
Abstract
Background
Three of entomopathogenic fungi, identified as Isaria javanica (Cjc-03 and Cjw-01) and Purpureocillium lilacinum (TS-01) were found naturally infecting the whiteflies, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) on chili and tomato plants. All the isolates were identified by morphological characterization and molecular identification (ITS region amplification). The virulence of the three isolates was evaluated against nymphs of B. tabaci at three different concentrations (1 × 106, 1 × 107, and 1 × 108 conidia/ml) under laboratory conditions and on adults of B. tabaci at one conidia concentration (1 × 107 conidia/ml) under laboratory and glasshouse conditions.
Results
Isaria javanica showed the highest virulence against B. tabaci nymphs and adults in both laboratory and glasshouse conditions. I. javanica exhibited the highest mortality in the laboratory against B. tabaci, nymphs at the highest concentration (1 × 108 conidia/ml) 8 days post-inoculation. Likewise, in the laboratory bioassay, B. tabaci, adults, Cjc-03 isolate exhibited maximum mortality (80.0%), followed by Cjw-01 isolate (77.5%), and TS-01 isolate (65.0%) at the concentration of 1 × 107 conidia/ml at 7 days post-inoculation. The highest mortality rate (63.45%) was recorded by the Cjc-03 isolate and TS-01 (54.22%) isolate at 3 days post-inoculation in glasshouse bioassay using the concentration of 1 × 107 conidia/ml of each of the 2 isolates (Cjc-03 and TS-01) against B. tabaci adults.
Conclusion
Overall, the findings showed that both isolates of I. javanica and P. lilacinum reduced the number of eggs, nymphs, and adults’ emergence of B. tabaci, following the application of 1 × 107 conidia/ml on tomato leaves in the glasshouse. The newly isolated strains could be developed as a potential commercial biopesticide for managing B. tabaci.
Similar content being viewed by others


Background
Sweet potato whitefly, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) is one of the major pests of vegetable and ornamental crops in many tropical and subtropical regions around the world (Singh and Kaur 2020). Adults and nymphs of B. tabaci cause damage to the plant directly during feeding and oviposition and indirectly through the transmission of plant. Moreover, sooty mold colonizes on the surface of plants due to honeydew secreted by B. tabaci affecting photosynthetic activity in plants, resulting in a considerable decrease in agricultural product quality (Shah et al. 2020). Plant viruses transmitted by B. tabaci, such as begomoviruses are estimated to cause yield losses ranging from 20 to 100% and losses worth millions of dollars (Gangwar and Charu 2018). Insecticides have been widely used to control the whitefly, but this approach has several disadvantages, including the development of insecticide resistance and food safety concerns (Gangwar and Charu 2018).
In recent years, there has been increasing research on using biological control agents (BCA) as alternative strategies to control B. tabaci. Over the past few decades, entomopathogenic fungi (EPF) have become one of the major components of integrated pest management (IPM) that are being investigated for the control of several insect pests, and more than 20 species documented to be effective against B. tabaci, including Beauveria bassiana, Isaria fumosoroseus, Metarhizium anisopliae, Ashersonia spp., Isaria javanica, Purpureocillium lilacinum, and Verticillium lecanii (Zafar et al. 2016). Previous findings showed that B. tabaci is susceptible to I. javanica and P. lilacinum infection under laboratory conditions (Du et al. 2021).
Native isolates or EPF strains from various hosts provided unique control programs to manage several indigenous pests because they are more effective to control pests of a particular environment (Sayed et al. 2018). Soil is also a natural habitat for EPF and acts as important reservoir of entomopathogens capable to survive for long time without the host insects. However, it is extremely challenging to isolate new species and strains of EPF from the soil in particular regions due to human activities that results in the frequent genetic connections among soil inhabitants (Dong et al. 2016). Moreover, the discovery of isolates from the insect host, the isolate characterization, and the virulence test are the first steps in producing novel EPF as BCA (Dayanti et al. 2018).
So far, three species of EPF; B. Isaria, and Metarhizium spp. have been discovered from infested insects in Peninsular Malaysia (Kin et al. 2017). Apart from I. fumosorosea documented by Eslamizadeh et al. (2013) in Malaysia, there are unaware other studies of EPF species or isolates isolated from B. tabaci. The effectiveness of this isolate I. fumosorosea isolate (Pf-UPM) was compared with the other nine indigenous Isaria isolates detected from the soil on B. tabaci, and the infectivity rate of Pf-UPM isolate for all life stages was significantly higher than the other nine isolates. Here, two new species of EPF, I. javanica and P. lilacinum from infected B. tabaci were successfully isolated. Therefore, the present study aimed to isolate and identify EPF infecting B. tabaci associated with vegetables as a contribution to the IPM of B. tabaci.
Methods
Isolation of entomopathogenic fungi from the whitefly samples
A survey for the collection of nymphal instars of whitefly was conducted in a glasshouse and field planted with vegetables in the Glasshouse and a research experimental station at the Faculty of Agriculture, UPM. Leaflets of five different types of vegetables (chili, eggplant, tomato, pepper, and okra) infested with nymphs were collected, place in a plastic container (15 × 9 cm) and brought to the laboratory for isolation of EPF. Nymphal instars were observed under a stereomicroscope (Leica Zoom 2000 Stereo Microscope, USA) for probable infection with EPF. To isolate fungal strains, infested leaves with second to fourth instars of B. tabaci were surface sterilized with 70% ethanol for 5 min and washed 5 times with sterile distilled water in a laminar flow. The treated leaves were air-dried on sterile tissue papers for 30 min. The surface-sterilized nymphs from the leaves were then picked using a surface sterile needle and placed directly onto the PDA media supplemented with 50 mg/ml of antibiotic, penicillin, streptomycin, and tetracycline. Two nymphs were placed in the opposite direction on each plate and ten plates were used for each vegetable infested with nymphs. The plates were incubated at 25 ± 2 °C for 12 days and the presence or absence of EPF on the plates inoculated with whitefly nymphs was observed daily until the fungal growth was observed.
Morphological identification of entomopathogenic fungi
The isolated fungal strains were transferred to PDA plates and cultured for 5 days at 25 °C. Pure cultures of the observed fungi were prepared using microscope glass slides. A sterile needle was used to pick fresh mycelia and placed in a single drop of sterile distilled water and/or lactophenol blue mounted on a clean glass slide. The identification of fungal isolates was based on the morphological characteristics as described by Watanabe (2010) and Humber (2012). Morphological features of the colony, conidial shape and size, and mycelia growth rate were studied using a light microscope (Olympus CX31 series, England), equipped with a digital Dinolite eye-piece camera (magnification 40×).
Molecular identification of entomopathogenic fungi
Direct PCR amplification was conducted according to the manufacturer’s protocols for the KOD FX Neo PCR master mix (Toyobo Co. Ltd., Japan). Mycelium of 7 days fungal cultures was scraped, suspended in 100 μl TE buffer and vortexed for 30 s. The fungal ITS region was amplified using the primers ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′ and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) (White et al. 1990). The PCR reaction was performed in 25 μl consisting 2 µl mycelium suspension from each sample, 12.5 μl of 2 × PCR buffer, 5ul dNTP mixture, 1.5 μl of each forward and reverse primers, 0.5 μl of Kod FX Neo,) and ITS4 and 3 µl sterile water. PCR was performed using Thermocycler Biometra (T-Persona, Germany). The PCR reactions were amplified with the following conditions: An initial denaturation temperature of 94 °C for 4 min; 35 cycles of denaturation at 94 °C for 1 min, annealing at 53.7 °C for 1 min, and extension at 72 °C for 2 min; and a final elongation of 72 °C for 10 min. The PCR products were subjected to electrophoresis on a 1.5% agarose gel stained with GelRed™ and run at 90 V. PCR products were visualized under UV illumination and photographed. PCR products were sent to Apical Scientific Sdn. Bhd. Malaysia for purification and direct sequencing. The forward and reverse DNA sequences were manually analyzed to remove the low-quality and ambiguous bases using chromos software. The sequences were identified using the Basic Local Alignment Search Tool (BLAST)n search of GenBank. (http://www.ncbi.nlm.nih.gov). Trimmed Clustal W alignment of sequences, top hit sequences from GenBank, and Metarhizium anisopliae outgroup sequence were used to create a maximum likelihood phylogenetic tree with 1000 bootstrap replicates using MEGA (version 7.0), and whitefly species was identified. The phylogenetic tree for ITS sequences was constructed by the maximum likelihood method with 1000 bootstraps replicates using MEGA version 7 (Kumar and Stecher 2016).
Whitefly colonies and host plant
A stock culture of whiteflies (B. tabaci) which colonized tomato plants in the greenhouse was collected, reared, and maintained in the greenhouse, Ladang 15, Faculty of Agriculture, UPM. The B. tabaci was identified and confirmed by PCR using the mitochondrial DNA cytochrome oxidase I COI (mt COI) gene with the primers (C1-J-2195 and L2-N-3014) as described by Frohlich et al. (1999). The sequencing data were registered and assigned the accession number; OM638559 (GenBank). The infested leaves used for bioassay were obtained by placing whitefly-free plants (21 day-old) close to the adult-infested tomato seedlings for 72 h. Adult whiteflies were then removed, and plants infested with eggs were transferred to another glasshouse and placed in a cage for 12–15 days until the nymphs entered the second instar. This method resulted in more than 40 eggs per leaf (Mascarin et al. 2013). B. tabaci colonies were maintained on tomato seedlings, which planted in 300 l plastic pots held in meshed cages at greenhouse conditions (25–30 °C and 60–80% RH with a photoperiod of 12L:12D light: dark. Tomato plants were checked daily, and the damaged seedlings were replaced with new ones.
Growth of the fungal isolates and production of conidia
Mycelial growth and sporulation of each 3 EPF isolates were examined, following the method described by Ali et al. (2010). Agar plugs (7 × 7 mm) containing mycelia of each EPF pure culture (Cjc-03, Cjw-01, and TS-01) were taken from 12 day-old culture with a cork borer and aseptically inoculated at the center of the Petri plates (90 mm) containing PDA media and incubated at 25 ± 2 °C. Each isolate had four replicates. The mycelial growth of each EPF culture was measured at 3, 5, 7, 9, and 12 days after inoculation (DAI) by measuring the average of two perpendiculars measured diameters of each colony (D1 and D2). To estimate spore concentration, a 7 mm agar disc of actively growing culture was cut using a sterilized cork borer and was placed into a test tube containing 10 ml sterile distilled water. One ml suspension was diluted with 9 ml sterile distilled water using a serial dilution to obtain concentrations of 1 × 106, 1 × 107, and 1 × 108 conidia/ml. The spore suspensions were vortexed for 3 min to obtain a homogeneous solution. For each suspension, spores were counted using a Neubauer hemacytometer (Sumikarsih et al. 2019).
Laboratory bioassay of EPF against B. tabaci nymphs
Leaves infested with second instar nymphs of B. tabaci were excised and dipped into three concentrations: 1 × 106, 1 × 107, and 1 × 108, conidia/ml for 30 s. Nymphal instars were recognized, following Naranjo and Ellsworth, (2017). Third and fourth instars were removed from the leaves by needles, and the number of the second instar was counted and adjusted to 20–25 before dipping into different concentrations. The excised leaves were air-dried and placed on the abaxial surface up in a dishes (90 mm diameter) line with a skinny layer of 1.5% agarose gel (Shah et al. 2020). The control excised leaves were dipped in 0.01% Tween 80, and all the treatments were incubated at 25 ± 2 °C. To prevent saprophytic fungi from growing on whitefly honeydew, aeration was established daily by opening each plate for 30 min (Santiago-Alvarez et al. 2006). Three experiments were conducted evaluating four treatments: (Cjc-03, Cjw-01, TS-01, and 0.01% Tween 80). Dead nymphs (discoloured, and/or desiccated, or developed symptoms with fungal growth or sporulation) were observed using a stereo microscope (Leica Zoom 2000 Stereo Microscope, USA) and mortality was recorded at 3, 5, and 8 days after inoculation. Each treatment consisted of three replicates. The entire experiment was conducted three times using a fresh conidial suspension and a fresh batch of nymphs. Treated whitefly was randomly chosen for confirming whether mortality was caused by the fungal grown on the dead whitefly. After eight days of inoculation, dead whitefly specimens were observed with a stereomicroscope (Leica Zoom 2000 Stereo Microscope, USA), and some of which were processed with a scanning electron microscope (SEM) procedure.
Laboratory bioassay of the EPF against adult whitefly
The experiment was conducted using the method of Mascarin et al. (2013) with minor modifications. Leaf disks (approximately 30 mm) were dipped into 1 × 107 conidia/ml of each fungal isolate and 0.01% tween 80 was served as control. Treated leaf disks were allowed to dry and placed on acrylic vials (77 × 38 mm) containing 10 ml of 1.5% agarose gel. Adult whiteflies ≤ 5 days old were exposed to treated leaf disks and confined using a mesh with a rubber band. The assessment for adult mortality was recorded at 7 DAT. The whole experiment was conducted two times with four treatments: (Cjc-03, Cjw-01, TS-01, and 0.01% Tween 80) and four replicates where each replicate consisted of one leaf disk.
Virulence of EPF against adult whitefly in greenhouse
Tomato seedlings, 30 day old were grown in an 800 l capacity pot. The substrate (sterilized peatmoss) from each pot was covered with aluminum foil to prevent the run-off of fungal suspension.
The seedlings were sprayed with 1 × 107 spore suspension until run-off and placed in a cage (an acrylic aquarium) (40 × 25 cm) held vertically, which serves as a bioassay cage. Control plants were sprayed with sterile distilled water having 0.01% Tween 80. One side of the cage was covered with fine mesh and treated seedlings were kept for 24 h. Following that, 50 to 60 adult whiteflies (≤ 5 days) were manually aspirated from the stock colony and released into the bioassay cage containing treated plants for 3 days. The EPF treatments and control were replicated six times within the greenhouse. Dead whiteflies were counted 3 days after the release of insects, and live insects were removed. To confirm if mycosis was the cause of mortality, all dead insects were placed on PDA plates, incubated for 5–7 days at room temperature, and fungal growth on the insect cadavers was observed.
Effect of EPF isolates on the number of eggs, nymphs, and adults’ emergence of B. tabaci on tomato leaves
The number of eggs laid per one square centimeter of leaflets was counted from each treated plant. Three leaflets per leaf were sampled and two leaves in the midsection of each treated plant were assessed (Barra-Bucarei et al. 2020). Thereafter, the treated seedlings were moved to a new cage and placed in another glasshouse for subsequent development of immature stages and adult emergence. The number of nymphs (instar III and IV) per leaflet was counted at 35 DAT. Eight leaflets (approximately 40 × 20 mm) per treated plant were sampled from four leaves of each treated plant and evaluated in the laboratory. At 42 DAT, adult emergence was calculated by counting the exuviae left by adults’ whitefly on the leaves. Ten leaflets (approximately 40 × 20 mm) per treated plant were sampled from three leaves of each treated plant and evaluated in the laboratory. This bioassay was conducted twice with three treatments: (Cjc-03, TS-01, and 0.01% Tween 80) and six replicates.
Data analysis
All data analyses were performed using R (version 3.6.1). Experimental design in the laboratory bioassay with the second instar nymphs was completely randomized, and responses (B. tabaci mortality) were analysed using a three-way (fungus, concentration, and time) factorial design using Fischer's least significant difference (LSD) test at α = 0.05. The effect of these isolates on adults’ mortality was analysed using one-way ANOVA under a randomized complete design (RCD). For the glasshouse study, adult mortality, the mean number of eggs, nymphs, and adults’ emergence were subjected to an ANOVA test. Mortality data were corrected by using Abbott’s formula (Abbott 1925). Comparisons of the mean treatment were performed using Fischer's least significant difference (LSD) test at α = 0.05.
Results
Fungal identification of entomopathogenic fungi: morphological identification
A total of three fungal isolates of EPF were identified from whitefly infesting two vegetable crops (chili and tomato) in Glasshouse and Research Experimental Station at the Faculty of Agriculture, UPM. The pure culture isolates were grown in PDA and identified based on macro and micro-morphological features. The morphological identification showed that the two isolates were I. javanica and the other isolate was characterized as P. lilacinum.
Isaria javanica
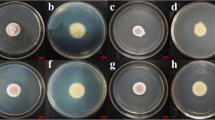
Two fungal isolates (Cjw-01 and Cjc-03) had morphological features related to I. javanica based on colony, colour, sized and conidial shape. On PDA, fungal colony of Cjw-01 was pinkish white cottony, with dense mycelia in the centre, formation of light-yellow pigment with a concentric ring pattern. However, the Cjc-03 colony was white to whitish-yellow (Fig. 1A). The conidia of both isolates were oval to spindle in shape, with a length of about 6 × 2.2 μm (length × width) (Fig. 1D and E).

Colony morphology of Isaria javanica and Purpureocillium lilacinum isolated from infested whitefly (Bemisia tabaci) on PDA media at 25 °C after 10 days of growth A Colony of I. javanica (Cjw-01) B Colony of I. javanica (Cjc-03) C Colony of P. lilacinum (TS-01) D Conidia of I. javanica (Cjw-01) E Conidia of I. javanica (Cjc-03) F Conidia of P. lilacinum (TS-01)
Purpureocillium lilacinum
The colony was round, white in the first three days, and gradually turned pink after 7 days (Fig. 1C). Conidia were oval and single-celled, and the size was 4.2 × 2.1 μm (Fig. 1F). During twelve days of incubation, the radial growth attained 41.15 ± 0.22 mm, 48.50 ± 0.38 mm, and 63.30 ± 0.18 mm for Cjw-01, Cjc-03, and TS-01 isolates respectively. The sporulation; 1.27 × 106, 1.38 × 106, and 3.99 × 106 conidia/ml were also recorded on Cjw-01, Cjc-03, and TS-01 isolates, respectively.
Molecular identification
Polymerase chain reaction amplification and DNA sequencing results indicated that the rDNA‐ITS gene of Cjw-01, Cjc-03, and TS-01 isolates were 594 bp (Fig. 2), following this the DNA sequence were submitted to GenBank; where the GenBank accession number MW857170, MW857169, and MW857173, respectively. BLASTn results in GenBank revealed that the ITS gene of Cjw-01 and Cjc-03 isolates were 99.81% homologous to the species I. javanica ex-type strain (GenBank accession number AY624186). Moreover, phylogenetic analysis showed that the isolate was closely clustered in the I. javanica clade (Fig. 3) this confirmed the results of the morphological identification that the Cjw-01 CJc-03 isolates were I. javanica strain. Sequencing of the ITS placed the TS-01 isolate in the species P. lilacinum and the ITS sequence showed 99% similarity with strain P. lilacinum in the GenBank (Table 1). However, phylogenetic analysis revealed that the two isolates (Cjc-03 and Cjw-01) and one isolate (TS-01) were closely clustered in the I. javanica and P. lilacinum clade, respectively, confirming the morphological identification of the I. javanica and P. lilacinum isolates.

PCR amplification from ITS region and amplification fragment showing approximately 594 bp. Lane M: 1 kb ladder; Lane 1–3: Cjw-01, Cjc-3, TS-01, Lane 4: Negative control (distilled water)

Maximum likelihood (ML) tree of Isaria javanica isolates and Purpureocillium lilacinum isolate based on ITS region. Metarhizium anisopliae was used as an outgroup
The tree was constructed by the Maximum Likelihood method based on the Kimura 2-parameter model (Kimura 1980).
Virulence of EPF isolates against second nymphal instar of B. tabaci in the laboratory
The result of the present study showed that all the three EPF strains exhibited a significant effect on the second instar nymphs of B. tabaci (F = 686.40, p < 0.01). However, the factorial analysis of variance yields significant effects of fungal concentration, time interval, and their interaction on the mean mortality of B. tabaci (F = .99.15, p > 0.001; F = 382.70, p > 0.001; F = 4.60, p > 0.05) (Table 2).
At 8 DAT, the highest mortality recorded in the highest concentration, 1 × 108 conidia/ml, was 91.1% in Cjc-03 isolate, 76.5% in Cjw-01 isolate and 62.7% in TS-01 isolate, and low mortality observed at the lowest concentration 1 × 106 conidia/ml were 67.7%, 55.6% and 42.89% for Cjc-03, Cjw-01 and TS-01, respectively. Accordingly, at the 5 DAT, the maximum mortality recorded (84.4, 61.3 and 55.6%) at the highest concentration 1 × 108 conidia/ml, while the mortality recorded at the lowest concentration, 1 × 106 conidia/ml (51.1, 35.6 and 35.36%) for Cjc-03, Cjw-01 and TS-01, respectively. On the other hand, the maximum mortality observed at the 3 DAT in the highest concentration 1 × 108 conidia/ml was 47.7% for Cjc-03 isolate, 39.4% for Cjw-01 isolate, and 33.3% for TS-01 isolate, and minimum mortality was recorded at the lowest concentration 1 × 106 conidia/ml Cjc-03 isolate (33.5%), Cjw-01 isolate (26.7%) and TS-01 isolate (14.8%) (Fig. 4). In all treatments, I. javanica isolates shown to be the most virulent against B. tabaci at 3rd, 5th and 8th day after infection.

Mortality (%) of Bemisia tabaci at different time intervals (3, 5, and 8 DAT) with 3 fungal isolates Isaria javanica, (Cjc-03, Cjw-01), and Purpureocillium lilacinum (TS-01) at three concentrations of each fungal isolates and control treatment. The column shows the % mortality. Treatment columns with different alphabets are significantly different from other treatments (LSD test at p = 0.05)
Based on the scanning electron microscope (SEM) analysis of the dead whitefly, it was shown that the conidia are attached and the mycelium penetrated to whitefly's body. Scanning electron microscopy of I. javanica isolates showed dense mycelium growing on the integument and on the cuticle surface of whitefly (Fig. 5). Furthermore, a thickening of the margins of the germ tubes was observed during fungal penetration, indicating the formation of appressoria (an infective structure) (Fig. 5D). The infection of healthy B. tabaci second instar nymphs treated with I. javanica and P. lilacinum were also observed under a stereomicroscope before and after the application (Fig. 6).

Scanning electron microscopy (SEM) of the nymph of Bemisia tabaci infected with Isaria javanica at 8 days post-inoculation A Conidia of I. javanica attached to the body of B. tabaci B Germinated conidia of I. javanica C Mycelium growing on the integument of infected nymph D Hyphal penetration of I. javanica into the integument of an infected nymph of B. tabaci

The infection of Bemisia tabaci second instar nymphs treated with Isaria javanica and Purpureocillium lilacinum observed under a stereomicroscope A Healthy whitefly nymph before application B B. tabaci nymph after 8 days of infection by I. javanica; C: B. tabaci nymph after 8 days of infection by P. lilacinum
Virulence of EPF isolates against adult B. tabaci in the laboratory
Each one of the two isolates of I. javanica, (Cjc-03 and Cjw-01) and one isolate of P. lilacinum (TS-01) tested at a single conidia concentration showed pathogenicity to adults of B. tabaci, caused more than 50% mortality, following the 7th day of infection. There was non-significant difference among the three isolates (Cjc-03, Cjw-01, and TS-01), but significantly differed than control treatment (F = 22.56 p < 0.001) (Fig. 7). The highest mortality (80.0%) was recorded for Cjc-03 isolate (Fig. 7) and I. javanica was the most effective against adults of B. tabaci.

Mortality of adult Bemisia tabaci at 7 DAT when treated with Isaria javanica (Cjc-03, Cjw-01) and Purpureocillium lilacinum (TS-01) at 1 × 107conidia/ml concentration. Different letters above the bars indicate statistical significance (p < 0.05, LSD test)
Virulence of EPF isolates against B. tabaci adults on tomato leaves at 3 days after treatment in the glasshouse
In the glasshouse, the virulence of isolated fungi against adult whitefly showed that the two isolates, I. javanica, (Cjc-03) and P. lilacinum (TS-01) had significantly suppressed the adults of B. tabaci with a single conidia concentration (i.e., 1 × 107) (F = 27.22 p < 0.001). When B. tabaci adults infected with I. javanica and P. lilacinum, the mortality rates were greater than 50% on the 3rd day after treatment (Fig. 8). Results revealed that the infection and pathogenicity of I. javanica isolate to B. tabaci, in glasshouse conditions, was higher than those with P. lilacinum.

Mortality (%) of Bemisia tabaci at 3 DAT when treated with 2 isolates (Cjc-03 and TS-01) at 1 × 107conidia/ml concentration. Different letters above the bars indicate statistical significance (p < 0.05)
Effect of EPF isolates on the number of eggs, nymphs, and adults’ emergence on tomato leaves in the glasshouse
The effect of I. javanica (Cjc-03) and P. lilacinum (TS-01) on the number of eggs, nymphs, and adults’ emergence were examined on the tomato leaves following application on adult B. tabaci. There were significant differences in the number of eggs observed in the leaves from the treated plant and control leaves at 3 DAT (F = 17.76 p = 0.0001) (Fig. 9). The seedlings treated with Cjc-03 and TS-01 exhibited the least number of eggs with means of (8.66 and 11.66 eggs) (N cm2 leaflet), respectively, when compared to seedlings of the control treatment 20.83 eggs (Fig. 9).

Mean numbers (± SE) of Bemisia tabaci eggs on tomato leaves when infected with Isaria javanica and Purpureocillium lilacinum at 3 DAT
As for the number of the nymphs, it was observed that tomato plants treated with a single conidia concentration (i.e., 1 × 107 conidia/ml) reduced the numbers of nymphs of B. tabaci on the leaves after six weeks of treatment as compared to the control. A significantly high mean number was recorded in the control treatment 6.5 nymphs in comparison to the plants treated with Cjc-03 and TS-0 with means of 1.33 and 2.5 nymphs (N cm2 leaflet), respectively (F = 36.05 p < 0.001) (Fig. 10). The adult emergence rate was also recorded from the exuviae left on the tomato leaves (leaflet 4 × 2 cm) of the treated plant at 42 days after application of single conidia concentration on adult B. tabaci. A significantly high mean number of adults emerged was recorded in the control treatment 18.66 adults in comparison to the plants treated with Cjc-03 and TS-0 with means of 8.33 and 11.83 adults (4 × 2 leaflet), respectively (F = 4.52 p = 0.029) (Fig. 10).

Effect of Isaria javanica (Cjc-03) and Purpureocillium lilacinum (TS-01) on the mean number of nymphs and adults of Bemisia tabaci A: nymphs (III and IV) B: adult emergence of B. tabaci on tomato leaves at 42 DAT
Discussion
In the present study, three EPF isolates, I. javanica (Cjw-01 and Cjc-03), and P. lilacinum (TS-01) were isolated and identified from immature B. tabaci individuals. Previous findings revealed that I. javanica and P. lilacinum could be isolated from infected B. tabaci and other insect pests and also potentially control many vegetable insect pests (Du et al. 2021). However, these two species were not previously isolated from insects in Malaysia and neighbouring countries. The first step in developing mycoinsecticides for the control of insect pests is to investigate them from a specific host. The isolation, identification, and virulence test of the fungal isolates are the important steps in developing EPF as BCA (Dayanti et al. 2018). Shah et al. (2020) had been previously isolated strains of I. javanica from the cadaver of B. tabaci sampled from the field of eggplant in Oman. Moreover, I. javanica had been isolated from several agricultural insect pests such as Asian citrus psyllid, Diaphorina citri Kuwayama, (Ou et al. 2019); Sunn pests (Eurygaster spp.); (Gül et al. 2021). P. lilacinum is well known to infect nematodes and little is known about its effect on insects pests (Toledo-Hernández et al. 2019). It was previously detected from mycoses instar of some certain insect pests such as Antiteuchus innocens Englemand & Rolston (Hemiptera: Pentatomidae) (Toledo-Hernández et al. 2019). To our knowledge, this is the new isolate of P. lilacinum isolated from field infested by B. tabaci. This is the fact that Purpureocillium spp., was a new divergent genus that was established in 2011 (Luangsa-Ard et al. 2011), and was more frequent and commonly isolated from soil samples as has been observed in several previous studies (Sun et al. 2021).
The growth rate of the three isolates was studied under laboratory conditions. Insect biocontrol depends on the fungal strain with good growth characteristics (Meng et al. 2017). All the three isolates grew optimally and produced spores that are similar and better than those previously reported (Du et al. 2021). Several studies have demonstrated the effectiveness of I. javanica and P. lilacinum for the control of B. tabaci (Sun et al. 2021). In Malaysia, the application of EPF as biological control of pests is currently in its early phase of evaluation. The study has been mainly focused on M. anisopliae. Despite the potentials of I. javanica and P. lilacinum as a BCA for a variety of insect pests (Sun et al. 2021), there has been little or no information on their use for the control of insect pests in Malaysia, compared to M. anisopliae and other species of EPF.
Based on the result of the laboratory study, both nymphs and adults B. tabaci were susceptible to all the three isolates (Cjw-01, Cjc-03, and TS-01). It is discernible from this study that second nymphal instar of B. tabaci were the most susceptible to infection than adults’ stage, confirming previous reports that the second instar of B. tabaci was highly susceptible to EPF infection ( Sain et al. 2019). The mortality rate of the second nymphal instar varied depending on the concentration of spores and period of application. It was noted that mortality increased by increasing the concentration of conidia and the time interval between applications. This result is in accordance with Abdulle et al. (2020) findings who demonstrated the effectiveness of three isolates of L. lecanii against B. tabaci under laboratory conditions and caused the highest mortality up to 93% in the highest concentration (1 × 107 conidia/ml) at 7 days post inoculation (dpi). Accordingly, Wu et al. (2020) reported the significant mortality of P. xylostella upon application of newly isolated Isaria spp. A significant effect of temperature, conidia concentration, and period of exposure in causing mortality of B. tabaci by B. bassiana and L. lecanii was observed by Keerio et al. (2020).
With the same conidia concentration and exposure time, it was observed that second instar mortality rate was higher in case of the I. javanica treatment than in the P. lilacinum treatment. This might be due to the variation in mechanical damage and production of toxins resulting in insect death (Singh et al. 2017). For example, Wu et al. (2021), conducted experiment to determine the virulence of I. javanica and other strains of B. bassiana and M. anisopliae against the 4th nymphal instar of B. tabaci. A strain of I. javanica was found to exhibit high virulence among all the strains tested.
Moreover, the single application of conidial concentration (1 × 107 conidia/ml) on adult B. tabaci resulted in more than 50% mortality for seven days post inoculation from all three isolates. A similar trend was also observed in previous research with P. lilacinum application. Glasshouse experiments are essential to confirm laboratory findings before moving on with full-scale commercial development (Cuthbertson et al. 2007). In this study, the glasshouse bioassay of one isolate each of I. javanica and P. lilacinum (Cjw-03 and TS-01) with single conidia concentration (1 × 107 conidia/ml) against adults B. tabaci caused mortality 63.5 and 54.2% over a period of 72 h, respectively.
The fungi evaluated significantly decreased the number of eggs and nymphs than control treatment. The conidial concentration of the EPF sprayed on the leaves was discovered to colonize a host plant completely, and retrieved from different parts (roots and shoots), demonstrating their potential to protect the entire plant from insect pests with a single application (Bamisile et al. 2019).
Furthermore, the adult emergence rate observed at six weeks after treatment was significantly lower in the seedlings treated with Cjc-03 and TS-01 isolates than to the control treatment. Other studies, e.g. Zhu and Kim (2011) reported that the application of fungal isolates, Beauveria. bassiana, I. fumosorosea, and L. lecanii against B. tabaci on eggplant significantly reduced the adults’ emergence of B. tabaci.
Conclusion
Three EPF isolates, I. javanica, (Cjw-01 and Cjc-03), and P. lilacinum (TS-01) were isolated and identified from immature B. tabaci individuals. All three EPF strains showed pathogenicity on the adults and immature stages of B. tabaci where the reduction numbers of eggs, nymphs and adults’ emergence were recorded. The locally isolated strains of I. javanica and P. lilacinum could be developed as potential biopesticides against B. tabaci although the development of proper formulation is still required.
Availability of data and materials
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Abbott WS (1925) A method of computing the effectiveness of an insecticides. J Econ Entomol 18:265–267
Abdulle YA, Nazir T, Keerio AU, Ali H, Zaman S, Anwar T (2020) In vitro virulence of three Lecanicillium lecanii strains against the whitefly, Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae). Egypt J Biol Pest Control 30(129):1–6
Ali S, Wu J, Huang Z, Ren SX (2010) Production and regulation of extracellular chitinase from the entomopathogenic fungus Isaria fumosorosea. Biocontrol Sci Technol 20(7):723–738. https://doi.org/10.1080/09583151003714091
Bamisile BS, Dash CK, Akutse KS, Qasim M, Aguila LCR, Wang F, Keppanan R, Wang L (2019) Endophytic Beauveria bassiana in foliar-treated citrus limon plants acting as a growth suppressor to three successive generations of Diaphorina citri kuwayama (Hemiptera: Liviidae). InSects. https://doi.org/10.3390/insects10060176
Barra-Bucarei L, González MG, Iglesias AF, Aguayo GS, Peñalosa MG, Vera PV (2020) Beauveria bassiana multifunction as an endophyte: growth promotion and biologic control of Trialeurodes vaporariorum, (westwood) (hemiptera: Aleyrodidae) in tomato. InSects 11(9):1–15. https://doi.org/10.3390/insects11090591
Cuthbertson AGS, Walters KFA, Northing P, Luo W (2007) Efficacy of the entomopathogenic nematode, Steinernema feltiae, against sweetpotato whitefly Bemisia tabaci (Homoptera: Aleyrodidae) under laboratory and glasshouse conditions. Bull Entomol Res 97(1):9–14. https://doi.org/10.1017/S0007485307004701
Dayanti AK, Sholahuddin YA, Subositi D (2018) First record of entomopathogenic fungi on autumn leaf caterpillar (Doleschallia bisaltide). IOP Conf Ser Earth Environ Sci. https://doi.org/10.1088/1755-1315/142/1/012040
Dong T, Zhang B, Jiang Y, Hu Q (2016) Isolation and classification of fungal whitefly entomopathogens from soils of Qinghai-Tibet Plateau and Gansu Corridor in China. PLoS ONE 11(5):1–12. https://doi.org/10.1371/journal.pone.0156087
Du C, Wu J, Cuthbertson AGS, Hamid Bashir M, Sun T, Ali S (2021) Morphological, molecular and virulence characterisation of six Cordyceps spp. isolates infecting the diamondback moth Pluttela xylostella. Biocontrol Sci Technol 31(4):373–386. https://doi.org/10.1080/09583157.2020.1854175
Eslamizadeh R, Sajap AS, Omar D, Adam N (2013) First Record of Isaria fumosorosea Wize (Deuteromycotina: Hyphomycetes) Infecting Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in Malaysia. J Entomol 10(4):182–190
Frohlich DR, Torres-Jerez I, Bedford ID, Markham PG, Brown JK (1999) A phylogeographical analysis of the Bemisia tabaci species complex based on mitochondrial DNA markers. Mol Ecol 8(10):1683–1691. https://doi.org/10.1046/j.1365-294X.1999.00754.x
Gangwar RK, Charu G (2018) Lifecycle, distribution, nature of damage and economic importance of whitefly, Bemisia tabaci (Gennadius). Acta Sci Agric 2(4):36–39
Gül E, Babaroğlu NE, Demirci F (2021) Characterization and virulence of entomopathogenic fungi from sunn pests in Turkey. J Asia Pac Entomol 24(2):215–223. https://doi.org/10.1016/j.aspen.2021.02.007
Humber RA (2012) Chapter VI Identification of entomopathogenic fungi. In: Lacey LA (ed) Manual of techniques in invertebrate pathology, 2nd edn. Academic Press, San Diego, CA, USA, pp 151–187
Keerio AU, Nazir T, Abdulle YA, Jatoi GH, Gadhi MA, Anwar T, Sokea T, Qiu D (2020) In vitro pathogenicity of the fungi Beauveria bassiana and Lecanicillium lecanii at different temperatures against the whitefly, Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae). Egypt J Biol Pest Control. https://doi.org/10.1186/s41938-020-00247-8
Kimura M (1980) A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 16:111–120
Kin PK, Azmi WA, Kamarudin N, Ali SRA, Moslim R (2017) The Occurrence of entomopathogenic fungi on mineral and peat soils in Peninsular Malaysia. Am J Agric Biol Sci 12(1):1–12. https://doi.org/10.3844/ajabssp.2017.1.12
Kumar S, Stecher GTK (2016) MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol 33:1870–1874
Luangsa-Ard J, Houbraken J, van Doorn T, Hong SB, Borman AM, Hywel-Jones NL, Samson RA (2011) Purpureocillium, a new genus for the medically important Paecilomyces lilacinus. FEMS Microbiol Lett 321(2):141–149. https://doi.org/10.1111/j.1574-6968.2011.02322.x
Mascarin GM, Kobori NN, Quintela ED, Delalibera I (2013) The virulence of entomopathogenic fungi against Bemisia tabaci biotype B (Hemiptera: Aleyrodidae) and their conidial production using solid substrate fermentation. Biol Control 66(3):209–218. https://doi.org/10.1016/j.biocontrol.2013.05.001
Meng X, Hu J, Ouyang G (2017) The isolation and identification of pathogenic fungi from Tessaratoma papillosa Drury (Hemiptera: Tessaratomidae). PeerJ 5:e3888. https://doi.org/10.7717/peerj.3888
Naranjo SE, Ellsworth PC (2017) Methodology for developing life tables for sessile insects in the field using the whitefly, Bemisia tabaci, in cotton as a model system. J vis Exp 129:1–12. https://doi.org/10.3791/56150
Ou D, Zhang LH, Guo CF, Chen XS, Ali S, Qiu BL (2019) Identification of a new Cordyceps javanica fungus isolate and its toxicity evaluation against Asian citrus psyllid. Microbiologyopen 8(6):1–9. https://doi.org/10.1002/mbo3.760
Sain SK, Monga D, Kumar R, Nagrale DT, Kranthi S, Kranthi KR (2019) Comparative effectiveness of bioassay methods in identifying the most virulent entomopathogenic fungal strains to control Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). Egypt J Biol Pest Control 29(31):1–11. https://doi.org/10.1186/s41938-019-0130-z
Santiago-Alvarez C, Maranhaõ EA, Maranhaõ E, Quesada-Moraga E (2006) Host plant influences pathogenicity of Beauveria bassiana to Bemisia tabaci and its sporulation on cadavers. Biocontrol 51:519–532. https://doi.org/10.1007/s10526-005-5737-1
Sayed SM, Ali EF, El-Arnaouty SA, Mahmoud SF, Amer SA (2018) Isolation, identification, and molecular diversity of indigenous isolates of Beauveria bassiana from Taif region, Saudi Arabia. Egypt J Biol Pest Control 28(1):1–6. https://doi.org/10.1186/s41938-018-0054-z
Shah R, Al-Sadi AM, Nasser Al-Sabahi J, Al-Raeesi AA, Khamis Said Al-Rawahi K, Saud Al-Rashdi A, FadhilMadad Al-Hinai S, Velazhahan R (2020) Efficacy of an Omani strain of Cordyceps javanica and its culture filtrate against whitefly (Bemisia tabaci) under laboratory conditions. All Life 13(1):615–622. https://doi.org/10.1080/26895293.2020.1835742
Singh H, Kaur T (2020) Pathogenicity of entomopathogenic fungi against the aphid and the whitefly species on crops grown under greenhouse conditions in India. Egypt J Biol Pest Control. https://doi.org/10.1186/s41938-020-00287-0
Singh D, Raina TK, Singh J (2017) Entomopathogenic fungi: an effective biocontrol agent for management of insect populations naturally. J Pharm Sci Res 9(6):830–839
Sumikarsih E, Herlinda S, Pujiastuti Y (2019) Conidial density and viability of Beauveria bassiana isolates from Java and Sumatra and their virulence against Nilaparvata lugens at different temperatures. AGRIVITA. J Agric Sci 41(2):335–350. https://doi.org/10.17503/agrivita.v41i2.2105
Sun T, Wu J, Ali S (2021) Morphological and molecular identification of four Purpureocillium isolates and evaluating their efficacy against the sweet potato whitefly, Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae). Egypt J Biol Pest Control 31(1):0–8. https://doi.org/10.1186/s41938-021-00372-y
Toledo-Hernández RA, Toledo J, Valle-Mora J, Holguín-Meléndez F, Liedo P, Huerta-Palacios G (2019) Pathogenicity and virulence of Purpureocillium lilacinum (Hypocreales: Ophiocordycipitaceae) on Mexican fruit fly adults. Florida Entomol 102(2):309–314. https://doi.org/10.1653/024.102.0204
Watanabe T (2010) Pictorial atlas of soil and seed fungi morphologies of cultured fungi and key to species, 3rd edn. CRC Press, London
White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols a guide to methods of applications. Academic Press, San Diego, pp 315–322
Wu S, Toews MD, Oliveira-Hofman C, Behle RW, Simmons AM, Shapiro-Ilan DI (2020) Environmental tolerance of entomopathogenic fungi: a new strain of Cordyceps javanica isolated from a whitefly epizootic versus commercial fungal strains. Insects 11(10):1–15. https://doi.org/10.3390/insects11100711
Wu S, Toews MD, Castrillo LA, Barman AK, Cottrell TE, Shapiro-Ilan DI (2021) Identification and virulence of Cordyceps javanica strain wf GA17 isolated from a natural fungal population in sweetpotato whiteflies (Hemiptera: Aleyrodidae). Environ Entomol. https://doi.org/10.1093/ee/nvab061
Zafar J, Freed S, Khan BA, Farooq M (2016) Effectiveness of Beauveria bassiana against cotton whitefly, Bemisia tabaci (Gennadius) (Aleyrodidae: Homoptera) on different host plants. Pakistan J Zool 48(1):91–99
Zhu H, Kim JJ (2011) Susceptibility of the tobacco whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae) biotype Q to entomopathogenic fungi. Biocontrol Sci Technol 21(12):1471–1483. https://doi.org/10.1080/09583157.2011.636482
Acknowledgements
The authors would like to express their gratitude to the Tertiary Education Trust Fund (TETFund), Nigeria for providing Ph.D. scholarship and Universiti Putra Malaysia (UPM) for the general support during the study.
Funding
This study was funded by the Fundamental Research Grant Scheme (FRGS) from the Ministry of Higher Education Malaysia (Grant No. 5540212) (FRGS/1/2019/WAB01/UPM/02/36).
Author information
Authors and Affiliations
Contributions
IS analyzed and interpreted the data regarding the virulence of entomopathogenic fungi, Isaria javanica and Purpureocillium lilacinum and was a major contributor in writing the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sani, I., Jamian, S., Saad, N. et al. Identification and virulence of entomopathogenic fungi, Isaria javanica and Purpureocillium lilacinum isolated from the whitefly, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in Malaysia. Egypt J Biol Pest Control 33, 14 (2023). https://doi.org/10.1186/s41938-023-00657-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-023-00657-4