
Abstract
Background
Morocco is considered one of the main biodiversity hotspots in the Mediterranean region and contains various plant species including wild and domestic Rosa. This genus is the most important among cultivated ornamental plants in the world, with a high economic value in cosmetics, pharmaceutical industries, and floriculture. In the present study, genetic diversity among the collected accessions of wild Rosa species in Morocco was assessed using Inter-Simple Sequence Repeat (ISSR) and Directed Amplification of Minisatellites DNA (DAMD) markers.
Results
Results confirmed that both markers used have a good efficiency to assess genetic diversity in wild roses. Ten ISSR and eight DAMD primers amplified 276 and 203 loci, with an average of 27.4 and 25 polymorphic alleles per primer, respectively. The polymorphic information content (PIC) values were 0.34 with ISSR and 0.31 with DAMD. Analysis of molecular variance (AMOVA) showed that genetic variation in wild rose occurs mainly within populations (86%) rather than between populations (14%). The region of Azrou (Middle Atlas of Morocco) is the area that registered the highest genetic diversity in the present study with He = 0.21. The 39 rose accessions were divided into three main groups with among-group similarity of 30%. Principal component analysis and the hierarchical classification were consistent with genetic relationships derived by structure analysis.
Conclusion
The findings revealed that the patterns of grouping are weakly correlated with geographical origin. ISSR and DAMD markers showed that the accessions have a good genetic diversity.
Similar content being viewed by others


Background
The genus Rosa represents one of the largest genera in the Rosaceae family and is comprised of more than 200 terrestrial species and more than 18,000 cultivars but only ten species have contributed to modern commercial roses [1]. Wild rose has extremely high economic, ornamental [2, 3], and medicinal values [4]. Rose cultivars are derived from eight to ten wild species hybridization [5]. Several researches reported that Rosa distribution concern the temperate and sub-tropical regions of the northern hemisphere, specifically in Europe, Asia, the Middle East, and North America [6, 7]. However in North Africa, particularly in Morocco, fifteen species of wild roses, which spread in the northern half of the country, particularly in the Atlas Mountains and the Rif, were reported by Fennane et al. (1999) [8].
Recent research has classified the genus Rosa into four subgenera: Hulthemia, Platyrhodon, Hesperhodos, which are monotypic or include two species, and Rosa genus, which contains all the remaining species, grouped into 10 sections [9]. Generally, the classification of the Rosa genus has posed many problems to taxonomists as a gradation between many subdivisions is slight, and in many instances, segregation is a matter of opinion [10]. Genetic relationships within this genus are confusing due to the variability of species and the weak barriers to intraspecific hybridization [11]. For this purpose, it is recommended to identify the species and the biological relationships within the Rosa genus for use in genetic improvement [12, 13]. Molecular markers were found to be more efficient in selection compared to conventional morphology-based methods. They provide easy and accurate access to genetic variability and determine polymorphism at the DNA level without environmental interference [14].
Since the mid-1980s, genome identification and selection has progressed rapidly with the help of PCR technology. A large number of marker protocols that are rapid and require only small quantities of DNA have been developed [15]. The widely-used PCR-based markers are RAPDs [16], SSRs or microsatellites [17], AFLPs [18], ISSRs [19], and DAMDs [20].
In the 1990s, molecular markers were developed for rose cultivar identification [21, 22] and several of these were tested to identify species relationships in Rosa [21, 22]. used RAPDs to examine the relationships among cultivars and a limited number of wild species. The work of [21] showed a distinction between a group of cultivars and that of wild species. But since 1994, a new molecular marker technique called inter simple sequence repeat (ISSR) has been available [19]. ISSR-PCR is a technique that originated from microsatellite regions and uses microsatellite sequences as primers (16–18 bp) in a polymerase chain reaction to generate multi-locus markers [23]. DAMD, as well, has been developed on the basis of preserved gene regions and can easily generate functional markers related to a given plant phenotype [24]. Both molecular markers are generated from the microsatellite-rich portions of the genome [25].
ISSR and DAMD markers are becoming important tools for genetic diversity analysis in plants. They collectively provide a comprehensive description of the nature and the extent of diversity, its genetic relationship, and germplasm management [26,27,28,29,30,31].
In Morocco, studies on wild roses are very rare even absent at morphological, anatomical, chemical, or genetic levels. The only studies provided to date concern morphological aspects [8]. To fill this gap, particular attention has been devoted to the genetic diversity of wild roses in order to know the potential of this natural resource as a natural heritage, to understand the species relationships with the aim of preservation and proposition of a reliable genetic improvement program in the context of current climate change.
A study of polymorphism among wild rose accessions was carried out using ISSR and DAMD markers. Therefore, the aim of this study is to characterize the genetic variability of wild rose species available in Morocco and the relationship between wild genetic pools. This will allow of performing breeding programs.
Methods
Collection of plant materials
Thirty-nine accessions of Rosa sp. including 4 accessions of hybrid rose (3 Rosa damascena (KMG1, KMG3, and KMG4) and unknown (TOGH)) were collected from different geographical provinces of Morocco (Ifrane, Midelt, Azilal, Errachidia, Tinghir, and Ouarzazate) ranging from 1260 to 2358 m altitude (Fig. 1, Table 1). The sampling mode used in this study is simple random.

Map of Morocco illustrating the geographic distribution of the 39 Moroccan Rose accessions. Zoned 1: South East, zoned 2: High Atlas, and zoned 3: Middle Atlas
DNA extraction and purification
Fresh young leaves (50 mg) were freeze-dried. Total genomic DNA was extracted from the young leaves using the CTAB method [32]. The leaves were crushed using 1 ml of preheated extraction buffer [PVP 0.2%, Tris-HCl 1 M pH 8.0, NaCl 5 M, 2% CTAB, EDTA 0.5 M, 0.2% β-mercaptoethanol] and incubated for 1 hour at 65°C. Then they were treated with an equal volume of chloroform mixture:isoamylic alcohol (24:1; v/v). Tubes were centrifuged at 13,000 rpm at 4°C for 15 min. After centrifugation, the aqueous phase obtained was transferred to a new tube and 700 μl cold isopropanol was added. DNA was precipitated by centrifuging at 13,000 rpm at 4°C for 10 min. Resulting pellets were washed twice with 70% ethanol. Isolated DNA was air-dried and dissolved in 200 μl of TE buffer. DNA quality was evaluated by spectrophotometry and visualized under UV light after electrophoresis on 1% agarose gel.
ISSR analysis
All 39 accessions were analyzed with 10 ISSR primers (Table 2). DNA amplification was performed in 10 μl per reaction. PCR reaction contained 25 ng of genomic DNA, 2.5 μl of 1X buffer (dNTP, MgCl2), 0.5 U of TaqDNA polymerase (BIOLINE, London, UK), 0.5 μM of each forward and reverse primers and 5.3 μl of pure water. Amplification was carried out in thermocycler (AerisTM) with the following program: an initial denaturing step of 5 min at 95 °C, followed by 35 cycles, each had a 95 °C denaturing step for 30 s, a hybridization at 52 to 58.5 °C (depending on primers) for 1 min and extension step at 72 °C for 2 min. The final elongation step was set to 72°C for 8 min. The amplification products were stored at 4 °C until visualization using polyacrylamide gel electrophoresis. After electrophoresis, the gel was removed from the plates and stained with ethidium bromide solution for 5 min. A 50-bp ladder (BIOLINE) was used to evaluate the approximate molecular weight of amplification products.
DAMD analysis
Using eight DAMD primers (Table 3), all 39 accessions were analyzed. DNA amplification was carried out in 10 μl per reaction. PCR reactions contained 25 ng of genomic DNA, 2.5 μl of 1X buffer (dNTP, MgCl2), 0.5 U of TaqDNA polymerase (BIOLINE, London, UK), 0.5 μM of each forward and reverse primers and 5.3 μl of pure water. Amplification was performed in thermocycler (AerisTM) with the following program: an initial denaturing step of 3 min at 94°C, followed by 35 cycles, each had a 92°C denaturing step for 45 s, a hybridization at 40 to 70°C (depending on primers) for 2 min and extension step at 72°C for 2 min. The final elongation step was set to 72°C for 5 min. The amplification products were stored at 4°C until visualization using polyacrylamide gel electrophoresis.
Statistical analysis
Genetic diversity and frequency analysis
ISSR and DAMD profiles were scored as present (1) or absent (0) for each entry and a binary qualitative data matrix was constructed. Very faint and non-reproducible bands were omitted from scoring. The genetic parameters such as observed number of alleles (na), effective number of alleles (ne) [33], Shannon's Information index (I) [34], expected heterozygosity (He), polymorphic band (PB), and percent of polymorphic band (PPB) were calculated by GenALEx program version 6.5. Analysis of molecular variance was calculated by SAS software version 9. Principal component analysis (PCoA) for the 39 accessions of roses was generated using GENSTAT 18th edition.
Marker efficiency analysis
The performance of the molecular markers was measured by calculating various parameters including polymorphism information content (PIC), effective multiplex ratio (EMR), marker index (MI), and resolving power (Rp) for each primer by program iMEC (https://irscope.shinyapps.io/iMEC/).
Genetic relationship analysis
Clustering was done to determine the genetic distance between individuals and to verify the consistency of population genetic variation. The dissimilarity index was performed for cluster analysis using WNJ (Weighted Neighbor-Joining) method in DARWIN program version 6.0.4. The bootstrap analysis running 1000 replication was employed to determine a sampling variance of the genetic similarities calculated from the data sets gained of different marker systems [35].
Genetic structure
The Bayesian-based model carried out the genetic makeup of rose accessions. This was analyzed by STRUCTURE (version 2.3.4) and was compressed into a single “Zip-Rar” file and then uploaded online to “Structure Harvester” (http://taylor0.biology.ucla.edu/structureHarvester/).
Results
A total of 13 ISSRs and 10 DAMDs were used. Ten ISSR primers (I1, I3, UBC811, UBC834, UBC841, UBC842, UBC880, UBC889, ISSR2, and ISSR15) and eight DAMD primers (HBV3, HBV5, HVR, INS, M13, URP2R, URP9F, and URP25F) were selected as they gave better amplification products and showed clear polymorphism (Fig. 2).

Amplification profile of M13 primer (A) and UBC841 (B) among 39 Rose accessions; M, DNA ladder
Polymorphism and efficiency of ISSR markers
ISSR primers produced 276 fragments among which 274 (99.27%) were polymorphic. A set of 10 ISSR primers detected polymorphic bands that varied from 13 (ISSR2) to 39 (UBC842) with an average of 27.4 bands per primer. Altogether, eight primers (I1, I3, UBC811, UBC841, UBC842, UBC880, UBC889, and ISSR2) presented 100% of polymorphic rates, while the lowest polymorphic percent (94.11%) was detected at UBC834. The highest PIC value was recorded for ISSR2 (0.37) followed by ISSR15 (0.36) while the lowest (0.29) was for UBC842 with an average of PIC of 0.34. Effective multiplex ratio (EMR) spanned from 3.9 (UBC834) to 12.1 (ISSR15) with 9.4 as the average. Marker index (MI) was calculated to recognize the usefulness of the ISSR primers system on Rosa sp. which maximum was for ISSR15 (4.42) followed by UBC811 (4.15) while the lowest for UBC834 (1.17) along with a mean of 3.05 per primer. Resolving power average value (Rp) was 11.95. UBC842 primer counted the highest value (16.92) while UBC834 showed the lowest value (5.74) (Table 4).
Polymorphism and efficiency of DAMD primers
Two hundred and three fragments were generated by DAMD primers among which 200 (98.52%) were polymorphic. A set of eight DAMD primers detected polymorphic bands which varied from 6 (URP2R) to 44 (M13) with an average of 25 bands per primer. However, all primers generated multiple patterns ranging from 9 to 44 with an average of 25.37 alleles per loci. Absolutely, seven primers (HBV3, HBV5, HVR, INS, M13, URP9F, and URP25F) registered 100% of polymorphic rates, while the polymorphic percent observed was as low as 66.66% for URP2R. URP2R recorded the highest PIC value of 0.374 followed by HBV3 (0.36) whereas the lowest was URP25F (0.25) with a mean of PIC of 0.31. Effective multiplex ratio (EMR) spanned from 2.77 (URP2R) to 11.77 (M13) with an average of 7.9 per primer. The marker index (MI) was maximum (3.82) for HBV5 followed by M13 with 3.71, while the lowest was 1.03 for URP2R with a mean of 2.11. The highest value counted for resolving power (Rp) is 16.15 for M13 while the lowest was 2.61 detected in URP2R with an average of 9.85 per primer (Table 4).
Genetic diversity analysis
Variations among and within the Rosa sp. populations were evaluated by molecular variance analysis (AMOVA) (Table 5). Based on combined markers data, the results revealed a higher variation within regions (86%), while 14% of the variation was recorded among the regions with a significant PhiPT value (PhiPT = 0.14, p = 0.001).
Percentage of polymorphic loci (PPL) of Roses using the combined ISSR and DAMD markers (Table 6) spanned from 21.92% (Ait Touda) to 73.07% (Azrou). The Azrou region recorded the highest value of the observed number of alleles (Na), effective number of alleles (Ne), Shannon’s information index (I), and expected heterozygosity (He) with values of 1.49, 1.34, 0.33, and 0.21, respectively. On the other hand, Ait Touda registered the lowest values (0.59, 1.15, 0.13, and 0.09, respectively).
Genetic distance
Table 7 revealed the genetic distance among 10 regions including 39 accessions of Rosa sp. based on the combined ISSR and DAMD matrix that ranged from a minimum of 0.05 to a maximum of 0.19. The highest genetic distance (0.19) was registered for the pair of regions such as Kela Mgona and Akka Ait Ouyed. However, the minimum genetic distance (0.05) was recorded for the pair regions of Bouadel and Agoudim.
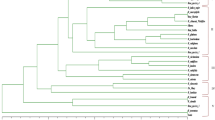
Cluster analysis
The dissimilarity index calculated with data from the combined matrix of ISSR and DAMD markers and used to build a Weighted Neighbor-Joining tree ranged from 0.26 to 0.84. At 70% of dissimilarity, the dendrogram divided the 39 accessions into three major clusters (Fig. 3). The first cluster consisted of 15 wild accessions from Azrou (8 accessions), Ait Touda (2 accessions), and Kela Mgona, Taoudaat, Akka Ait Ouyed, Tirghist, and Tamalout (1 accession). The second cluster comprised 20 wild accessions springing from Bouadel (7 accessions), Agoudim (4 accessions), Tirghist (2 accessions), Tounfite (3 accessions), Tamalout (2 accessions), and Taoudaat and Akka Ait Ouyed (1 accession). Four accessions were classified in the third cluster and comprised 4 hybrid accessions from Kela Mgona (3 accessions) and Tounfite (1 accession).

Dendrogram based on the dissimilarity index values of 39 rose accessions using ISSR and DAMD markers. Accessions are as indicated in Table 1. Bootstrap values (%, based on 1000 replications)
Principal component and population structure analyses
The principal component analysis (PCoA) made it possible to graphically represent the relationship between the ISSR and DAMD markers, and the different accessions (Fig. 4). Derived on the basis of ISSR and DAMD markers data, the first three axes describe a total variation of 26.26%. The first axis (PC1) expressed the largest variation with 12.78% and, the second (PC2) and the third (PC3) accounted for 7.33 and 6.15%, respectively. PCoA showed a significant overlap between accessions, except for 4 accessions from Kela Mgona and 1 accession from Tounfite which was gathered far from other regions.

Principal coordinate analyses (PCoA) for 39 Moroccan Rosa sp. accessions based on ISSR and DAMD data
Using the structure software, we can determine the best value of k in order to characterize the genetic structure of 39 accessions. The program detected the highest value of delta k obtained from the structure harvester, k=3. Based on the best k=3, each assessed accession is often positioned into three different subgroups (Fig. 5). Subgroup 1 consisted of all accessions from Kela Mgona, 4 accessions from Azrou, 1 accession from Tounfite and Akka Ait Ouyed, whereas all accessions from Ait Touda and Taoudaat, 4 accessions from Azrou, 2 accessions from Tamalout and 1 accession from Agoudim and Tirghist were grouped into subgroup 2. All accessions from Bouadel, Tounfite (3), Agoudim (3), Tirghist (2), Akka Ait Ouyed, and Tamalout (1) were allocated to subgroup 3.

Population structure of 39 rose accessions based on ISSR and DAMD data
Discussion
The classical phylogenetic approach relies on the morphological characteristics of an organism. Rosa also have a wide and overlapping range of morphological variations which are influenced by the environment. Classification based on morphology alone is not adequate [36]. Therefore, for these reasons, several molecular markers have been applied in the genetic studies of roses, notably AFLP, RAPD, ISSR, SSR, SCoT, URP, CDDP, and DAMD. The choice of the molecular marker is an important step for researchers and especially geneticists and breeders. Our study analyzed the genetic diversity of the Moroccan rose population, using two PCRs marker molecular ISSR and DAMD.
ISSR and DAMD markers produced high levels of polymorphism, such as the average number of scored bands, polymorphism rates, and PIC were slightly higher with the ISSR primers (27.3, 99.08%, and 0.34, respectively) compared to those of the DAMD primers (25.37, 95.83%, and 0.31, respectively). The high rate of polymorphism revealed by both markers may have been due to the difference in ploidy level, mutations, recombination, natural hybridization, or random segregation of heterozygous chromosomes in the process of meiosis [37]. The polymorphism rate obtained using ISSR and DAMD primers during this study was close to those observed with 47 genotypes of Rosa in Iran (90%) by [15]; this rate was higher than those reported by [13] using ISSR on 7 species of Rosa and by [23] on 23 genotypes of Rosa canina, reaching 75% and 77%, respectively. The mean of polymorphic band obtained in this study using ISSR and DAMD were 27.4 and 25, respectively, and this is much higher than the results reported by [23] using ISSR with an average of 3 and by [38] using DAMD with a mean of 5.87; this is probably due to the fact that we used polyacrylamide gel which revels more band than agarose gel.
In our study, the efficiency of markers was determined by effective multiplex ratio (EMR), marker index (MI), and resolving power (Rp). The values detected for EMR, MI and Rp (8.95, 3.05, and 11.95, respectively) using the ISSR primers are higher than those obtained by DAMD primers (6.68, 2.11, and 9.85, respectively). These results show that the ISSR marker is very efficient compared to the DAMD marker. In accordance with [39], the results revealed that ISSR profiling is an effective tool for assessing genetic diversity.
According to the AMOVA analysis, 86% of genetic variations within regions were shown by the combined ISSR and DAMD markers, indicating that the variation within accessions was higher (86%) than among them (14%). Many researches obtained similar results. Indeed, [40] using URP and SCoT markers on 40 Damask rose genotypes and [41] using 12 pairs of universal cpDNA primers on 12 wild populations revealed within population values of 93% and 77%, respectively, while they obtained 7% and 23% among population. The authors reported that the genetic variation within the collected populations was higher than the variation among them. This indicates that the genetic variation in wild rose mainly occurred within populations.
Maximum values of indices in relation to genetic diversity (Na, Ne, I, He, and PPL) were reported for the Azrou region using combined data. This result indicates that this region could be an important source of diversity for breeding projects which can benefit from new alleles and genes of interest.
Variable genetic distances among rose populations were registered in this study. Results revealed that regions with the lowest distance (Bouadel and Agoudim: 0.05) are found in adjacent geographic regions. This shows a possible existence of seed movement among these areas; unlike populations Kela Mgona (included hybrid accessions) and Akka Ait Ouyed, which exhibited large genetic distances (0.19).
The dissimilarity index calculated with data from the combined matrix of ISSR and DAMD markers ranged from 0.265 to 0.84 with a similarity of 30%. Various studies revealed a high similarity value compared to our findings [42]. using RAPD and SSR markers on 71 Indian local varieties of roses, [15] using ISSR markers, [38] using SCoT, CCDP, and DAMD primers, and [43] using SSR markers detected similarity values with 63%, 52%, 56%, and 67%, respectively. This proved a high genetic variability of roses in Morocco.
Three clusters were recorded in this study. In cluster I, there is an overlap between the regions Ait Touda, Taoudaat, Tamalout, Tirghist, and Akka Ait Ouyed. On the other hand, the accessions of the Azrou region are grouped together in a single sub-clade with the integration of KMG2 accession from Kela Mgona. As a result, the region of Azrou where the endemic species of Morocco Rosa mesatlantica [8] was included is a genetically conserved area that can be considered as a source of variability. Moreover, this indicates that this region could be an important source of diversity. This result leads us to the exploitation of these wild resources and to carry out more research on them. In cluster II, there is also an overlap between the seven regions. Therefore, there is no genetic conservation within these regions. The four hybrids of Kela Mgona and Tounfite are grouped together in the third cluster. This is normal due to the morphological and genetic differences between domesticated and wild rose species. The results of clustering showed that geographical origin was not the main factor causing the genetic differentiation among the wild rose populations.
In the present studies, the results of PCoA analysis were consistent with those of the cluster analysis. The accessions were well distributed in the biplot, which could be due to the relevance of primers used and the wide variety between accessions. The PCoA revealed that Bouadel, Tirghist, Tounfite, Agoudim, and Akka Ait Ouyed regions are very close to each other, which indicates that there is a high genetic flow among them, while this did not follow their origins. The results of PCoA indicated that the wild rose populations were expanded. There may be multiple refuges for wild rose, such as the Azrou and Bouadel populations.
Roses usually cross-pollinate and are self-incompatible which makes them more genetically diverse between and within populations [44,45,46]. analyzed 55 genotypes of roses in Brandenburg (Germany) which were grouped in twelve clusters, whereas the high genetic variation was observed within Rosa canina populations. These authors stated that this variation was due to the polyploidy, outcrossing, and seed dispersal systems. That is in accordance with our study which also showed that most of the species present in Morocco belong to the caninae section [8].
Using the structure harvester software, the best K were identified. The best level of genetic structure in the combined data system was K=3. In this study, high gene flow was detected among rose accessions. This is probably because of plant transmissions by humans or genetic flow and displacement by natural factors [47]. This genetic variation should be considered as a valuable genetic source for breeding programs.
Conclusion
This study showed the genetic diversity of Moroccan wild rose accessions using ISSR and DAMD markers. These markers detected a high level of polymorphism in these accessions. The PIC average values for both markers were 0.34 and 0.31, respectively, which indicates the efficiency of used markers in revealing polymorphism among wild rose accessions. Furthermore, the findings confirmed the efficiency of combined data in estimating the genetic diversity among the populations. The study of genetic diversity is a very important and crucial step that can help us to think of how to use this high level of variation in the breeding program and also how to manage and conserve those wild genetic resources. This study shows also that it is important to exploit wild resources for the genetic improvement of hybrid roses, especially Rosa damascena in Morocco, especially since this country is known as one of the main producers of these roses in the world.
Availability of data and materials
All data generated or analyzed during this study are included in this article.
Abbreviations
- PCR:
-
Polymerase chain reaction
- RAPD:
-
Random amplified polymorphic DNA
- SSR:
-
Simple sequence repeat
- RFLP:
-
Restriction fragment length polymorphism
- SCoT:
-
Start codon targeted
- URP:
-
Universal rice primers
- CCDP:
-
Conserved DNA-derived polymorphism
- iMEC:
-
Marker efficiency calculator
References
Gudin S (2000) Rose: genetics and breeding. In: Janick J (ed) Plant Breeding Reviews. Wiley, New York, pp 159–189
He Y, Dong H, Xue GX (2018) Study on the appearance characteristics and the changing tendency of wild Rosa rugosa Thunb. North Fruits 05:5–9
Xie J, Li MX, Du ZZ (2020) Chemical compositions and their mechanism in cosmetics of Rosa rugosa Thunb. Nat Prod Res Dev 32(02):341–349
Thao NP, Luyen BTT, Jo SH (2014) Triterpenoid saponins from the roots of Rosa rugosa Thunb. as rat intestinal sucrase inhibitors. Arch Pharm Res 37(10):1280–1285
Gudin S (2003) Breeding/Overwiew. In: Roberts AV, Debener T, Gudin S (eds) Encyclopedia of rose science. Elsevier, Academic Press, Oxford
Nilsson OR (1997) In: Davis PH (ed) Flora of Turkey and the east Aegean islands, 4th edn. Edinburgh University Press, Edinburgh, pp 106–128
Samiei L, Naderi R, Khalighi A, Bushehri A, Mozaffarian V, Esselink GD, Smulders MJM (2010) In search of genetic diversity in Rosa foetida Herrmann in Iran. Acta Horticult 836:25–30
Fennane M, Ibn Tattou M, Mathiez J, El Oualidi J, Ouyahya A (1999) Flore pratique du Maroc, vol 1, pp 499–502
Wissemann V (2017) Conventional Taxonomy (Wild Roses). Reference Module in Life Sciences. Elsevier, London, pp 1–6
Takeuchi S, Nomura K, Uchiyama H, Yoneda K (2000) Phylogenetic relationship in the genus Rosa based on the restriction enzyme analysis of the chloroplast DNA. J Jpn Soc Hort Sci 69:598–604
Koopman WJM, Wissemann V, De Cock K, Huylenbroeck JV, De Riek J, Sabatino GJH, Visser D, Vosman B, Ritz CM, Maes B, Werlemark G, Nybom H, Debener T, Linde M, Smulders MJM (2008) AFLP markers as a tool to reconstruct complex relationships: a case study in Rosa (Rosaceae). Am J Bot 95:353–366
Matsumoto S, Kouchi M, Yabuki J, Kusunoki M, Ueda Y, Fukui H (1998) Phylogenetic analysis of the genus Rosa using matK sequence: molecular evidence for the narrow genetic background of modern roses. Sci Hortic 77:73–82
Jabbarzadeh Z, Khosh-Khui M, Salehi H, Saberivand A (2010) Phylogenetic Relationships among Seven Old Rose Species Grown in Iran Revealed by ISSR Markers. Hort Environ Biotechnol 51(3):189–192
Do Val ADB, Ferreira JL, VieriraNeto J, Pasqual M, de Oliviera AF, Borém A, Cançado GMA (2012) Genetic diversity of Brasilian and introduced olive germplasm based on microsatellite markers. Genet Mol Res 11(1):556–571
Jabbarzadeh Z, Khosh-khui M, Salehi H, Shahsavar AR, Saberivand A (2013) Assessment of Genetic Relatedness in Roses by ISSR Markers. World Appl Sci J 28(12):2085–2090
Williams JGK, Kubelik AR, Liavak KJ, Rafalski JA, Tingey SV (1990) DNA polymorphisms amplified by arbitary primers are useful as genetic markers. Nucleic Acids Res 18:6531–6535
Tautz D (1989) Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucleic Acids Res 17:6463–6471
Vos PR, Hogers M, Bleeker M, Reijans T, Van der Lee M, Hornes A, Fritjers J, Pot J, Peleman M, Kuiper ZM (1995) AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res 23:4407–4414
Zietkiewicz E, Rafalski A, Labuda D (1994) Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 20:176–183
Heath DD, Iwana GK, Delvin RH (1993) PCR primed with VNTR core sequences yields species specific patterns and hypervariable probes. Nucleic Acids Res 21:5782–5785
Debener T, Bartels CH, Mattiesch L (1996) RAPD analysis of genetic variation between a group of rose cultivars and selected wild rose species. Mol Breed 2:321–327
Millan F, Osuma F, Cobos S, Torres A, Cuber JI (1996) Using RAPDs to study phylogenetic relationships in Rosa. Theor Appl Genet 92:273–277
Jamali M, Ghanbari A, Estaji A, Giglou MT, Saidi M (2019) Genetic diversity of dog rose (Rosa canina L.) using ISSR markers. Iran J Genet Plant Breed 8(2):1–8
Poczai P, Varga I, Laos M, Cseh A, Bell N, Valkonen JP, Hyvönen J (2013) Advances in plant gene–targeted and functional markers: a review. Plant Methods 9(1):6
Kumar A, Mishra P, Singh SC, Sundaresan V (2014) Efficiency of ISSR and RAPD markers in genetic divergence analysis and conservation management of Justicia adhatoda L., a medicinal plant. Plant Syst Evol 300:1409–1420
Bhattacharya E, Dandin SB, Ranade SA (2005) Single primer amplification methods reveal exotic and indigenous mulberry varieties are similarly diverse. J Biosci 30:669–677
Kumar S, Kumaria S, Sharma SK, Rao SR, Tandon P (2011) Genetic diversity assessment of Jatropha curcas L. germplasm from Northeast India. Biomass Bioenergy 35:3063–3070
Kumar S, Kumaria S, Tandon P (2013) SPAR methods coupled with seed-oil content revealed intra-specific natural variation in Jatropha curcas L. from Northeast India. Biomass Bioenergy 54:100–106
Ranade SA, Rana TS, Narzary D (2009) SPAR profile and genetic diversity amongst pomegranate (Punica granatum L.) genotypes. Physiol MolBiolPlants 15:61–70
Sharma SK, Rawat D, Kumar S, Kumar A, Kumaria S, Rao SR (2010) Single primer amplification reaction (SPAR) reveals intra-specific natural variation in Prosopis cineraria (L.) Druce. Trees Struct Funct 24:855–864
Sharma SK, Kumaria S, Tandon P, Rao SR (2011) Single primer amplification reaction (SPAR) reveals inter-and intra-specific natural genetic variation in five species of Cymbidium (Orchidaceae). Gene 483:54–62
Doyle JJ, Doyle JL (1990) Isolation of plant DNA from fresh tissue. Focus 12:13–15
Kimura M, Weiss GH (1964) The stepping stone model of population structure and the decrease of genetic correlation with distance. Genetics 49:561–576
Lewontin RC (1972) Testing the theory of natural selection. Nature 236:181–182
Perrier X, Flori A, Bonnot F (2003) Methods for data analysis. Genetic diversity of cultivated tropical plants. Science Publishers, Inc. and CIRAD, Montpellier, pp 31–63
Lewis WH (1957) Revision of the genus Rosa in Eastern North America: a review. Am Rose Annu 42:116–126
Smith JF, Burke CC, Wagner WL (1996) Interspecific hybridization in natural population of Crytandra (Gesneriaceae) on the Hawaiian Islands: evidence from RAPD markers. Plant Syst Evol 200:61–77
Saidi A, Eghbalnegad Y, Hajibarat Z (2017) Study of genetic diversity in local rose varieties (Rosa spp.) using molecular markers. Banat’s. J Biotechnol VIII(16):148–157
Namita PS, Sonah H, Singh SP, Sharma TR (2013) Genetic diversity analysis of marigold (Tagetes sp) genotypes using RAPD and ISSR markers. Indian J Agric Sci 83(5):484–490
Mostafavi AS, Omidi M, Azizinezhad R, Etminan A, Badi HN (2021) Genetic diversity analysis in a mini core collection of Damask rose (Rosa damascena Mill.) germplasm from Iran using URP and SCoT markers. J Genet Eng Biotechnol 19(144):1–14
Xu J, Zang F, Wu Q, Wang Y, Wang B, Huang P, Zang D, Ma Y, Zheng Y (2021) Analysis of the genetic diversity and molecular phylogeography of the endangered wild rose (Rosa rugosa) in China based on chloroplast genes. Glob Ecol Conserv 28:e01653
Kaul K, Karthigeyan S, Dhyani D, Kaur N, Sharma RK, Ahuja PS (2009) Morphological and molecular analyses of Rosa damascena × R. bourboniana interspecific hybrids. Sci Hortic 122(2):258–263
Gaurav AK, Namita RDVS, Ramkumar MK, Singh MK, Singh B, Krishnan SG, Panwar S, Sevanthi AM (2021) Genetic diversity analysis of wild and cultivated Rosa species of India using microsatellite markers and their comparison with morphology-based diversity. J Plant Biochem Biotechnol 31:61–70
Cole P, Melton B (1986) Self-and cross-compatibility relationships among genotypes and between ploidy of the rose. J Am Soc Hort Sci 111(1):122–125
Charlesworth D (2003) Effects of inbreeding on the genetic diversity of populations. Philos Trans R Soc Lond B Biol Sci 358(1434):1051–1070
Jürgens R, Ball A, Verster A (2009) Interventions to reduce HIV transmission related to injecting drug use in prison. Lancet Infect Dis 9(1):57–66
Percifield RJ, Hawkins JS, Mccoy JA, Widrlechner MP, Wendel JF (2007) Geneticdiversity in Hypericum perforatum and AFLP markers for species–specific identification of H. perforatum L. Plant Med 739:1614–1621
Acknowledgements
The authors would like to acknowledge Dr. FOKAR Mohamed from Texas Tech University (USA) for the English revision of the manuscript
Funding
Not applicable
Author information
Authors and Affiliations
Contributions
KS, GD, YA, and RA conceived the research idea and designed the experiments. KS, YA, GD, RA, and DI collected the plant materials and revised and approved the final manuscript. KS, NL, GD, RA, YA, and MI conceptualized and supervised the experiment. FG and YE analyzed the data. The authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Saghir, K., Abdelwahd, R., Iraqi, D. et al. Assessment of genetic diversity among wild rose in Morocco using ISSR and DAMD markers. J Genet Eng Biotechnol 20, 150 (2022). https://doi.org/10.1186/s43141-022-00425-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43141-022-00425-1