pathol 59:415–423
Bekheit RA, Aboulthana WM, Serag WM (2023) Bio- and phyto-chemical study on Nannochloropsisoculata algal extract incorporated with gold nanoparticles, in vitro study. Front Sci Res Technol 5(1):1–17
Kunnaja P, Chansakaow S, Wittayapraparat A, Yusuk P, Sireeratawong S (2021) In vitro antioxidant activity of Litseamartabanica root extract and its hepatoprotective effect on chlorpyrifos-induced toxicity in rats. Molecules 26:1906
Heikal TM, El-Sherbiny M, Hassan SA, Arafa A, Ghanem HZ (2012) Antioxidant effect of selenium on hepatotoxicity induced by chlorpyrifos in male rats. Int J Pharm Pharmaceut Sci 4:603–609
Tanvir EM, Afroz R, Chowdhury MAZ, Khalil MI, Hossain MS, Rahman A, Rashid H, Gan SH (2015) Honey has a protective effect against chlorpyrifos induced toxicity on lipid peroxidation, diagnostic markers and hepatic histoarchitecture. Eur J Integr Med 7:525–533
Mohamed WR, Mehany ABM, Hussein RM (2018) Alpha lipoic acid protects against chlorpyrifos-induced toxicity in Wistar rats via modulating the apoptotic pathway. Environ Toxicol Pharmacol 59:17–23
Kasteel EEJ, Nijmeijer SM, Darney K, Lautz LS, Dorne J-LCM, Kramer NI, Westerink RHS (2020) Acetylcholinesterase inhibition in electric eel and human donor blood: an in vitro approach to investigate interspecies differences and human variability in toxicodynamics. Arch Toxicol 94:4055–4065
Meligi NM, Hassan HF (2017) Protective effects of Erucasativa (rocket) on abamectin insecticide toxicity in male albino rats. Environ Sci Pollut Res 24:9702–9712
Saoudi M, Badraoui R, Rahmouni F, Jamoussi K, El Feki A (2021) Antioxidant and protective effects of Artemisiacampestris essential oil against chlorpyrifos-induced kidney and liver injuries in rats. Front Physiol 12:618582
Uzun FG, Kalender Y (2013) Chlorpyrifos induced hepatotoxic and hematologic changes in rats: the role of quercetin and catechin. Food Chem Toxicol 55:549–556
Ali N, Rashid S, Nafees S, Hasan SK, Shahid A, Majed F, Sultana S (2017) Protective effect of chlorogenic acid against methotrexate induced oxidative stress, inflammation and apoptosis in rat liver: an experimental approach. Chem Biol Interact 272:80–91
Perumal SS, Ekambaram SP, Dhanam T (2017) In vivo antiarthritic activity of the ethanol extracts of stem bark and seeds of Calophylluminophyllum in Freund’s complete adjuvant induced arthritis. Pharm Biol 55(1):1330–1336
Saoudi M, Badraoui R, Bouhajja H, Ncir M, Rahmouni F, Grati M, Jamoussi K, El Feki A (2017) Deltamethrin induced oxidative stress in kidney and brain of rats: protective effect of Artemisiacampestris essential oil. Biomed Pharm 94:955–963
Zhu S, Jiang X, Boudreau MD, Feng G, Miao Y, Dong S, Wu H, Zeng M, Yin J-J (2018) Orally administered gold nanoparticles protect against colitis by attenuating Toll-like receptor 4- and reactive oxygen/nitrogen species-mediated inflammatory responses but could induce gut dysbiosis in mice. J Nanobiotechnol 16:86
Zangeneh MM, Zhaleh M, Salmani S, Zangeneh A, Almasi M, Khedri MR, Rashidi K (2019) Assessment of the anti-anemic effect of aqueous extract of Alliumeriophyllum Boiss leaf in phenylhydrazine-treated Wistar male rats. Com Clin Pathol 28(2):427–434
Patil UK, Saraf S, Dixit VK (2004) Hypolipidemic activity of seeds of Cassiatora Linn. J Ethnopharmacol 90:249–252
Saoudi M, Ncir M, Ben Ali M, Grati M, Jamoussi K, Allouche N, El Feki A (2017) Chemical components, antioxidant potential and hepatoprotective effects of Artemisiacampestris essential oil against deltamethrin-induced genotoxicity and oxidative damage in rats. Gen Physiol Biophys 36:331–342
Ibrahim KA, Khwanes SA (2018) The protective role of propolis against chlorpyrifos induced oxidative stress and DNA damage in the liver of male albino rats. Egypt J Agric Res 96(1):85–94
Ojha A, Gupta YK (2017) Study of commonly used organophosphate pesticides that induced oxidative stress and apoptosis in peripheral blood lymphocytes of rats. Hum Exp Toxicol 36(11):1158–1168
Saoudi M, Hmida IB, Kammoun W, Rebah FB, Jamoussi K, Feki AE (2018) Protective effects of oil of Sardinella pilchardis against subacute chlorpyrifos-induced oxidative stress in female rats. Arch Environ Occup Health 73:128–135
Kanbur M, Eraslana G, Silicib S (2009) Antioxidant effect of propolis against exposure to propetamphos in rats. Ecotoxicol Environ Safety 72:909–915
Dib I, Fauconnier ML, Sindic M, Belmekki F, Assaidi A, Berrabah M, Mekhfi H, Aziz M, Legssyer A, Bnouham M, Ziyyat A (2017) Chemical composition, vasorelaxant, antioxidant and antiplatelet effects of essential oil of Artemisia campestris L. from oriental Morocco. BMC Complement Altern Med 17(1):82
Aboulthana WM, Youssef AM, Seif MM, Osman NM, Sahu RK, Ismael M, El-Baz HA, Omar NI (2021) Comparative study between Crotontiglium seeds and Moringaoleifera leaves extracts, after incorporating silver nanoparticles, on murine brains. Egypt J Chem 64(4):1709–1731
Tanvir EM, Afroz R, Chowdhury MAZ, Gan SH, Karim N, Islam MN, Khalil MI (2016) A model of chlorpyrifos distribution and its biochemical effects on the liver and kidneys of rats. Hum Exp Toxicol 35(9):991–1004
Turk E, Kandemir FM, Yildirim S, Caglayan C, Kucukler S, Kuzu M (2019) Protective effect of hesperidin on sodium arsenite-induced nephrotoxicity and hepatotoxicity in rats. Biol Trace Elem Res 189(1):95–108
Abolaji AO, Ojo M, Afolabi TT, Arowoogun MD, Nwawolor D, Farombi EO (2017) Protective properties of 6-gingerol-rich fraction from Zingiberofficinale (ginger) on chlorpyrifos-induced oxidative damage and inflammation in the brain, ovary and uterus of rats. Chem Biol Interact 270:15–23
Jang Y, Lee AY, Jeong S-H, Park K-H, Paik M-K, Cho N-J, Kim J-E, Cho M-H (2015) Chlorpyrifos induces NLRP3 inflammasome and pyroptosis/apoptosis via mitochondrial oxidative stress in human keratinocyte HaCaT cells. Toxicology 338:37–46
Liang Y, Zhan J, Liu D, Luo M, Han J, Liu X, Liu C, Cheng Z, Zhou Z, Wang P (2019) Organophosphorus pesticide chlorpyrifos intake promotes obesity and insulin resistance through impacting gut and gut microbiota. Microbiome 7:19
Zhang Y, Jia Q, Hu C, Han M, Guo Q, Li S, Bo C, Zhang Y, Qi X, Sai L, Peng C (2021) Effects of chlorpyrifos exposure on liver inflammation and intestinal flora structure in mice. Toxicol Res (Camb) 10(1):141–149
Albasher G, Almeer R, Al-Otibi FO, Al-Kubaisi N, Mahmoud AM (2019) Ameliorative effect of Betavulgaris root extract on chlorpyrifos-induced oxidative stress, inflammation and liver injury in rats. Biomolecules 9:261
Joshi AKR, Rajini P (2012) Hyperglycemic and stressogenic effects of monocrotophos in rats: evidence for the involvement of acetylcholinesterase inhibition. Exp Toxicol Pathol 64:115–120
Imam A, Sulaiman NA, Oyewole AL, Amin A, Shittu S-TT, Ajao MS (2018) Pro-neurogenic and antioxidant efficacy of Nigellasativa oil reduced vulnerability to cholinesterase dysfunction and disruption in amygdala dependent behaviors in CPF exposure. JKIMSU 7:1–12
Rahmouni F, Badraoui R, Amri N, Elleuch A, El-Feki A, Rebai T, Saoudi M (2019) Hepatotoxicity and nephrotoxicity in rats induced by carbon tetrachloride and the protective effects of Teucriumpolium and vitamin. Toxicol Mech Meth 29(5):313–321
Sumbayev VV, Yasinska IM, Garcia CP, Gilliland D, Lall GS, Gibbs BF, Bonsall DR, Varani L, Rossi F, Calzolai L (2013) Gold nanoparticles downregulate interleukin-1β-induced pro-inflammatory responses. Small 9(3):472–477
Dkhil MA, Bauomy AA, Diab MS, Al-Quraishy S (2015) Antioxidant and hepatoprotective role of gold nanoparticles against murine hepatic schistosomiasis. Int J Nanomedicine 10:7467–7475
Almazroo OA, Miah MK, Venkataramanan R (2017) Drug metabolism in the liver. Clin Liver Dis 21:1–20
Altun S, Özdemir S, Arslan H (2017) Histopathological effects, responses of oxidative stress, inflammation, apoptosis biomarkers and alteration of gene expressions related to apoptosis, oxidative stress, and reproductive system in chlorpyrifos-exposed common carp (Cyprinuscarpio L.). Environ Pollut 230:432–443
Latuszynska J, Luty S, Raszewski G, Przebirowska D, Tokarska-Rodak M (2003) Neurotoxic effect of dermally applied chlorpyrifos and cypermethrin. Reversibility of changes. Ann Agric Environ Med 10(2):197–201
Ghahremani S, Soodi M, Atashi A (2018) Quercetin ameliorates chlorpyrifos-induced oxidative stress in the rat brain: possible involvment of PON2 pathway. J Food Biochem 42:e12530
Alanazi AZ, Alqahtani F, Mothana RAA, Mohany M, Abuohashish HM, Ahmed MM, Al-Rejaie SS, Balemba OB (2020) Protective role of Loranthusregularis against liver dysfunction, inflammation, and oxidative stress in streptozotocin diabetic rat model. Evid-Based Complement Altern Med 2020:1–8
Pan P, Lin S, Wang Y, Chen W, Chuang Y, Wu C, Chen C (2014) Protective effects of rutin on liver injury induced by biliary obstruction in rats. Free Radic Biol Med 73:106–116
De Araújo Júnior RF, de Araújo AA, Pessoa JB, Neto FPF, da Silva GR, Oliveira ALCSL, de Carvalho TG, Silva HFO, Eugênio M, Sant’Anna C, Gasparotto LHS (2017) Anti-inflammatory, analgesic and anti-tumor properties of gold nanoparticles. Pharmacol Rep 69(1):119–129
Shah PI, Bhatt PA, Patel PS, Ghori VL, Vaghela PD (2010) Study of electrophoretic pattern in serum of multiple myeloma patients. Int J Pharm Health Sci 1(2):116–131
Aboulthana WM, Ismael M, Farghaly HS (2016) Assessment of mutagenicity induced by toxic factors affecting ovarian tissue in rats by electrophoresis and molecular dynamic modeling. Int J Curr Pharm Rev Res 7:347–359
Harrouk W, Codrington A, Vinson R, Robaire B, Hales BF (2000) Paternal exposure to cyclophosphamide induces DNA damage and alters the expression of DNA repair genes in the rat preimplantation embryo. Mutat Res 461:229–241
Cheshenko K, Pakdel F, Segner H, Kah O, Eggen RIL (2008) Interference of endocrine disrupting chemicals with aromatase CYP19 expression or activity and consequences for reproduction of teleost fish. Gen Comp Endocrinol 155:31–62
Senturk M, Ceyhun SB, Erdogan O, Kufrevioglu OI (2009) Invitro and invivo effects of some pesticides on glucose-6-phosphate dehydrogenase enzyme activity from rainbow trout (Oncorhynchusmykiss) erythrocytes. Pestic Biochem Phys 95:95–99
Javed A, Ashwini LS, Muralidhar TS, Sagar S, Medam SK (2014) Effect of quercetin on cyclophosphamide induced biochemical profiles in rat liver. Int J Res Stud Biosci 2(10):40–46
Hassan HA, Yousef MI (2010) Ameliorating effect of chicory (Cichorium intybus L.)-supplemented diet against nitrosamine precursors-induced liver injury and oxidative stress in male rats. Food Chem Toxicol 48(8–9):2163–2169
Yuvaraj S, Premkumar VG, Vijayasarathy K, Gangadaran SG, Sachdanandam P (2007) Ameliorating effect of coenzyme Q10, riboflavin and niacin in tamoxifen-treated postmenopausal breast cancer patients with special reference to lipids and lipoproteins. Clin Biochem 40:623–628
El-Sayed AB, Aboulthana WM, El-Feky AM, Ibrahim NE, Seif MM (2018) Bio and phyto-chemical effect of Amphoracoffeaeformis extract against hepatic injury induced by paracetamol in rats. Mol Biol Rep 45(6):2007–2023
Surowiak P, Maciejczyk A, Materna V, Drag-Zalesinska M, Wojnar A, Pudelko M, Kedzia W, Spaczynski M, Dietel M, Zabel M, Lage H (2007) Unfavourable prognostic significance of S100P expression in ovarian cancers. Histopathology 51(1):125–128
Abulyazid I, Abd Elhalim SA, Sharada HM, Aboulthana WM, Abd Elhalim STA (2017) Hepatoprotective effect of carob pods extract (Ceratonia siliqua L.) against cyclophosphamide induced alterations in rats. Int J Curr Pharm Rev Res 8(2):149–162
Aboulthana WM, Ibrahim NE, Osman NM, Seif MM, Hassan AK, Youssef AM, El-Feky AM, Madboli AA (2020) Evaluation of the biological efficiency of silver nanoparticles biosynthesized using Croton tiglium L. seeds extract against azoxymethane induced colon cancer in rats. Asian Pac J Cancer Prev 21(5):1369–1389
Aboulthana WM, Youssef AM, El-Feky AM, Ibrahim NE, Seif MM, Hassan AK (2019) Evaluation of antioxidant efficiency of Croton tiglium L. seeds extracts after incorporating silver nanoparticles. Egypt J Chem 62(2):181–200
Kim MS, Kim HR, So HS, Lee YR, Moon HC, Ryu DG, Yang SH, Lee GS, Song JH, Kwon KB (2014) Crotonis fructus and its constituent, croton oil, stimulate lipolysis in OP9 adipocytes. Evid Based Complement Alternat Med. 2014(780385):5
Ramanathan M, Jaiswal AK, Bhattacharya SK (1999) Superoxide dismutase, catalase and glutathione peroxidase activities in the brain of streptozotocin induced diabetic rats. Indian J Exp Biol 37(2):182–183
Mansouri M, Pirouzi M, Saberi MR, Ghaderabad M, Chamani J (2013) Investigation on the interaction between cyclophosphamide and lysozyme in the presence of three different kind of cyclodextrins: determination of the binding mechanism by spectroscopic and molecular modeling techniques. Molecules 18:789–813
Codrington AM, Hales BF, Robaire B (2007) Chronic cyclophosphamide exposure alters the profile of rat sperm nuclear matrix proteins. Biol Reprod 77:303–311
Najafian M, Jahromi MZ, Nowroznejhad MJ, Khajeaian P, Kargar MM, Sadeghi M, Arasteh A (2011) Phloridzin reduces blood glucose levels and improves lipids metabolism in streptozotocin-induced diabetic rats. Mol Biol Rep 1(1):1–8
Udia PM, Takem LP, Ufot UF, Antai AB, Owu DU (2016) Insulin and alpha amylase levels in alloxan-induced diabetic rats and the effect of Rothmannia hispida (K. Schum) Fagerl leaf extract. J Phytopharmacol 5:1–5
Benjamin S, Pradeep S, Josh MKS, Kumar S, Masai E (2015) A monograph on the remediation of hazardous phthalates. J Hazard Mater 298:58–72
Srividhya R, Gayathri R, Kalaiselvi P (2012) Impact of epigallo catechin-3-gallate on acetylcholine-acetylcholine esterase cycle in aged rat brain. Neurochem Int 60:517–522
Seif MM, Ahmed-Farid OA, Aboulthana WM (2017) Evaluation of the protective effect of Acaciasenegal extract against di-(2-ethylhexyl phthalate) induced hepato- and neurotoxicity in rats. Annu Res Rev Biol 19:1–17
El-Zayat EM (2007) Isoenzyme pattern and activity in oxidative stress-induced hepatocarcinogenesis: the protective role of selenium and vitamin E. Res J Med Sci 2:62–71
Fraga CG, Galleano M, Verstraeten SV, Oteiza PI (2010) Basic biochemical mechanisms behind the health benefits of polyphenols. Mol Aspects Med 31(6):435–445
Singh V, Panwar R (2014) In vivo antioxidative and neuroprotective effect of 4-allyl-2-methoxyphenol against chlorpyrifos-induced neurotoxicity in rat brain. Mol Cell Biochem 388(1–2):61–74
Jiang W, Li S, Hu S, Wang J, Song W, Ji L, Wang Y (2015) Fucosylated chondroitin sulfate from sea cucumber inhibited islets of Langerhans apoptosis via inactivation of the mitochondrial pathway in insulin resistant mice. Food Sci Biotechnol 24:1105–1113
El Gamal AA, AlSaid MS, Raish M, Al-Sohaibani M, Al-Massarani SM, Ahmad A, Hefnawy M, Al-Yahya M, Basoudan OA, Rafatullah S (2014) Beetroot (Beta vulgaris L.) extract ameliorates gentamicin-induced nephrotoxicity associated oxidative stress, inflammation, and apoptosis in rodent model. Mediat Inflamm 2014:983952
Abdel-Halim AH, Fyiad AA, Aboulthana WM, El-Sammad NM, Youssef AM, Ali MM (2020) Assessment of the anti-diabetic effect of Bauhiniavariegata gold nano-extract against streptozotocin induced diabetes mellitus in rats. J Appl Pharm Sci 10(05):077–091
Zhou Y-T, He W, Wamer WG, Hu X, Wu X, Lo YM, Yin J-J (2013) Enzyme-mimetic effects of gold@platinum nanorods on the antioxidant activity of ascorbic acid. Nanoscale 5:1583–1591
Acknowledgements
The authors wish to thank the National Research Centre, Dokki, Giza, Egypt, for the facilities provided and all technical support to accomplish this research work. They wish to thank Dr. Esraa Refaat and Dr. Sally Eid Khaled, Pharmacognosy Department, Pharmaceutical and Drug Industries Research Institute, National Research Centre, for their role in the preparation of the plant extract. Also, they wish to thank Prof. Dr. Ahmed Mahmoud Youssef, Packaging Materials Department, National Research Centre, for his role in the biosynthesis of Au-NPs from the plant extract.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
W.M.A., N.E.I., and A.K.H. suggested the idea and collected the previously published papers related to it. Also, they put the practical plan. H.A. and H.A.T. were concerned with preparation of plant extract and chlorpyrifos solution injected to rats. W.M.A., N.E.I., and W.K.B. were concerned with undergoing both biochemical and molecular experiments. K.A.A. was responsible for undergoing the histopathological examination. All authors gave their approval to the final draft of the manuscript. W.M.A. communicated with journals’ editors asking to publish the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The experimental animal design was carried out in accordance with the protocol published in the “Guide for the care and use of laboratory animals” and authorized by the Institutional Animal Ethics Committee of the National Research Centre, Dokki, Giza, Egypt (No.: 131512012023).
Consent for publication
Not applicable.
Competing interests
The authors declare that no conflicts of interest (financial or non-financial) were among them.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplementary Table 1.
The primers with the sequences suitable to be used for qRT-PCR. Supplementary Table 2. Effect of C. equisetifolia extract incorporated with gold nanoparticles (Au-NPs) against toxicity induced by chlorpyrifos (CPF) on different hematological measurements in rats. Data were calculated from five replicates and expressed as mean ± SE. aSignificant versus control group, bSignificant versus toxic (CPF) group at P≤0.05. Supplementary Table 3. Effect of C. equisetifolia extract incorporated with gold nanoparticles (Au-NPs) against toxicity induced by chlorpyrifos (CPF) on different biochemical measurements in rats. Data were calculated from five replicates and expressed as mean ± SE. aSignificant versus control group, bSignificant versus toxic (CPF) group at P≤0.05. Supplementary Figure 1. Effect of C. equisetifolia extract incorporated with gold nanoparticles (Au-NPs) against toxicity induced by chlorpyrifos (CPF) on a) the change in body weights, b) the relative organ weights (organ/body weights ratio) of rats. Data were calculated from five replicates and expressed as mean ± SE. aSignificant versus control group, bSignificant versus toxic (CPF) group at P≤0.05.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Aboulthana, W.M., Ibrahim, N.ES., Hassan, A.K. et al. The hepato- and neuroprotective effect of gold Casuarina equisetifolia bark nano-extract against Chlorpyrifos-induced toxicity in rats. J Genet Eng Biotechnol 21, 158 (2023). https://doi.org/10.1186/s43141-023-00595-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43141-023-00595-6