
Abstract
Background
Arbuscular mycorrhizal fungi (AMF) and Trichoderma harzianum are effective bioagents against Fusarium oxysporum and Verticillium dahliae in tomato plants. The objective of the research was to evaluate the in vivo antagonistic activity of AMF and T. harzianum against Verticillium and Fusarium wilt by enhancing the growth and resistance of tomato plants. A completely randomized experimental design was used, consisting of twelve treatments with nine replicates for each treatment. The treatments included combinations of AMF and T. harzianum inoculation, infection or non-infection by F. oxysporum and V. dahliae, while also considering individual and combined treatments. Mycorrhization rates, growth parameters, disease severity, disease progression, and the impact on disease mitigation were evaluated.
Results
The study revealed the superiority of AMF over T. harzianum, resulting in a significant enhancement in the overall extent of mycorrhizal colonization in tomato plants co-inoculated with T. harzianum. Moreover, AMF treatments and the AMF + T. harzianum consortium contributed to the improvement in growth among all plants infected with V. dahliae and F. oxysporum. Both AMF and T. harzianum significantly reduced the progression of Fusarium wilt, resulting in reductions of 45.14 and 44.91%, respectively, than the untreated plants infected with F. oxysporum (initial disease severity of 75.54%). T. harzianum demonstrated greater efficacy in reducing V. dahliae infection, with a reduction of 34.45% compared to 28.26% for AMF, starting from an initial disease severity of 69.85%. Thus, T. harzianum demonstrated greater effectiveness in controlling disease, particularly Verticillium wilt.
Conclusion
The target application of disease control methods in tomato plants revealed the effectiveness of both AMF and T. harzianum in mitigating Fusarium wilt. Furthermore, T. harzianum demonstrated a higher level of effectiveness against Verticillium wilt. These findings emphasize the potential of AMF and T. harzianum as sustainable alternatives in agriculture, providing a viable option to decrease dependence on fungicides.
Similar content being viewed by others


Background
Crop pests, such as diseases, insects, and weeds, result in substantial annual crop yield losses worldwide, even with the extensive application of chemical pesticides. Nonetheless, the reliance on pesticides poses risks to both human health and the environment, while also fostering the emergence of pesticide-resistant pests (Tudi et al. 2021).
To address these challenges, alternative biological control approaches have emerged, including the utilization of symbiotic endophytic fungi such as arbuscular mycorrhizal fungi (AMF) and certain species of the Trichoderma genus. These approaches have demonstrated their potential in controlling a wide range of plant pathogens, allowing for more environmentally friendly agricultural practices.
AMF, which are classified within the phylum Glomeromycota, play a vital role in plant development by not only stimulating growth but also enhancing the plants resistance against a range of pests (Boutaj et al. 2022). Moreover, commercial products based on Trichoderma harzianum such as T. Trianum (Kthiri et al. 2020) or researched strains like T. virens (Jogaiah et al. 2018) and T. pubescens (Behiry et al. 2023) have shown promising results as biocontrol agents (BCA).
Tomato (Lycopersicon esculentum Mill.) production holds significant importance in Algeria's agricultural economy. Nonetheless, this crop faces considerable susceptibility to a variety of pathogens. On the other hand, climate change and human activities are causing ecosystem alterations, resulting in reduced biodiversity and creating favorable conditions for the proliferation of pathogens (Cavicchioli et al. 2019). Diseases such as Verticillium wilt, caused by the soil-borne fungal pathogen V. dahliae, is particularly challenging to control. However, promising results have been observed in combating this disease with antagonists from the Trichoderma genus (Kong et al. 2022) and AMF (Boutaj et al. 2022).
Similarly, the biological control of Fusarium wilt caused by Fusarium oxysporum is facilitated by some Trichoderma species (Erazo et al. 2021) and AMF (Devi et al. 2022). AMF act as beneficial biocontrol agents, enhancing plant immunity against pathogens, while contributing to plant nutrition and modulating the microbial balance in the mycorrhizosphere (Vishwakarma et al. 2022). Furthermore, certain species of Trichoderma exert control over pathogens by directly impacting them. They stimulate plant innate immunity through the use of natural and synthetic elicitors (Takehara et al. 2023). Trichoderma species also employ mechanisms such mycoparasitism (Panchalingam et al. 2022). To enhance the effectiveness of biocontrol agents, researchers have been investigating the use of BCA consortia to achieve synergistic effects. These consortia often possess multiple biocontrol mechanisms, resulting in enhanced biocontrol outcomes. Furthermore, the coexistence of symbiosis and root pathogenesis within the same plant is a common occurrence in nature. Interestingly, studies have shown that certain strains of Trichoderma have the ability to influence the activity of AMF (Martinez et al. 2004). However, the current understanding of the in vivo interaction between these two processes and their impact on plant development remains limited.
The objective of this study is evaluating the potential of AMF and T. harzianum both individually and in combination, as bio-protectors of tomato plants in vivo, under greenhouse conditions. The focus will be on controlling two common pathogens, Verticillium dahliae and Fusarium oxysporum.
Methods
Tomato material and growth conditions
The experiment was conducted in the greenhouse of Badji Mokhtar University, locality of Sidi Ammar, Annaba province, Algeria (36° 81′ 58′′ N; 7° 71′ 36′′ E). The Rio Grande tomato variety was sterilized with 2% sodium hypochlorite and then sown in autoclaved peat for 1 h at 120 °C, two consecutive times. The seedlings were then placed in individual pots and kept in the greenhouse under daylight conditions at a temperature of 20–25 °C and a relative humidity of 60–70%. The tomato plants were watered twice a week. One-month-old plants were used for the experiment.
Fungal material
Four fungal strains were utilized in the study. Two antagonists, including arbuscular mycorrhizal fungi (AMF) and T. harzianum, as well as the two pathogens V. dahliae and F. oxysporum.
Arbuscular mycorrhizal fungi
The AMF used were provided in the form of a commercial inoculum, Symbivit (INOCULUM plus, France). Symbivit is a granular formulation containing propagules of six AMF species (Claroideoglomus claroideum, Claroideoglomus etunicatum, Funneliformis geosporum, Funneliformis mosseae, Glomus micro-aggregatum and Rhizophagus intraradices) that have been sporulated and stored on an inert support (clay) in the presence of technological additives. The AM fungal inoculum contains 20 infective propagules g−1 (SYMBIVIT Database, July 2023).
Trichoderma harzianum
The T. harzianum strain was provided by Valorization and Conservation of Biological Resources (VALCOR) laboratory, M'Hamed Bougara University of Boumerdes. The T. harzianum isolate was sequenced and identified as Trichoderma harzianum, listed in the GenBank database on NCBI under the code OL587563 (Reghmit et al. 2022). It was characterized by its high in vitro capacity to control V. dahliae. Multiplication of T. harzianum was performed on potato dextrose medium (PDB) supplemented with 50 mg/l streptomycin. T. harzianum spores were added to the culture medium and incubated at 150 rpm and 25 ± 1°C for 7 days. The density of the suspensions was adjusted to 106 CFU/ml.
Fusarium oxysporum and Verticillium dahliae
Strains of the pathogens V. dahliae and F. oxysporum were provided by the VALCOR laboratory. The pathogenicity test for these two species was previously conducted by the VALCOR laboratory, with V. dahliae (R1) tested by Tihar-Benzina et al. (2016) and F. oxysporum tested by Bennacer et al. (2022). Multiplications were carried out separately on potato dextrose medium (PDB) supplemented with 50 mg/l streptomycin, following the same procedure as for T. harzianum, with incubation at 150 rpm and 25 ± 1 °C for a period of 14 days.
In vivo antagonistic assay
The experiment was conducted using a completely randomized block design. Tomato seeds were germinated on January 18, 2020, and the young seedlings were transplanted in two-liter polyethylene bags on February 18, 2020. Each plant was inoculated with 200 infectious fungal propagules of AM, which were deposited near the roots. The application of T. harzianum and both pathogens (V. dahliae and F. oxysporum) was carried out at 13 and 14 weeks from the start of the experiment, respectively. Inoculation of these plants was performed by infecting each tomato plant with 10 ml/plant of a suspension containing V. dahliae and F. oxysporum spores (106 spore ml−1) through single applications. A total of twelve treatments were conducted, each repeated nine times. All the possible treatments, including individual treatment (AMF + V. dahliae, AMF + F. oxysporum, T. harzianum + V. dahliae, T. harzianum + F. oxysporum, and AMF + T. harzianum) as well combination treatments (AMF + T. harzianum + V. dahliae and AMF + T. harzianum + F. oxysporum), were applied using the two bio-inoculants (AMF and T. harzianum). Additionally, one control treatment (without pathogens), positive treatments with pathogens inoculated (V. dahliae and F. oxysporum), and antagonist treatments (AMF, T. harzianum) were conducted.
Plant parameters
After 18 weeks of transplanting, on July 20, 2020, nine tomato plants were considered for measurement and evaluation of the following parameters: mycorrhizal colonization intensity of tomato roots, growth parameters (above-ground and root fresh biomass) and diseases incidence on tomato plants inoculated with V. dahliae and F. oxysporum.
The intensity of mycorrhizal colonization in the root system (M% and A%) was determined using staining technique described by Phillips and Hayman (1970). Root samples from five plants per treatment were used, and observations were made on 30 root fragments of 1 cm. The intensity of colonization of the root cortex was estimated in relation to the entire root system and expressed as a percentage. Arbuscular colonization was also evaluated and expressed as a percentage. The MYCOCALC program and the method described by Trouvelot et al. (1986) were used for annotation and evaluation.
- Growth biomass increase (GB) was calculated for above-ground and root fresh biomass by comparing the mean biomass of plants inoculated with antagonistic agents against control and positive treatments. The calculations were performed for individual treatments (AMF + V. dahliae, AMF + F. oxysporum, T. harzianum + V. dahliae, T. harzianum + F. oxysporum, and AMF + T. harzianum) as well as the combination treatments (AMF + T. harzianum + V. dahliae and AMF + T. harzianum + F. oxysporum). GB was expressed as a percentage and calculated using the formula:
- Length of the branches showing symptoms of diseases (DS), such as yellowing, browning or wilting of the leaves, or the presence of necrosis, was measured at the end of the experiment. Disease incidence (%) was calculated for each plant by the following formula:
Statistical analysis
Intensity and arbuscular colonization of the root system by AMF were assessed according to two factors: infection with V. dahliae or F. oxysporum pathogens in presence or absence of T. harzianum. The data obtained were analyzed using a two-way ANOVA with a general linear model. Post-hoc pairwise comparisons were conducted using Tukey’s honestly significant difference (HSD) test in the R software, version 4.0.2, with the R commander package version 2.8-0 (Downie 2016). The growth parameters were subjected to a three-way ANOVA (AMF * T. harzianum * V. dahliae) and (AMF * T. harzianum * F. oxysporum). Mean comparisons between different treatments were performed at a significance level of 5% using Tukey’s HSD test with R 4.0.2 and Rcmdrpackage, version 2.8-0.
Results
AMF root cortex colonization of tomato plants
Estimation of tomato root colonization by AMF revealed varying levels among the different applied treatments, including AMF, AMF + V. dahliae, AMF + F. oxysporum, AMF + T. harzianum, AMF + T. harzianum + V. dahliae and AMF + T. harzianum + F. oxysporum. The intensity of root cortex colonization was high in all treatments, ranging from 52.09% in the AMF treatment to 67.13% in the AMF + V. dahliae treatment (Fig. 1a). A significant effect was observed between the AMF treatment and all other treatments, except for the AMF + T. harzianum + F. oxysporum treatment. Arbuscules were more abundant in plants inoculated with T. harzianum and infected by V. dahliae or by F. oxysporum (Fig. 1b). The lowest values were recorded in plants co-inoculated with T. harzianum and infected with F. oxysporum (3.2%).

Arbuscular mycorrhizal fungi (AMF) colonization of tomato plants, mean values with different letters are significantly different (P > 0.05), a Arbuscular mycorrhizal fungi (AMF) hyphal root colonization rate, b AMF arbuscular root colonization rate
Biocontrol activity of F. oxysporum and V. dahliae wilt by biostimulation of tomato growth
Above-ground biomass growth
In this study, various treatments were applied to tomato plants to evaluate their effects on above-ground and root biomass. The analysis of mean biomass growth in tomato plants inoculated with AMF, T. harzianum, and AMF + T. harzianum, both in the presence and absence of V. dahliae or F. oxysporum pathogens, showed fluctuating biomass values (Fig. 2). When comparing the above-ground biomass measurements of treated groups to the untreated control group, inoculation with AMF resulted in a significant increase of 52.68%. Similarly, the treatment with T. harzianum improved above-ground biomass, with a gain of 21.25%. However, the combined inoculation of AMF and T. harzianum did not show a significant difference compared to the control.

Effect of arbuscular mycorrhizal fungi (AMF) and Trichoderma harzianum on tomato growth in the presence of fungal disease (Verticillium and Fusarium wilt), mean values with different letters are significantly different (P > 0.05)
In the case of plants infected with V. dahliae, notable growth stimulation was observed in both the AMF and AMF + T. harzianum treated groups. These treatments resulted in significant increases in above-ground biomass, with gains of 50.31 and 52.09%, respectively, when compared to the untreated plants and the positive control group infected with V. dahliae. However, T. harzianum did not have a significant effect on above-ground biomass growth. When considering plants infected with F. oxysporum, the above-ground biomass showed substantial improvements, with gains of 52.94, 23.6, and 32.06% for the AMF, T. harzianum, and AMF + T. harzianum treatments, respectively, as compared to the F. oxysporum-infected plants (Table 1).
Root biomass growth
In terms of root biomass, the AMF + T. harzianum treatment demonstrated the highest effectiveness than the untreated control group. This treatment significantly stimulated root biomass growth, resulting in an increase of 56.62%. When comparing root biomass measurements of plants infected with V. dahliae, the AMF + T. harzianum treatment was the most efficient, resulting in root biomass augmentation of 62.66%. Similarly, the AMF treatment increased root biomass, by 36.27%. However, T. harzianum treatment recorded the lowest value. For the plants infected with F. oxysporum, the best treatments were AMF, AMF + T. harzianum, (13.72 g and 17.38 g, respectively), corresponding to increases of 128.66 and 189.66%. Plants infected with F. oxysporum showed the lowest values (Fig. 2).
Incidence of disease in tomato plants inoculated with V. dahliae and F. oxysporum
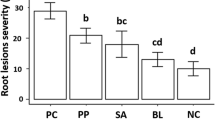
Tomato plants infected with V. dahliae and F. oxysporum without any treatment, showed high disease incidence rates (Fig. 3). However, a mitigation in disease incidence was observed for all the applied treatments, including AMF, T. harzianum, and their synergistic effect (AMF + T. harzianum). For tomato plants infected with V. dahliae, the mean disease incidence rate was 69.85%, with only 30.15% of healthy stems. Notably, maximum protection was observed in tomato plants treated with T. harzianum, which showed a 32.45% decrease in disease incidence. AMF and AMF + T. harzianum treated plants exhibited a disease mitigation, respectively by 28.26 and 30.47%. The combination of AMF and T. harzianum did not show additional effects compared to individual treatments. Furthermore, tomato plants inoculated with F. oxysporum presented the highest mean disease incidence rate, surpassing that of V. dahliae, with 75.54% of affected stems and only 24.46% remaining healthy. However, disease mitigation was observed for AMF, T. harzianum, and AMF + T. harzianum, resulting in decreases of 45.14, 44.91 and 43.62% in disease incidence rates, respectively.

Incidence rate of Verticillium and Fusarium wilt and antagonistic effect of arbuscular mycorrhizal fungi (AMF) and Trichoderma harzianum
Discussion
In this study, co-inoculation of AMF and T. harzianum in greenhouse conditions has been shown to be effective in controlling diseases caused by V. dahliae and F. oxysporum. However, it is important to note that the effectiveness of these symbiotic association in enhancing resistance or tolerance may vary between AMF symbionts and T. harzianum. They probably operate through multiple and various mechanism. AMF ensures protection through competition control. The presence of T. harzianum and both pathogenic fungi led to a significant increase in overall mycorrhizal colonization in the plants. However, plants inoculated solely with AMF displayed the lowest colonization rates. Furthermore, the presence of T. harzianum and the pathogens resulted in a higher intensity of arbuscules. The activation of specific plant defense mechanisms in response to AMF colonization is one of the ways mycorrhizal fungi provide protection to plants. A study showed that by regulating the competitive balance, AMF exclude harmful strains or species that can be detrimental to the plant (Harman et al. 2021). However, it’s worth noting that AMF and soil-borne plant diseases compete for root tissues, which may lead to direct competition for space. Martínez-Medina et al. (2009) reported that multi-taxa AMF inoculation significantly increase root colonization, and the presence of T. harzianum also enhanced AMF colonization. Furthermore, inoculation with T. harzianum showed better results than the colonization by wild populations of microbes alone (Hafiz et al. 2022).
Several studies have demonstrated that increased mineral uptake in AMF colonized plants is associated with greater disease resistance (Li et al. 2013). When nutrient concentrations were high, an accompanying increase in plant biomass was observed (Bhantana et al. 2021). The productivity of plants depends on their overall health and interactions with rhizospheric microorganisms, whether symbiotic or pathogenic. Under the present experimental conditions, in the absence of disease, tomato plants inoculated with AMF and T. harzianum showed improved growth than uninoculated plants, particularly in terms of above-ground biomass. AMF treatment was the most effective. However, co-inoculation (AMF + T. harzianum) did not have a significant effect on above-ground biomass compared to the control, which is consistent with the findings of Molina-Lores et al. (2022). The authors reported the superior performance of AMF (Glomus cubense) in enhancing plant nutrition than T. harzianum and its combined effect (Glomus cubense + T. harzianum), attributing it to structural improvements in the plant, including an increase in stem diameter (Molina-Lores et al. 2017). AMF treatment induced better growth of above-ground and root biomass, even in plants infected with V. dahliae or F. oxysporum leading to improved resistance against these pathogens.
In the case of Fusarium wilt, all treatments (AMF, T. harzianum and AMF + T. harzianum) effectively improved above-ground biomass. For plants affected by Verticillium wilt, the most effective treatment was AMF, either alone or in combination with T. harzianum, resulting in enhanced above-ground biomass compared to T. harzianum treatment. Regarding root systems, T. harzianum did not show any increase in biomass in plants infected with these two diseases. Root biomass was significantly boosted by the AMF + T. harzianum. Mycorrhizae compensated for the loss of biomass and root function caused by pathogen infection by enhancing nutrient and water uptake, indirectly reducing pathogen damage and improving disease resistance in host plants (Ma et al. 2021). Previous studies by Lee and Whang (2016) and Benzina-Tihar et al. (2020) linked poor root system growth in tomato plants inoculated with certain Trichoderma species to the production of indole acetic acid (IAA), which promotes stem and coleoptile growth and inhibits root growth. Kakabouki et al (2021) reported that greater root colonization by arbuscular mycorrhizal fungi following T. harzianum inoculation, suggesting that T. harzianum contributes to the growth-enhancing activity of AMF. Linderman (1994) suggested that microbial antagonists of pathogenic fungi do not interfere with AMF, and may even promote the development of mycosymbionts and facilitate AMF formation.
Furthermore, few studies have demonstrated the sensitivity of a pathogen strain to biological control agents, depending on their modes of action. Buck and Jeffers (2004) reported variability in the susceptibility of pathogenic strains to biocontrol agents that act through nutrient competition. V. dahliae, for instance, was found less susceptible to T. harzianum than AMF that act through nutrient competition.
In this study, the incidence of wilt caused by F. oxysporum was higher than to V. dahliae. The application of AMF and T. harzianum, either alone or in combination, effectively reduced the severity of Verticillium and Fusarium wilt. T. harzianum showed greater effectiveness in reducing Verticillium wilt compared to AMF treatments, while both AMF and T. harzianum demonstrated similar effectiveness against Fusarium wilt. The application of AMF + T. harzianum consortium did not show any additional effect compared to individual treatments. Molina-Lores et al. (2022) stated that the combined presence of T. harzianum, known for its biopesticide effect, and Glomus cubense, known for its bio-fertilizing effect, yields stronger outcomes compared to their individual applications. Linderman (1994) highlighted the potential of harnessing the prophylactic abilities of AM fungi in conjunction with other rhizosphere microorganisms that exhibit antagonistic properties against root pathogens used in biological control practices. This involves utilizing Trichoderma species in combination with mycorrhizae, resulting in a positive and synergistic impact on the overall health of specific plants. Guzmán-Guzmán et al. (2023) have also emphasized that Trichoderma remains the preferred choice in the development of biological control programs.Trichoderma acts directly on the pathogen population in the rhizosphere by releasing metabolites that impact pathogen growth, including extracellular enzymes capable of deteriorating their cell wall (Rao et al. 2016). Furthermore, Trichoderma showcases its protective capabilities through various mechanisms, such as the synthesis of silver nanoparticles and the development of innovative formulations like nano-emulsions, as highlighted by Konappa et al. (2021). Moreover, inoculating plants with T. harzianum elicits a range of defense responses at the histopathological, biochemical, and gene expression levels, effectively providing protection against diseases, as outlined by De Britto and Jogaiah (2022).
Conclusions
The findings further enhance our understanding of the potential of arbuscular mycorrhizal fungi and T. harzianum as biocontrol agents for the management of wilt diseases in tomato plants. Their utilization revealed distinct defense mechanisms. AMF demonstrated a protective effect by increasing the mycorrhizal surfaces, thereby improving plant nutrition and promoting robust growth and ultimately strengthening their resistance against pathogens. On the other hand, T. harzianum acted directly on the pathogens. While all treatments, including the application of AMF, T. harzianum, and their combined synergistic treatment exhibited a slowdown in disease progression for both wilt diseases, the distinct synergistic effect was not prominently observed. This study highlighted the specificity of biocontrol agents in their interaction with pathogens. Both AMF and T. harzianum exhibited effectiveness in combating Fusarium wilt, which demonstrated a higher incidence rate than to Verticillium wilt. Additionally, T. harzianum demonstrated superior control in the case of Verticillium wilt. The results taken together emphasize the potential of AMF and T. harzianum as sustainable alternatives in agriculture, offering a viable solution to reduce reliance on fungicides and promote environmentally and health-conscious practices.
Availability of data and materials
The datasets used and analyzed during the current study are indicated in the manuscript and are available from the corresponding author on reasonable request.
Abbreviations
- AMF:
-
Arbuscular mycorrhizal fungi
- BCA:
-
Biocontrol agents
- DS:
-
Symptoms of diseases
- PDB:
-
Potato dextrose medium
- F. oxysporum :
-
Fusarium oxysporum
- GB:
-
Growth biomass increase
- T. harzianum :
-
Trichoderma harzianum
- VALCOR laboratory:
-
Valorization and conservation of biological resources laboratory
- V. dahliae :
-
Verticillium dahliae
References
Behiry S, Soliman SA, Massoud MA, Abdelbary M, Kordy AM, Abdelkhalek A, Heflish A (2023) Trichoderma pubescens elicit induced systemic resistance in tomato challenged by Rhizoctonia solani. J Fungi 9(2):167
Bennacer A, Sahir-Halouane F, Aitslimane-Aitkaki S, Oukali Z, Oliveira IV, Rahmouni N, Aissaoui M (2022) Structural characterization of phytochemical content, antibacterial, and antifungal activities of Juglans regia L. leaves cultivated in Algeria. Biocatal Agric Biotechnol 40:102304
Benzina-Tihar F, Reghmit A, Oulebsir-Mohandkaci H, Sahir-Halouane F (2020) Les champignons endophytes de l’olivier (Olea europaea cv. europea) du Nord algérien et criblage de leurs activités antagonistes contre le Verticillium dahliae. IOBC-WPRS Bull 151:7–15
Bhantana P, Malla R, Vista SP, Rana MS, Mohamed G, Joshi BD, Hu CX (2021) Use of arbuscular mycorrhizal fungi (AMF) and zinc fertilizers in an adaptation of plant from drought and heat stress. Biomed J Sci Tech Res 38:30357–30373
Boutaj H, Meddich A, Roche J, Mouzeyar S, El Modafar C (2022) The effects of mycorrhizal fungi on vascular wilt diseases. Crop Prot 155:105938
Buck JW, Jeffers SN (2004) Effect of pathogen aggressiveness and vinclozolin on efficacy of Rhodotorula glutinis PM4 against Botrytis cinerea on geranium leaf disks and seedlings. Plant Dis 88(11):1262–1268
Cavicchioli R, Ripple WJ, Timmis KN, Azam F, Bakken LR, Baylis M, Webster NS (2019) Scientists’ warning to humanity: microorganisms and climate change. Nat Rev Microbiol 17(9):569–586
De Britto S, Jogaiah S (2022) Priming with fungal elicitor elicits early signalling defence against leaf spot of broccoli underlying cellular, biochemical and gene expression. Microbiol Res 263:127143
Devi NO, Tombisana Devi RK, Debbarma M, Hajong M, Thokchom S (2022) Effect of endophytic Bacillus and arbuscular mycorrhiza fungi (AMF) against Fusarium wilt of tomato caused by Fusarium oxysporum f. sp. lycopersici. Egypt J Biol Pest Control 32(1):1–14
Downie T (2016) Using the R commander: a point-and-click interface for R. J Stat Softw 75:1–4
Erazo JG, Palacios SA, Pastor N, Giordano FD, Rovera M, Reynoso MM, Torres AM (2021) Biocontrol mechanisms of Trichoderma harzianum ITEM 3636 against peanut brown root rot caused by Fusarium solani RC 386. Biol Control 164:104774
Guzmán-Guzmán P, Kumar A, de los Santos-Villalobos S, Parra-Cota FI, Orozco-Mosqueda MDC, Fadiji AE, Santoyo G (2023) Trichoderma species: our best fungal allies in the biocontrol of plant diseases. Plants 12(3):432
Hafiz FB, Moradtalab N, Goertz S, Rietz S, Dietel K, Rozhon Rozhon W, Humbeck K, Geistlinger J, Neumann G, Schellenberg I (2022) Synergistic effects of a root-endophytic Trichoderma fungus and Bacillus on early root colonization and defence activation against Verticillium longisporum in rapeseed. Mol Plant-Microbe Interact 35(5):380–392
Harman G, Khadka R, Doni F, Uphoff N (2021) Benefits to plant health and productivity from enhancing plant microbial symbionts. Front Plant Sci 11:610065
Jogaiah S, Abdelrahman M, Tran LSP, Ito SI (2018) Different mechanisms of Trichoderma virens-mediated resistance in tomato against Fusarium wilt involve the jasmonic and salicylic acid pathways. Mol Plant Pathol 19(4):870–882
Kakabouki I, Tataridas A, Mavroeidis A, Kousta A, Karydogianni S, Zisi C, Papastylianou P (2021) Effect of colonization of Trichoderma harzianum on growth development and CBD content of hemp (Cannabis sativa L.). Microorganisms 9(3):518
Konappa N, Udayashankar AC, Dhamodaran N, Krishnamurthy S, Jagannath S, Uzma F, Pradeep CK, De Britto S, Chowdappa S, Jogaiah S (2021) Ameliorated antibacterial and antioxidant properties by Trichoderma harzianum mediated green synthesis of silver nanoparticles. Biomolecules 11(4):535
Kong WL, Ni H, Wang WY, Wu XQ (2022) Antifungal effects of volatile organic compounds produced by Trichoderma koningiopsis T2 against Verticillium dahliae. Front Microbiol 13:1013468
Kthiri Z, Jabeur MB, Machraoui M, Gargouri S, Hiba K, Hamada W (2020) Coating seeds with Trichoderma strains promotes plant growth and enhance the systemic resistance against Fusarium crown rot in durum wheat. Egypt J Biol Pest Control 30(1):1–10
Lee JC, Whang KS (2016) Optimization of indole-3-acetic acid (IAA) production by Bacillus megaterium BM5. Korean J Soil Sci Fert 49(5):461–468
Li Y, Liu Z, Hou H, Lei H, Zhu X, Li X, Tian C (2013) Arbuscular mycorrhizal fungi-enhanced resistance against Phytophthora sojae infection on soybean leaves is mediated by a network involving hydrogen peroxide, jasmonic acid and the metabolism of carbon and nitrogen. Acta Physiol Plant 35:3465–3475
Linderman RG (1994) Role of VAM fungi in biocontrol. In: Pfleger FL, Linderman RG (eds), Mycorrhizae and plant health, APS, St. Paul, Minn, pp 1–25
Ma A, Zhang D, Wang G, Wang K, Li Z, Gao Y, Qi J (2021) Verticillium dahliae effector VDAL protects MYB6 from degradation by interacting with PUB25 and PUB26 E3 ligases to enhance Verticillium wilt resistance. Plant Cell 33(12):3675–3699
Martinez A, Obertello M, Pardo A, Ocampo JA, Godeas A (2004) Interactions between Trichoderma pseudokoningii strains and the arbuscular mycorrhizal fungi Glomus mosseae and Gigaspora rosea. Mycorrhiza 14:79–84
Martínez-Medina A, Pascual JA, Lloret E, Roldan A (2009) Interactions between arbuscular mycorrhizal fungi and Trichoderma harzianum and their effects on Fusarium wilt in melon plants grown in seedling nurseries. J Sci Food Agric 89(11):1843–1850
Molina-Lores LB, Montero-Limonta G, Rizo-Mustelier M, Bell-Mesa TD, Vuelta-Lorenzo DR (2017) Influencia de alternativas nutricionales sobre posturas injertadas de mango (mangifera indical) en el vivero sabana ingenio, santiago de cuba. Ciencia En Su PC 2:71–82
Molina-Lores LB, Lussón-Puncet JA, Ávila-Góngora Y, Cutiño-Mendoza A, Bell-Mesa TD (2022) Efecto de Trichoderma harzianum y Glomus cubense en la producción de plántulas de tomate. Revista Transdiciplinaria De Estudios Sociales y Tecnológicos 2(3):42–48
Panchalingam H, Powell D, Adra C, Foster K, Tomlin R, Quigley BL, Kurtböke Dİ (2022) Assessing the various antagonistic mechanisms of Trichoderma strains against the brown root rot pathogen Pyrrhoderma noxium infecting heritage fig trees. J Fungi 8(10):1105
Phillips JM, Hayman DS (1970) Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br Mycol Soc 55:158–161
Rao KLN, Raju KS, Ravisankar H (2016) Cultural conditions on the production of extracellular enzymes by Trichoderma isolates from tobacco rhizosphere. Braz J Microbiol 47:25–32
Reghmit A, Benzina-Tihar F, Djeziri M, Hadjouti R, Oukali Z, Sahir-Halouane F (2022) Effective biofertilizer Trichoderma spp. isolates with enzymatic activity and metabolites enhancing plant growth. Int Microbiol 25(4):817–829
Takehara Y, Fijikawa I, Watanabe A, Yonemura A, Kosaka T, Sakane K, Imada K, Sasaki K, Kajihara H, Sakai S, Mizukami Y, Haider MS, Jogaiah S, Ito SI (2023) Molecular analysis of MgO nanoparticle-induced immunity against Fusarium Wilt in Tomato. Int J Mol Sci 24(3):2941
Tihar-Benzina F, Sahir-Halouane F, Hameed KM (2016) Algerian isolates of fluorescent Pseudomonas spp. as potential biological control against wilt pathogen Verticillium dahliae. Plant Omics 9(1):48–60
Trouvelot A, Kough JL, Gianinazzi-Pearson V (1986) Mesure du taux de mycorhization VA d'un système radiculaire. Recherche de méthodes d'estimation ayant une signification fonctionnelle. In: Gianinazi-Pearson V, Gianinazzi S (eds), Physiology and genetics aspects of mycorrhizae, proceeding of the 1st European symposium on mycorrhizae, Paris, France, pp 217–221
Tudi M, Daniel Ruan H, Wang L, Lyu J, Sadler R, Connell D, Chu C, Phung DT (2021) Agriculture development, pesticide application and its impact on the environment. Int J Environ Res Public Health 18(3):1112
Vishwakarma SK, Ilyas T, Malviya D, Shafi Z, Shahid M, Yadav B, Singh HV (2022) Arbuscular Mycorrhizal Fungi (AMF) as potential biocontrol agents. In: Rhizosphere microbes: biotic stress management. Springer, Singapore, pp 197–222
Acknowledgements
Not applicable.
Funding
No funding.
Author information
Authors and Affiliations
Contributions
MHA contributed in all experiments, data analysis and interpretation and writing the manuscript; MC performed investigation; BF contributed to identification and donor of the fungal material isolates, methodology, and revision of the manuscript; HN contributed to data analysis and graphical display; RN prepared fungi material and performed identification of T. harzianum isolate; ZH wrote the first draft manuscript; and KH performed writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors have read and agreed to the published version of the manuscript.
Competing interests
The authors declare that they have no competing interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Meddad-Hamza, A., Benzina, F., Meddad, C. et al. Biological control of arbuscular mycorrhizal fungi and Trichoderma harzianum against Fusarium oxysporum and Verticillium dahliae induced wilt in tomato plants. Egypt J Biol Pest Control 33, 91 (2023). https://doi.org/10.1186/s41938-023-00737-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-023-00737-5