
Abstract
Background
Inflammation is known to involve in many pathological processes of different diseases, but the current therapy causes adverse effects. Thus, there is a great interest for the discovery of flavonoids as a valuable alternative to classical analgesic and anti-inflammatory agent with dual-inhibitory action, especially on both COX-2 and 5-LOX which can minimize or overcome this problem.
Results
In the present work, drug-likeness properties of the synthesized flavonoids via Lipinski’s Rule of Five were predicted using QikProp prior to evaluation of their COX and LOX inhibitory activities using enzyme assays. Subsequently, molecular docking was performed using GLIDE to analyse their binding behaviour. The results showed that all compounds obeyed the Lipinski’s Rule of Five. NPC6 and NPC7 had displayed better selectivity towards COX-2 as compared to Indomethacin with less than 50% inhibition against COX-1. In addition, these compounds also inhibited activity of 5-LOX. Their selectivity to COX-2 was due to the binding to hydrophobic region and extends to lobby region near the entrance of COX binding site forming hydrogen bond with Ser530. Interestingly, these compounds showed a similar binding mode as Zileuton in the active site of 5-LOX and formed hydrogen bond interaction with Ala424.
Conclusion
NPC6 and NPC7 had potential as dual inhibitor of COX-2 and 5-LOX. The scaffolds of these chemical entities are useful to be as lead compounds for the dual inhibition of COX-2 and 5-LOX.
Similar content being viewed by others
1 Background
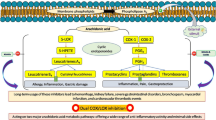
Arachidonic acid (AA) and its metabolites (prostaglandins and leukotrienes) are important intracellular messengers which play an important role in pain and inflammation regulatory pathways [1]. AA is a 20-carbon polyunsaturated fatty acid which is released from membrane phospholipid by phospholipase A2 (PLA2). It is then metabolized by cyclooxygenase (COX) and 5-lipoxygenase (5-LOX) for biosynthesis of prostaglandins and leukotrienes, respectively. COX catalyses the conversion of AA to prostaglandin G2 (PGG2) and PGG2 to PGH2. PGH2 is then transformed into a number of prostaglandins (PGE2, PGD2, PGF2α, PGI2) and thromboxane A2 (TXA2) involved in fever, pain, swelling, inflammation, and platelet aggregations [2]. COX-1 is involved in the synthesis of prostaglandins that is necessary for haemostatic integrity, gastric mucosal protection, renal function and platelet aggregation. Meanwhile, prostaglandin which is derived from COX-2 catalytic pathway is produced when inflammation occurs. Therefore, COX-2 selective NSAIDs inhibit prostanoids that are involved in the inflammation without disrupting COX-1 functions and avoid gastrointestinal and renal toxicity theoretically [3].
5-LOX is the other main pathway in the AA cascade. It catalyses the oxidation of AA to 5-hydroperoxyeicosatetratenoic acid (5-HPETE), which is converted into leukotrienes (LT) such as LTB4, LTC4, LTD4 and LTE4 [4]. The leukotrienes mediators such as LTB4 regulate the innate immune response and play roles in chronic inflammation such as rheumatoid arthritis, osteoporosis and asthma [5,6,7,8]. Targeting several arachidonic acid pathways may lead to effective treatment while maintaining normal physiological function. Therefore, concurrent inhibition of COX-2 and 5-LOX but not COX-1 is a good strategy to produce potential anti-inflammatory drugs with fewer side effects.
Non-steroidal anti-inflammatory drugs (NSAIDs) are widely used as an analgesic and anti-inflammatory agent to treat inflammatory-related diseases including dysmenorrhea, osteoarthritis, rheumatoid arthritis, gout, ocular inflammation, ankylosing spondylitis, actinic keratosis, tendinitis, and bursitis. However, long-term treatment with traditional NSAIDs (tNSAIDs) such as ibuprofen, indomethacin, diclofenac, ketoprofen, naproxen, piroxicam and nabumetone are associated with gastrointestinal bleeding, ulceration and perforation [9, 10]. Therefore, several approaches have been considered to target COX-2 and 5-LOX in order to block the formation of prostaglandins and leukotrienes induced during inflammation. This approach is trendy in drug discovery to develop drugs with multi-targets and has a superior safety profile but minimal adverse effects on gastro, renal and cardiovascular system [11].
Recent research has focused on the identification of potential COX-2 as well as 5-LOX inhibitors based on the scaffold of flavonoids [12, 13]. Flavonoids consist of a benzene ring (A) that is condensed with a six membered ring (C) and carry a phenyl ring (B) at the 2-position [14]. Its subclasses, chalcones, and flavanones, had been reported to exhibit anti-inflammatory activity in both proliferative and exudative phases of inflammation via inhibition of various enzymes such as nitric oxide synthase, xanthine oxidase, aldose reductase, LOX, and COX [15, 16]. In the continuous effort to find a dual inhibitor of COX-2/5-LOX, the present study investigated the anti-cyclooxygenase and anti-lipoxygenase properties of chalcone and flavanone derivatives using an enzymatic assay. The derivatives of flavonoids were synthesized based on our previous study that reported their potential as anti-inflammatory agents [17]. The study also analysed the binding mode of the active compounds and predicted the drug-likeness properties of the compounds. As far as we know, this is the first time that these derivatives were tested for their dual COX-2/5-LOX inhibitory activities.
2 Methods
2.1 Chemicals
The chemicals used were analytical grade and commercially available. All chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise stated. The chalcone, flavone, pinostrobin, and pinocembrin derivatives were obtained from the Institute of Science (IOS), UiTM (Table 1) [18]. These compounds were synthesized using the previous method with some modifications [19].
2.2 Drug-likeness properties
The drug-likeness properties of the synthetic compounds were performed using Qikprop module in Schrödinger package.Footnote 1 It was evaluated based on Lipinski’s Rule of Five, where the properties are molecular weight (MW) ≤ 500 Dalton, the number of hydrogen-bond donors (HBD) ≤ 5, the number of hydrogen-bond acceptors (HBA) ≤ 10 and the octanol/water partition coefficient (log P) < 5. The compounds that violated more than one of Lipinski’s Rule of Five were excluded from further study. Besides that, the number of rotatable bonds was also calculated where the acceptable range was between 0 and 15.
2.3 COX peroxidase assay
The inhibitory activity of COX was determined by measuring the formation of oxidized N,N,N’,N’-tetramethyl-p-phenylenediamine (TMPD) spectrophotometrically at 550 nm. The experiment was carried out based on a previous study with modifications [20]. The inhibitory activity of the synthetic compounds was tested against the human COX-1 and human recombinant COX-2 enzymes whilst the compounds were replaced with dimethyl sulphoxide (DMSO) as the total activity of the enzymes. Reactions were initiated in a 96-well plate at a final volume of 220 µL. COX inhibition reaction was performed by 10 min of incubation at room temperature in the presence of reaction buffer (Tris buffer, 0.1 M, pH 8.0, 150 µL), haem (2.2 mM in DMSO, 10 µL), COX-1 or COX-2 enzymes and tested compounds (100 µM, diluted in DMSO, 10 µL). The reaction was initiated by adding 20 μL of freshly prepared TMPD and followed by the addition of 20 μL of AA (diluted with ethanol). The absorbance was measured at 550 nm after 5 min of incubation at room temperature and the calculations were performed as in Eq. 1. Indomethacin (COX-2 inhibitor) and Celecoxib (COX-1 inhibitor) were used as reference drugs in this assay.
where AO = The total activity of the control (enzyme), AT = The activity of the tested compounds.
2.4 LOX assay
All synthetic compounds were screened against 5-LOX at a concentration of 100 μM and compared with Zileuton as a reference drug. The assay was carried out as described by Pufahl et al. [21] with minor modifications. Basically, each well contained a mixture at a volume of 50 μL including buffer (50 mM Tris, 2 nM EDTA, 2 mM CaCI2, pH 7.5), human recombinant 5-LOX (0.1 U, Merck), 2’,7’-dichlorofluorescin diacetate (H2DCFDA, 10 μM), tested compounds (100 μM), AA (3 μM, Cayman Chemicals) and adenosine triphosphate (ATP, 10 μM, Sigma-Aldrich). In brief, H2DCFDA and 5-LO were incubated for 5 min before the addition of the tested compounds. This was followed by a second incubation with the tested compounds for 10 min. The reaction was initiated by the addition of a substrate solution containing AA and ATP. To measure the total activity of 5-LOX, the compounds/inhibitors were replaced with DMSO. Then, the reaction was measured using a POLARstar Omega (BMG LabTech GmbH, Ortenberg, Germany) microplate reader with fluorescence signal at excitation and emission wavelengths of 485 and 520 nm, respectively. The signals were measured at 0 and 60 min and the percentage of inhibition was calculated as in Eq. 2. All the steps were carried out at room temperature.
where ∆ABS Compounds = The difference of absorbance of the compounds/drug at the time interval, ∆ABS Control = The difference of absorbance of the enzyme at the time interval.
2.5 Molecular docking
Docking studies were performed for the most active compounds which had the potential to be a dual inhibitor of COX-2/5-LOX in the binding site of COX-2 and 5-LOX enzymes using GLIDE software (Schrödinger, LLC, Oregon, USA) [22]. The docking study was performed to evaluate a possible binding mode of active compounds in the COX and 5-LOX binding sites. The crystal structure of human COX-1, human COX-2, and human 5-LOX with PDB ID of 6Y3C, 5F19, and 3O8Y, respectively, were retrieved from the Protein Database Bank (PDB) (https://www.rcsb.org/pdb/home/home.do) [23]. 5F19 was chosen to represent COX-2 in this study because it has the highest resolution number (2.04 Å). 5F19 was found to bind to aspirin and it inhibits COX-2 by acetylation of Ser530. Therefore, the modified residue (OAS) at position 530 was changed to its mother residue (SER) prior to molecular docking.
All protein structures were prepared using the Protein Preparation Wizard (Schrödinger, LLC, Oregon, USA).Footnote 2 The water molecules were removed, all hydrogens were added, and their bond orders were assigned. Molecular docking study was performed in extra precision (XP) mode of GLIDE and the grid generation was generated at the centre of selected amino acid residues with a grid spacing of 12 Å [24]. For the COX enzyme (COX-1 and COX-2), the amino acid residues that had been selected were His90, Arg120, Gln192, Tyr355, Tyr385, Trp387, Val423, Arg513, Phe518, Val523, and Ser530. Meanwhile, for 5-LOX, the amino acid residues were based on a description by Gilbert et al. where the grid box was set at the coordinates of X = − 2.7017, Y = 24.5204, and Z = 0.4547 within 14 Å radius [25].
3 Results
3.1 Drug-likeness properties
The drug-likeness properties including the solubility of the tested compound and the number of rotatable bonds are presented in Table 2. Based on the predicted values, MW of the compounds are between 256 and 363, and the log P values of the compounds are in the range of 2.337–4.538. The HBD and HBA are < 2 and < 5, respectively. Thus, all in-house datasets of compounds obeyed the Lipinski’s Rule of Five with the number of rotatable bonds less than 8. These indicated that the synthetic compounds are druggable and suitable to be administered as oral drugs.
3.2 Dual-inhibitory activity of cyclooxygenase and lipoxygenase
The ability of the synthesized compounds to inhibit COX-2 and 5-LOX was evaluated using a biological assay as shown in Fig. 1 and Table 3, respectively. All compounds were tested for COX-1 and COX-2 inhibition at high concentration (100 µM) to determine the percentage of inhibitory activity and selectivity of tested compounds (percentage of COX-2 inhibition/percentage of COX-1 inhibition). Eight (8) tested compounds were found to have a higher selectivity for COX-2 over COX-1 compared to Indomethacin and slightly inhibited 5-LOX. These compounds were NP6, NP9, NPC4, NPC5, NPC6, NPC7, NPC8, and NC2. However, the results revealed that none of the tested compounds were able to exhibit inhibitory activity similar to or higher than Celecoxib. Among the compounds, NPC7 showed the most promising COX-2 inhibitory activity with a selectivity index towards COX-2 at 5.661.

Inhibitory activities of COX and selectivity index (SI) for COX-2 of the tested compounds
For 5-LOX activity, the compounds were evaluated using a high dose concentration (100 µM) but the majority of the compounds exhibited less than 40% of inhibitory activities. Only compound NF4, NF5, NC1, NC5, NC6, and NC7 did not show any inhibitory activity towards 5-LOX. Meanwhile, nine (9) compounds known as NPC5, NPC6, NPC7, NPC8, NP4, NP6, NP7, NP9, and NF7 showed more than 10% inhibition. Six (6) compounds known as NPC3, NPC4, NP5, NF2, NC2, and NC8 were shown to have less than 10% of 5-LOX inhibitory activity. Thus, it can be summarized that NPC6 and NPC7 are potential dual inhibitors of COX-2 and 5-LOX. It is also worth revealing that these tested compounds are more selective against COX-2 with less than 50% inhibitory activity against COX-1.
3.3 Molecular docking study
The binding interactions of NPC6 and NPC7 in the binding pockets of COX-2 and 5-LOX were further analysed through docking studies. Figure 2 shows the preferred binding poses of both compounds in the catalytic active site of COX-2 and 5-LOX. Noteworthy, both compounds exhibited different binding positions as Celecoxib in COX-2 but similar binding mode as Zileuton in 5-LOX. Additionally, in COX-2, the orientation of these compounds was opposite to each other where chromanone of NPC6 protruded near the opening site of COX-2 while chromanone of NPC7 buried inside the hydrophobic side pocket of COX-2.

Binding mode of the compounds inside the active site. a COX-2, b 5-LOX. Celecoxib (turquoise), Zileuton (green), NPC6 (blue) and NPC7 (red) are represented in stick form. Ferum is depicted as sphere
Based on the docking results (Table 4), Celecoxib had the highest number of hydrogen bonds with three hydrogen bonds while only one hydrogen bond was observed between NPC7 and the binding site of COX-2. In contrast, no hydrogen bond was formed in the binding pocket of COX-2 for NPC6. Besides, the highest selectivity of Celecoxib against COX-2 was contributed by the π-cation interaction with Arg120. For both tested compounds, there was also π–π interaction with Tyr355. The position of Celecoxib and the tested compounds inside the catalytic site of COX-2 was stabilized by surrounded hydrophobic interactions.
In the case of 5-LOX (Table 5), there were two hydrogen bonds were established between Zileuton and interacting amino acids of 5-LOX. In contrast, NPC7 formed only one hydrogen bonds between its chromanone group and Ala424. Similarly, NPC6 created a hydrogen bond in the active site of 5-LOX but it was stabilized by surrounded hydrophobic and polar interactions which contributed to a slightly higher 5-LOX inhibition as compared to NPC7.
4 Discussion
Understanding the structure of COX and differentiating between COX isoforms are necessary in designing novel and selective COX inhibitors. Active site of COX is comprised of long hydrophobic channel with a narrow entrance at the membrane-binding domain. Even though the active site is highly similar, their binding cavity is different where COX-2 has a larger binding cavity than COX-1. This lead to rooms for the development of selective COX inhibitors [26]. The active site of COX is divided into three regions: entrance of the active site which comprised of Arg120 and Tyr355; hydrophobic pocket which located beneath the haem group; and side pocket which determined the selectivity of COX inhibitors. The entrance of the active site and the hydrophobic pocket are highly conserved regions, but the side pocket is a non-conserved region where a few amino acid residues are different and give rise to extra pocket to COX-2.
Both active sites of COX-1 and COX-2 are very similar but in COX-2, there is side pocket which is located above the entrance of COX binding site (Arg120 and Tyr355) [27]. Further, a crystal structure analysis of the non-conserved region identified that the side pocket in COX-2 is raised due to a single amino acid residue which is different at position 523; near to Arg120, and it is important for the selectivity of the drugs. The changes of a single methyl group of isoleucine (Ile) in COX-1 to valine (Val) in COX-2 at position 523 leaves a space in the lining of the binding site and allows drugs or inhibitors to access the side pocket [27, 28]. The accessibility of the side pocket in COX-2 allows the additional interactions with Arg513 (substitution of His513 in COX-1) which contributes to COX-2 selectivity [29]. The orientation of Leu384 also contributes to the selectivity of COX-2. The presence of Phe503 causes the Leu384 side chain to point into the active site in COX-1. Meanwhile, in COX-2, the small size of Leu503 allows Leu384 to move away from the active site and increases accessible space in COX-2 binding site [30].
For this study, both the crystal structures of human COX-1 and COX-2 are required, and they had been prepared using the Protein Preparation Wizards tool in the Schrödinger package before proceeding to molecular docking. The COX-1 protein structure is necessary for this study to identify the compounds that are selective toward COX-2 protein. Previous studies had used homology modelling for COX-1 since no crystal structure of human COX-1 is available in the PDB database [31, 32]. However, there was an apoprotein of human COX-1 deposited in February 2020 [33]. Thus, the study performed docking analysis using this crystal structure (PDB ID: 6Y3C).
In the meantime, human COX-2 crystal structure is only available in PDB starting from March 2016. Most of COX-2 previous studies used murine crystal structure (PDB ID: 3LN1/1CX2/3NT1/6COX) because its binding site is similar to the human COX-2 (except for 5KIR) and the differences are not in the binding site [27, 34]. In the case of PDB ID: 5KIR where Rofecoxib is bound to the human COX-2, the crystal structure of COX-2 has approximately 20% larger unit cell volume and this is due to the different method of crystallization of COX-2 used [34]. Meanwhile, there were also some studies that used a homology model to build a crystal structure of the human COX-2, especially before crystal structures of the human COX-2 were deposited in the PDB [35].
LOX is another pathway that is involved in the AA cascade. It is a non-haem iron-containing enzyme that is involved in the conversion of AA into 5-HPETE, which is responsible for the production of LTB4 and cysteinyl leukotrienes. The 5-LOX binding site consists of a deep bent-shaped which is mostly occupied by hydrophobic amino acids. The 5-LOX binding site was reported to extend from Phe177 and Tyr181 in the upper part of the cleft to Trp599 and Leu420 at the bottom of the cleft [36]. Besides that, the entrance of 5-LOX consists of Lys409, a polar amino acid, and the study also claimed that several amino acid residues such as Tyr181, Leu414, Asn425, Arg411, and Phe421 are important residues for 5-LOX interaction.
Based on the biological assay, NPC6 and NPC7 displayed significant dual COX-2/5-LOX inhibitory activity and were chosen to proceed for docking studies. These compounds were docked into the COX-1, COX-2, and 5-LOX active sites using the GLIDE software. These compounds were found to be “preferential COX-2 inhibitors” compared to “highly selective COX-2 inhibitors”. This is because the inhibitory activities and the binding energy for COX activities were less than Celecoxib. These findings were in agreement with the other “preferential COX-2 inhibitor” such as meloxicam, etodolac, and nimesulide [37, 38].
Through docking studies, compounds NPC6 and NPC7 possessed different binding pose in comparison to Celecoxib where the compounds mostly occupied the hydrophobic region in COX-2 binding site instead of the side pocket region. This is due to the different chemistry scaffold between Celecoxib and these derivatives [39]. Besides that, the tested compounds displayed higher selectivity to COX-2 in docking scores and it was in agreement with experimental enzyme assay. This also had been observed in 5-LOX inhibitory activity where the result of the enzyme inhibition and the docking score were in agreement.
COX-2 selectivity of these active compounds is attributed to their (i) hydrogen bond interactions with key amino acids such as Ser530 in the COX catalytic domain (active site); and (ii) π–π interaction between ring B of flavanone moiety with Tyr355 at the entrance of COX binding site. The binding had been strengthened through hydrophobic interaction at the hydrophobic region and lobby region of the COX binding site of the COX enzyme. For 5-LOX inhibition, the activity of NPC6 and NPC7 was contributed by a hydrogen bond interaction with Ala424. For comparison, Zileuton formed hydrogen bonding interactions with Asn425 and His600. On the other hand, hydrophobics and polar interactions were the main contributors for the activity of NPC6 and NPC7 with Tyr181, Leu 414 and Phe421 are important residues in the binding interactions of 5-LOX.
5 Conclusion
The present work described the potential of chalcone, flavone and flavanone derivatives as dual COX-2/5-LOX inhibitors. While eight (8) compounds possessed proportionate COX-2 inhibitory activity as Celecoxib, none of them had higher selectivity index to COX-2 than Celecoxib. However, these compounds showed inhibitory activity against 5-LOX where they exhibited up to 40% of inhibitory activity at high concentration (100 µM). On top of that, NPC6 and NPC7 have displayed dual COX-2/5-LOX inhibitory activity. The results were consistent with docking studies of these compounds in COX and 5-LOX binding site. Furthermore, most of the compounds occupied the hydrophobic region and extend to the lobby region near the entrance of COX binding site, whereas structural modifications are needed by extending the substitutions at para-position of benzene ring to increase the selectivity towards 5-LOX. Overall, the scaffolds of these chemical structures have potential to further optimise as lead compounds for the dual inhibition of COX-2 and 5-LOX.
Availability of data and materials
Not applicable.
Notes
QikProp. Schrödinger Release 2017–1: QikProp. 2017.
Protein Preparation Wizard. Schrödinger Suite 2017–1: Protein Preparation Wizard. 2017.
Abbreviations
- COX-2:
-
Cyclooxygenase-2
- 5-LOX:
-
5-Lipoxygenase
- AA:
-
Arachidonic acids
- PLA2:
-
Phospholipase A2
- PGG2 :
-
Prostaglandin G2
- TXA2 :
-
Thromboxane A2
- 5-HPETE:
-
5-Hydroperoxyeicosatetratenoic acid
- LT:
-
Leukotrienes
- NSAIDs:
-
Non-steroidal anti-inflammatory drugs
- tNSAIDs:
-
Traditional NSAIDs
- IOS:
-
Institute of Science
- TMPD:
-
N,N,N’,N’-tetramethyl-p-phenylenediamine
- DMSO:
-
Dimethyl sulphoxide
- H2DCFDA:
-
2’,7’-Dichlorofluorescin diacetate
- ATP:
-
Adenosine triphosphate
- PDB:
-
Protein database bank
- XP:
-
Extra precision mode
References
Wang B, Wu L, Chen J et al (2021) Metabolism pathways of arachidonic acids: mechanisms and potential therapeutic targets. Signal Transduct Target Ther 6(1):94
Hofer M, Hoferová Z, Falk M (2019) Brief story on prostaglandins, inhibitors of their synthesis, hematopoiesis and acute radiation syndrome. Molecules 24(22):4019
Qureshi O, Dua A (2022) COX inhibitors. StatPearls Publishing, Treasure Island
Wisastra R, Dekker FJ (2014) Inflammation, cancer and oxidative lipoxygenase activity are intimately linked. Cancers (Basel) 6:1500–1521
Kazani S, Planaguma A, Ono E et al (2013) Exhaled breath condensate eicosanoid levels associate with asthma and its severity. J Allergy Clin Immunol 132:547–553
Dixit N, Wu DJ, Belgacem YH et al (2014) Leukotriene B4 activates intracellular calcium and augments human osteoclastogenesis. Arthritis Res Ther 16:496
Lin HC, Lin TH, Wu MY et al (2014) 5-lipoxygenase inhibitors attenuate TNF-α-Induced inflammation in human synovial fibroblasts. PLoS ONE 9:e107890
Bouchareychas L, Grössinger EM, Kang M et al (2017) Critical role of LTB 4/BLT1 in IL-23–induced synovial inflammation and osteoclastogenesis via NF-κB. J Immunol 198:452–460
Praveen Rao PN, Knaus EE (2008) Evolution of nonsteroidal anti-inflammatory drugs (NSAIDs): cyclooxygenase (COX) inhibition and beyond. J Pharm Pharm Sci 11:81s–110s
Crofford LJ (2013) Use of NSAIDs in treating patients with arthritis. Arthritis Res Ther 15:1–10
Jaismy Jacob P, Manju SL, Ethiraj KR et al (2018) Safer anti-inflammatory therapy through dual COX-2/5-LOX inhibitors: a structure-based approach. Eur J Pharm Sci 121:356–381
Hanáková Z, Hošek J, Kutil Z et al (2017) Anti-inflammatory activity of natural geranylated flavonoids: cyclooxygenase and lipoxygenase inhibitory properties and proteomic analysis. J Nat Prod 80:999–1006
Ibrahim SRM, Mohamed GA, Alshali KZ et al (2018) Lipoxygenase inhibitors flavonoids from Cyperus rotundus aerial parts. Braz J Pharmacogn 28:320–324
Feng W, Hao Z, Li M (2017) Isolation and structure identification of flavonoids. In: Justino GC (ed) Flavonoids. IntechOpen, Rijeka, p 2
Jiang X, Yu J, Zhou Z et al (2019) Molecular design opportunities presented by solvent-exposed regions of target proteins. Med Res Rev 39:2194–2238
Kondhare DD, Gyananath G, Tamboli Y et al (2017) An efficient synthesis of flavanones and their docking studies with aldose reductase. Med Chem Res 26:987–998
Yusof MIM, Salleh MZ, Kek TL et al (2013) Activity-guided isolation of bioactive constituents with antinociceptive activity from Muntingia calabura L. leaves using the formalin test. Evid Based Complement Altern Med 2013:1–27
Nyokat N (2019) Phytochemical studies and biological evaluation of Artocarpus odoratissimus and Artocarpus sarawakensis, and some synthesis of pinostrobin pinocembrin derivatives. Universiti Teknologi MARA, Thesis
Shenvi S, Kumar K, Hatti KS et al (2013) Synthesis, anticancer and antioxidant activities of 2,4,5-trimethoxy chalcones and analogues from asaronaldehyde: structure–activity relationship. Eur J Med Chem 62:435–442
Vernieri E, Gomez-Monterrey I, Milite C et al (2013) Design, synthesis, and evaluation of new tripeptides as COX-2 inhibitors. J Amino Acids 2013:606282
Pufahl RA, Kasten TP, Hills R et al (2007) Development of a fluorescence-based enzyme assay of human 5-lipoxygenase. Anal Biochem 364:204–212
Friesner RA, Banks JL, Murphy RB et al (2004) Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J Med Chem 47:1739–1749
Berman HM, Westbrook J, Feng Z et al (2000) The protein data bank. Nucleic Acids Res 28:235–242
Friesner RA, Murphy RB, Repasky MP et al (2006) Extra precision glide: docking and scoring incorporating a model of hydrophobic enclosure for protein-ligand complexes. J Med Chem 49:6177–6196
Gilbert NC, Bartlett SG, Waight MT et al (2011) The structure of human 5-lipoxygenase. Science 331(6014):217–219
Simmons DL, Botting RM, Hla T (2004) Cyclooxygenase isozymes: the biology of prostaglandin synthesis and inhibition. Pharmacol Rev 56:387–437
Kurumbail RG, Stevens AM, Gierse JK et al (1996) Structural basis for selective inhibition of cyclooxygenase-2 by anti-inflammatory agents. Nature 384:644–648
Gierse JK, McDonald JJ, Hauser SD et al (1996) A single amino acid difference between cyclooxygenase-1 (COX-1) and -2 (COX-2) reverses the selectivity of COX-2 specific inhibitors. J Biol Chem 271:15810–15814
Wong E, Bayly C, Waterman HL et al (1997) Conversion of prostaglandin G/H synthase-1 into an enzyme sensitive to PGHS-2-selective inhibitors by a double His513 → Arg and Ile523 → Val mutation. J Biol Chem 272:9280–9286
Bayly CI, Black WC, Léger S et al (1999) Structure-based design of COX-2 selectivity into flurbiprofen. Bioorg Med Chem Lett 9:307–312
Fiser A (2010) Template-based protein structure modelling. Methods Mol Biol 673:73–94
Md Idris MH, Mohd Amin SN, Selvaraj M et al (2018) High-throughput structure-based drug design of chalcones scaffolds as dual inhibitor of cyclooxygenase-2 and microsomal prostaglandin E synthase-1. J Pharm Sci Emerg Drugs 6:1–14
Miciaccia M, Belviso BD, Iaselli M et al (2021) Three-dimensional structure of human cyclooxygenase (hCOX)-1. Sci Rep 11:4312
Orlando BJ, Malkowski MG (2016) Crystal structure of rofecoxib bound to human cyclooxygenase-2. Acta Crystallogr Sect Struct Biol Commun 72:772–776
Dhanjal JK, Sreenidhi AK, Bafna K et al (2015) Computational structure-based de novo design of hypothetical inhibitors against the anti-inflammatory target COX-2. PLoS ONE 10:e0134691
Charlier C, Hénichart JP, Durant F et al (2006) Structural insights into human 5-lipoxygenase inhibition: combined ligand-based and target-based approach. J Med Chem 49:186–195
Suleyman H, Cadirci E, Albayrak A et al (2008) Nimesulide is a selective COX-2 inhibitory, atypical non-steroidal anti-inflammatory drug. Curr Med Chem 15:278–283
Xu S, Hermanson DJ, Banerjee S et al (2014) Oxicams bind in a novel mode to the cyclooxygenase active site via a two-water-mediated H-bonding Network. J Biol Chem 289:6799–6808
Boittier ED, Tang YY, Buckley ME et al (2020) Assessing molecular docking tools to guide targeted drug discovery of CD38 inhibitors. Int J Mol Sci 21:5183
Acknowledgements
The authors would like to thank the Institute of Science (IOS), UiTM for providing the synthesized compounds. All the compounds were submitted for application of patent (Application Number: PI 2018704123).
Funding
The research was supported by Ministry of Education Malaysia through the Transdisciplinary Research Grant Scheme (Grant No.: 600-RMI/TRGS 5/3 (1/2014)) and Integrative Pharmacogenomics Institute (iPROMISE), UiTM (Grant No.: 241910/2015/MZS/7).
Author information
Authors and Affiliations
Contributions
SNMA and SNMA conducted the experiments and analysed the data. MHMI interpreted the data and produced the manuscript. NN carried out the synthesis. MS clarified the computational modelling results. HYK, ZS and ASH guided the synthesis and reviewed the manuscript. ZAZ, LKT and MZS designed the study and reviewed the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests in this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Md Idris, M.H., Mohd Amin, S.N., Mohd Amin, S.N. et al. Flavonoids as dual inhibitors of cyclooxygenase-2 (COX-2) and 5-lipoxygenase (5-LOX): molecular docking and in vitro studies. Beni-Suef Univ J Basic Appl Sci 11, 117 (2022). https://doi.org/10.1186/s43088-022-00296-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43088-022-00296-y