
Abstract
Backgrounds
Plant-plant interactions are among the most important factors affecting the natural recovery of vegetation. While the impacts of nurse plants on species composition and biodiversity are well documented, the effects of different nurse’s growth forms on all biodiversity components including taxonomic, functional, and phylogenetic diversity have been less studied and compared, especially for their effects on different times after fire disturbance. This research was focused on comparing the effects of a perennial grass (Elymus hispidens), a perennial herb (Phlomis cancellata), and a high shrub species (Lonicera nummulariifolia) on species composition and the biodiversity components, and how these impacts change across five sites with short-term (1 and 4 years sites), long-term (10 and 20 years sites) times since last fire and a control site where no fire was known in recorded history in semi-arid shrublands of Fereizi Chenaran located in Northeast of Iran. The changes of species composition and taxonomic, functional, and phylogenetic diversity were calculated with respect to the presence/absence of nurse’s growth forms, fire history, and their interactions.
Results
Nurse shrubs affected species composition and all biodiversity components, whereas all indices were reduced when considering Elymus grass as nurse plant. On the other hand, the herb Phlomis enhanced species composition and taxonomic diversity, while it had a negative effect on functional and phylogenetic diversity. Such specific effects of nurse types were mostly observed under long timescales (i.e., 10- and 20-year sites). Interestingly, the relative importance of nurse types and time since the last fire largely explained the variation of species composition and biodiversity components, with larger effects of nurse types on all biodiversity components. However, we found a significant contribution of fire explaining variation of species composition and phylogenetic diversity.
Conclusions
These results indicated nurse plants can affect the post-fire recovery of vegetation by providing specific mechanisms controlling beneficiary relatedness depending on their growth forms and time scales since the last fire. Therefore, these findings suggest perennial plants in the form of nurse species as a useful factor to develop techniques of active restoration in burned ecosystems.
Resumen
Antecedentes
Las interacciones planta-planta están entre los factores más importantes que afectan la recuperación natural de la vegetación. Mientras que los impactos de las plantas nodrizas en la composición de especies y biodiversidad han sido bien documentadas, los efectos de las formas de crecimiento de plantas nodrizas en todos los componentes de la biodiversidad, incluyendo diversidad taxonómica, funcional y filogenética ha sido menos estudiada y comparada, en especial por sus diferentes efectos en diferentes períodos de tiempo luego de disturbios como el fuego. Esta investigación se enfocó en comparar los efectos de un pasto perenne (Elymus hispidens), una hierba perenne (Phlomis cancellata), y una especie de arbusto alto (Lonicera nummulariifolia) en la composición de especies y en los componentes de la diversidad, y sobre cómo esos impactos cambian a lo largo de cinco sitios con períodos cortos (sitios con 1 a 4 años), períodos largos (sitios con 10 y 20 años) después del último fuego, y un sitio control en el cual no había registros históricos de incendios, en arbustales semiáridos de Fereizi Chenaran, un área ubicada en el noreste de Irán. Los cambios en la composición de especies y en la diversidad taxonómica, funcional y filogenética fueron calculados en relación a la presencia/ausencia de las formas de crecimiento de las plantas nodriza, la historia del fuego y sus interacciones.
Resultados
Las plantas nodriza afectaron la composición de especies y todos los componentes de la biodiversidad, mientras que todos los índices fueron reducidos cuando se consideró al pasto Elymus como planta nodriza. Por otra parte, la hierba Phlomis aumentó la composición de especies y la diversidad taxonómica, mientras que tuvo un efecto negativo en la diversidad funcional y filogenética. Los efectos específicos de los tipos de nodriza se observaron mayoritariamente en los sitios con escalas de tiempo de ocurrencia del fuego más antiguas (i.e., 10 a 20 años desde el último incendio). De manera interesante, la importancia relativa de los tipos de nodriza y el tiempo desde el último incendio explican mayoritariamente la variación en la composición de especies y los componentes de la biodiversidad, con efectos más amplios de tipos de nodrizas en todos los componentes de la biodiversidad. De todas maneras, encontramos una contribución del fuego para explicar la variación en la composición de especies y la diversidad filogenética.
Conclusiones
Estos resultados indican que las plantas nodrizas pueden afectar la recuperación post-fuego de la vegetación mediante la provisión de mecanismos específicos que controlan la relación benéfica dependiente de sus formas de crecimiento y la escala de tiempo desde el último fuego. Por lo tanto, estos resultados sugieren a las plantas perennes en la forma de especies nodriza como un factor útil para desarrollar técnicas de restauración activa en ecosistemas quemados.
Similar content being viewed by others

Background
Wildfires are major natural disturbances impacting plant communities and ecosystems (Bowd et al. 2018). A large body of evidence suggests that fire has pronounced effects on plant biodiversity and the functioning of terrestrial ecosystems (Ghasempour et al. 2022; Omidipour et al. 2021; Bowman et al. 2016; Hurteau et al. 2014; Pausas and Ribeiro 2013). For instance, fire can significantly change taxonomic composition and promote functional/phylogenetic divergence or convergence (Gómez-González et al. 2011; Hernandez-Serrano´ et al. 2013) through diversification of plant lineages (Bytebier et al. 2011; He et al. 2012; 2019). However, such important impacts of fire on vegetation are closely related to different time scales (Keeley et al. 2012; Gosper et al. 2013), presence of dominant perennial plants (Padilla and Pugnaire 2006), and ecosystem type (McLauchlan et al. 2020; He et al. 2019). The complex relationships between these important factors have posed challenges for fire ecologists in formulating clear theories and hypotheses regarding post-fire vegetation recovery (Mucina et al. 2019). Therefore, the evidence suggesting key factors and mechanisms that promote natural post-fire vegetation recovery has been widely embraced in fire ecology, fostering ecological knowledge and facilitating the development of active restoration practices.
Natural vegetation recovery is a lengthy process that occurs without support and has significant impacts on biodiversity, ecosystem functions, and services in degraded ecosystems (Sandell Festin et al. 2019; Bashirzadeh et al. 2022b; Wortley et al. 2013; Wilgan et al. 2011). In this context, plant biodiversity and composition have been identified as crucial indicators of the natural recovery of vegetation (Wortley et al. 2013; Badano and Cavieres 2006). Understanding key factors and mechanisms of this process through plant biodiversity and species composition can potentially inform the development of active restoration practices in burned ecosystems (Padilla and Pugnaire 2006). However, plant ecologists face major challenges when considering plant biodiversity, which encompasses various components such as taxonomic, functional, and phylogenetic diversity (Swenson et al. 2006; Cadotte and Davies 2016). Making connections between the results of these components and their interpretations can be difficult. Therefore, numerous studies in fire ecology have focused only on the floristic composition and taxonomic component (Bahalkeh et al. 2021), while less attention has been given to other components such as functional diversity (Abedi et al. 2022) and phylogenetic diversity (Bashirzadeh et al. 2023; Hipp et al. 2015; Keeley and Fotheringham 2003). It has been shown that natural vegetation recovery is observable not only in variations of floristic composition, but also in functional traits and evolutionary relatedness of plant species within communities (Keeley et al. 2012; Dantas et al. 2013). Thus, the overall outcomes of post-fire vegetation recovery are significantly dependent on biodiversity components.
Natural post-fire vegetation recovery is influenced by several major factors such as the presence of perennial plants (Ren et al. 2008; Niknam et al. 2018), spatial scale considered (Gomez-Aparicio 2009), and time scales since the last fire occurrence (He et al. 2019). A wide range of studies have recognized the positive effects of perennial plants, acting as nurse plants, on vegetation structure and soil seed bank in severe ecosystems such as alpine environments and global drylands (Bashirzadeh et al. 2022a; Erfanzadeh et al. 2022; Pashirzad et al. 2019a; Pashirzad et al. 2019b; Soliveres et al. 2015; Soliveres et al. 2014; Soliveres and Maestre 2014; He et al. 2012; Gomez-Gonz ´ alez ´ et al. 2011) and degraded ones (Gomez-Aparicio 2009; Padilla and Pugnaire 2006; Bashirzadeh et al. 2022b). The findings of such studies have provided valuable insights into natural vegetation recovery (Bond et al. 2005; Calitz et al. 2015; Bashirzadeh et al. 2022b), and have contributed to the development of restoration practices (Padilla and Pugnaire 2006; Ren et al. 2008). These findings and theories have primarily defined the impacts of nurse plants on the recruitment and survival of less tolerant plant species by creating suitable microenvironments under their canopies.
As a result, they contribute to increasing the taxonomic diversity of the community (e.g., Soliveres and Maestre 2014; Le Bagousse-Pinguet et al. 2019; Madrigal-Gonzalez et al. 2020). However, the specific effects of nurse species are not solely limited to adding new species and increasing the species pool. Differences in functional traits of plant species have been observed between plant species that grow under nurse species and those that can thrive in their absence (Rafiee et al. 2023; Soliveres et al. 2014). Furthermore, the maintenance of such traits across phylogeny has been observed in plant communities in relation to the presence of nurse species (Keeley et al. 2016; 2012; Abedi et al. 2022, He et al. 2019). However, our understanding of the mechanisms promoted by nurse species in the recovery of natural vegetation in burned ecosystems, particularly in relation to fires, still has major gaps. This is because, in addition to the general effects of fire as a major disturbance factor on vegetation, there are also specific effects of fire that mediate facilitative plant interactions in burned ecosystems (Vega-Alvarez et al. 2019; Keeley et al. 2012). These fire effects may activate adaptive mechanisms that promote natural recovery of vegetation by increasing the distribution of traits related to post-fire recruitment and growth strategies (Alam et al. 2020; Lavorel et al. 2007; Pausas et al. 2003).
Interestingly, the impacts of nurse species on vegetation recovery in disturbed ecosystems can vary depending on their growth forms, study scale considered, and time intervals after disturbance (Gomez-Aparicio 2009; Erfanzadeh et al. 2014; He et al. 2012). In the case of alpine environments, there is strong evidence that shrubs have the most important effects on taxonomic and phylogenetic diversity (Butterfield et al. 2013; Cavieres et al. 2013; Camac et al. 2013; Svriz et al. 2013). On the other hand, in arid ecosystems, nurse grasses can strongly affect functional diversity by providing suitable microhabitats for beneficiary species with distant relatedness (Gastauer et al. 2018; Navarro-Cano et al. 2018; Bashirzadeh et al. 2022b). However, such effects of nurse growth forms are highly scale-dependent and should be considered across the scale of vegetation patches to the composition of the entire community, where the most effective theories and empirical perspectives in plant ecology are structured (Soliveres et al. 2015). Additionally, besides considering nurse growth forms and scale, the time interval since the last likely fire influences vegetation recovery in burned ecosystems (Paniw et al. 2017). In this context, species diversity is expected to recover significantly over time following a fire, as the time since the last fire increases (Abedi et al. 2022; Bashirzadeh et al. 2023). Herath et al. (2009) found that nearly all plant species in fire-prone ecosystems that were present in the pre-fire communities, were evident for 12 months after the fire. This phenomenon is consistent with the “autosuccession” model (Hanes 1971) and the “initial floristic composition” model (Enright et al. 2007) of community succession (He et al. 2019). However, it is not clear which forms of nurse growth and at what intervals contribute to this phenomenon in burned ecosystems. Therefore, further research studies are needed to clarify the relative impacts of nurse types on the natural recovery of all biodiversity components considering times scales and the proper scale of burned ecosystems.
Here, we analyzed the effects of three dominant perennial plants with different growth forms (Elymus hispidus subsp. hispidus, Phlomis cancellata, and Lonicera nummularifolia with perennial grass, perennial forb, and sprawling high shrub growth forms, respectively) on species composition and taxonomic, functional and phylogenetic diversity, over different time periods since the last fire (1, 4, 10, and 20 years since the last fire), and compared these to a control site, where no fire was known in recorded history. This research study was conducted in semi-arid scrublands of the Fereizi region in northeastern Iran, which can serve as an empirical example of post-fire vegetation recovery in scrublands. In addition, previous studies only addressed either the effects of fire in terms of fire intensity (Abedi et al. 2022; Heydari et al. 2017) or the presence of nurse plants on taxonomic diversity and species composition at point basis scales (i.e., vegetation patch scales) (Abedi et al. 2022; Bahalkeh et al. 2021; Paniw et al. 2017). Therefore, some important issues such as how nurse species affect all biodiversity components (i.e., taxonomic, functional, and phylogenetic diversity), which mechanisms are used to facilitate their natural recovery, and relevant contributions of the nurse species across time scales in burned ecosystems have remained unclear. In this regard, it is of great importance to consider some key functional characteristics of nurse species, including growth forms, when assessing their impacts in order to fill the gaps in understanding of the natural vegetation recovery after fire, and to provide important insights for the development of active restoration practices. Therefore, we obtained information on nurse plant growth forms, functional trait values, and phylogenetic relationships among plant species, and addressed the following questions: (i) Does each biodiversity component exhibit significant variation in relation to nurse plant types across different times since the last fire? (ii) Are the specific effects of each nurse plant on biodiversity correlated to times since the last fire, the specific biodiversity facet considered, and functional groups of plant species? (iii) To what extent do nurse plant types, time since the last fire, and their interaction explain the variance in different biodiversity components and species composition?
Methods
Study area
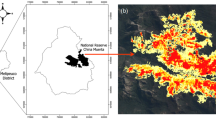
The study area consists of semi-arid shrublands of the Fereizi region located in northeastern Iran (between 36° 35′ and 36° 48′ N, and 58° 24′ and 58° 52′ E) (Bashirzadeh et al. 2023) (Fig. 1a). Elevation ranges from 1980 to 2323 m, generally increasing from south to north. Mean annual precipitation ranges from 285 to 312 mm with a cold semi-arid climate based on Emberger’s method (Additional file 1: Appendix S1). The rain season is from Autumn (late October) to early Spring, with the main rain season occurring from February through April, with an average temperature of 7.5° C (Memariani et al. 2009). Altitude,, and temperature values were obtained from the WorldClim dataset in 30″ resolution (Hijmans et al. 2005). Plant communities are typically dominated by herbaceous plants and shrubs (Bashirzadeh et al. 2023).

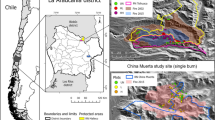
a Characteristics of the study sites across different times since last fire. Within each study site, two transects 200 m in length and approximately 500 m apart were established, and 20 plots each nurse were placed on each ones. We sampled vegetation beneath nurse individuals and nearby paired open areas. b Five study sites were surveyed in the Fereizi-Chenaran region including (a) unburned site (i.e. control site without fire occurrence), (b) 1-year sites (i.e. site with most recent fire occurring in the last year), (c) 4-year site (i.e. site with most recent fire occurring 4 years prior), (d) 10-year site and (e) 20-year site (i.e. sites with last fire occurring between 10 and 20 years prior)
The dominant plant species are typically Lonicera nummulariifolia Jaub. & Spach. (Caprifoliaceae), Elymus hispidens (Opiz) Melderis (Poaceae), Phlomis cancellata Bunge (Lamiaceae), Juniperus polycarpos K. Koch (Cupressaceae), Cotoneaster nummularius Lindl. (Rosaceae), Poa bulbosa L. (Poaceae), Taeniatherum caput-medusae (L.) Nevski (Poaceae) and Astragalus verus Olivier. (Fabaceae). However, other plant species including Artemisia kopetdaghensis (Poljakov) Y.R.Ling (Asteraceae), Artemisia turkomanica Gand. (Asteraceae), Acantholimon peterostegium Bunge (Plumbaginaceae), Acanthophyllum glandolusum Bunge ex Boiss. (Caryophyllaceae), Salvia abrotanoides (Kar.) Sytsma (Lamiaceae) and Rosa beggeriana Schrenk (Rosaceae) were frequently found in the study area (Bashirzadeh et al. 2023; Memariani et al. 2009). Our analyses focused on the three most dominant plant species as nurse plants, L. nummulariifolia, P. cancellata, and E. hispidens, which occurred at all sample sites. L. nummulariifolia is a sprawling high shrub species (plant height: 3–5 m) with a wide crown (Barzegaran et al. 2022). Especially under moderate disturbance conditions, it became clear that such broad-crowned shrubs have a positive effect on understory plant species (Jankju and Ejtehadi 2016; Soliveres and Maestre 2014; Soliveres et al. 2012). Sprawling shrubs generally create suitable microenvironments for understory species by increasing soil moisture and reducing radiation and air temperature compared to open areas. P. cancellata was considered as a perennial herbaceous plant based on Raunkiaer’s system of classification, previous studies conducted in northeastern Iran (Erfanian et al. 2021; Atashgahi et al. 2018; Memariani et al. 2016), and also according to our assessments of this plant species in the study region although it was highlighted as a sub-scrub plant in a reference (https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:453832-1). We included this plant species as a nurse plant in our further analyses because it promotes the growth, recruitment, and survival of other species (Soliveres et al. 2014; Jankju 2013; Jankju et al. 2008). E. hispidens is a perennial grass species that can grow up to 1.5 m tall, and its limited crown provides suitable habitat for some plant species with specific niches (Gomez-Aparicio 2009; 2004).
Data collection
Site selection
We selected five sites based on different times since the last fire in the study area. They included a control site where no fire was known in recorded history with natural vegetation, a 1-year site in which the last fire occurred in the last year (i.e., in 2021), a 4-year site, a 10-year site and a 20-year site, in which fires occurred 4, 10, and 20 years ago, respectively (Fig. 1a). These sites encompass similar ecological and topographical conditions, ensuring minimal variations in terms of climatic and topographical variables. These variables include annual precipitation and altitude, which range from 285 mm and 1980 m in the 4-year site to 312 mm and 2323 m in the control site (Additional file 1: Appendix S1). Furthermore, all the sites are located on the same geological substrate composed of limestone (Memariani et al. 2009). From all these sites, we extracted comparable data on altitude and precipitation from the WorldClim dataset (http://www.worldclim.org; Hijmans et al. 2005). To examine fire occurrence, we extracted the time since the last fire for each site from aerial photos and from information on fire time recorded by the staff of the environment department in the Fereizi region.
We estimated fire intensity for each site based on classifications from Heydari et al. (2017). Within these classifications, fire intensity is calculated using some specific indicators. Such specific indicators are (i) apparent fire; (ii) herbaceous cover; (iii) the percentage of shrubs, herbs, and trees killed; and (iv) stem charring of tree height (Heydari et al. 2017). In the 1-year plot, we found ground fires and burning lower tree branches. Herbaceous plants and some shrubs were killed. The stems of some plants were burned up to 70% of the whole total tree height. Additionally, between 25 and 30% of shrubs and herbs had been dried because of the fire (Fig. 1b). Such observations were also made at other burned sites by the staff working in the environment department at the Fereizi region. Finally, in the control site, no evidence of vegetation destroyed by fire was found, and no charring on tree stems was found as well. Based on such findings, all burned sites indicated an intermediate or moderate level of fire (Fig. 1b).
Vegetation sampling
To assess nurse plant effects on vegetation recovery across different times since the last fire, two transects in each site with 100 m length and approximately 500 m apart were established (Fig. 1a) on north-facing slopes, where similar ecological and topographical conditions were mostly found across transects. Then, on each transect, 20 individuals of each dominant plant species (i.e., L. nummulariifolia, E. hispidens, and P. cancellata) were randomly selected as potential nurses, and 20 1 m × 1 m quadrats were surveyed under the canopies of these dominant plant species, and the same number of quadrats in open areas far from such potential nurses. Then, the vegetation within the quadrats was surveyed as a percent coverage of present taxa from early May 2021 to late June 2021. To identify different microclimatic enhancement mechanisms of nurse species, it is essential to survey the vegetation beneath nurse plants and their open spaces using the same number and size of quadrats. This is because the measured plant biodiversity varies greatly depending on the sampling effort (Erfanzadeh et al. 2022; Pashirzad et al. 2019a; 2019b; Soliveres et al. 2012). Therefore, the number of transects and quadrats laid out was determined based on two main criteria: (i) the extent of study sites and (ii) species-area curves (Bashirzadeh et al. 2022b). We identified a total of 75 plant taxa in our study sites. To avoid the effects of rare species, we excluded 20 (out of the 75) with a frequency below five in the entire dataset (Bahalkeh et al. 2021) (see details in Additional file 2: Appendix S2 in Supporting Information).
Functional trait information
We assessed eight functional traits for plant species in our sites, including leaf dry matter content (LDMC) (g), specific leaf area (SLA) (m2 kg − 1), leaf nitrogen (mg g − 1) and carbon content (mg g − 1), plant height (m), seed mass (mg), growth form, and life span. These functional traits were introduced as key functional traits for plant species in burned ecosystems, and are dependent on biotic interactions and disturbance conditions (Rafiee et al. 2022; Lienin and Kleyer 2012; Lavorel et al. 2007; Meng et al. 2015; Abedi et al. 2022). For example, SLA and LDMC reflect plant growth rates in fire-prone ecosystems. Fire generally increases SLA and decreases LDMC because fire increases the abundance of fast-growing and ruderal species compared with perennial species (Davies et al. 2009; Rhodes et al. 2010). In addition, other traits related to plant regeneration including seed mass, plant height, and plant growth also represent plant response to fire (Cruz et al. 2022). Life span represents plant strategies and abilities including competitive or facilitative abilities, and leaf nitrogen/carbon content and life span can indicate post-fire resprouting in shrubs (Anacker et al. 2011; Sakschewski et al. 2015), and differentiate annual from perennials plant species in herbaceous communities (Keeley et al. 2012).
We obtained information on these plant functional traits by measuring some traits including plant height and life span in field and others from publicly available trait datasets (BIEN package in R (Maitner et al. 2018), TRY (Kattge et al. 2020), LEDA (Kleyer et al. 2008) and TR8. To measure plant height in the field, we selected 10 individuals of each plant species and measured plant height as the mean of these individuals, focusing on the vegetative part. This approach was chosen to minimize bias introduced by the reproductive parts, which can vary at different growth stages, particularly in grasses (Cavieres et al. 2013). For other functional traits, we averaged observations of plant species individuals to create a species mean trait value, when multiple measurements per species were available in these databases. For some plant species, we used genus-level means when either species-level data were not available or plants were identified only to genus level (Lamanna et al. 2014) (see Supporting Information Additional file 2: Appendix S2).
Phylogenetic information
We obtained a phylogeny of 55 plant species present in all study sites based on the most up-to-date megaphylogeny for seed plants. We standardized species names in our dataset according to The Plant List using the R package “Taxonstand” (Cayuela et al. 2012). Then, we used the R function V.PhyloMaker (Jin and Qian 2019) to link the species names in our dataset with those in the megaphylogeny, and the scenario 3 approach (Qian and Jin 2016), to add species to the phylogeny. This phylogenetic tree was then used as reference lists from which phylogenetic diversity could be calculated for our communities in the dataset.
Measures of taxonomic, functional, and phylogenetic diversity
To measure the effects of nurse’s growth forms on biodiversity components in different times since the last fire, we created some communities in relation to the presence of nurse species such as L. nummulariifolia, P. cancellata, and E. hispidens by randomly substituting 50% of the quadrats sampled in open microsites per 50 m transect, with the same number of randomly selected quadrats at the same transect beneath Lonicera, Phlomis, and Elymus areas, respectively. While the latter does not consider the relative proportion of nurse/open areas at each site, using this standard proportion allowed us to create different communities on transects (two communities on each transect) and compare our results across sites with different times since the last fire (Bashirzadeh et al. 2022a). In addition, this procedure allows us to understand the effects of nurse plants in communities, which is a field of plant ecology where robust theories and findings are often made. We achieve this by comparing communities that include nurse quadrats to exclusively open communities (i.e., communities without nurse effects) (Bashirzadeh et al. 2022a). In this context, we considered communities with the presence of L. nummulariifolia, P. cancellata, and E. hispidens as Lonicera, Phlomis, and Elymus communities and their absences as Open communities for further analyses (i.e., four communities per 50 m transect). Then, we measured biodiversity components for these communities in each site. To measure taxonomic diversity, we used the first two Hill numbers to estimate species richness (q = 0) and species diversity as the exponential of Shannon’s entropy (q = 1; referring to Shannon diversity) (Chao et al. 2014) for Lonicera, Phlomis, Elymus, and Open communities. The calculation was based on the percent coverage of plant taxa, which is less affected by differences in total sampling effort than other methods (Chao and Jost 2012).
To measure functional and phylogenetic diversity, we assessed abundance-weighted mean pairwise distance (MPD) (Tucker et al. 2017; Webb et al. 2002) as the most robust measure of the phylogenetic and functional relatedness for Lonicera, Phlomis, Elymus, and Open communities. This index in the abundance‐weighted case is equivalent to Rao's Q and Hill numbers (Webb et al. 2002). We calculated the standardized effect sizes of abundance-weighted MPD for functional (FSES.mpd) and phylogenetic (PSES.mpd) diversity to produce a phylogenetic and functional index of diversity that is independent of species richness. We used the independent-swap algorithm to draw a null distribution based on 999 randomizations and created standard effect sizes of mpd by comparing the observed community diversity to the null distribution of randomly assembled communities. Positive (SES values > 1.96) and negative (SES values < − 1.96) values of SES.mpd indicate significant functional/phylogenetic over-dispersion and clustering patterns in the community, respectively. We calculated SES.mpd with the “ses.mpd” functions in R package picante (Kembel et al. 2010).
Measures of relative interaction index (RII) in communities considering different nurse types
We calculated the Relative Interaction Index (RII) for species cover to quantify the effect of each nurse species on other species in different combinations of the three treatments (time since the last fire, forms of nurse growth, and their interactions), following Armas et al. (2004):
where XLonicera, XPhlomis, XElymus, and Xopen represent the total plant cover in the four different patches. This index is symmetric about zero (no significant interaction) and varies between 0 and 1 for facilitation and between 0 and − 1 for competition. It was calculated using the summed total plant cover of all species present in communities considering the presence/absence of nurse types.
Measures of species composition and functional groups
We performed a Correspondence Analysis (CA) on Lonicera, Phlomis, Elymus, and Open communities to measure species composition for these plant communities. Correspondence Analysis was performed using the “ca” function in R package ca (Greenacre et al. 2016). Lonicera, Phlomis, Elymus, and open communities were plotted at CA as the distribution of their quadrats (i.e., nurse and open quadrats) with respect to time since the last fire. Then, the amount of variation among the communities on CA axis 1 and CA axis 2 (i.e., percentage of Eigen values for CA axis 1 and CA axis 2) was plotted with respect to times since the last fire.
In addition to the CA ordination of communities in terms of the dispersion of their quadrats, variations in the species composition of communities in CA spaces were determined based on the dispersion of their plant species (Bahalkeh et al. 2021). We then performed a cluster analysis of the functional traits matrix, where the plant species present in the communities were presented in rows and the functional traits in columns to identify groups of species with similar functional identities. Cluster analysis was performed using the “hclust” function based on the Ward.D method in the R package vegan (Oksanen et al. 2016). Before performing the cluster analysis, we measured the functional dissimilarity between plant species using the Gower distance index. Finally, cluster groups were plotted on CA species space to assess the presence of plant species and cluster groups in the communities in relation to time since the last fire.
Statistical analyses
We analyzed variation in RIIs of nurse types and the taxonomic, functional, and phylogenetic diversity relative to the presence/absence of nurse types, different times since the last fire, and their interactions. We analyzed the response of RIIs and biodiversity facets (i.e., taxonomic, functional, and phylogenetic diversity) (using separate analyses for each) relative to the presence/absence of Lonicera nummulariifolia, Phlomis cancellata, and Elymus hispidus in communities by developing linear mixed effect models. The models included the presence/absence of nurse’s growth forms, different times since the last fire and their interactions as fixed effects, and study area as a random factor to account for the lack of independency between communities including different nurse’s growth forms and open communities. The models were fitted using the lmer () function in the R package lmerTEST (Kuznetsova et al. 2017), and their marginal R2 values were obtained using the function “r.squaredGLMM” provided in the package “MuMIn” (Barton 2013). Marginal R2 is the proportion of variance explained solely by our fixed factors (presence of nurse types and times since the last fire). Finally, we calculated p-value for individual treatments (i.e., fixed effects) from F statistics of type III sum of squares with the Satterthwaite approximation to estimate the denominator degrees of freedom using function “summary” provided in the package “lmerTEST” (Kuznetsova et al. 2017).
To examine the relative importance of times since the last fire and the presence of nurse’s growth forms in explaining the variation in each diversity index, we performed variation partitioning based on partial linear regression using the “varpart” function (Oksanen et al. 2016). The total percentage of variation explained was divided into a unique and shared contribution for two sets of predictors: (i) times since last fire (i.e., fire factor), (ii) presence of nurse’s growth forms (i.e. nurse factor), and (iii) their interactions (i.e., shared environment between these factors).
To test whether the presence/absence of nurse’s growth forms, different times since the last fire and their interactions significantly influence species composition, a linear mixed effect model was performed on CA quadrat scores for Lonicera, Phlomis, and Elymus communities (i.e., 50% nurse + 50% open quadrats) and Open communities (i.e., only open quadrats), separately for each CA axis, with presence/absence of nurse’s growth forms, different times since last fire and their interactions treated as fixed effects, and the communities nested within the sites as the random effects. Finally, to assess the effect of these treatments on the cover of plant species in each cluster group, a linear mixed effect model, similar to previous ones, was performed on CA species scores. All linear mixed-effects models were fitted with the function lmer in the R package lmerTest (Kuznetsova et al. 2017), and P-values were calculated from F-statistics of type III sum of squares with the Satterthwaite approximation to estimate the denominator degrees of freedom. Dependent variables were checked for normality and log-transformed before performing linear mixed effect models (Legendre and Legendre 2012).
Results
Effects of nurse’s growth forms on relative interaction index values across times since the last fire
Time scale since the last fire, the presence/absence of nurse’s growth forms and their interactions had highly significant effects on RIIs values for the whole understory community which refers to the presence of all beneficiary species in a given area (Fig. 2). We found negative RII values for the entire understory species in Elymus communities across all times since the last fire, including both short-term and long-term times. However, RII was much more negative in earlier time periods (i.e., 1-year and 4-year sites) compared to long-term times (i.e., 10- and 20-year sites) in the Elymus communities (Fig. 2; Elymus). In contrast, positive RII values were found for understory communities in Phlomis and Lonicera communities across all time scales (Fig. 2; Phlomis and Lonicera). Notably, more positive RII values were obtained for understory communities in Lonicera compared to Phlomis communities across long-term times (10-year and 20-year sites) (Fig. 2).

Mean ± SE (n = 20) of different nurse types (i.e. Elymus, Lonicera and Phlomis) Relative Interaction Index (RII) cover of the whole understory community at sites with different times since last fire (i.e. 1-year, 4-year, 10-year and 20-year sites) and unburned site (control site). Significant treatment effects in two-way linear mixed-effects models on the effects of the time since last fire (TF), presence/absence of nurse type (N) treatments and their interactions (TF x N) are shown in the graphs p < 0.05; ***, p < 0.0001
Effects of nurse’s growth forms on species composition across times since the last fire
All Lonicera communities located in different times since the last fire were scattered on the left-hand side of CA axis 1, and showed significant correlations with CA axis 2 (Fig. 3). In contrast, Elymus and Phlomis communities were scattered across CA axis 1, with more positive CA scores than negative scores (only observed in Elymus and Phlomis across 20-year site). Additionally, they exhibited stronger correlations with CA axis 1 than CA axis 2 (Fig. 3). Overall, plant communities, with respect to the presence of nurse’s growth forms, occupied a wider range of spaces with diverse CA scores depending on times since the last fire. In this regard, communities at longer times (i.e., 10 and 20 years) exhibited larger CA spaces with diverse scores compared to communities at short-term times (i.e., 1 and 4 years) and the control site, whereas all nurse communities (except Elymus communities) were more scattered along CA axis 2 resulting in narrower CA spaces at short-term communities and the control site (Fig. 3). We found significant variation in species composition in relation to presence of nurse plants with different growth forms and time scales since last fire (Fig. 3). In terms of species composition on CA axis 1, the presence of nurse’s growth forms, times since last fire and their interaction significantly explained the observed variation. On the other hand, the time scale and its interaction with nurse plants were the most important factors in explaining variations in species composition on CA axis 2 (Fig. 3; Axis 1 and Axis 2).

CA diagrams of plots (above diagram; nurse and open communities are represented as red circles) and species (below diagram). The communities diagram represents the mean CA scores for the communities ± SE of the three treatment combinations (i.e. time since last fire, nurse types and their interactions). Results of two-way linear mixed-effects models on the effects of the nurse’s growth forms (i.e. nurse types), times since last fire and their interactions are shown below axis 1 and at the left of axis 2: **, p < 0.01. (b) CA1 and CA2 scores are displayed for species cluster groups (A, B, C, D, E)
Five groups were identified in the cluster analysis conducted on the functional trait matrix (see details in Additional file 3: Appendix S3). Group A included only annual herbaceous species with plant height less than 0.50 m (e.g., Callipeltis cucularis, Galium aparine, Asperula glomerata, and Ixilirion tataricum), located on the upper negative side of the CA axis 1, where Elymus and Phlomis communities with long-term times (i.e., 10- and 20-year communities) were scattered on the ordination space (Fig. 3, Additional file 2: Appendix S2). Group B included annual herbaceous and grass species ranging in height from 0.014 to 0.53 m, including Bromus tectorum, Scandix stellata, Filago arvensis, Stipa arabica, Achillea wilhelmsii, Taeniathrum caput-medisae, and Vicia ervilia, most of which were located on the relatively upper left side of CA -axis 1, where mainly Lonicera was scattered at long-times and Elymus and Phlomis at short-term times, as well as the control area in the ordination space (Fig. 3, Additional file 2: Appendix S2). Group C included perennial shrub and cushion species with plant height greater than 0.50 m (e.g., Lonicera nummularifolia, Rosa beggeriana, Acanthophyllum glandulosum, Ziziphora clinopodioides m, and Lactuca orientalis), scattered mainly on upper positive side of CA axis 1, where Lonicera and Phlomis communities occupied the ordination space in long-term times (Fig. 3, Group C). Groups D and E included perennial herbaceous species such as Phlomis cancellata, Eryngium bungei, Hymenocrater calycinus, Noaea mucrunata, and Thalictrum sultanabadense, with perennial herbs greater than 0.5 m tall in group D and those with less than 0.5 m were located in group E (Fig. 3, Additional file 2: Appendix S2). In addition, these plant species were located on the lower positive side of CA -axis 1, where communities were found in short-term periods (i.e., 1, 4-year communities) than in long-term times (Fig. 3). The linear mixed effect models indicated important effects of nurse plants and their interactions with time since the last fire on variation in functional groups on CA axis 1 (Fig. 3). In addition, time scale and its interaction with nurse plants had highly significant effects on CA axis 2 (Fig. 3).
Effects of nurse’s growth forms on biodiversity facets across times since the last fire
Taxonomic indices responded significantly to the presence of nurse plants and time scales since the last fire (Fig. 4; q0 = mR2adj = 0.172, q1 = mR2adj = 0.276). In this context, an increase in taxonomic diversity based on both diversity indices (i.e., q0 and q1) was observed in Lonicera and Phlomis communities at longer times (10 years and 20 years, respectively). In contrast, Elymus communities indicated a decrease in taxonomic diversity in compared to open communities under long-term times since the last fire. The control site illustrated higher taxonomic diversity in open communities (Fig. 4; q0 and q1 panels). Interestingly, nurse plants and their interactions with time scale since the last fire were recognized as the most important predictors of variations in q0 (species richness) and q1 indices between sites (Fig. 4; q0 and q1 panels).

Bar plots showing taxonomic (i.e. species richness (q0) and Shannon’s diversity) (q1)) (above panels), functional (FSES.mpd) and phylogenetic (PSES.mpd) (below panels) diversity at the studied sites with respect to presence of nurse plants with grass (Elymus hispidens), herb (Phlomis cancellata) and shrub (Lonicera nummulariifolia) growth forms and different times since last fire. Variation in taxonomic, functional and phylogenetic indices across times since last fire and presence of nurse types were measured via linear mixed effect models. The marginal adjusted R2 shows the variation explained by the fixed factors in our statistical models (mR2adj. are shown in each case). Significant treatment effects shown from two-way linear mixed-effects models on the effects of the time since last fire (TF), presence/absence of nurse type (N) treatments and their interactions (TF x N) are shown in the graphs. *, p < 0.05; **, p < 0.01; ***, p < 0.0001
Significant and consistent effects of nurse’s growth forms and their interaction were observed for functional and phylogenetic diversity. In addition, time since the last fire indicated the significant effects on these biodiversity indices especially on variations of phylogenetic diversity (Fig. 4; FSES.mpd = mR2adj = 0.433; PSES.mpd = mR2adj = 0.389; please see mR2adj values and significance levels for the treatments on biodiversity indices in Fig. 4). In this regard, higher functional and phylogenetic diversity (i.e., functional and phylogenetic overdispersion) were observed in Lonicera communities in all times since last fire except in the control site (Fig. 4; Lonicera). In contrast, Phlomis and Elymus communities indicated a significant decrease in functional and phylogenetic diversity (i.e., functional/phylogenetic clustering) under all times especially in short-term times (i.e., 1 and 4-year sites) for Phlomis communities (Fig. 3; Phlomis) and under long-term times for Elymus communities (Fig. 3; Elymus and Control). However, stronger and consistent decreases in functional and phylogenetic diversity were observed for Elymus communities than Phlomis communities, especially under long times (Fig. 4).
Relative importance of fire and nurse’s growth forms on biodiversity facets
The amount of variance explained for all diversity facets increased when considering nurse plants (orange fraction in Fig. 5). This was particularly true for all biodiversity indices, especially for functional and phylogenetic diversity (please see orange fraction (nurse factor) in Fig. 5). In addition, time since last fire was a particularly strong predictor of variation in all biodiversity facets important for phylogenetic diversity (Fig. 5; yellow fraction (Fire factor)). Interaction between the presence of nurse plants and time since the last fire (see shared area between orange and yellow fractions) indicated contributions to explain variations in biodiversity components (Fig. 5; see shared area between orange and yellow fractions for all biodiversity facets). Although such contributions were not the strongest for biodiversity components, this suggests that the effects of nurse’s growth forms on biodiversity facets are relatively sensitive to time since the last fire.

The relative contribution of nurse types (NURSE in orange) and times since last fire (FIRE in yellow) to taxonomic (q0 and q1 indices), functional (FSES.mpd) and phylogenetic (PSES.mpd) diversity. Values represent the adjusted R2‐values
Discussion
The influence of nurse plants on biodiversity and species composition, particularly in severe ecosystems such as drylands and alpine and mountainous semi-arid ecosystems, has received some attention in the last two decades (Bashirzadeh et al. 2022a; Rafiee et al. 2022; Pashirzad et al. 2019a; 2019b; Butterfield and Briggs 2011; Valiente-Banuet and Verdu 2007; Cavieres et al. 2005). However, we still do not have enough information regarding nurse plant effects on post-fire recovery of vegetation in relation to important factors such as growth form, time since fire, and at an applicable scale for burned ecosystems and plant successions (He et al. 2019; Keeley et al. 2012). Here, we analyzed the effects of select dominant perennial plants representing grass, herb and shrub growth forms on species composition and taxonomic, functional and phylogenetic diversity across different time-scales in a semi-arid shrublands of the Fereizi region located in Northeast of Iran.
Our results show the specific effects of perennial plants on the natural recovery of vegetation depending on their growth forms and time intervals. For example, Lonicera nummulariifolia shrubs significantly enhanced species composition and all components of biodiversity especially under long times since the last fire, whereas Elymus hispidus grasses showed a negative effect on all biodiversity components through recruiting the only grasses and herbaceous plants. Phlomis communities indicated an increase in taxonomic diversity, while they had a negative effect on functional and phylogenetic diversity through including herbaceous and shrub species. Interestingly, even though the specific effects of nurse’s growth forms on biodiversity components depending on their morphological characteristics, all the effects of nurse’s growth forms peaked under long times since the last fire (i.e., 10- and 20-year sites). In addition, fire solely and in the form of interaction with nurse types was recognized as one of the important factors in explaining the variation of species composition and biodiversity facets in our study area. Based on these results, these factors have served as strong explanations for the major importance of nurse plants with different growth forms as drivers of vegetation restoration in burned ecosystems (Paniw et al. 2017; He et al. 2019). In addition, our results strongly indicate that fire, as an agent of disturbance and significant ecological and evolutionary force, should have an influence on biodiversity across various spatiotemporal scales through its interactions with different nurse plants (e.g., Schnitzler et al. 2011; Cramer and Verboom 2017; Pausas and Lamort 2018).
Nurse plants influence species composition and biodiversity components depending on their growth forms in burned communities
The effects of different nurse plants on species composition and some specific biodiversity components were associated with growth form, suggesting that the morphological features of nurse plants, as well as some processes controlling beneficiary relatedness beneath canopies of nurse plants, likely have impacts (Gomez-Aparicio 2009). This finding suggests the presence of different ecological conditions within these communities, which can be attributed to the varying effects of nurse plants in creating favorable microclimates based on their specific characteristics (He et al. 2012). There are hypotheses suggesting facilitative impacts of nurse shrubs driven by their specific morphological features, such as a low root-to-shoot ratio (i.e., a less competitive strategy) (Piston et al. 2016; Gomez-Aparicio 2009), to support a high abundance of beneficiaries with different functional identities. Following this hypothesis, our results indicated an increase in taxonomic diversity consistent with the presence of post-fire recruiting plant species, which are usually short-lived herbs, grasses, and subshrubs that were mostly observed in Lonicera communities. Such observations could introduce drought-resilient shrubs as ecosystem engineers, promoting biodiversity by facilitating the survival and growth of other long-lived shrubs, herbs, and grasses (reviewed in Gómez-Aparicio et al. 2004; Calvo et al. 1999). On the other hand, these positive effects of shrubs on the recruitment of other plant species can provide new insights into developing plant restoration practices (Gómez-Aparicio et al. 2004; Castro et al. 2006; Siles et al. 2008; Padilla and Pugnaire 2006) by understanding the mechanisms promoted by nurse shrubs in the recovery of plant biodiversity. In this context, we found that nurse shrubs allow for a specific trade-off; competition among closely related taxa and facilitation among distantly related taxa (Soliveres et al. 2012). The significant increase in functional and phylogenetic diversity among beneficiary species in communities including Lonicera nummulariifolia shrubs represents the presence of plant species with different functional identities (i.e., those with complete differentiation in realized niches) (Bannister et al. 2019; Pashirzad et al. 2018; Bahalkeh et al. 2017).
In contrast, the nurse grass (Elymus hispidus) and herb (Phlomis cancellata) strongly influenced species composition, leading to a decrease in all biodiversity components, especially over long periods of time. Additionally, the nurse herb had a less extensive impact on functional and phylogenetic diversity, particularly at short-term sites. These results highlight the differences in mechanisms promoted by nurse grass compared to nurse herb, depending on their morphological characteristics. Grasses generally have a larger negative effect than herbs, which supports previous studies showing grasses to be better competitors with higher establishment and growth rates than herbs (Goldberg et al. 1999; Pywell et al. 2003). Grasses have fibrous roots and large root-to-shoot ratios, which allow them to compete efficiently for numerous resources. Due to this strong competitive ability, nurse grasses tend to restrict the presence of beneficiary species, ultimately leading to the dominance of stress-tolerant species. These stress-tolerant species are capable of tolerating the dual environmental filtering imposed by nurse grasses, such as annual and perennial forbs and grasses (Harrington and Johns 1990; Puerta-Piñero et al. 2007). These specific mechanisms alter the overall species composition of plant communities by introducing plant species that would not otherwise be present (Butterfield et al. 2013; He et al. 2012; Soliveres et al. 2015). Thus, our findings have revealed innovative features of nurse grass in the recovery of vegetation. These benefits of using grasses as nurse plants have not been commonly identified in restoration practices, except in wetlands (Egerova et al. 2003; McKee et al. 2007) and semi-arid steppes (Maestre et al. 2001).
Nurse plants influence species composition and biodiversity facets depending on time since last fire
We found that nurse species with different growth forms had a significant impact on species composition and biodiversity, especially at intermediate and long-time intervals since the last fire (i.e., at 10 and 20-year sites). These results provide strong empirical support for the intermediate disturbance hypothesis (IDH) (Huston 2014), which suggests the positive effects of nurse plants on biodiversity, resulting in an increase in biodiversity, consistent with an increase in time since the last fire. Furthermore, the positive effects of nurse plants on biodiversity reach their peak under moderate levels (Dee and Menges 2014; Beckage and Stout 2000). It is worth noting that nurse plants may only effectively restore vegetation in burned ecosystems under moderate and long-time intervals since the last fire (Tepley et al. 2017). Therefore, the occurrence of fire and its impacts on vegetation play a crucial role in determining the post-fire recovery of vegetation by influencing the mechanisms promoted by nurse plants (Castro et al. 2006; Gomez-Aparicio 2009; 2004). In this regard, our results demonstrate the positive effects of fire on increasing the competitive and facilitative abilities of nurse grasses and shrubs, respectively. This finding contradicts previous research, which mostly argues that fire decreases competition (Bahalkeh et al. 2021). Therefore, we conclude that nurse types and time since the last fire are important drivers of the variance in species composition and biodiversity facets.
Furthermore, it was expected that fire would explain a larger amount of variance in phylogenetic diversity compared to other biodiversity facets, despite the strong influence of nurse plants on phylogenetic diversity (e.g., He et al. 2019; Paniw et al. 2017; Khapugin et al. 2016). Previous studies have highlighted the presence of strong phylogenetic conservatism among plant species in post-fire recovery, indicating that these lineages have evolved in response to fire disturbances (He et al. 2019). Therefore, fire acts as a significant driver of diversification by promoting mutation, selection, divergence, and speciation (Stevens-Rumann et al. 2018). This leaves a strong fingerprint on current phylogenetic diversity and may explain the stronger effect of fire on this biodiversity component compared to other components (see variance partitioning analyses in Fig. 3; PSES.mpd). Consequently, analyzing biodiversity responses to nurse plants collectively and considering the time since the last fire offers strong potential for linking fire to the evolutionary history of post-fire vegetation and the mechanisms of nurse plants (Keeley et al. 2012; Verdu and Pausas 2007).
Management implications
The findings of this research study, which primarily focus on the significance of nurse plant growth forms in the natural recovery of vegetation, are consistent with the recent resurgence of facilitation as the dominant interaction in intermediate disturbance levels (Callaway 2007; Brooker et al. 2008). These findings could provide strong support for the use of plant-based techniques in the restoration practices of burned ecosystems. Implementing such procedures would be considered cost-effective and environmentally friendly, aligning with recent conceptual models that hypothesize plant facilitation as an exploitable interaction in active restoration (Navarro-Cano et al. 2018; Cuevas et al. 2013; Ren et al. 2008). In this regard, nurse plants were identified as a key factor in managing the conservation of vegetation and preventing the loss of plant biodiversity, particularly in the short time since the last fire. Therefore, we recommend the planting of seedlings or seeds of dominant perennial plants such as shrubs and herbaceous species as an effective measure for restoring burned ecosystems. This recommendation is based on our comprehensive evaluation of the effects of nurse plants on the natural recovery of vegetation, taking into account the different growth forms of nurse species, the appropriate scale of vegetation (i.e., community scale, where the most influential theories were developed), and the various time scales since the last fire.
Conclusion
In this study, we observed that all growth forms of nurse plants influenced species composition and various biodiversity components, regardless of the time since the last fire. Lonicera communities exhibited enhanced biodiversity facets alongside a significant change in species composition. Conversely, we found a decrease in taxonomic, functional, and phylogenetic diversity in Elymus communities. Additionally, we found that nurse herb species had a positive effect on species composition and taxonomic diversity in Phlomis communities, while it had a negative effect on functional and phylogenetic diversity. Interestingly, these specific effects of nurse types were predominantly observed in intermediate and long time since the last fire sites (i.e., 10 and 20 years). Based on these findings, we conclude that nurse plants have stronger and more consistent effects on species composition and overall biodiversity than the time since the last fire. However, fire made a significant contribution to phylogenetic diversity. These findings demonstrate the substantial effects of fire on the evolutionary history of vegetation during post-fire recovery in burned ecosystems. Therefore, this study suggests that nurse plants can restore species composition and plant biodiversity, particularly under long times since the last fire, aligning with the intermediate disturbance hypothesis. Consequently, nurse plants can be considered a practical factor for developing active restoration practices in burned ecosystems.
Availability of data and materials
Additional supporting information may be found in the online version of the article at the publisher’s website.
References
Abedi, M., R. Omidipour, S.V. Hosseini, K. Bahalkeh, and N. Gross. 2022. Fire disturbance effects on plant taxonomic and functional β-diversity mediated by topographic exposure. Ecology and Evolution 12: e8552.
Alam, M.A., S.V. Wyse, H.L. Buckley, G.L. Perry, J.J. Sullivan, N.W. Mason, R. Buxton, S.J. Richardson, and T.J. Curran. 2020. Shoot flammability is decoupled from leaf flammability, but controlled by leaf functional traits. Journal of Ecology 108 (2): 641–653. https://doi.org/10.1111/1365-2745.13289.
Anacker, B., N. Rajakaruna, D. Ackerly, S. Harrison, J. Keeley, and M. Vasey. 2011. Ecological strategies in California chaparral: Interacting effects of soils, climate, and fire on specific leaf area. Plant Ecology and Diversity 4 (2–3): 179–188. https://doi.org/10.1080/17550874.2011.633573.
Armas, C., R. Ordiales, and F.I. Pugnaire. 2004. Measuring plant interactions: a new comparative index. Ecology 85: 2682–2686. https://doi.org/10.1890/03-0650.
Atashgahi, Z., H. Ejtehadi, M. Mesdaghi, and F. Ghasemzadeh. 2018. Plant diversity of the Heydari Wildlife Refuge in northeastern Iran, with a checklist of vascular plants. Phytotaxa 340: 101–127.
Badano, E.I., and L.A. Cavieres. 2006. Impacts of ecosystem engineers on community attributes: Effects of cushion plants at different elevations of the Chilean Andes. Diversity and Distributions 12: 388–396.
Bahalkeh, K., M. Abedi, and G. Dianati Tilaki. 2017. Microclimate changes of cushion species Onobrychis cornuta affected by fire in Golestan National Park Grasslands. Iranian Journal of Ecohydrology 3: 623–630.
Bahalkeh, K., M. Abedi, D. Tilaki, and R. Michalet. 2021. Fire slightly decreases on the short-term the competitive effects of a thorny cushion shrub in a semi-arid mountain steppe. Applied Vegetation Science 24: e12575. https://doi.org/10.1111/avsc.12575.
Bannister, J.A., N. Travieso, M. Acevedo, et al. 2019. Shrub influences on seedling performance when restoring the slow-growing conifer Pilgerodendron uviferum in southern bog forests. Restoration Ecology. https://doi.org/10.1111/rec.13090.
Barton, K. 2013. Package ‘MuMIn’. Model Selection and Model Averaging Based on Information Criteria. R package version 1.15.11. http://CRAN.R-project.org/package=MuMIn. Accessed 11 Jan 2023.
Barzegaran, F., R. Erfanzadeh, S. Saber Amoli. 2022. The effect of woody plants on the understory soil parameters is different between grazed and ungrazed areas. Community Ecology 23: 187–196. https://doi.org/10.1007/s42974-022-00090-x.
Bashirzadeh, M., S. Soliveres, M. Farzam, and H. Ejtehadi. 2022a. Plant–plant interactions determine taxonomic, functional and phylogenetic diversity in severe ecosystems. Global Ecology and Biogeography 31 (4): 649–662.
Bashirzadeh, M., M. Abedi, and M. Farzam. 2022. Plant–plant interactions determine natural restoration of plant biodiversity over time, in a degraded mined land. Ecology and Evolution 12: e8878. https://doi.org/10.1002/ece3.8878.
Bashirzadeh, M., M. Abedi, R.P. Shefferson, and M. Farzam. 2023. Post-Fire Recovery of Plant Biodiversity Changes Depending on Time Intervals since Last Fire in Semiarid Shrublands. Fire 6 (3): 103. https://doi.org/10.3390/fire6030103.
Beckage, B., and I.J. Stout. 2000. The effects of repeated burning on species richness in Florida sandhills: A test of the intermediate disturbance hypothesis. Journal of Vegetation Science 11: 113–122.
Bond, W.J., F.I. Woodward, and G.F. Midgley. 2005. The global distribution of ecosystems in a world without fire. New Phytologist 165: 525–538.
Bowd, E.J., D.B. Lindenmayer, S.C. Banks, and D.P. Blair. 2018. Logging and fire regimes alter plant communities. Ecological Applications: A Publication of the Ecological Society of America 28: 826–841. https://doi.org/10.1002/eap.1693.
Bowman, D.M.J.S., G.L. Perry, S.I. Higgins, C.N. Johnson, S.D. Fuhlendorf, and B.P. Murphy. 2016. Pyrodiversity is the coupling of biodiversity and fire regimes in food webs. Philosophical Transactions of the Royal Society B 371: 20150169.
Brooker, R.W., F.T. Maestre, R.M. Callaway, C.L. Lortie, L.A. Cavieres, G. Kunstler, et al. 2008. Facilitation in plant communities: The past, the present, and the future. Journal of Ecology 96: 18–34.
Butterfield, B.J., and J.M. Briggs. 2011. Regeneration niche differentiates functional strategies of desert woody plant species. Oecologia 165 (2): 477–487. https://doi.org/10.1007/s00442-010-1741-y.
Butterfield, B, J., L.A. Cavieres, R.M. Callaway, B.J. Cook, Z. Kikvidze, C.J. Lortie, R. Michalet,… R.W. Brooker. 2013. Alpine cushion plants inhibit the loss of phylogenetic diversity in severe environments. Ecology Letters 16 478-486.
Bytebier, B., A. Antonelli, D.U. Bellstedt, and H.P. Linder. 2011. Estimating the age of fire in the Cape flora of South Africa from an orchid phylogeny. Proceedings of the Royal Society B 278: 188–195.
Cadotte, M.W., and T.J. Davies. 2016. Phylogenies in ecology: A guide to concepts and methods. Princeton, NJ: Princeton University Press. https://doi.org/10.1515/9781400881192.
Calitz, W., A.J. Potts, and R.M. Cowling. 2015. Investigating species level flammability across five biomes in the Eastern Cape, South Africa. South African Journal of Botany 101: 32–39.
Callaway, R.M. 2007. Positive interactions and interdependence in plant communities. Springer.
Calvo, A.M., L.L. Hinze, H.W. Gardner, and N.P. Keller. 1999. Sporogenic effect of polyunsaturated fatty acids on development of Aspergillus spp. Applied and Environment Microbiology 65: 3668–3673.
Camac, J., R.J. Williams, C.H. Wahren, W.K. Morris, and J.W. Morgan. 2013. Post-fire regeneration in alpine heathland: Does fire severity matter? Austral Ecology 38 (2): 199–207.
Castro, J., R. Zamora, and J.H. Hódar. 2006. Restoring Quercus pyrenaica forests using pioneer shrubs as nurse plants. Applied Vegetation Science 9: 137–142.
Cavieres, L.A., E.I. Badano, A. Sierra-Almeria, S. Gomez-Gonzalez, and M.A. Molina-Montenegro. 2005. Positive interactions between alpine plant species and the nurse cushion plant Laretia acaulis do not increase with elevation in the Andes of central Chile. New Phytologist 196 (1): 59–69.
Cavieres, L.A., R.W. Brooker, B.J. Butterfield, B.J. Cook, Z. Kikvidze, C.J. Lortie, R. Michalet, F.L. Pugnaire, C. Schöb, S. Xiao, and F. Anthelme. 2013. Facilitative plant interactions and climate simultaneously drive alpine plant diversity. Ecology Letters 17 (2): 193–202.
Cayuela, L., I. Granzow-de la Cerda, F.S. Albuquerque, and D.J. Golicher. 2012. taxonstand: An r package for species names standardization in vegetation databases. Methods in Ecology and Evolution 3 (6): 1078–1083.
Chao, A., and L. Jost. 2012. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 93 (12): 2533–2547. https://doi.org/10.1890/11-1952.1.
Chao, A., N.J. Gotelli, T.C. Hsieh, E.L. Sander, K.H. Ma, R.K. Colwell, and A.M. Ellison. 2014. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecological Monographs 84: 45–67. https://doi.org/10.1890/13-0133.1.
Cramer, M.D., and G.A. Verboom. 2017. Measures of biologically relevant environmental heterogeneity improve prediction of regional plant species richness. Journal of Biogeography 44: 579–591.
Cruz, O., S.F. Riveiro, M. Casal, and O. Reyes. 2022. Effect of fire factors (smoke, ash, charcoal and heat) on seeds of plant species. MethodsX 9: 101679.
Cuevas, J.G., S.I. Silva, P. León-Lobos, and R. Ginocchion. 2013. Nurse effect and herbivory exclusion facilitate plant colonization in abandoned mine tailings storage facilities in north-central Chile. Revista Chilena De Historia Natural 86: 63–74. https://doi.org/10.4067/S0716-078X2013000100006.
Dantas, V.L., M. Batalha, and J.G. Pausas. 2013. Fire drives functional threshold on the savanna–forest transition. Ecology 94 (11): 2454–2463.
Davies, K.W., T.J. Svejcar, and J.D. Bates. 2009. Interaction of historical and nonhistorical disturbances maintains native plant communities. Ecological Applications 19: 1536–1545. https://doi.org/10.1890/09-0111.1.
Dee, J.R., and E.S. Menges. 2014. Gap ecology in the Florida scrubby flatwoods: Effects of timesince-fire, gap area, gap aggregation and microhabitat on gap species diversity. Journal of Vegetation Science 25: 1235–1246.
Egerova, J., C.E. Proffitt, and S.E. Travis. 2003. Facilitation of survival and growth of Baccharis halimifolia L. by Spartina alterniflora Loisel. in a created Louisiana salt marsh. Wetlands 23: 250–256.
Enright, N.J., E. Mosner, B.P. Miller, N. Johnson, and B.B. Lamont. 2007. Soil versus canopy seed storage and plant species coexistence in species-rich shrublands of southwestern Australia. Ecology 88: 2292–2304.
Erfanian, M.B., F. Memariani, Z. Atashgahi, M. Mesdaghi, M. Saeedi, M. Darrudi, M. Hamedian, S. Hosseini, and H. Ejtehadi. 2021. Unpalatable plants induce a species-specifc associational efect on neighboring communities. Scientific Reports 11: 14380.
Erfanzadeh, R., R. Shahbazian, and H. Zali. 2014. Role of plant patches in preserving flora from the soil seed bank in an overgrazed high-mountain habitat in northern Iran. Journal of Agricultural Sciences and Technology 16: 229–238.
Erfanzadeh, R., F. Barzegaran, S. Saber Amoli, and J. Petillon. 2022. The effect of shrub community on understory soil seed bank with and without livestock grazing. Community Ecology 23 (4): 1–11.
Gastauer, M., J.R. Silva, C.F. Caldeira Junior, S.J. Ramos, P.W.M. Souza Filho, A.E. Furtini Neto, and J.O. Siqueira. 2018. Mine land rehabilitation: Modern ecological approaches for more sustainable mining. Journal of Cleaner Production 172: 1409–1422. https://doi.org/10.1016/j.jclepro.2017.10.223.
Ghasempour, M., R. Erfanzadeh, and P. Torok. 2022. Fire effects on soil seed banks under different woody plant species in Mazandaran province. Iran. Ecological Engineering 183: 106762.
Goldberg, D.E., T. Rajaniemi, J. Gurevitch, and A. Stewart-Oaten. 1999. Empirical approaches to quantifying interaction intensity: Competition and facilitation along productivity gradients. Ecology 80: 1118–1131.
Gómez-Aparicio, L., R. Zamora, J.M. Gómez, J.A. Hódar, J. Castro, and E. Baraza. 2004. Applying plant facilitation to forest restoration: A meta-analysis of the use of shrubs as nurse plants. Ecological Applications 14: 1128–1138.
Gomez-Aparicio, L. 2009. The role of plant interactions in the restoration of degraded ecosystems: A meta-analysis across life-forms and ecosystems. Journal of Ecology 97: 1202–1214. https://doi.org/10.1111/j.1365-2745.2009.01573.
Gomez-Gonz ´ alez ´, S, C. Torres-Díaz, C. Bustos-Schindler, and E. Gianoli. 2011. Anthropogenic fire drives the evolution of seed traits. Proceedings of the National Academy of Sciences of the United States of America 108: 18743–18747.
Gosper, C.R., S.M. Prober, and C.J. Yates. 2013. Multi-century changes in vegetation structure and fuel availability in fire-sensitive eucalypt woodlands. Forest Ecology and Management 310: 102–109.
Greenacre, M., O. Nenadic, M. Friendly. 2016. Package ‘ca’. Simple, Multiple and joint correspondence analysis. Version 0.64. Available at: http://cran.r-project.org/web/packages/ca/ca.pdf. Accessed 11 Jan 2023.
Hanes, T.L. 1971. Succession after fire in the chaparral of southern California. Ecological Monographs 41: 27–52.
Harrington, G.N., and G.G. Johns. 1990. Herbaceous biomass in a eucalyptus-savanna woodland after removing trees and/or shrubs. Journal of Applied Ecology 27: 775–787.
He, T., J.G. Pausas, C.M. Belcher, D.W. Schwilk, and B.B. Lamont. 2012. Fire-adapted traits of Pinus arose in the fiery cretaceous. New Phytologist 194: 751–759.
He, T., B.B. Lamont, and J.G. Pausas. 2019. Fire as a key driver of Earth’s biodiversity. Biological Review 94: 1983–2010.
Herath, D.N., B.B. Lamont, N.J. Enright, and B.P. Miller. 2009. Impact of fire on plant-species persistence in post-mine restored and natural shrubland communities in southwestern Australia. Biological Conservation 142: 2175–2180.
Hernandez-Serrano´, A., M, M. Verdu’, S.C. Gonzalez-Martínez’, and J.G. Pausas’s. 2013. Fire structures pine serotiny at different scales. American Journal of Botany 100: 2349–2356.
Heydari, M., R. Omidipour, M. Abedi, and C. Baskin. 2017. Effects of fire disturbance on alpha and beta diversity and on beta diversity components of soil seed banks and aboveground vegetation. Plant Ecology and Evolution 150: 247–256. https://doi.org/10.5091/plecevo.2017.1344.
Hijmans, R.J., S.E. Cameron, J.L. Parra, P.G. Jones, and A. Jarvis. 2005. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25 (15): 1965–1978. https://doi.org/10.1002/joc.1276.
Hipp, A.L., D.J. Larkin, R.S. Barak, M.L. Bowles, and M.W. Cadotte. 2015. Phylogeny in the service of ecological restoration. American Journal of Botany 102: 647e648. https://doi.org/10.3732/ajb.1500119.
Hurteau, M.D., J.B. Bradford, P.Z. Fulé, A.H. Taylor, and K.L. Martin. 2014. Climate change, fire management, and ecological services in the southwestern US. Forest Ecology and Management 327: 280–289. https://doi.org/10.1016/j.foreco.2013.08.007.
Huston, M.A. 2014. Disturbance, productivity, and species diversity: Empiricism vs. logic in ecological theory. Ecology 95: 2382–2396.
Jankju, M. 2013. Role of nurse shrubs in restoration of an arid rangeland: Effects of microclimate on grass Establishment. Journal of Arid Environment 89: 103–109. https://doi.org/10.1016/j.jaridenv.2012.09.008.
Jankju, M., and H. Ejtehadi. 2016. Effects of drought and slope aspect on canopy facilitation in a mountainous rangeland. Journal of Plant Ecology 10 (4): 626–633.
Jankju, M., A. Delavari, and A. Ganjali. 2008. Interseeding Bromus kopetdaghensis, in shrublands. Iranian Journal of Rangland Science 2: 314–328.
Jin, Y., and H. Qian. 2019. V.PhyloMaker: An R package that can generate very large phylogenies for vascular plants. Ecography 42: 1353–1359.
Kattge, J., G. Bönisch, S. Díaz, S. Lavorel, I.C. Prentice, P. Leadley, ... M. Cuntz. 2020. TRY plant trait database–enhanced coverage and open access. Global Change Biology, 26(1), 119–188.
Keeley, J.E., and C.J. Fotheringham. 2003. Species–area relationships in mediterranean-climate plant communities. Journal of Biogeography 30: 1629–1657.
Keeley, J.E., W.J. Bond, R.A. Bradstock, et al. 2012. Fire in Mediterranean ecosystems: Ecology, evolution and management. Cambridge: Cambridge University Press Cambridge.
Keeley, J.E., J.G. Pausas, P.W. Rundel, W.J. Bond, and R.A. Bradstock. 2016. Fire as an evolutionary pressure shaping plant traits. Trends in Plant Science 16: 406–411.
Kembel, S.W., P.D. Cowan, M.R. Helmus, W.K. Cornwell, H. Morlon, D.D. Ackerly, S.P. Blomberg, and C.O. Webb. 2010. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 26 (11): 1463–1464. https://doi.org/10.1093/bioinformatics/btq166.
Khapugin, A.A., E.V. Vargot, and G.G. Chugunov. 2016. Vegetation recovery in fire-damaged forests: A case study at the southern boundary of the taiga zone. Forestry Studies 64: 39–50. https://doi.org/10.1515/fsmu-2016-0003.
Kleyer, M., Bekker, R. M., Knevel, I. C., Bakker, J. P., … & Peco, B. 2008. The LEDA Traitbase: a database of life‐history traits of the Northwest European flora. Journal of Ecology, 96 (6), 1266–1274.
Kuznetsova, A., L. Hartmann, and A. Burkert. 2017. The Astrophysical Journal 836: 190.
Lamanna, C., B. Blonder, C. Violle, N.J.B. Kraft, B. Sandel, I. Imova, J.C. Donoghue, J. C. Svenning, B. J. McGill, B. Boyle, V. Buzzard, S. Dolins, P.M. Jorgensen, A. Marcuse-Kubitza, N. Morueta-Holme, R.K. Peet, W.H. Piel, J. Regetz, M. Schildhauer , …B.J. Enquist. 2014. Functional trait space and the latitudinal diversity gradient. Proceedings of the National Academy of Sciences of the United States of America 111(38): 13745–13750. https://doi.org/10.1073/pnas.1317722111.
Lavorel, S., S. Díaz, J.H.C. Cornelissen, E. Garnier, S.P. Harrison, S. McIntyre, J.G. Pausas, N. Pérez-Harguindeguy, C. Roumet, and C. Urcelay. 2007. Plant functional types: Are we getting any closer to the Holy Grail? In Terrestrial ecosystems in a changing world, ed. J. Canadell, L.F. Pitelka, and D. Pataki, 149–164. Springer.
Le Bagousse-Pinguet, Y., S. Soliveres, N. Gross, R. Torices, M. Berdugo, and F.T. Maestre. 2019. Phylogenetic, functional, and taxonomic richness have both positive and negative effects on ecosystem multifunctionality. Proceedings of the National Academy of Sciences 116 (17): 8419–8424.
Legendre, P., and L. Legendre. 2012. Numerical Ecology, 3rd ed. Amesterdam, NL: Elsevier.
Lienin, P., and M. Kleyer. 2012. Plant trait responses to the environment and effects on ecosystem properties. Basic and Applied Ecology 13 (4): 301–311. https://doi.org/10.1016/j.baae.2012.05.002.
Madrigal-Gonzalez, J., C. Cano-Barbacil, J. Kigel, P. Ferrandis, and A.L. Luzuriaga. 2020. Nurse plants promote taxonomic and functional diversity in an arid Mediterranean annual plant community. JournAl of Vegetation Science 31 (4): 658–666.
Maestre, F.T., S. Bautista, J. Cortina, J. Bellot, and J. 2001. Potential of using facilitation by grasses to establish shrubs on a semiarid degraded steppe. Ecological Applications 11: 1641–1655.
Maitner, B. S., B. Boyle, N. Casler, R. Condit,….B.J. Enquist. 2018. The bien r package: A tool to access the Botanical Information and Ecology Network (BIEN) database. Methods in Ecology and Evolution 9 (2): 373-379.
McKee, K.L., J.E. Rooth, and I.C. Feller. 2007. Mangrove recruitment after forest disturbance is facilitated by herbaceous species in the caribbean. Ecological Applications 17: 1678–1693.
McLauchlan, K. K., P.E. Higuera, J. Miesel, B.M. Rogers, J. Schweitzer, J. K. Shuman, A.J. Tepley, J. M. Varner, T.T. Veblen, S.A. Adalsteinsson, J.K. Balch, P. Baker, E. Batllori, E. Bigio, P. Brando, M. Cattau, M., M.L. Chipman, J. Coen, R. Crandall, … A.C. Watts. 2020. Fire as a fundamental ecological process: Research advances and frontiers. Journal of Ecology 108(5): 2047–2069. https://doi.org/10.1111/1365-2745.13403.
Memariani, F., M.R. Joharchi, H. Ejtehadi, and K.H. Emadzadeh. 2009. A cintribution of the flora and vegetation of Binalood mountain range. NE Iran: Floristic and Chrological Studies in Fereizi, Ferdowsi University International Journal of Biological Science 1: 1–17.
Memariani, F., V. Zarrinpour, H. Akhani. 2016. A review of plant diversity, vegetation and phytogeography of the Khorassan-Kopet Dagh floristic province in the Irano-Turanian region (northeastern Iran-southern Turkmenistan), Phytotaxa (1): 249, PP: 8–30.
Meng, X., Z. Huang, J. Di, D. Mu, Y. Wang, X. Zhao, H. Zhao, W. Zhu, X. Li, L. Kong, and L. Xing. 2015. Expression of Human epidermal growth factor receptor-2 in resected rectal cancer. Medicine 94 (47): 1–8. https://doi.org/10.1097/MD.0000000000002106.
Mucina, L., J.L. Tsakalos, and P.D. Macintyre. 2019. Ecology, biodiversity and mining: science and solving the challenges. In Mine Closure 2019: Proceedings of the 13th International Conference on Mine Closure, ed. A.B. Fourie and M. Tibbett, 19–34 Australian Centre for Geomechanics.
Navarro-Cano, J.A., M. Goberna, and M. Verdú. 2018. Data from: Trait based selection of nurse plants to restore ecosystem functions in mine tailings. Journal of Applied Ecology 55: 1195–1206.
Niknam, P., R. Erfanzadeh, H. Ghelichnia, and A. Cerda. 2018. Spatial Variation of Soil Seed Bank under Cushion Plants in a Subalpine Degraded Grassland. Land Degradation and Development 29: 4–14.
Oksanen, J., F. G. Blanchet, M. Friendly, R. Kindt, P. Legendre, D. McGlinn, P.R. Minchin, R. B. O’Hara, G. L. Simpson, P. Solymos, M.H.H. Stevens, E. Szoecs, H. Wagner. 2016. vegan: Community Ecology Package. R package version 2.4–1. https://CRAN.R-project.org/package=vegan. Accessed 15 Jan 2023.
Omidipour, R., P. Tahmasebi, M.F. Faizabadi, M. Faramarzi, and A. Ebrahimi. 2021. Does β diversity predict ecosystem productivity better than species diversity? Ecological Indicators 122: 107212. https://doi.org/10.1016/j.ecolind.2020.107212.
Padilla, F.M., and F.I. Pugnaire. 2006. The role of nurse plants in the restoration of degraded environments. Frontiers in Ecology and Environment 4 (4): 196–202.
Paniw, M., R. Salguero-Gómez, and F. Ojeda. 2017. Transient facilitation of resprouting shrubs in fire-prone habitats. Journal of Plant Ecology 11 (3): 475–483.
Pashirzad, M., H. Ejtehadi, J. Vaezi, and R.P. Sheferson. 2018. Spatial scale dependent phylogenetic signal in species distributions along geo graphic and elevation gradients in a mountainous rangeland. Ecology and Evolution 00: 1–11. https://doi.org/10.1002/ece3.4293.
Pashirzad, M., H. Ejtehadi, J. Vaezi, and R.P. Sheferson. 2019a. Plant–plant interactions influence phylogenetic diversity at multiple spatial scales in a semi-arid mountain rangeland. Oecologia 189: 745–755.
Pashirzad, M., H. Ejtehadi, J. Vaezi, and R.P. Sheferson. 2019b. Multiple processes at different spatial scales determine beta diversity patterns in a mountainous semi-arid rangeland of Khorassan-Kopet Dagh floristic province, NE Iran. Plant Ecology 220: 829–844.
Pausas, J.G., and B.B. Lamort. 2018. Ecology and biogeography in 3D: The case of the Australian Proteaceae. Journal of Biogeography 45: 1469–1477.
Pausas, J.G., and E. Ribeiro. 2013. The global fire-productivity relationship. Global Ecology and Biogeography 22: 728–736.
Pausas, J.G., N. Ouadah, A. Ferran, T. Gimeno, and R. Vallejo. 2003. Fire severity and seedling establishment in Pinus halepensis woodlands, eastern Iberian Peninsula. Plant Ecology 169: 205–213.
Piston, N., C. Schob, C. Armas, and I. Prieto. 2016. Contribution of co-occurring shrub species to community richness and phylogenetic diversity along an environmental gradient. Perspective in Plant Ecology, Evolution and Systematics 19: 30–39.
Puerta-Piñero, C., J.M. Gómez, and F. Valladares. 2007. Irradiance and oak seedling survival and growth in a heterogeneous environment. Forest Ecology and Management 242: 462–469.
Pywell, R.F., J.M. Bullock, D.B. Roy, L.I.Z. Warman, K.J. Walker, and P. Rothery. 2003. Plant traits as predictors of performance in ecological restoration. Journal of Applied Ecology 40: 65–77.
Qian, H., and Y. Jin. 2016. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. Journal of Plant Ecology 9 (2): 233–239. https://doi.org/10.1093/jpe/rtv047.
Rafiee, F., H. Ejtehadi, M. Farzam, H. Zare, and M. Bashirzadeh. 2022. Changes in plant biodiversity facets of rocky outcrops and their surrounding rangelands across precipitation and soil gradients. Scientific Reports 12: 12518.
Rafiee, F., M. Bashirzadeh, H. Zare, H. Ejtehadi, and M. Farzam. 2023. Nurse shrubs influence plant biodiversity across environmental gradients in rocky outcrops and surrounding rangelands. Journal of Vegetation Science 34 (3): e13188.
Ren, H., L. Yang, and N. Liu. 2008. Nurse plant theory and its application in ecological restoration in lower subtropics in China. Progress in Natural Science 18: 137–142.
Rhodes, E.C., J.D. Bates, R.N. Sharp, and K.W. Davies. 2010. Fire effects on cover and dietary resources of sage-grouse habitat. The Journal of Wildlife Management 74: 755–764. https://doi.org/10.2193/2009-143.
Sakschewski, B., W. Von Bloh, A. Boit, A. Rammig, J. Kattge, L. Poorter, J. Peñuelas, and K. Thonicke. 2015. Leaf and stem economics spectra drive diversity of functional plant traits in a dynamic global vegetation model. Global Change Biology 21 (7): 2711–2725. https://doi.org/10.1111/gcb.12870.
Sandell Festin, E., M. Tigabu, M.N. Chileshe, S. Syampungani, and P.C. Odén. 2019. Progresses in restoration of post-mining landscape in Africa. Journal of Forestry Research 30: 381–396. https://doi.org/10.1007/s11676-018-0621-x.
Schnitzler, J., T.G. Barraclough, J.S. Boatwright, P. Goldblatt, J.C. Manning, M.P. Powell, T. Rebelo, and V. Savolainen. 2011. Causes of plant diversification in the Cape biodiversity hotspot of South Africa. Systematic Biology 60: 343–357.
Siles, G., P.J. Rey, J.M. Alcántara, and J.M. Ramírez. 2008. Assessing the long-term contribution of nurse plants to restoration of Mediterranean forests through Markovian models. Journal of Applied Ecology 45: 1790–1798.
Soliveres, S., and F.T. Maestre. 2014. Plant-plant interactions, environmental gradients and plant diversity: A global synthesis of community level studies. Perspectives in Plant Ecology, Evolution and Systematics 16: 154–163.
Soliveres, S., R. Torices, and F.T. Maestre. 2012. Environmental conditions and biotic interactions acting together promote phylogenetic randomness in semi-arid plant communities: New methods help to avoid misleading conclusions. Journal of Vegetation Science 23 (5): 822–836.
Soliveres, S., C. Smit, and F.T. Maestre. 2015. Moving forward on facilitation research: Response to changing environments and effects on the diversity, functioning and evolution of plant communities. Biological Reviews 90 (1): 297–313.
Soliveres, S., F. T. Maestre, M. A. Bowker, R. Torices, J. L. Quero, M. Garcia-Gomez,… Z. Noumi. 2014. Functional traits determine plant co-occurrence more than environment or evolutionary relatedness in global drylands. Perspective in Plant Ecology, Evolution and Systematics 16 (4): 164-173.
Stevens-Rumann, C.S., K.B. Kemp, P.E. Higuera, B.J. Harvey, M.T. Rother, D.C. Donato, P. Morgan, and T.T. Veblen. 2018. Evidence for declining forest resilience to wildfires under climate change. Ecology Letters 21: 243–252. https://doi.org/10.1111/ele.12889.
Svriz, M., M.A. Damascos, H. Zimmermann, and I. Hensen. 2013. The exotic shrub Rosa rubiginosa as a nurse plant. Implications for the restoration of disturbed temperate forests in Patagonia. Argentina. Forest Ecology and Management 289: 234–242.
Swenson, N.G., B.J. Enquist, J. Pither, J. Thompson, and J.K. Zimmerman. 2006. The problem and promise of scale dependency in community phylogenetics. Ecology 87: 2418–2424. https://doi.org/10.1890/0012-9658(2006)87[2418:TPAPOS]2.0.CO;2.
Tepley, A.J., J.R. Thompson, H.E. Epstein, and K.J. Anderson-Teixeira. 2017. Vulnerability to forest loss through altered postfire recovery dynamics in a warming climate in the Klamath Mountains. Global Change Biology 23: 4117–4132.
Tucker, C.M., M.W. Cadotte, S.B. Carvalho, T.J. Davies, S. Ferrier, S.A. Fritz, R. Grenyer, M.R. Helmus, L.S. Jin, A.O. Mooers, S. Pavoine, O. Purschke, D.W. Redding, D.F. Rosauer, M. Winter, and F. Mazel. 2017. A guide to phylogenetic metrics for conservation, community ecology and macroecology. Biological Reviews 92 (2): 698–715. https://doi.org/10.1111/brv.12252.
Valiente-Banuet, A., and M. Verdu. 2007. Facilitation can increase the phylogenetic diversity of plant communities. Ecology Letters 10: 1029–1036. https://doi.org/10.1111/j.1461-0248.2007.01100.x.
Vega-Alvarez, J., J. Garcia-Rodriguez, and L. Cayuela. 2019. Facilitation beyond specie richness. Journal of Ecology 107 (2): 722–734. https://doi.org/10.1111/1365-2745.13072.
Verdu, M., and J.G. Pausas. 2007. Fire drives phylogenetic clustering in Mediterranean Basin woody plant communities. Journal of Ecology 95: 1316–1323.
Webb, C.O., D.D. Ackerly, M.A. McPeek, and M.J. Donoghue. 2002. Phylogenies and community ecology. Annual Review of Ecology and Systematics 33 (1): 475–505. https://doi.org/10.1146/annurev.ecolsys.33.010802.150T.448.
Wilgan, B.W.V., V. Govender, G.G. Forsyth, and T. Krajii. 2011. Towards adaptive fire management for biodiversity conservation: Experience in South African National Parks. Koedoe - African Protected Area Conservation and Science 53 (2): 1–9.
Wortley, L., J. Marc, and M. Howes. 2013. Evaluating Ecological Restoration Success: A Review of the Literature. Restoration Ecology 21 (5): 537–543.
Acknowledgements
The authors wish to thank Iran National Science Foundation (INSF) for supporting the authors in conducting the current research study, and Dr. Richard P. Shefferson for his support in reviewing the paper and his suggestions to improve the quality of the paper.
Funding
This research was done as part of the Post Doctorate Program of M. B. under the supervision of M.F which was financially supported by Iran National Science Foundation: INSF (Grant No. 4000679).
Author information
Authors and Affiliations
Contributions
M. F. designed the study; M.B. and M.F. provided the data; data analyses and preparation of figures and tables were done by M.B.; the paper was written by M.B. and M.A.; and all authors substantially contributed to the subsequent drafts.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Appendix S1.
Characteristics of the studied sites with more of five points: Longitude, Latitude, Altitude, Precipitation and Time since last fire.
Additional file 2: Appendix S2.
A list of plant species present in the communities and their functional trait values.
Additional file 3: Appendix S3.
Functional groups based on cluster analysis on trait matrix for plant species present in studied communities
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bashirzadeh, M., Abedi, M. & Farzam, M. Plant-plant interactions influence post-fire recovery depending on fire history and nurse growth form. fire ecol 20, 9 (2024). https://doi.org/10.1186/s42408-024-00246-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42408-024-00246-2