
Abstract
Background
Brown spot disease is one of the most destructive foliar diseases of rice. Biological control of this disease is considered more environmentally safe, but the biological control agents have limited availability. The fungus Trichoderma can be used as a biocontrol agent for various plant diseases, including brown spots. Indigenous Trichoderma species from West Timor were evaluated in this study to reveal their effectiveness in reducing brown spot disease of upland rice. The study was conducted under a screen house condition using a factorial treatment design laid out in a completely randomized design. The two factors were the brown spot disease control method (T. viride, T. harzianum, T. hamatum, fungicide Trivia 73 WP, and a control treatment) and upland rice genotype (Inpago 7 and Breun Senaren 2).
Results
The results showed that Trichoderma spp. and the fungicide Trivia 73 WP caused significantly lower AUDPC and apparent infection rate than the control treatment. The total phenol content and agronomic characters (number of reproductive tillers per plant, number of filled grains per panicle, grain yield per plant) were higher in the Trichoderma and fungicide treatments than the control treatment. Meanwhile, tannin and saponin levels were non-significantly different among treatments. Trichoderma and the fungicide Trivia treatments effectively suppressed brown spot disease of upland rice, and the efficacy was dependent on the rice variety, with a mean efficacy range over the two varieties of 27.05–36.42%. The Trichoderma and the fungicide Trivia treatments increased the grain yield per plant differentially in the 2 rice varieties, with a mean grain yield increase over the 2 varieties of 13.0% (T. viride), 23.5% (T. harzianum), 35.0% (T. hamatum), and 19.9% (Trivia 73 WP).
Conclusion
Efficacy of local Trichoderma of West Timor and the fungicide Trivia treatments were varietal dependent with T. hamatum being the most effective in reducing the brown spot disease and producing the highest grain yield increase over the 2 rice varieties. The local Trichoderma species of West Timor have a future potential as biocontrol agents of upland rice brown spot disease, but it is necessary to further evaluate their effectiveness against the disease in replicated field trials.
Similar content being viewed by others

Background
Leaf brown spot caused by the fungus Drechslera oryzae (Breda de Haan), Subram & Jain [syn. Helminthosporium oryzae, Bipolaris oryzae (Breda de Haan) Shoemaker], is one of the primary diseases of upland rice that causes high yield losses (IRRI 2009).
In East Nusa Tenggara (ENT) Province, Indonesia, leaf brown spot has long been associated with upland rice farming. The traditional cultivation practices and the poor dry land soil nutrition are the factors that cause the disease to occur more frequently. Consequently, most of the local upland rice cultivars in the region are susceptible and moderately susceptible to leaf brown spot disease, and the yield loss caused by the disease ranged from 10.46 to 56.15% (Mau et al. 2020). Leaf brown spot is, therefore, considered one of the factors contributing to the low upland rice productivity (2 t/ha) in ENT Province, while under controlled and optimum field conditions, the local cultivars can produce a grain yield of 5–6 t/ha (Ndiwa and Mau 2019). Therefore, seeking effective control measures for leaf brown spot disease is necessary to prevent high yield losses in upland rice and hence increase rice productivity.
Brown spot disease can be controlled using various methods (Asghar et al. 2019), including chemical spray, disease-free seeds, use of resistant varieties, biological control, etc. Resistant varieties are the most economical and environmentally sound control measure (Magar 2015). However, the brown spot resistant varieties are limited, and the resistance is frequently short-lived due to the frequent emergence of new virulent races (Arshad et al. 2008). Thus, an integrated control strategy that involves a more environmentally friendly method, such as biological control agents, also needs to be implemented to tackle the disease effectively. For example, the fungus Trichoderma sp. is a biological control agent proven to control various plant diseases effectively.
Trichoderma is a highly active fungus in the rhizosphere, soil, and the plant canopy environment, and it can also be a parasite to other fungi (Harman et al. 2004). Trichoderma produces several types of extracellular proteins and compounds that are fungi toxic and fungi-static to several fungal pathogens and also can stimulate plant growth, so this fungus has received serious attention as a biological control agent (Puyam 2016).
The ability of Trichoderma to suppress pathogen and disease development occurs through various mechanisms such as competition (Khalili et al. 2012), mycoparasitism (Dix and Webster 1995), antibiosis (Khalili et al. 2012) and also by inducing the host plant’s resistance (Levy et al. 2015) and promoting the host plant growth (Hassan et al. 2017). Induction of host plant resistance can occur through the production of secondary metabolites such as phenol, saponin, and tannin (Yanti et al. 2018), while plant growth promotion may occur through the production of plant growth hormones, etc. (Alison and Robert 2014).
In an in vitro preliminary study, that several indigenous Trichoderma species of West Timor were found inhibiting the aerial growth of the pathogenic fungus D. oryzae with an efficacy of 35–50% (Mau et al. 2021). The results indicated the potential of local Trichoderma isolates as biological control agents of rice brown spot disease in rice. Evaluating these local Trichoderma isolates along with a fungicide spray on different upland rice varieties may lead to finding more effective ways of controlling brown spot disease in upland rice. The objectives of this study were to elucidate the effectiveness of indigenous Trichoderma species of West Timor against the brown spot disease on two upland rice genotypes and observe their effects on the agronomic characters on the tested upland rice genotypes.
Methods
Study locations and materials
The study was conducted at the Plant Disease Laboratory, Faculty of Agriculture, Universitas Nusa Cendana, and in a screen house in Lasiana Village, Kupang District, East Nusa Tenggara Province, Indonesia. The study was conducted from April to November 2021. Research materials used in the study included T. viride, T. harzianum, T hamatum (obtained from a collection of Plant Disease Laboratory, Universitas Nusa Cendana), D. oryzae, the fungicide Trivia 73 WP, and 2 Indonesian upland rice varieties, i.e., Inpago 7 and Breun Senaren 2.
Experimental design
The experiment employed a completely randomized design consisting of a factorial treatment design of 2 factors, i.e., upland rice genotype (G) and disease control method (C). The upland rice genotype consisted of 2 varieties (Inpago 7 and Breun Senaren 2), while the disease control method consisted of 5 treatments, i.e., T. viride, T. harzianum, T. hamatum, the fungicide Trivia 73 WP (active ingredients: Fluopicolide 6% and Propineb 66.7%), and a control treatment (without disease control). A total of 15 treatment combinations were evaluated, consisting of 3 replicates.
Procedures
Plant cultivation
Rice plants were grown in polybags of 40 cm × 30 cm (height × width) containing soil taken from the surrounding uncultivated field. The soil was taken from 1 – 20 cm depth, cleaned from the debris, and air-dried for 3 days before being finely ground and filtered to obtain a uniform soil particle. Next, 10 kg air-dried filtered soil was placed into a polybag and then watered to a field capacity level. Four upland rice seeds were planted in each polybag, and only one plant was retained from 2 weeks of age until harvest. A standard upland rice agronomic practice was applied throughout the plant growing cycle. All the plants were maintained in a screen house made from a transparent para net to allow maximum sunlight intensity.
Preparation of Trichoderma solution and foliar application of the treatment
Before being used in the experiment, the Trichoderma culture was purified by a single spore method on a potato dextrose agar (PDA) medium for a week at room temperature. Then, the Trichoderma was re-identified to confirm their identity by comparing their colony appearance and microscopic characters with the manual description provided by Kubicek dan Harman (2002) and Watanabe (2002). The treatments were applied to the rice plants in a liquid formulation of Trichoderma and fungicide through foliar spraying. The Trichoderma isolate was grown on a potato dextrose agar (PDA) growth medium for 7 days. Then, 5 disks (Ɵ = 0.6 cm) of the Trichoderma were taken and placed into a 250-ml flask containing 100 ml PD broth medium. The flasks containing the PD broth culture were then continuously shaken at room temperature for 7 days at 150 rpm. The colonized PD broth medium was filtered using a 0.45 µm mesh membrane paper. The filtered filtrate was adjusted to a final concentration of 108 conidia/mL and used as a working solution. The fungicide Trivia 73 WP working solution was prepared by dissolving one gram of the fungicide powder into 1 L distilled water and shaken thoroughly to obtain a homogenous working solution with a final concentration of 1 g/L.
Trichoderma and the fungicide Trivia 73 WP were administrated by foliar spray 19 days after planting (2 days before plant inoculation using D. oryzae). The working solutions of Trichoderma and fungicide Trivia were sprayed onto the rice plant's leaves until runoff occurred. The foliar application was carried out at around 8 to 9 am. The treatment applications were conducted 4 times on a two-weekly basis. Plants that received no Trichoderma and fungicide sprays served as the control (no disease control treatment).
Preparation of D. oryzae suspension and plant inoculation
The fungus D. oryzae was obtained from infected rice fields. The diseased leaf disks were cultured on a PDA medium for 7 days, and a single spore was isolated and further grown on a PDA medium for another 7 days under room temperature for purification and identification of D. oryzae. Identification was done by comparing the colony appearance and microscopic characters with the authentic description provided by Ellis (1971) and Agarwal et al. (1989). A PDA medium was used to propagate the D. oryzae isolate for plant inoculation. After 14 days, the fungus mycelia mats were gently scraped with a spatula and added with 10 ml sterile water containing 0.02% Tween 20 to obtain the conidial suspension. The conidial concentration of the stock solution was determined using a hemocytometer and then adjusted to 1 × 106 conidia/ml for plant inoculation. Plant inoculation was conducted 21 days after planting (2 days after foliar sprays of Trichoderma sp. and the fungicide). All tested plants were inoculated by foliar spraying until runoff occurred. Inoculation was done from 8 to 9 am.
Observation
Observation was done on individual plants, including disease severity, phenol content, tannin and saponin levels, the number of reproductive tillers per plant, the number of filled grains per panicle, and grain yield per plant. Disease severity was assessed every week from 7 days after inoculation until the early flowering stage. The disease was assessed based on a disease scale according to the standard evaluation system of IRRI (2013) and was then used to calculate the disease severity level using the formula:
where DS = disease severity, n = number of leaves with a certain disease score, v = disease score, Z = the highest observed disease score, and N = the total number of leaves observed in each plant. The mean disease severity of each plant was averaged from all leaves in the plant.
All disease severity recordings were used to calculate the area under the disease progress curve (AUDPC) based on the formula of Campbell and Madden (1990).
where Yi = disease severity at the ith observation, n = the last disease assessment (number of assessment), and ti = time period of assessment at the ith observation.
The mean apparent infection rate was calculated based on the time-varying apparent rate (Madden et al. 2007) as follows:
where yt = disease severity at the tth observation, and tt = time period of assessment at the tth observation.
Total phenol content and tannin and saponin levels were determined on leaves of the tested rice plants 3 days before the final disease assessment. Five grams of fresh leaf sample of each plant was taken and used for the analysis in the Laboratory of Biosains, Universitas Nusa Cendana. Total phenol content was determined based on the method of McDonald et al. (2001), while saponin and tannin levels were qualitatively assessed based on the method of Syahadat and Azis (2012).
Agronomic data were recorded at harvest. The number of reproductive tillers was obtained by counting the reproductive tiller per plant, while the number of grains per panicle was averaged from the number of grains of all panicles within a plant. The grain yield was measured as grain weight (± 14% moisture content) per plant.
Data analysis
Quantitative data such as AUDPC, apparent infection rate, total phenol content, and agronomical data were subjected to ANOVA followed by an LSD post hoc test, while saponin and tannin levels were descriptively analyzed. Correlation analysis was performed to see the associations among variables. The ANOVA was performed using GenStat 12th (VSN International 2009), while the correlation analysis was conducted using PAST ver. 3 (Hammer et al. 2001).
Results
Brown disease severity and efficacy of the treatments
Ten recordings of brown spot disease severity were made during the study. The disease symptoms started to appear from the first week after inoculation. At the last assessment, the disease severity ranged on the 2 tested varieties which were 19.1–26.6 and 19.6–26.1%, respectively.
Data in Table 1 summarize the results of factorial ANOVA of epidemiological and agronomical parameters. The single factor genotype and disease control method significantly or highly significantly affected the AUDPC and apparent infection rate. In contrast, the interaction between the 2 factors caused non-significant effect on AUDPC but significantly affected the apparent infection rate.
Data (*) in Table 2 show significant AUDPC among treatments within every single factor of rice genotype (G) and disease control method (C). The overall mean AUDPC over the 2 varieties (mean G) was highest in the control treatment (1345.03%.day), while the lowest was in T. hamatum (855.20%.day), but did not differ significantly from that of T. harzianum (908.59%.day). Consistent with the mean genotype, the highest AUDPC within each genotype was observed on the control treatment, and the lowest was on T. hamatum and T. harzianum applications. The overall mean AUDPC of the disease control method factor was higher in Breun Senaren 2 (1093.41%.day) than Inpago 7 (926.68%.day). Within each disease control method, Inpago 7 consistently showed lower AUDPC than Breun Senaren 2, indicating that the former had a higher brown spot resistance than the latter.
Apparent infection rates also varied significantly among treatments. The treatment combination of Breun Senaren 2 × T. viride showed the lowest apparent infection rate (0.0077 disease/unit/day), while the highest was on Inpago 7 × control treatment (0.0343 disease/unit/day). On average, the apparent infection rate in Inpago 7 (0.019 disease/unit/day) was higher than Breun Senaren 2 (0.016 disease/unit/day). The mean apparent infection rate of the disease control method over the 2 rice genotypes was lowest in T. harzianum (0.0093 disease/unit/day), and the highest was on the control treatment (0.0297 disease/unit/day).
Data in Table 2 also show that the disease control method using Trichoderma sp. and fungicide sprays effectively reduced the disease AUDPC than the control treatment. The efficacy of the Trichoderma and the fungicide Trivia varied significantly between the 2 rice varieties. Over the 2 varieties, the highest efficacy (36.42%) was obtained from T. hamatum spray. In comparison, the effectiveness of the other 3 treatments was 27.05% (T. viride), 28.62% (fungicide Trivia 73 WP), and 32.45% (T. harzianum), which did not significantly differ from each other. Efficacy of Trichoderma and the fungicide Trivia was statistically similar on Inpago 7 but was significantly different on Breun Senaren 2 with T. hamatum the most effective.
Phenol, tannin, and saponin levels
Total phenol content was highly significantly affected by the interaction of disease control method and rice genotype (Table 1). Similarly, the single factor treatments showed a highly significant effect on the total phenol content. Data in Table 3 show that the treatment combination of T. hamatum × Inpago 7 produced the highest phenol content (2.40 mg GAE/g), followed by T. harzianum × Inpago 7 (2.07 mg GAE/g) and the fungicide Trivia 73 WP × Breun Senaren 2 (2.00 mg GAE/g) in the second place. Meanwhile, the lowest phenol content was recorded on the control × Inpago 7 treatment combination (1.42 mg GAE/g). On the overall mean of the 2 varieties, T. harzianum spray produced the highest phenol content (2.01 mg GAE/g), which was significantly higher than other treatments, while the lowest was observed on the control treatment (1.61 mg GAE/g). The mean phenol content of the the five disease control treatments over the two varieties differed significatly from each other. The overall mean phenol content of disease control method treatment was higher in Inpago 7 (1.91 mg GAE/g) than Breun Senaren 2 (1.76 mg GAE/g). In contrast to total phenol content, tannin was equally produced in all treatment combinations at a high level in both varieties (Table 3). Meanwhile, saponin was produced in low to medium levels in both varieties. The control treatment had a low level of saponin in both varieties, which was also similar to the treatment combinations of T. hamatum × Inpago 7, T. viride × Breun Senaren 2, and T. harzianum × Breun Senaren 2, while the rest treatment combinations produced medium levels of saponin.
Number of reproductive tiller per plant and number of filled grain per panicle
Interaction of rice variety (G) and disease control method (C) caused non-significant effect on the number of reproductive tillers per plant. In contrast, both the single factor treatments caused a highly significant effect on the trait (Table 1). Mean reproductive tillers per plant over the 2 genotypes were highest in the fungicide Trivia 73 WP (21.33 tillers), but it was non-significantly different from Trichoderma sp. applications (Table 4). In comparison, the lowest number of reproductive tillers was recorded on the control treatment (18.50 tillers).
On average, the number of reproductive tillers per plant of Inpago 7 (24.47) was higher than Breun Senaren 2 (15.47 tillers). A similar situation did happen within each disease control method (Table 4).
Interaction of disease control method and rice variety highly affected (P < 0.01) the number of filled grains per panicle (Table 1). The highest number of grain per panicle was observed on the treatment combination of Trivia 73 WP × Inpago 7 (140.33 grains), followed by T. harzianum × Inpago 7 (130.44 grains) and Trivia 73 WP × Breun Senaren 2 (121.22 grains) in the second and the third places, respectively (Table 4). Meanwhile, the lowest number of grains per panicle was observed on the treatment combination of no disease control × Inpago 7 (109.44 grains) and no disease control × Breun Senaren 2 (101.44 grains) (Table 4).
On the overall mean of the 2 varieties, the fungicide spray using Trivia 73 WP produced the highest number of grains per panicle (131.56 grains), followed by T. viride (119.78 grains) in the second, and T. harzianum and T. hamatum in the third place. On the other hand, the control treatment produced the lowest number of grains per panicle (105.44 grains). On the overall means of disease control method, Inpago 7 produced a higher number of grains per panicle (122.67 grains) than Breun Senaren 2 (110.40 grains) (Table 4).
Grain yield per plant and increase (%) of grain yield per plant
The ANOVA results revealed that the interaction between the disease control method and rice genotype significantly affected grain yield per plant (Table 1). Mean grain yield per plant is presented in Table 4.
On the Inpago 7 genotype, T. harzianum produced the highest mean grain yield/plant (25.50 g), followed by T. hamatum (22.50 g) and Trivia 73 WP (21.17 g). In contrast, the control treatment produced the lowest grain yield/plant (17.83 g). T. hamatum had the highest mean grain yield plant/plant on Breun Senaren 2 (22.33 g), followed by T. viride (19.0 g), Trivia 73 WP (18,67 g), T. harzianum (16.33 g), and the control treatment (15.67 g). On the overall mean of the two varieties, T. hamatum produced the highest grain yield/plant (22.42 g), followed by T. harzianum (20.92 g), Trivia 73 WP (19.92 g), and T. viride (18.75 g). The control treatment produced the lowest grain/plant (16.75 g).
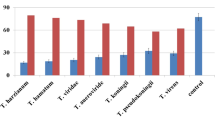
The results of grain yield per plant indicate that foliar applications of Trichoderma species and fungicide Trivia 73 WP could increase the grain yield of tested upland rice varieties compared to no disease control treatment. The percentage of grain yield increase is presented in Fig. 1, where the percentage of grain increase caused by Trichoderma and Trivia fungicide applications was dependent on the variety. On Inpago 7, T. hamatum caused the highest increase of grain yield (42.8%), while on Breun Senaren 2, T. hamatum produced the highest increase of grain yield per plat (42.9%). Over the 2 varieties, the percentage of grain yield increase was highest on T. hamatum, while T. viride, T. harzianum, and Trivia 73 WP caused a statistically similar percentage of grain yield per plant.

Mean percentage of grain yield increase under combined treatments of disease control method and upland rice variety. Lowercase denotes comparison among disease control methods within the same variety or mean variety; uppercase denotes comparison between Inpago 7 and Breun Senaren 2. Values/bars within the same group with the identical lowercase or uppercase are not significantly different based on the LSD (0.05) post hoc test
Correlation analysis
Correlation analysis was performed to see the association among the studied variables, employing the replicated variable’s data of the treatment combinations (Table 5). AUDPC had a positive correlation with apparent infection rate (r = 0.47), and it had a negative correlation with the reproductive tiller (r = − 0.49), the number of filled grain (r = − 0.49), and grain yield (r = − 0.55), but it had non-significant correlation with phenol content (r = − 0.33). The apparent infection rate was negatively correlated with phenol content (r = − 0.49) and AUDPC, but it did not correlate with other variables. Total phenol had no association with other variables except with apparent infection rate (a negative correlation).
The number of reproductive tillers correlated positively with both the number of filled grain (r = 0. 53) and grain yield (r = 0.49), but it negatively correlated with AUDPC. Grain yield per plant correlated positively with both the number of filled grains per panicle (r = 0.49) and the number of filled grain per panicle (r = 0.541), but it was negatively correlated with AUDPC (Table 5).
Discussion
In this study, foliar applications of local Trichoderma species and the fungicide Trivia 73 WP were investigated for their effectiveness in suppressing the brown leaf spot in two genotypes of upland rice. The disease control treatments using Trichoderma sp. and fungicide treatments were able to suppress the leaf brown spot disease progress on two upland rice varieties as indicated by their lower AUDPC than in the control treatment. In addition, the lower AUDPC in the Trichoderma sp. and the fungicide applications was consistent with the lower mean apparent infection rate of these treatments than the control treatment. The efficacy of Trichoderma sp. and the fungicide treatments ranged from 27.05 to 36.42%. Foliar application of T. hamatum was the most effective against the disease, while T. viride and T. harzianum were as effective as fungicide spray using Trivia 73 WP. The ability of Trichoderma sp. to suppress leaf brown spot in the current study is consistent with previous study results by Khalili et al. (2012). The results also confirmed the effectiveness of Trichoderma application against blast diseases of rice (Chou et al. 2020) and various crop diseases (Soesanto et al. 2019).
Results of the present study revealed that total phenol content was higher in plants receiving Trichoderma applications than in the control treatment. The results also showed that Trichoderma application reduced the AUDPC and apparent infection rate, supported by the positive correlation between the 2 parameters (r = 0.47). Meanwhile, there was a significantly negative correlation between phenol content and the apparent infection rate (r = − 0.49), indicating the involvement of phenol in suppressing the disease progression rate. Although previous studies had reported phenol involvement in induced resistance against D. oryzae (Hassan et al. (2017), the novelty of the present study results is that phenol production of local Trichoderma of West Timor and Trivia applications was varietal dependent. This suggests that phenol involvement in suppressing leaf brown spots depends on the rice variety and the Trichoderma species, implying different mechanisms of host–pathogen interaction, resulting in various resistance levels.
All treatment combinations produced similarly high-level tannin content, indicating that tannin may not involve in the host plant response against D. oryzae and, presumably, the high-level tannin content was produced constitutively, which is in line with Dixon (2001). In contrast, Yanti et al. (2018) reported the involvement of tannin in resistance against dieback disease (Botryodiplodia theobromae) of cocoa seedlings. Unlike tannin, saponin was produced in low to medium levels with no apparent pattern among rice genotype and disease control treatments, which renders its involvement in resistance to brown spot disease inconclusive. Nevertheless, previous studies demonstrated that saponin is involved in biochemical resistance against various plant diseases (Yanti et al. 2018).
The number of productive tillers per plant was negatively correlated (r = − 0.49) with AUDPC, indicating that an increase in AUDPC will be followed by a decrease in productive tiller number and vice versa. Presumably, the increase in AUDPC caused a decline in the plant's photosynthetic capacity since the brown spot symptoms mainly occurred on rice leaves, which reduces the photosynthetic area of the plant. This will, in turn, cause a decline in the number of reproductive tillers (Dariush et al. 2020). The ability of Trichoderma and fungicide applications to produce a higher number of tillers than the control treatment was presumably related to the lower AUDPC, and hence a higher leaf photosynthetic capacity they possessed. The genetic factor may contribute to the significant difference between varieties in the reproductive tiller number as the ability to produce reproductive tiller is varietal dependent (Ndiwa and Mau 2019).
The treatment combinations highly affected the number of filled grains, implying that this trait is highly variable among the disease control treatments and the rice variety. The differences in disease severity may cause different photosynthetic capacities of the rice plants, which resulted in a significant grain number per panicle among treatments. A previous study by Hassan et al. (2017) showed that Trichoderma foliar application reduced the brown spot infection and caused an increase in agronomic characters, including the number of discolored (filled) grain.
The grain yield was affected significantly by the disease control method and rice genotype interaction, which may reflect the genetic potential of the tested rice genotypes. The expression of this genetic potential is further influenced by environmental factors, i.e., disease control treatments using Trichoderma and the fungicide. The grain yield of the 2 varieties in the Trichoderma and fungicide treatments was significantly higher than the control treatment. The lower grain yield in the control treatment may be caused by the decrease in both reproductive tiller number and number of grains per panicle as the direct effect of leaf infection by the disease, which reduced the photosynthetic capacity of rice plants (Pantha et al. 2017). This presumption is supported by the negatively significant correlation between AUDPC with, respectively, grain yield (r = − 0.55), the number of the reproductive tiller (r = − 0.49), the number of filled grains per panicle (r = − 0.46), and the positive correlation between grain yield with, respectively, the number of the reproductive tiller (r = 0.49) and the number of filled grains (r = 0.54).
A previous study by Hassan et al. (2017) showed that Trichoderma foliar application reduced the brown spot infection but increased the agronomic characters (leaf chlorophyll content, plant height, panicle length, the number of filled grain, and grain yield) as compared to the non-treated plants. The ability of Trichoderma to improve the agronomic characters is consistent with the previous findings that Trichoderma can stimulate the host plant growth (Hassan et al. 2017).
The present study results revealed that the efficacy of Trichoderma and fungicide treatments against brown spot ranged from 27.05 to 36.42%, while the grain yield of the plants receiving the treatments was about 13–35% higher than the control. This discrepancy indicated that the grain yield observed in this study may reflect the genetic potential of the tested rice genotypes, which may or may not be directly related to leaf brown spot severity. The higher grain yield in the Trichoderma-treated plants than the control might be occurred through both the reduced leaf brown spot infection and the mechanism of plant growth stimulation. These included an increased number of reproductive tillers and filled grain. As a comparison, previous study (Dariush et al. 2020) confirmed that brown spot infection reduced the grain yield by reducing the reproductive tiller number and the number of filled grains. However, the host plant’s growth stimulation was not examined physiologically in the present study, which is the limitation of the current study that needs to be considered in future works.
Conclusions
The indigenous Trichoderma species isolated from West Timor and the fungicide Trivia 73 WP effectively reduced brown spot disease of upland rice, and the efficacy was dependent on the rice variety, with a mean efficacy range over the 2 varieties from 27.05 to 36.42%. The Trichoderma and the fungicide Trivia treatments increased the grain yield per plant differentially in the 2 rice varieties, with a mean grain yield increase over the 2 varieties of 13.0% (T. viride), 23.5% (T. harzianum), 35.0% (T. hamatum), and 19.9% (Trivia 73 WP). The local Trichoderma species of West Timor have a future potential as biocontrol agents of brown spot disease of upland rice as they were able to suppress the disease and increase the grain yield of upland rice under a screen house condition. However, before being further employed as potential biological agents for brown spot disease, future works are needed to evaluate the effectiveness of this local Trichoderma against the disease in replicated field trials.
Availability of data and materials
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- ENT:
-
East Nusa Tenggara
- WP:
-
Wettable powder
- DS:
-
Disease severity
- AUDPC:
-
Area under the disease progress curve
- ANOVA:
-
Analysis of variance
- AIR:
-
Apparent infection rate
- TPC:
-
Total phenol content
- NRTP:
-
Number of the reproductive tiller per plant
- NFGP:
-
Number of filled grain per panicle
- GYP:
-
Grain yield per plant
- LSD:
-
Least significant difference
References
Agarwal PC, Mortensen CN, Mathur SB (1989) Seed-borne diseases and seed health testing of rice. Phytopathol Pap 30:106
Alison S, Robert H (2014) Applications of Trichoderma in plant growth promotion. In: Biotechnology and biology of Trichoderma, pp 415–428. https://doi.org/10.1016/B978-0-444-59576-8.00031-X
Arshad HMI, Khan JA, Jamil FF (2008) Screening of rice germplasm against blast and brown spot diseases. Pak J Phytopathol 20(1):52–57
Asghar M, Baig MMQ, Chaudhary S, Iqbal MF, Ali MA (2019) Evaluation of difenoconazole along with macronutrients spray for the control of brown leaf spot (Bipolaris oryzae) disease in rice (Oryza sativa) crop. Sarhad J Agric 35(1):1–6. https://doi.org/10.17582/journal.sja/2019/35.1.1.6
Campbell CL, Madden LV (1990) Introduction to plant disease epidemiology. Wiley, New York
Chou C, Castilla N, Hadi B, Tanaka T, Chiba S, Sato I (2020) Rice blast management in Cambodian rice fields using Trichoderma harzianum and a resistant variety. Crop Prot 135:104864. https://doi.org/10.1016/j.cropro.2019.104864
Dariush S, Darvishnia M, Ebadi AA, Padasht-Dehkaei F, Bazgir E (2020) Screening rice genotypes for brown spot resistance along with yield attributing characters and its association with morphological traits. J Crop Prot 9(3):381–393
Dix NJ, Webster J (1995) Fungal ecology. Chapman & Hall
Dixon RA (2001) Natural products and plant disease resistance. Nature 411:843–847
Ellis MB (1971) Dematiaceous hyphomycetes. CMI, Kew, England.
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. http://palaeo-electronica.org/2001_1/past/issue1_01.htm. Accessed 26 Oct 2019
Harman GE, Howell CR, Viterbo A, Chet I, Lorito M (2004) Trichoderma species—opportunistic, avirulent plant symbionts. Nat Rev 2:43–56
Hassan AA, Kalboush ZA, Saleh MM (2017) Efficacy of Trichoderma spp as biocontrol agents against rice brown spot disease and biochemical approaches. J Plant Prot Path Mansoura Univ 8(12):677–686
IRRI (2009) Brown spot (rice facts sheet). International Rice Research Institute, Los Banos
IRRI (2013) Standard evaluation system for rice, 5th edition. International Rice Research Institute, Los Banos
Khalili E, Sadravi M, Naeimi S (2012) Biological control of rice brown spot with native isolates of three Trichoderma species. Braz J Microbiol 43(1):297–305. https://doi.org/10.1590/S1517-838220120001000035
Kubicek CP, Harman GE (2002) Trichoderma and Gliocladium. Vol. 1. Basic Biology, Taxonomy and Genetics. Taylor & Francis
Levy NO, Harel YM, Haile ZM, Elad Y, Rav-David E, Jurkevitch E, Katan J (2015) Induced resistance to foliar diseases by soil solarization and Trichoderma harzianum. Plant Pathol 64(2):365–374. https://doi.org/10.1111/ppa.12255
Magar PB (2015) Screening of rice varieties against brown leaf spot disease at Jyotinagar, Chitwan, Nepal. Intl J Appl Sci Biotechnol 3(1):56–60. https://doi.org/10.3126/ijasbt.v3i1.12014
Mau YS, Ndiwa ASS, Oematan SS (2020) Brown spot disease severity, yield and yield loss relationships in pigmented upland rice from East Nusa Tenggara, Indonesia. Biodiversitas 21(4):2625–2634
Mau YS, Hahuly MV, Henuk JBD, Widinugraheni S (2021) Efficacy of Trichoderma species from West Timor against Dreschlera oryzae in the laboratory. Research Report. Research and Community Service Institute, Universitas Nusa Cendana, Indonesia (in Indonesian)
McDonald S, Prenzler PD, Autolovich M, Robards K (2001) Phenolic content and antioxidant activity of olive extracts. Food Chem 73:73–84. https://doi.org/10.1016/S0308-8146(00)00288-0
Ndiwa ASS, Mau YS (2019) Yield and yield component performances of local pigmented upland rice cultivars from East Nusa Tenggara, Indonesia in three locations. Int J Trop Drylands 3(2):49–55
Pantha P, Shrestha SM, Manandhar HK, Gaire SP, Aryal L, Yadav DR (2017) Evaluation of rice genotypes for resistance against brown spot disease caused by Bipolaris oryzae. Int J Curr Res 9(04):48562–48569
Puyam A (2016) Advent of Trichoderma as a bio-control agent—a review. J Appl Nat Sci 8(2):1100–1109. https://doi.org/10.31018/jans.v8i2.927
Soesanto L, Mugiastuti E, Manan A (2019) Raw secondary metabolites application of two Trichoderma harzianum isolates towards vascular streak dieback on cocoa seedlings. Pelita Perkebunan 35(1):22–32 ((in Indonesian))
Syahadat RM, Aziz SA (2012) Effect of media composition and fertigation with organic fertilizer to bioactive contained of orange jassmine (Murraya paniculata (L.) Jack) leaf in seedling. Bull Littro 23(2):142–147 ((in Indonesian))
VSN International (2009) GenStat for windows, 12th edn. VSN International Ltd., Hertfordshire
Watanabe T (2002) Pictorial atlas of soil and seed fungi: morphologies of cultured fungi and key to species, 2nd edn. CRC Press LLC, Boca Raton
Yanti LA, Achmad A, Khumaida N (2018) Resistance mechanisms of white jabon seedlings (Anthocephalus cadamba) against Botryodiplodia theobromae causing dieback disease. Biodiversitas 19(4):1441–1459. https://doi.org/10.13057/biodiv/d190434
Acknowledgements
We are very grateful to the Laboratory of Plant Disease, Universitas Nusa Cendana, for providing the Trichoderma isolates, the Indonesian Rice Research Institute for providing the upland rice seeds of Inpago 7 variety, and East Flores Department of Agriculture for providing the seeds of Breun Senaren 2 variety. We extend special thanks to Mrs. Rina Sula for her excellent assistance in the laboratory work.
Funding
This research was funded by Universitas Nusa Cendana through the professorship research scheme year 2021 (Contract number: 178/PP/2021).
Author information
Authors and Affiliations
Contributions
YSM organized the research team, provided the research supplies, and contributed to all experiment stages. RSP, KDN, and HK contributed to all stages of the experiment. JBDH and SW assisted in designing the experiment, performed laboratory analysis, and performed data analysis and interpretation. MVH assisted in data analysis and interpretation and assisted in drafting and finalizing the manuscript, and YRYG assisted in designing the experiment and assisted in data analysis and interpretation. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
The manuscript has not been published completely or in part elsewhere.
Competing interests
The authors declare no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mau, Y.S., Prayetno, R.S., Kaka, H. et al. Efficacy of indigenous Trichoderma isolates of West Timor, Indonesia, as biocontrol agents of brown spot (Drechslera oryzae) on two upland rice varieties. Egypt J Biol Pest Control 32, 62 (2022). https://doi.org/10.1186/s41938-022-00559-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-022-00559-x