
Abstract
Osteoarthritis (OA) is a degenerative joint disease that is common among the middle-aged and older populations, causes patients to experience recurrent pain in their joints and negatively affects their quality of life. Currently, therapeutic options for patients with OA consist of medications to alleviate pain and treat the symptoms; however, due to typically poor outcomes, patients with advanced OA are unlikely to avoid joint replacement. In recent years, several studies have linked disrupted homeostasis of the joint cavity microenvironment to the development of OA. Recently, extracellular vesicles (EVs) have received increasing attention in the field of OA. EVs are natural nano-microcarrier materials with unique biological activity that are produced by cells through paracrine action. They are composed of lipid bilayers that contain physiologically active molecules, such as nucleic acids and proteins. Moreover, EVs may participate in local and distal intercellular and intracellular communication. EVs have also recently been shown to influence OA development by regulating biochemical factors in the OA microenvironmental. In this article, we first describe the microenvironment of OA. Then, we provide an overview of EVs, summarize the main types used for the treatment of OA, and describe their mechanisms. Next, we review clinical studies using EVs for OA treatment. Finally, the specific mechanism underlying the application of miRNA-enriched EVs in OA therapy is described.
Similar content being viewed by others
Introduction
OA is a degenerative joint disease that occurs in the knee joint. OA involves structural changes in hyaline cartilage, subchondral bone, ligaments, the joint capsule, the synovial membrane, and periarticular muscles. Local damage to the articular cartilage centered on weight-bearing areas is a representative pathological feature of OA. Worldwide, 9.6% of men and 18.0% of women over 60 years of age experience OA symptoms. Approximately 80 percent of people with OA have substantially reduced mobility, and 25 percent are unable to perform major daily tasks [1]. Currently, a gold standard therapy is unavailable for OA, although surgery and medications have achieved some success in treating patients. However, the safety and risks associated with surgery, as well as the side effects of drugs, remain clinical issues. An understanding of the pathogenic processes that drive OA might provide opportunities for future development of therapies to address this unmet clinical need.
OA is a low-grade inflammatory disease that causes cartilage degradation, synovial inflammation, subchondral bone alterations, osteophyte development, ligament degeneration, joint capsule hypertrophy, and proangiogenic characteristics [2, 3]. Synovitis refers to inflammatory alterations to the synovium, such as hyperplasia of the synovial lining, inflammatory cell infiltration, neoangiogenesis, and fibrosis [4,5,6]. Synovitis affects 70% of OA patients, and the severity of this condition is related to pain and cartilage loss [7, 8]. Synovial tissue from early OA patients shows elevated production of proinflammatory mediators, suggesting that acute synovitis is one of the first joint alterations to occur [9].
Many processes and substances, including transcription factors, epigenetic changes, cytokines, and proteases, govern joint tissue homeostasis, which is disturbed with OA [6]. This disturbance produces widespread alterations and prevents the synovial joint from facilitating frictionless and smooth mobility. Inflammation and thickening of synovial tissue are caused by this disruption [10], but proinflammatory mediators produced by OA immune cells from the synovium and infrapatellar fat pad (IPFP) also contribute to cartilage destruction [11]. Pattern recognition receptors such as Toll-like receptors recognize distinct pathogen-associated molecular patterns and damage-associated molecular patterns [including extracellular matrix (ECM) degeneration and products of cellular stress], prompting the cells present in the OA joint to release large amounts of inflammatory mediators. Activation of pattern recognition receptors induces cell signaling, resulting in the production of proinflammatory cytokines and chemokines such as interleukin (IL)-6, IL-8, IL-1, and tumor necrosis factor-α (TNF-α), as well as proteases such as matrix metalloproteinases (MMP)-1, MMP-3, and MMP-13, that degrade the structural components of cartilage ECM (primarily aggrecan (ACAN) and collagen) and alter chondrocyte viability and glycosaminoglycan(GAG)release [6, 11,12,13,14,15,16,17]. In patients with OA, large numbers of proinflammatory macrophages (M1) are activated, and these activated M1 macrophages cause further damage to the articular cartilage. Promoting the polarization of M1 macrophages to anti-inflammatory macrophages (M2) is a beneficial approach to protect articular cartilage and promote cartilage regeneration and repair. An imbalance in remodeling mediated by bone resorption by osteoclasts and bone creation by osteoblasts results in a decrease in tissue mineralization, a loss of stiffness, and thickening of the subchondral bone [18].
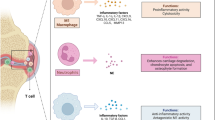
The abovementioned factors involved in the pathogenesis of OA cause an imbalance in the microenvironmental homeostasis of the joint cavity, which further aggravates the disease. This microenvironment has been summarized as presenting dysregulation of anti-inflammatory and proinflammatory factors, an imbalance in the immune system, disruption of the chondrogenic factor, and an increase in the amount of destructive factors (Fig. 1). Therefore, accurate regulation of the OA microenvironment and restoration of homeostasis are essential to protect articular cartilage and slow the development of OA. EVs and their regulatory potential have been the subject of many recent studies. EVs participate in cell-to-cell communication. They are produced by cells and are considered natural nano-microcarrier materials that, unlike other biological materials, inherit the biological characteristics of their precursors. EVs also have low toxicity and excellent selectivity, as well as the ability to penetrate biological membranes and transport a large number of bioactive molecules between cells. EVs were linked to OA in recent studies, as EVs regulate the inflammatory response and promote M2 macrophage polarization, cartilage production, and tissue healing [19,20,21,22]. In addition to attracting interest for other therapeutic applications, EVs may play a role in the treatment of OA by controlling the OA microenvironment. Moreover, the creation of nanomaterials based on EVs or their derivatives will be a new avenue for future cell-free OA biotherapy.

Comparison of the microenvironment between normal and OA joint cavities. Synovial thickening, synovial inflammation, massive activation of inflammatory M1 macrophages, articular cartilage erosion, and the release of a large number of inflammatory factors (including TNF-α, IL-1, IL-12, IL-6, and IL-16), increase in the levels factors responsible for cartilage destruction, and destruction of factors responsible for cartilage formation occur. B cells、T cells、mast cells and NK cells infiltrate into the joint cavity. The imbalance in the homeostasis of the OA joint cavity microenvironment promotes the formation and progression of OA
In this review, we first present the microenvironment of OA. Then, we will provide an overview of EVs and the main types of EVs used to treat OA. Finally, we review the main mechanisms of EVs in the treatment of OA.
The microenvironment of OA
OA is a disease that can affect the cartilage and surrounding tissues of any joint, although it most typically affects major joints, such as the knee, hip, and wrist. According to epidemiological data, OA affects approximately 4% of the global population, with basically no sex or geographic disparities. The annual incidence of OA-related disability accounts for 2% of all disability rates. Men are more likely than women to experience OA before the age of 45 years, while women are more likely to experience OA after the age of 45 years. The incidence increases progressively with age, reaching up to 40% or more in some areas in adults over the age of 60 years, 20% of whom will experience substantial symptoms at a certain stage. With the aging of the population, the incidence of OA will increase, thus increasing the costs of medical treatment. Therefore, to the identification of effective strategies to prevent and cure OA has become increasingly essential. The next section provides an overview of the microenvironmental changes that occur in the joints throughout the pathogenesis of OA.
Cytokines
The OA microenvironment is incredibly complex, containing many different cells types that release a variety of cytokines. Macrophages, which are categorized into either the M1 or M2 type, are the most important of these cell types. A considerable imbalance in the ratio of M1 to M2 macrophages has been observed in the OA microenvironment, with an increase in the M1 population and a reduction in the M2 population, which contributes to the development of OA.
Cytokines stimulate cells, which in turn activate the local cells (synovial cells, chondrocytes, osteoblasts, osteoclasts, etc.), causing pathological changes such as cartilage degradation, vascular proliferation, and bone production through endocrine, autocrine, and paracrine signaling [23]. IL-1β and TNF-α are the most important of these cytokines. The most potent cytokine for cartilage degradation in the OA microenvironment is IL-1β [24], which is produced by chondrocytes, osteoblasts, synoviocytes, and leukocytes. IL-1β acts independently or synergistically with other cytokines to cause articular cartilage degradation and joint inflammatory responses [25]. According to Piotr Wojdasiewicz et al. [26], IL-1β activates its downstream transcription factors to promote synovial vasodilation, hyperplasia, and joint discomfort, resulting in the expression of hundreds of genes, some of which produce inflammatory mediators, including nitric oxide (NO) and prostaglandin E2 (PGE2). By inducing the generation of interstitial collagenase, matrix lysin 1 and collagenase 3 [27], and platelet-reactive protein motif-containing disintegrin-like metalloproteinases, IL-1β disturbs chondrocytes and the ECM, damaging the cartilage structure and even inducing localized abnormalities in the articular cartilage. These changes further amplify the activity of IL-1β by inducing the production of TNF-α, other cytokines, such as IL-6 and IL-8, chemokine (C–C motif) ligand 5 (CCL5), and different adhesion molecules by chondrocytes, osteoblasts, synoviocytes, and leukocytes in an autocrine manner. IL-1β also induces the generation of reactive oxygen species (ROS) during disease progression [28], which produces different peroxides that directly degrade articular cartilage and exacerbate OA symptoms.
TNF-α is the another inflammatory cytokine that triggers an inflammatory response in people with OA [29]. TNF-α stimulates the release of MMP-1, MMP-3, and MMP-13 from cartilage, the synovium, and subchondral bone layer-associated cells, resulting in a gradual decrease in the cartilage collagen and proteoglycans contents and inhibition of proteoglycan and collagen II (COL II) synthesis. These changes ultimately indirectly leads to chondrocyte death and disturbs the homeostatic balance between cartilage damage and repair, resulting in varying degrees of chondrocyte death and disruption [30]. TNF-α can induces the proliferation of vascular endothelial cells by stimulating tissue expression of growth factors such as vascular endothelial growth factor, basic fibroblast growth factor, and platelet-derived growth factor, which promotes vasodilation and subchondral bone and synovial tissue proliferation and exacerbates the inflammatory symptoms of OA, as noted by Wang et al. [31]. TNF-α may also reduce the activity of the protein kinase CK2 [32]. Furthermore, TNF-α may cause chondrocyte death through both apoptosis and autophagy, worsening cartilage degradation in individuals with OA.
Cytokines have bidirectional functions in tissues such as articular cartilage, the synovium, and subchondral bone. The overexpression of cytokines may lead to the overexpression of downstream molecules such as MMPs, which can contribute to pathological alterations, including cartilage deterioration. As a result, one of the features of the OA microenvironment is the disturbance of the homeostatic equilibrium maintained by cytokines.
Proteinases
Several proteases that have been implicated in early OA articular cartilage degradation are considered diagnostic and therapeutic criteria for OA. Cellular tissues and immune cells release inflammatory mediators such as IL-1, IL-7, and TNF-α after external activation of the joint [33]. These inflammatory mediators increase the production of several proteases, such as MMPs, a disintegrin and metalloprotease (ADAM), and ADAM with thrombospondin motifs (ADAMTS), which can cause structural damage to articular cartilage and the adjacent joint tissues [34]. The major components of cartilage ECM are ACAN and COL II, which have the primary purposes of maintaining the mechanical structure and biochemical characteristics of articular cartilage.
MMPs are zinc-dependent endonucleases that regulate the composition of the cellular matrix during the normal physiological processes of the organism. MMPs are the most prominent proteases involved in the destruction of cartilage ECM in the OA microenvironment [35]. MMP-13, which is generated by chondrocytes and fibroblasts, is the most effective collagen-degrading enzyme in the MMP family [33]. Furthermore, MMP-13 is a highly active protease involved in the loss of cartilage ECM in the OA microenvironment due to its dual effects on degrading both proteoglycans and the ECM [36, 37]. As a result, the OA microenvironment overexpresses the MMP family of enzymes.
Similar to MMPs, the ADAMTS family of proteases are zinc metalloproteases with platelet-responsive protein motifs. ADAMTS gene expression contributes to the organism homeostasis under normal conditions. However, in the OA microenvironment, inflammatory cytokines such as IL-1 and TNF-α cause a significant increase in the levels of ADAMTS family members, which leads to the development of nonclassical OA inflammation via a downstream chain reaction [38]. Early cartilage deterioration in OA is assumed to involve the hydrolysis of cartilage ACAN by ADAMTS. According to the literature, ADAMTS-5, a member of the ADAMTS family of proteases, is the main hydrolase responsible for the degradation of ACAN in the ECM of OA articular cartilage [39]. Because ADAMTS is involved in total tissue regeneration, increased ADAMTS expression may be linked to not only the degeneration and deterioration of the joint but also the persistence of OA inflammation and injury, implying that OA is an uncontrolled healing process [40].
Immune cell infiltration
According to Liu et al. [41], the incidence and progression of OA may be associated with the increased infiltration of memory B cells, mast cells, and macrophages and the reduced infiltration of memory CD4 T cells and activated NK cells. By releasing inflammatory mediators and antibodies, B cells control ECM degradation [42, 43]. Mast cell-derived trypsin, according to Wang et al., causes inflammation, chondrocyte death, and cartilage degradation [44]. Proteoglycan degradation has been observed in cocultures of activated mast cells and chondrocytes, according to Woolley et al. [45]. These findings suggest that mast cells causes cartilage deterioration. De Lange-Brokaar, et al. [46] found a significantly greater numbers of mast cells higher in OA samples than in RA’s,and these cells were associated with structural damage in patients with OA, suggesting a role for mast cells in this disease. Macrophages may control the severity of OA and joint inflammation by secreting a variety of mediators.Apparently, regulating the functional phenotype of macrophages may effectively cure OA or promote cartilage repair and regeneration [47, 48]. By secreting cytokines and growth factors, T cells cause the ECM degradation and remodeling [49]. As shown in the study by Ezawa et al.,anincrease in the number of memory CD4 T cells is a common phenomenon observed in the local inflammatory response of OA joints and plays a role in the development of OA [50]. NK cells can modulate the immune system. Based on accumulating evidence,NK cells are key to promoting immune cells involved in OA, and their interaction is facilitated by the CXCL10/CXCR3 axis. NK cells have the ability to alter subchondral bone metabolism and repair in addition to causing cartilage loss [51]. It was discovered that IL-2-activated NK cells may lyse both allogeneic and autologous mesenchymal stem cells [52]. Additionally, NK cells that have been activated can promote osteoclast development [53]. According to the studies reviewed above, OA is caused by the actions of B cells, mast cells, macrophages, T cells, and NK cells. These findings imply that the infiltration of different immune cells, which is a component of the OA microenvironment, is crucial for the pathogenesis of OA.
Cartilage injury and degeneration
Articular chondrocytes are dormant cells that do not multiply after maturation unless a traumatic or pathogenic event occurs [54]. Cellular degeneration is a natural aging event in which cells stop growing, regress, and lose their ability to proliferate. The lack of neovascularity in articular cartilage inhibits its capacity to regenerate, increasing the difficultly of repair after injury. The increased levels of damaging cytokines and proteases in OA causes articular cartilage erosion, extensive cartilage ECM degradation, and joint surface unevenness. Additionally, the cytokines and proteases mentioned above operate on chondrocytes, altering their biological function and hastening their degeneration and senescence. Degenerated and senescent cells produce important substances in the tissue milieu, which may alter the tissue microenvironment and damage nearby tissues, such as articular cartilage.
Overview of EVs
EVs biogenesis
Pericellular vesicles were initially discovered in mammalian tissues and bodily fluids in the late 1960s [55, 56]. The term "extracellular vesicles" was first used in 2011 to characterize all extracellular structures surrounded by lipid bilayers. The three types of EVs that are classified according to their size, are ectosomes, exosomes (Exos), and apoptotic cell-derived EVs (ApoEVs). The sizes, contents, and formation mechanisms of the three types of extracellular vesicles are all different (Fig. 2). Ectosomes (100–1000 nm in diameter), comprising microvesicles (MVs), microparticles, and large and small vesicles, are very small vesicles that are expelled externally through the plasma membrane. Exos are endosomal vesicles of 50–150 nm in diameter that are produced by repetitive plasma membrane invaginations. Multivesicular bodies (MVBs) containing intraluminal vesicles (ILVs) are formed after the early creation of cup-like structures, early sorted endosomes (ESEs), and late sorted endosomes (LSEs). Both Exos and MVs have the ability to mediate intercellular communication and immunological control.

The sizes, contents, and formation mechanisms of the three types of EVs. a The size ranges of three types of EVs. b EVs biogenesis. c The contents of EVs from healthy cells
Ectosomes and Exos are two types of EVs that are generated by healthy cells, although apoptotic cells can also release a variety of EVs. ApoEVs are divided into two types: large membrane-bound vesicles (apoptotic vesicles, 1000–5000 nm) and very small apoptotic MVs (less than 1000 nm). ApoEVs have been proven in several investigations to exhibit activities comparable to those of healthy cell-derived EVs. The primary tasks of ApoEVs include removing apoptotic cells and providing a method of intercellular communication. The therapeutic functions of EVs produced from live cells in the treatment of OA are the topic of this review.
EVs enrichment
Ultracentrifugation, gradient ultracentrifugation, coprecipitation, size-exclusion chromatography, field flow fractionation, and affinity capture are the methods being used for EVs enrichment and purification. EVs have also been isolated and extracted using techniques such as microporous filtering, microfluidics, and high-performance liquid chromatography.
The prominent approach for used for EVs separation is ultracentrifugation [57]. This method feasibly separates particles with varying settling rates and then eliminates the undesired components during each centrifugation cycle by increasing the centrifugation speed and/or time in a stepwise manner. Although ultracentrifugation is the most widely used technique for EVs isolation, it has several drawbacks, including bulkiness, a requirement for expensive instrumentation, being time-consuming, carrying the risk of contamination with particles of aggregated proteins and ribonucleoproteins, and a requirement for large amounts of sample. Gradient ultracentrifugation, size-exclusion chromatography, and field flow fractionation all face difficulties when separating particles based on particle density or size. In contrast to these physical-based isolation approaches, coprecipitation is a polymer coprecipitation-based EVs enrichment technology. Typically, this approach decreases the hydration of EVs, resulting in their precipitation. A low centrifugal force enables simple and repeatable separation of precipitated EVs products, eliminating the need for a time-consuming ultracentrifugation process [58, 59]. However, this technique is costly and lacks specificity for EVs. Multiphase polymer particles, as well as coprecipitated lipoproteins and argonaut-2 (Ago-2) RNA complexes, are common byproducts of this method. Using phase interactions between markers on the EVs surface and captured molecules linked to distinct carriers, affinity capture enables the extraction of EVs with greater purity but lower yield [60].
Each approach has benefits and drawbacks, and a combination of methods may be the best option to extract the separated EVs. The features of several EVs separation techniques are summarized in Table 1.
EVs characterization
Isolated EVs must be properly characterized according to the International Society for Extracellular Vesicles (ISEV) minimal standard report for EVs characterization. Complete EVs characterization encompasses an assessment of both general and specific vesicle characteristics. The surface protein indicators of EVs are often characterized using Western blotting or enzyme-linked immunosorbent assay (ELISA). At least three positive and one negative EVs protein indicators should be described according to the ISEV. Moreover, at least one transmembrane/lipid binding protein (e.g., CD63 and CD9) and one cytoplasmic protein (e.g., TSG101 and ALIX) should be positive protein markers. Imaging methods and biophysical characterization are needed to characterize single vesicles. However, the only imaging techniques capable of acquiring high-resolution EVs images are electron microscopy (EM) and atomic force microscopy (AFM), which are methods that include transmission electron microscopy, scanning electron microscopy, and cryo-electron microscopy. Immunogold electron microscopy is commonly utilized to visualize the staining of certain EVs markers. Nanoparticle tracking analysis (NTA), tunable resistive pulse sensing (TRPS), dynamic light scattering (DLS), and flow cytometry (FC) are all examples of biophysical characterization techniques [67].
A light microscopic single EVs analysis approach (SEA) that enables a reliable assessment of numerous protein biomarkers in a single vesicle was described in a recent study [68]. With this method, EVs are immobilized in a microfluidic chamber, immunostained, and photographed. When vesicles are immobilized on the chip surface, the signal-to-noise ratio is generally greater than that when the vesicles are free-floating or flowing in solution. However, the chemical composition of each EVs and how EVs subpopulations are characterized are still unknown [69].
Main EVs types used to treat OA
Natural EVs used in OA therapy
Natural EVs are generally formed from unmodified mesenchymal stem cells (MSCs), which not only have the same biological functions as MSCs but also possess features that MSCs lack. As a result, EVs (including MVs and Exos) have been recommended as a replacement for standard cell-based OA treatments due to the following benefits: (1) a small size, (2) improved safety and fewer side effects due to their natural lipid and surface protein composition [70], (3) lower immunogenicity [70], and (4) protection from degradation and therapeutic site targeting of the therapeutic substances (nucleic acids and proteins) [71, 72], (5) the capacity to overcome various biological hurdles that MSCs are unable to overcome [72], (6) elimination of the need for cell injection, and (7) easy preservation procedures with fewer ethical concerns. EVs for OA research have been extracted from MSCs generated from a variety of sources, including bone marrow, adipose tissue, umbilical cords, synovial membranes/fluid, embryonic stem cells, and induced pluripotent stem cells [70]. Table 2 summarizes and compares the characteristics, size, safety, efficacy, derivation, dose, and animal models used with synovial mesenchymal stem cell-derived EVs (S-MSC-EVs), adipose-derived mesenchymal stem cell-derived EVs (AD-MSC-EVs), bone marrow mesenchymal stem cell-derived EVs (BM-MSC-EVs), and human umbilical cord mesenchymal stem cell-derived EVs (hUMSC-EVs). To make the article more rational, we also summarize the differences in MSCs isolated from different tissue sources in Table 3.
S-MSC-EVs
S-MSCs were extracted from the synovium surrounding a joint for the first time in 2001 [88]. S-MSCs specifically regenerate cartilage [89] and are presumed to be the most promising cells for stimulating cartilage regeneration. In vitro, S-MSCs exhibit excellent chondrogenic differentiation potential [90,91,92].An intra-articular injection of S-MSCs substantially improves cartilage regeneration in experimental animal models. These cells have also been utilized to treat joint-related disorders such as OA [93,94,95,96]. According to several studies have recently, S-MSCs-EVs can successfully stimulate cartilage regeneration and delay the development of OA [73, 74, 97]. Tao et al. [73] discovered that human S-MSCs-Exosomes (S-MSCs-Exos) expressing wingless/integrated (Wnt) 5a and Wnt5b reduced ECM secretion by activating Yes-associated protein (YAP) via alternate Wnt signaling pathways while increasing chondrocyte proliferation and migration. MiR-140-5p-Exos inhibited this adverse effect by targeting RalA. In vitro, human S-MSC-140-Exos increased articular cartilage proliferation and migration without interfering with ECM secretion. However, in vivo, human S-MSC-140-Exos effectively prevented OA in a rat model. Additionally, S-MSC-Exos increased chondrocyte proliferation and migration while inhibiting apoptosis, but had no effect on ECM production or secretion, according to Wang et al. [75]. Qiu et al. discovered that miR-129-5p expression was downregulated in OA patients and IL-1-induced chondrocytes, but high mobility group protein (HMGB) 1 was substantially upregulated [76]. S-MSC-Exos enriched in miR-129-5p decreased chondrocytes apoptosis, whereas S-MSCs-Exos enhanced both the IL-1-mediated inflammatory response and apoptosis in chondrocytes. Upon further investigation of this process, miR-129-5p was shown to bind the 3' untranslated region (3'UTR) end of HMGB1 and suppresses IL-1-mediated HMGB1 overexpression. Overall, this study revealed that miR-129-5p present in S-MSCs-Exos may prevent IL-1-induced OA by blocking HMGB1 release.
AD-MSC-EVs
AD-MSCs have been shown to have significant abilities to control cartilage regeneration and inflammation. They are regarded as a good source of cells for the treatment of OA [98,99,100,101,102,103]. However, the mechanism by which AD-MSCs stimulate cartilage repair is unknown. AD-MSCs control the local microenvironment mainly by secreting paracrine trophic factors, to promote repair and regeneration, reduce cartilage degradation, and enhance joint function [100]. According to Tofino-Vian et al., EVs, including MVs and Exos, mostly mediate the paracrine effects of AD-MSCs on osteoblasts in individuals with OA [77]. Wu et al. [78] examined the function of IPFP MSCs-derived Exos (MSCs-IPFP-Exos)in OA and the underlying processes. MSCs-IPFP generate large number of Exos and that these MSCs-IPFP-Exos exhibit an Exos-like morphology. MSCs-IPFP-Exos have been shown to ameliorate OA in vivo by inhibiting apoptosis. Moreover, MSCs-IPFP-Exos increase matrix secretion and decrease the expression of degradation-related factors. Furthermore, by blocking the mammalian target of the rapamycin (mTOR) pathway, MSCs-IPFP-Exos may substantially increase chondrocyte autophagy. Tofio-Vian et al. [79] isolated and identified MVs from human AD-MSCs (hAD-MSCs). Then, they studied the chondroprotective role of these MVs and discovered that they reduced the production of the inflammatory mediators TNF-α, IL-6, PGE2, and NO in IL-1-stimulated OA chondrocytes. When OA chondrocytes were treated with these MVs, the measured MMP activity and MMP-13 expression were reduced, but the expression of the anti-inflammatory cytokines IL-10 and COL II increased considerably. Woo et al. [80] examined the therapeutic potential of hAD-MSCs-derived small EVs (hAD-MSCs-sEVs) in the treatment of OA and the corresponding mechanism. hAD-MSCs-sEVs not only increased human chondrocyte proliferation and migration but also reduced the expression of MMP-1, MMP-3, MMP-13, and ADAMTS-5 by increasing COL II production in the presence of IL-1. An intra-articular injection of hAD-MSCs-sEVs dramatically slowed the development of OA and prevented cartilage degeneration in rats treated with sodium monoiodoacetate and mice with medial surgical damage.
BM-MSCs-EVs
EVs derived from BM-MSCs have been shown to affect cell fate, including apoptosis, proliferation, invasion, and migration [104, 105]. Furthermore, BM-MSCs-EVs control many physiological and pathological processes, such as the immune response, osteogenesis, fibrosis, and angiogenesis [106,107,108,109]. In several studies, BM-MSCs-EVs were shown to stimulate the regeneration and repair of injured tissues, including cartilage and subchondral bone [107, 110,111,112,113,114,115]. Wang et al. [81] discovered that miR-135b-Exos suppress the expression of transcription factor SP1 in chondrocytes. MiR-135b-Exos promote chondrocyte proliferation and accelerate OA cartilage repair by negatively regulating Sp1 expression. This study may provide a new direction for OA treatment. Li et al. [116] investigated the effect of BM-MSCs-Exos on the etiology and behavioral symptoms of mice with lumbar facet joint OA (LFJ OA). They used BM-MSCs-Exos to treat mice with LFJ OA and detected changes in aberrant nerve invasion in the cartilage and subchondral bone. They discovered that BM-MSCs-Exos may alleviate pain by removing abnormal calcitonin gene-related peptide (CGRP)-positive nerves and abnormal H-vascular development in LFJ subchondral bone. BM-MSCs-Exos also suppress the expression of anti-tartaric acid phosphatase and activation of the receptor activator of nuclear factor-κB ligand (RANKL)-receptor activator of nuclear factor-κB (RANK)-tumor necrosis factor receptor-associated factor 6 (TRAF6) signaling pathway. In addition, subchondral bone remodeling was increased. Qi et al. [82] demonstrated that BM-MSCs-Exos can promote chondrocyte proliferation and significantly inhibit IL-1-induced chondrocyte apoptosis by inhibiting p38 and ERK1/2 phosphorylation and stimulating the Akt signaling pathway, indicating that BM-MSCs-Exos can effectively maintain chondrocyte viability in an inflammatory environment. Cosenza et al. [83] discovered that BM-MSCs-Exos and BM-MSC-derived microparticles can increase the expression of chondrocyte markers (COL II and ACAN) while suppressing catabolism (MMP-13, ADAMTS-5) and inflammation (iNOS) markers. BM-MSCs-Exos and BM-MSCs-derived microparticles also protect chondrocytes from apoptosis and suppress inflammatory macrophage activation. Rong et al. [84] extracted sEVs after the hypoxic stimulation of BM-MSCs. These sEVs were capable of initiating the fast repair and regeneration of osteochondral defects and reducing the development of OA. sEVs can deliver miR-216a-5p to cartilage cells. MiR-216a-5p-sEVs can downregulate Janus kinase 2 (JAK2), promote chondrocyte proliferation and migration, and inhibit apoptosis. In vitro and in vivo, kartogenin (KGN)-BM-MSCs-sEVs treatment resulted in more effective cartilage repair and matrix production than treatment with KGN [117]. In conclusion, BM-MSCs-EVs are a viable therapeutic approach for OA.
hUMSC-EVs
hUMSCs have the advantages of a large tissue supply, a high growth capacity, a painless collection technique and excellent biological characteristics. According to previous research, hUMSCs may develop into osteoblasts, chondrocytes, adipocytes, and a variety of other cell types [118,119,120]. In recent investigations, MSCs-EVs have been shown to stimulate cartilage formation [73, 121, 122]. Hu et al. [85] examined the role and mechanism of hUMSCs-sEVs in cartilage regeneration. They found that hUMSCs-sEVs could enhance chondrocyte and human bone marrow mesenchymal stem cells (hBM-MSCs) migration, proliferation, and differentiation. An miRNA microarray revealed that miR-23a-3p was the most abundant miRNAs expressed in hUMSCs-sEVs. After transferring miR-23a-3p, hUMSCs-sEVs could suppress phosphatase and tensin homolog (PTEN) expression while increasing protein kinase B (Akt) expression, therefore enhancing cartilage regeneration. Yan et al. [86] revealed that hUMSCs-Exos may function as a natural carrier of the long noncoding RNA (lncRNA) H19. The lncRNA H19 can increase chondrocyte proliferation, migration, and matrix secretion and inhibit chondrocyte death and senescence. The corresponding mechanism is that lncRNA H19-Exos compete with miR-29b-3p and upregulate forkhead box O3 (FOXO3) expression in chondrocytes. An intra-articular injection of hUMSCs-Exos substantially enhances the healing of cartilage abnormalities. Furthermore, hUMSCs-Exos derived from three-dimensional (3D) culture were more beneficial for cartilage regeneration than those derived from traditional two-dimensional (2D) culture [87].
EVs derived from other cells
Sang et al. discovered [123] that hydrogels containing chondrocyte-derived Exos can promote cartilage regeneration and repair by controlling the levels of inflammatory factors in the OA microenvironment and polarizing macrophages. Zheng,et al. [124] found that compared with IL-1β stimulated chondrocytes, Exos from normal chondrocytes can prevent the development of OA by reversing mitochondrial dysfunction and polarizing macrophages to the M2 phenotype. According to Wa et al. [125], M2 macrophage-derived Exos exert a therapeutic effect on rats with knee OA (KOA) by suppressing the PI3K/Akt/mTOR pathway and reducing the inflammatory response and pathological damage to the articular cartilage. Tan et al. [126] found that the lncRNA H19 present in fibroblast-like cell-derived Exos can target the mir-106b-5p /TIMP2 axis, increase OA chondrocyte proliferation and migration, block ECM degradation, and attenuate the development of OA.
Engineered EVs for OA therapy
Engineered EVs have been the focus of scientists in recent years to increase the EVs target specificity and achieve precise control. Engineered EVs outperform natural EVs in terms of therapeutic potential. Engineering procedures (Fig. 3) (e.g., transfection, coincubation, electroporation, sonication, freeze–thaw cycles, extrusion, the use of saponins) are utilized to load EVs with suitable cargo to obtain superior therapeutic effects. Many preclinical experiments analyzing EVs-based medications or molecular delivery have yielded promising results. EVs-encapsulated therapeutic molecules and medications are more stable in the circulation, traverse physiological barriers more easily and have higher biological activity and lower systemic toxicity than their corresponding free molecules. Researchers have devised novel methods to create high-purity and high-yielding EVs and to construct drug or molecule delivery systems with a high loading efficiency, targeting ability, and regulated drug or molecule release to increase the usability of EVs in OA therapy. Liang et al. combined the lysosome-associated membrane protein 2 (LAMP-2B) gene with chondrocyte affinity peptide(CAP) for transfection into dendritic cells to create chondrocyte-targeting EVs and to soften and promote cartilage regeneration. MiR-140 was then transported to chondrocytes deep in the joint [127]. Xu et al. fused the MSC-binding peptide E7 to the EVs membrane protein LAMP-2B to create EVs with E7 peptide (E7-EVs) and synovial fluid mesenchymal stem cell (SF-MSC) targeting capabilities. KGN supplied with E7-EVs enters SF-MSCs more effectively and induces a greater degree of chondrogenic differentiation than KGN provided with EVs alone without E7, indicating that these EVs might be a promising advanced OA stem cell treatment [128].

The most important EVs engineering methods. Strategies for designing donor cells are shown on the left. Co-incubation and gene transfection methods are utilized to deliver cargos into donor cells. EVs engineering strategies are shown on the right. Sonication 、electroporation、freeze and thaw、co-incubation and extrusion techniques are utilized to insert cargos into EVs
Main mechanisms of EVs in the treatment of OA
Immunomodulation
The principal function of the immune system has long been presumed to be a part of the body's defense mechanism against viruses and the external environment. However, the immune system clearly exerts a significant effect on tissue healing. Proinflammatory cytokines, anti-inflammatory cytokines, and bidirectional factors all exist in the joint cavity, and the dynamic balance between these three variables maintains the normal physiological metabolism of articular cartilage. When this dynamic equilibrium is perturbed, the joint microenvironment is disrupted, which leads to the development of OA. Proinflammatory cytokines such as IL-1, IL-6, and IL-8, as well as MMP-3, are implicated in cartilage injury-induced matrix degradation and joint degeneration [129]. Proinflammatory cytokines, including IL-6 and IL-1 as well as the nuclear factor kappa B (NF-kB) pathway, have been found to exert a significant effect on synovial inflammation and cartilage degradation in OA patients. According to Xia et al. [130],and Zhao et al. [131], EVs produced from AD-MSCs were able to downregulate IL-6 expression and alter the expression of components of the NF-kB pathway.
Synovial and immunological cells, such as macrophages, produce proinflammatory cytokines and MMPs, which contribute to the development of OA [132]. According to recent research, M1 macrophages in OA synovial tissue limit MSC chondrogenic development in vitro via IL-6 [133], and M2 macrophages improve transplant cartilage survival by generating the anti-inflammatory cytokine IL-10 to decrease unfavorable inflammatory responses [134]. As a result, in cartilage regeneration therapy, the proinflammatory milieu of cartilage degeneration or OA must be controlled. Previously, macrophages were divided into two phenotypes: proinflammatory M1 and anti-inflammatory M2 phenotypes. Interferon-γ (IFN-γ), TNF-α, or pathogen-associated molecular patterns activate M1 macrophages. These activated macrophages then release proinflammatory cytokines such as IL-1, IL-6, IL-12, and iNOS. On the other hand, M2 macrophages are activated by different pathways. Transforming growth factor (TGF)-β1 and arginase-1 (Arg-1) are two growth- and angiogenesis-related substances secreted by M2 macrophages that decrease inflammation and promote tissue remodeling [135, 136]. The spatial and temporal distribution of M1 and M2 macrophages is critical to coordinate inflammation and tissue regeneration [137, 138].
MSCs-EVs produce large quantities of the anti-inflammatory cytokines IL-10 and TGF-β1 while suppressing the production of the proinflammatory mediators IL-1, IL-6, TNF-α, and IL-12 (Fig. 4). Furthermore, MSC-EVs decrease macrophage activation and promote the M1 to M2 conversion, which is important in many inflammatory illnesses. MSC-EVs exert immunomodulatory effects, according to Zhang et al. They can increase M2 macrophage infiltration into OA cartilage defects and synovial membranes, reduce M1 macrophage infiltration, and downregulate the inflammatory factors IL-1β and TNF-α, resulting in an overall decrease OA inflammatory responses [121]. Although the immunomodulatory effects of EVs on OA are unknown, the presence of EVs in serum was recently shown to protect human OA cartilage from GAG loss in the presence of the inflammatory factor IL-1β. Additionally, the number of M2 macrophages increases following EVs therapy, increasing cartilage regeneration in immunoreactive rats, which has led to our hypothesis that MSCs-EVs might cure OA.

As a new type of natural nanomaterial, EVs secreted by stem cells from various tissues (bone marrow, umbilical cord, synovium, and adipose tissue) regulate the OA microenvironment through various mechanisms to restore the homeostasis of the joint cavity. EVs can be injected directly into the joint cavity in batches or combined with acellular scaffold materials to inhibit inflammatory factor release, and promote the polarization of M1 macrophages to M2 macrophages. Moreover, they also reduce the production of cartilage destruction factors, and promote the synthesis of factors involved in cartilage formation
MSCs-EVs also exert anti-inflammatory immunomodulatory effects on T cells, NK cells and B cells. Studies have shown that MSCs-Exo exerts an anti-inflammatory effect on T cells and B cells by reducing the number of CD4 + T cells and CD8 + T cells, increasing the amount of Treg cells, and alleviating the inflammatory response of collagen-induced arthritis [139]. Moreover, MSCs-Exo can promote the transformation of effector T cells into Treg cells and increase the expression level of CTLA-4 in Treg cells. CTLA-4 is an important factor in the immunosuppressive effect of MSCs-Exo [140]. Human dendritic cells (DCs) can release large extracellular vesicles (lEVs), which can effectively induce the activation of CD4 + T cells in vitro. Among EVs secreted by immature DCs, lEVs promote the secretion of Th2 cytokines such as IL-4, IL-5 and IL-13 [141]. In addition, immature dendritic cell-derived exosomes (imDECs) can attenuate the inflammatory response and reduce the infiltration of CD4 + T cells [142]. Tumor-derived exosomes (TDEs) can inhibit the recruitment and migration of NK cells to the tumor environment, while suppressing the secretion of cytokines IFN-γ and TNF-α by NK cells, leading to immune escape and tumor progression [143]. Studies have revealed that MSCs-Exo can inhibit the proliferation of B cells and the differentiation into immunoglobulin-secreting plasma cells, and CCL2 in exosomes directly inhibits the secretion of immunoglobulin antibodies by plasma cells [144]. In conclusion, EVs can exert their anti-inflammatory immunomodulatory effects by regulating T cells, NK cells and B cells. However, there are currently few experimental studies on how EVs regulate T cells, NK cells, and B cells in OA, lacking sufficient basic theory, which is also the focus of the future research.
Chondrocyte regulation
Inflammation exacerbates the degeneration of damaged/diseased cartilage in OA, resulting in cell death, matrix degradation, and finally a loss of structure and function [129, 132]. Chondrocyte apoptosis is linked to cartilage deterioration and the progression of OA [32], and EVs may help prevent apoptosis in these cells [87, 121]. Cell migration and proliferation have also been reported to be facilitated by MSCs-EVs [121, 145].
Chondrocyte migration and proliferation are two critical cartilage health mechanisms that are both suppressed in OA. EVs derived from various sources have been shown to increase osteoarthritic chondrocyte proliferation, migration, and viability in a dose-dependent manner [19, 80, 82, 121, 146]. For example, as the EVs dose increases, proliferation occurs sooner; notably, a dose of 10 g of EVs was sufficient to induce chondrocyte migration. Moreover, some studies have examined the proteins involved in chondrocyte adhesion, migration, and proliferation that are regulated by EVs produced from MSCs [147, 148]. EVs alter the expression of genes such as fibroblast growth factor (FGF)-2, survivin, and Bcl2/Bax to control cell proliferation or reverse the inhibitory effects of TNF-α and IL-1β on cell migration and proliferation [87, 117, 121, 146]. By enhancing s-GAG synthesis and suppressing NO and MMP-13 production to maintain stromal homeostasis in a TMJ-OA model, hMSC-EVs attenuated the decrease in proliferation and migration. EVs derived from bone marrow stem cells, adipose stem cells, and synovial stem cells also enhance cartilage regeneration in chondrocytes by increasing GAG synthesis and COL II protein expression, and adipose stem cell-derived EVs exert the most significant effect [149]. Furthermore, these EV protein sources influence ECM stability and actin cytoskeletal dynamics, indirectly increasing chondrocyte proliferation and migration. Moreover, by releasing nucleic acids such as miRNAs, MSCs-EVs can control cell proliferation and migration. The uses of several miRNAs to treat OA are summarized in Table 4.
Induction of ECM synthesis
Changes in the composition and organization of the ECM are characteristic of OA. Because COL II and proteoglycans are two of the most important components of the ECM of articular cartilage that contribute to the creation of a healthy cartilage matrix, degradation of cartilage ECM proteins leads to cartilage degeneration [38]. MMP-13 and ADAMTS-4 and ADAMTS-5 are able to reduce the levels of COL II and proteoglycans in the OA joint cavity microenvironment. According to recent research, MSC-EVs can reverse ECM degradation by increasing the expression of matrix proteins and other cartilage formation-related genes while decreasing the levels of matrix-degrading enzymes. Tofio-Vian et al. [79] isolated and identified MVs from hAD-MSCs. When OA chondrocytes were treated with MVs, MMP activity and MMP-13 expression were reduced, but COL II expression increased considerably. Woo et al. [80] also discovered that hAD-MSC-sEVs can reduce the expression of MMP-1, MMP-3, MMP-13, and ADAMTS-5 and increase COL II production.
Many studies have recently examined the role of miRNAs in MSC-EVs to control the ECM. Overexpression of miR-92a-3p in BM-MSCs-EVs increased the expression of cartilage formation-related genes such aggrecan, SRY box gene-9 (SOX9), COL9A1, COL2A1, and cartilage oligomeric matrix protein (COMP) while decreasing the expression of COL10A1, Runt-related transcription factor 2 (Runx2), and MMP-13, according to Mao et al. [151]. Protection provided by EVs is beneficial not only because of the microRNAs contained within EVs but also because of EVs proteins. S-MSC-EVs, for example, contain miR-140-5p, which restores ECM secretion by regulating RalA expression, thereby rescuing SOX9 expression [73]. However, the underlying mechanism remains a mystery. As a result, additional research is needed to confirm the aforementioned findings and determine the mechanisms of cartilage matrix catabolism and anabolism.
Microenvironmental homeostasis
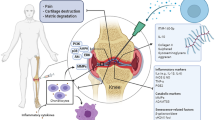
MSCs have been proven to have great promise in the treatment of OA in both preclinical and clinical trials. MSCs are involved in tissue homeostasis, free radical scavenging, immunomodulation, and cell proliferation [152, 153]. Substantial data show that the therapeutic effects of MSCs are primarily mediated by paracrine pathways and that MSCs-EVs exert a critical therapeutic effect [21, 73, 121, 154, 155]. Therefore, MSC-EVs maintain homeostasis within the joint microenvironment during OA treatment (Fig. 5).

MSCs-EVs target articular chondrocytes and synoviocytes to maintain homeostasis in the articular microenvironment. Cartilage degeneration, synovitis, and subchondral bone degradation are all pathological processes associated with OA. Synovial fluid component contents are altered by MSCs-EVs. Arrows pointing downward indicate downregulation, whereas arrows pointing upward indicate upregulation
MSCs-EVs have mostly been used in mice and rats for preclinical therapy of OA and osteochondral abnormalities. MSCs-EVs suppress synovial inflammation, protect subchondral bone, reduce oxidative stress and osteoblast senescence, prevent cartilage and bone from deterioration, correct gait anomalies, stimulate cartilage regeneration, and slow OA development [19, 78, 82, 83, 121]. Table 5 summarizes the in vitro and in vivo data on MSC-EVs.
Although MSCs-EVs are comparable to MSCs in terms of treating OA and osteochondral abnormalities, they are not the same. Notably, certain drawbacks to using EVs as a clinical translation tool in regenerative medicine have been documented. First, an isolation method that maintain the qualities of EVs in the long term is unavailable. Second, the large numbers of EVs needed for animal investigations and human clinical trials are difficult to attain [158]. Only a few mice can be treated with approximately 1–2 mg (protein content) of EVs generated from a total of approximately 60 million MSCs [145]. MSCs-EVs, on the other hand, are a simpler, safer, more practical, and easier-to-regulate OA therapeutic option than direct cell transplantation.
In conclusion, MSCs-EVs can affect intra-articular cells by controlling cartilage matrix anabolism and catabolism, subsequently enhancing the intra-articular inflammatory milieu, changing intra-articular homeostasis, and curing OA.
Clinical trial
Due to the advantages of MSC-EVs, many achievements have been reported, and clinical trials have been conducted in other disease fields, including Alzheimer's disease, lung infections, acute respiratory distress syndrome (ARDS), COVID-19, dry eye syndrome, etc. MSC-EVs will inevitably facilitate important advancements in the field of medicine in the future. However, research on the use of EVs as a biological alternative treatment is still in its early stages. The therapeutic use of EVs in the treatment of OA is limited by a variety of issues, as described below: 1. In terms of the illness itself, OA has a complicated etiology that may be caused by a number of different factors, necessitating additional research. 2. From the perspective of EVs, this industry still has certain bottleneck issues: ①for extensive pharmaceutical uses, EV isolation and purification techniques, yield, and purity have not been standardized; ②exosome composition heterogeneity and preservation are difficult problems for industrialization development and ③targeted cells internalization of EVs alters their chemical composition, making subsequent treatment results unpredictable. ④After EVs enter recipient cells, their subsequent biological distribution, pharmacokinetics and specificity of targeted delivery to the specific organ, as well as the therapeutic mechanism of OA disease, have not been fully elucidated. ⑤The location, duration of residency, and biological effects of EVs injected into the articular cavity on normal cells remain unclear.⑥Finally, experimental support for therapy in large animals is insufficient.
Conclusions and future perspectives
The whole joint, including the cartilage and subchondral bone, is affected by OA. The microenvironment of OA is complicated, and a complete understanding of this microenvironment will be extremely helpful to treat this disease. Because of their unique roles and properties, EVs may control the microenvironmental changes that coordinate the progression of OA, hence delaying disease progression. Furthermore, EVs contain a large number of proteins, miRNAs, and other bioactive molecules that are important for tissue repair and have a wide range of therapeutic applications in the etiology, diagnosis, and treatment of OA. MSCs-EVs inherit the basic activities of their parental cells, and their therapeutic benefits mediated by immunomodulation, tissue cell repair, and regenerative effects may become a key strategy for the treatment of OA. However, these OA studies are currently focused on animal models. The processes are difficult, time-consuming, and expensive, which are the main roadblocks to their usage. As a result, more clinical trials will be required in the future to validate these findings. Engineered EVs have recently become the focus of scientific research to improve the targeting specificity of EVs and enable more precise control. Engineered EVs outperform natural EVs in terms of their therapeutic potential. The study of EVs in OA, including their role, mode of action, and diagnostic/therapeutic applications, is still in its infancy, and many questions remain unresolved. We propose that natural nanomaterial-EVs will be employed as an effective therapeutic strategy for OA patients in the future as technology advances.
Availability of data and materials
Not applicable.
Abbreviations
- OA:
-
Osteoarthritis
- EVs:
-
Extracellular vesicles
- IPFP:
-
Infrapatellar fat pad
- ECM:
-
Extracellular matrix
- IL:
-
Interleukin
- TNF-α:
-
Tumor necrosis factor-α
- MMP:
-
Matrix metalloproteinases
- ACAN:
-
Aggrecan
- GAG:
-
Glycosaminoglycan
- M1:
-
Proinflammatory macrophages
- M2:
-
Anti-inflammatory macrophages
- NO:
-
Nitric oxide
- PGE2:
-
Prostaglandin E2
- CCL5:
-
Chemokine (C–C Motif) Ligand 5
- ROS:
-
Reactive oxygen species
- COL II:
-
Collagen II
- ADAM:
-
A disintegrin and metalloprotease
- ADAMTS:
-
ADAM with thrombospondin motifs
- Exos:
-
Exosomes
- ApoEVs:
-
Apoptotic cell-derived EVs
- MVs:
-
Microvesicles
- MVBs:
-
Multivesicular bodies
- ILVs:
-
Intraluminal vesicles
- ESEs:
-
Sorted endosomes
- LSEs:
-
Late sorted endosomes
- Ago-2:
-
Argonaut-2
- ISEV:
-
The International Society for Extracellular Vesicles
- ELISA:
-
Enzyme-linked immunosorbent assay
- EM:
-
Electron microscopy
- AFM:
-
Atomic force microscopy
- NTA:
-
Nanoparticle tracking analysis
- TRPS:
-
Tunable resistive pulse sensing
- DLS:
-
Dynamic light scattering
- FC:
-
Flow cytometry
- SEA:
-
Single EVs analysis approach
- MSCs:
-
Mesenchymal stem cells
- S-MSC-EVs:
-
Synovial mesenchymal stem cell-derived EVs
- AD-MSC-EVs:
-
Adipose-derived mesenchymal stem cell-derived EVs
- BM-MSC-EVs:
-
Bone marrow mesenchymal stem cell-derived EVs
- hUMSC-EVs:
-
Human umbilical cord mesenchymal stem cell-derived EVs
- S-MSCs-Exos:
-
S-MSCs-Exosomes
- Wnt:
-
Wingless / integrated
- HMGB:
-
High mobility group protein
- 3'UTR:
-
3' untranslated region
- MSCs:
-
IPFP-ExosIPFP MSCs-derived Exos
- mTOR:
-
Mammalian target of rapamycin
- hAD-MSCs:
-
Human AD-MSCs
- hAD-MSCs-sEVs:
-
hAD-MSCs-derived small EVs
- LFJ OA:
-
Lumbar facet joint OA
- CGRP:
-
Calcitonin gene related peptide
- RANKL:
-
Receptor activator of nuclear factor-κ B ligand
- RANK:
-
Receptor activator of nuclear factor-κ B
- TRAF6:
-
Tumor necrosis factor receptor-associated factor 6
- iNOS:
-
Inducible nitric oxide synthase
- JAK2:
-
Janus kinase 2
- KGN:
-
Kartogenin
- PTEN:
-
Phosphatase and tensin homolog
- Akt:
-
Protein kinase B
- lncRNA:
-
Long noncoding RNA
- FOXO3:
-
Forkhead box O3
- 3D:
-
Three-dimensional
- 2D:
-
Two-dimensional
- LAMP-2B:
-
Lysosome-associated membrane protein 2
- CAP:
-
Chondrocyte affinity peptide
- SF-MSC:
-
Synovial fluid mesenchymal stem cell
- NF-kB:
-
Nuclear factor kappa B
- IFN-γ:
-
Interferon-γ
- TGF:
-
Transforming growth factor
- Arg-1:
-
Arginase-1
- FGF:
-
Fibroblast growth factor
- SOX9:
-
SRY box gene-9
- COMP:
-
Cartilage oligomeric matrix protein
- Runx2:
-
Runt-related transcription factor 2
References
Lourido L, et al. Secretome analysis of human articular chondrocytes unravels catabolic effects of nicotine on the joint. Proteomics Clin Appl. 2016;10(6):671–80.
Mobasheri A. The future of osteoarthritis therapeutics: emerging biological therapy. Curr Rheumatol Rep. 2013;15(12):385.
Loeser RF, et al. Osteoarthritis: a disease of the joint as an organ. Arthritis Rheum. 2012;64(6):1697–707.
Szekanecz Z, et al. New insights in synovial angiogenesis. Joint Bone Spine. 2010;77(1):13–9.
Sellam J, Berenbaum F. Is osteoarthritis a metabolic disease? Joint Bone Spine. 2013;80(6):568–73.
Scanzello CR, Goldring SR. The role of synovitis in osteoarthritis pathogenesis. Bone. 2012;51(2):249–57.
Krenn V, et al. Synovitis score: discrimination between chronic low-grade and high-grade synovitis. Histopathology. 2006;49(4):358–64.
Ayral X, et al. Synovitis: a potential predictive factor of structural progression of medial tibiofemoral knee osteoarthritis – results of a 1 year longitudinal arthroscopic study in 422 patients. Osteoarthritis Cartilage. 2005;13(5):361–7.
Benito MJ, et al. Synovial tissue inflammation in early and late osteoarthritis. Ann Rheum Dis. 2005;64(9):1263–7.
Manferdini C, et al. From osteoarthritic synovium to synovial-derived cells characterization: synovial macrophages are key effector cells. Arthritis Res Ther. 2016;18:83.
Schelbergen RF, et al. Alarmins S100A8 and S100A9 elicit a catabolic effect in human osteoarthritic chondrocytes that is dependent on Toll-like receptor 4. Arthritis Rheum. 2012;64(5):1477–87.
Chadha R. Revealed aspect of metabolic osteoarthritis. J Orthop. 2016;13(4):347–51.
Liu-Bryan R. Synovium and the innate inflammatory network in osteoarthritis progression. Curr Rheumatol Rep. 2013;15(5):323.
Sohn DH, et al. Plasma proteins present in osteoarthritic synovial fluid can stimulate cytokine production via Toll-like receptor 4. Arthritis Res Ther. 2012;14(1):R7.
Piccinini AM, Midwood KS. DAMPening inflammation by modulating TLR signalling. Mediators Inflamm. 2010;2010:672395.
Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. 2010;11(5):373–84.
Kim HA, et al. The catabolic pathway mediated by Toll-like receptors in human osteoarthritic chondrocytes. Arthritis Rheum. 2006;54(7):2152–63.
Goldring SR, Goldring MB. Clinical aspects, pathology and pathophysiology of osteoarthritis. J Musculoskelet Neuronal Interact. 2006;6(4):376–8.
Liu Y, et al. MSC-derived exosomes promote proliferation and inhibit apoptosis of chondrocytes via lncRNA-KLF3-AS1/miR-206/GIT1 axis in osteoarthritis. Cell Cycle. 2018;17(21–22):2411–22.
Mao G, et al. Exosomal miR-95-5p regulates chondrogenesis and cartilage degradation via histone deacetylase 2/8. J Cell Mol Med. 2018;22(11):5354–66.
Wang Y, et al. Exosomes from embryonic mesenchymal stem cells alleviate osteoarthritis through balancing synthesis and degradation of cartilage extracellular matrix. Stem Cell Res Ther. 2017;8(1):189.
Kato T, et al. Exosomes from IL-1beta stimulated synovial fibroblasts induce osteoarthritic changes in articular chondrocytes. Arthritis Res Ther. 2014;16(4):R163.
Rogoveanu OC, et al. Association of cytokine gene polymorphisms with osteoarthritis susceptibility. Exp Ther Med. 2018;16(3):2659–64.
Vincent TL. IL-1 in osteoarthritis: time for a critical review of the literature. F1000Res. 2019;8:F1000 Faculty Rev-934.
Attur M, et al. Low-grade inflammation in symptomatic knee osteoarthritis: prognostic value of inflammatory plasma lipids and peripheral blood leukocyte biomarkers. Arthritis Rheumatol. 2015;67(11):2905–15.
Wojdasiewicz P, Poniatowski LA, Szukiewicz D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediators Inflamm. 2014;2014:561459.
Wang T, He C. Pro-inflammatory cytokines: The link between obesity and osteoarthritis. Cytokine Growth Factor Rev. 2018;44:38–50.
Afonso V, et al. Reactive oxygen species and superoxide dismutases: role in joint diseases. Joint Bone Spine. 2007;74(4):324–9.
Singh M, et al. Promoter polymorphisms in IL-6 gene influence pro-inflammatory cytokines for the risk of osteoarthritis. Cytokine. 2020;127:154985.
Kim HR, et al. Macrophage migration inhibitory factor upregulates angiogenic factors and correlates with clinical measures in rheumatoid arthritis. J Rheumatol. 2007;34(5):927–36.
Wang Y, et al. TNF-alpha-induced LRG1 promotes angiogenesis and mesenchymal stem cell migration in the subchondral bone during osteoarthritis. Cell Death Dis. 2017;8(3):e2715.
Charlier E, et al. Insights on Molecular Mechanisms of Chondrocytes Death in Osteoarthritis. Int J Mol Sci. 2016;17(12):2146.
Mehana EE, Khafaga AF, El-Blehi SS. The role of matrix metalloproteinases in osteoarthritis pathogenesis: An updated review. Life Sci. 2019;234:116786.
Alonso B, et al. Osteoarthritis-related biomarkers profile in chronic anterior cruciate ligament injured knee. Knee. 2020;27(1):51–60.
Duan L, et al. Infrapatellar fat pads participate in the development of knee osteoarthritis in obese patients via the activation of the NFkappaB signaling pathway. Int J Mol Med. 2020;46(6):2260–70.
Li H, et al. New insights on the MMP-13 regulatory network in the pathogenesis of early osteoarthritis. Arthritis Res Ther. 2017;19(1):248.
Chan CM, et al. Cytokine-induced MMP13 Expression in Human Chondrocytes Is Dependent on Activating Transcription Factor 3 (ATF3) Regulation. J Biol Chem. 2017;292(5):1625–36.
Malemud CJ. Inhibition of MMPs and ADAM/ADAMTS. Biochem Pharmacol. 2019;165:33–40.
Yang CY, Chanalaris A, Troeberg L. ADAMTS and ADAM metalloproteinases in osteoarthritis - looking beyond the “usual suspects.” Osteoarthritis Cartilage. 2017;25(7):1000–9.
Bay-Jensen AC, et al. Aggrecanase degradation of type III collagen is associated with clinical knee pain. Clin Biochem. 2018;58:37–43.
Liu Z, et al. Evaluation of Biomarkers and Immune Microenvironment of Osteoarthritis: Evidence From Omics Data and Machine Learning. Front Genet. 2022;13:905027.
Lin XC, et al. B-cell-specific mammalian target of rapamycin complex 1 activation results in severe osteoarthritis in mice. Int Immunopharmacol. 2018;65:522–30.
Jasin HE. Autoantibody specificities of immune complexes sequestered in articular cartilage of patients with rheumatoid arthritis and osteoarthritis. Arthritis Rheum. 1985;28(3):241–8.
Wang Q, et al. IgE-mediated mast cell activation promotes inflammation and cartilage destruction in osteoarthritis. Elife. 2019;8:e39905.
Woolley DE, Tetlow LC. Mast cell activation and its relation to proinflammatory cytokine production in the rheumatoid lesion. Arthritis Res. 2000;2(1):65–74.
de Lange-Brokaar BJ, et al. Characterization of synovial mast cells in knee osteoarthritis: association with clinical parameters. Osteoarthritis Cartilage. 2016;24(4):664–71.
Zhang H, Cai D, Bai X. Macrophages regulate the progression of osteoarthritis. Osteoarthritis Cartilage. 2020;28(5):555–61.
Wu CL, et al. The role of macrophages in osteoarthritis and cartilage repair. Osteoarthritis Cartilage. 2020;28(5):544–54.
Ziadlou R, et al. Anti-Inflammatory and Chondroprotective Effects of Vanillic Acid and Epimedin C in Human Osteoarthritic Chondrocytes. Biomolecules. 2020;10(6):932.
Ezawa K, et al. Comparative analysis of CD45RA- and CD45RO-positive CD4+T cells in peripheral blood, synovial fluid, and synovial tissue in patients with rheumatoid arthritis and osteoarthritis. Acta Med Okayama. 1997;51(1):25–31.
Benigni G, et al. CXCR3/CXCL10 Axis Regulates Neutrophil-NK Cell Cross-Talk Determining the Severity of Experimental Osteoarthritis. J Immunol. 2017;198(5):2115–24.
Spaggiari GM, et al. Mesenchymal stem cell-natural killer cell interactions: evidence that activated NK cells are capable of killing MSCs, whereas MSCs can inhibit IL-2-induced NK-cell proliferation. Blood. 2006;107(4):1484–90.
Takeda H, et al. Effect of IL-15 and natural killer cells on osteoclasts and osteoblasts in a mouse coculture. Inflammation. 2014;37(3):657–69.
Rahmati M, et al. Aging and osteoarthritis: Central role of the extracellular matrix. Ageing Res Rev. 2017;40:20–30.
Anderson HC. Vesicles associated with calcification in the matrix of epiphyseal cartilage. J Cell Biol. 1969;41(1):59–72.
Wolf P. The nature and significance of platelet products in human plasma. Br J Haematol. 1967;13(3):269–88.
Gardiner C, et al. Techniques used for the isolation and characterization of extracellular vesicles: results of a worldwide survey. J Extracell Vesicles. 2016;5:32945.
Rekker K, et al. Comparison of serum exosome isolation methods for microRNA profiling. Clin Biochem. 2014;47(1–2):135–8.
Alvarez ML, et al. Comparison of protein, microRNA, and mRNA yields using different methods of urinary exosome isolation for the discovery of kidney disease biomarkers. Kidney Int. 2012;82(9):1024–32.
Zhu L, et al. Isolation and characterization of exosomes for cancer research. J Hematol Oncol. 2020;13(1):152.
Greening DW, et al. A protocol for exosome isolation and characterization: evaluation of ultracentrifugation, density-gradient separation, and immunoaffinity capture methods. Methods Mol Biol. 2015;1295:179–209.
Heinemann ML, et al. Benchtop isolation and characterization of functional exosomes by sequential filtration. J Chromatogr A. 2014;1371:125–35.
Boing AN, et al. Single-step isolation of extracellular vesicles by size-exclusion chromatography. J Extracell Vesicles. 2014;3(1):2001–3078.
Tauro BJ, et al. Comparison of ultracentrifugation, density gradient separation, and immunoaffinity capture methods for isolating human colon cancer cell line LIM1863-derived exosomes. Methods. 2012;56(2):293–304.
McNamara RP, et al. Large-scale, cross-flow based isolation of highly pure and endocytosis-competent extracellular vesicles. J Extracell Vesicles. 2018;7(1):1541396.
Shao H, et al. New Technologies for Analysis of Extracellular Vesicles. Chem Rev. 2018;118(4):1917–50.
Thery C, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750.
Lee K, et al. Multiplexed Profiling of Single Extracellular Vesicles. ACS Nano. 2018;12(1):494–503.
Varderidou-Minasian S, Lorenowicz MJ. Mesenchymal stromal/stem cell-derived extracellular vesicles in tissue repair: challenges and opportunities. Theranostics. 2020;10(13):5979–97.
Kim GB, et al. Mesenchymal Stem Cell-Derived Exosomes and Their Therapeutic Potential for Osteoarthritis. Biology (Basel). 2021;10(4):285.
Mendt M, et al. Generation and testing of clinical-grade exosomes for pancreatic cancer. JCI Insight. 2018;3(8):e99263.
Alvarez-Erviti L, et al. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol. 2011;29(4):341–5.
Tao SC, et al. Exosomes derived from miR-140-5p-overexpressing human synovial mesenchymal stem cells enhance cartilage tissue regeneration and prevent osteoarthritis of the knee in a rat model. Theranostics. 2017;7(1):180–95.
Zhu Y, et al. Comparison of exosomes secreted by induced pluripotent stem cell-derived mesenchymal stem cells and synovial membrane-derived mesenchymal stem cells for the treatment of osteoarthritis. Stem Cell Res Ther. 2017;8(1):64.
Wang Z, et al. Exosomes derived from miR-155-5p-overexpressing synovial mesenchymal stem cells prevent osteoarthritis via enhancing proliferation and migration, attenuating apoptosis, and modulating extracellular matrix secretion in chondrocytes. Cell Biol Toxicol. 2021;37(1):85–96.
Qiu M, Liu D, Fu Q. MiR-129–5p shuttled by human synovial mesenchymal stem cell-derived exosomes relieves IL-1beta induced osteoarthritis via targeting HMGB1. Life Sci. 2021;269:118987.
Tofino-Vian M, et al. Extracellular Vesicles from Adipose-Derived Mesenchymal Stem Cells Downregulate Senescence Features in Osteoarthritic Osteoblasts. Oxid Med Cell Longev. 2017;2017:7197598.
Wu J, et al. miR-100–5p-abundant exosomes derived from infrapatellar fat pad MSCs protect articular cartilage and ameliorate gait abnormalities via inhibition of mTOR in osteoarthritis. Biomaterials. 2019;206:87–100.
Tofino-Vian M, et al. Microvesicles from Human Adipose Tissue-Derived Mesenchymal Stem Cells as a New Protective Strategy in Osteoarthritic Chondrocytes. Cell Physiol Biochem. 2018;47(1):11–25.
Woo CH, et al. Small extracellular vesicles from human adipose-derived stem cells attenuate cartilage degeneration. J Extracell Vesicles. 2020;9(1):1735249.
Wang R, Xu B, Xu H. TGF-beta1 promoted chondrocyte proliferation by regulating Sp1 through MSC-exosomes derived miR-135b. Cell Cycle. 2018;17(24):2756–65.
Qi H, et al. Exosomes derived from mesenchymal stem cells inhibit mitochondrial dysfunction-induced apoptosis of chondrocytes via p38, ERK, and Akt pathways. In Vitro Cell Dev Biol Anim. 2019;55(3):203–10.
Cosenza S, et al. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci Rep. 2017;7(1):16214.
Rong Y, et al. Hypoxic pretreatment of small extracellular vesicles mediates cartilage repair in osteoarthritis by delivering miR-216a-5p. Acta Biomater. 2021;122:325–42.
Hu H, et al. miR-23a-3p-abundant small extracellular vesicles released from Gelma/nanoclay hydrogel for cartilage regeneration. J Extracell Vesicles. 2020;9(1):1778883.
Yan L, Liu G, Wu X. The umbilical cord mesenchymal stem cell-derived exosomal lncRNA H19 improves osteochondral activity through miR-29b-3p/FoxO3 axis. Clin Transl Med. 2021;11(1):e255.
Yan L, Wu X. Exosomes produced from 3D cultures of umbilical cord mesenchymal stem cells in a hollow-fiber bioreactor show improved osteochondral regeneration activity. Cell Biol Toxicol. 2020;36(2):165–78.
De Bari C, et al. Multipotent mesenchymal stem cells from adult human synovial membrane. Arthritis Rheum. 2001;44(8):1928–42.
Jones BA, Pei M. Synovium-derived stem cells: a tissue-specific stem cell for cartilage engineering and regeneration. Tissue Eng Part B Rev. 2012;18(4):301–11.
Miyamoto C, et al. Osteogenic protein-1 with transforming growth factor-beta1: potent inducer of chondrogenesis of synovial mesenchymal stem cells in vitro. J Orthop Sci. 2007;12(6):555–61.
Kurth T, et al. Chondrogenic potential of human synovial mesenchymal stem cells in alginate. Osteoarthritis Cartilage. 2007;15(10):1178–89.
Shirasawa S, et al. In vitro chondrogenesis of human synovium-derived mesenchymal stem cells: optimal condition and comparison with bone marrow-derived cells. J Cell Biochem. 2006;97(1):84–97.
Kondo S, et al. Transplantation of Aggregates of Autologous Synovial Mesenchymal Stem Cells for Treatment of Cartilage Defects in the Femoral Condyle and the Femoral Groove in Microminipigs. Am J Sports Med. 2019;47(10):2338–47.
Enomoto T, et al. Timing of Intra-Articular Injection of Synovial Mesenchymal Stem Cells Affects Cartilage Restoration in a Partial Thickness Cartilage Defect Model in Rats. Cartilage. 2020;11(1):122–9.
Mak J, et al. Intra-articular injection of synovial mesenchymal stem cells improves cartilage repair in a mouse injury model. Sci Rep. 2016;6:23076.
Ozeki N, et al. Not single but periodic injections of synovial mesenchymal stem cells maintain viable cells in knees and inhibit osteoarthritis progression in rats. Osteoarthritis Cartilage. 2016;24(6):1061–70.
Koizumi K, et al. Synovial mesenchymal stem cells from osteo- or rheumatoid arthritis joints exhibit good potential for cartilage repair using a scaffold-free tissue engineering approach. Osteoarthritis Cartilage. 2016;24(8):1413–22.
Lee WS, et al. Intra-Articular Injection of Autologous Adipose Tissue-Derived Mesenchymal Stem Cells for the Treatment of Knee Osteoarthritis: A Phase IIb, Randomized, Placebo-Controlled Clinical Trial. Stem Cells Transl Med. 2019;8(6):504–11.
Kim YS, Koh YG. Comparative Matched-Pair Analysis of Open-Wedge High Tibial Osteotomy With Versus Without an Injection of Adipose-Derived Mesenchymal Stem Cells for Varus Knee Osteoarthritis: Clinical and Second-Look Arthroscopic Results. Am J Sports Med. 2018;46(11):2669–77.
Damia E, et al. Adipose-Derived Mesenchymal Stem Cells: Are They a Good Therapeutic Strategy for Osteoarthritis? Int J Mol Sci. 2018;19(7):1926.
Skalska U, Kontny E. Adipose-derived mesenchymal stem cells from infrapatellar fat pad of patients with rheumatoid arthritis and osteoarthritis have comparable immunomodulatory properties. Autoimmunity. 2016;49(2):124–31.
Maumus M, et al. Adipose mesenchymal stem cells protect chondrocytes from degeneration associated with osteoarthritis. Stem Cell Res. 2013;11(2):834–44.
Manferdini C, et al. Adipose-derived mesenchymal stem cells exert antiinflammatory effects on chondrocytes and synoviocytes from osteoarthritis patients through prostaglandin E2. Arthritis Rheum. 2013;65(5):1271–81.
Xu Y, et al. microRNA-16-5p-containing exosomes derived from bone marrow-derived mesenchymal stem cells inhibit proliferation, migration, and invasion, while promoting apoptosis of colorectal cancer cells by downregulating ITGA2. J Cell Physiol. 2019;234(11):21380–94.
Xie C, et al. Exosomes derived from microRNA-101-3p-overexpressing human bone marrow mesenchymal stem cells suppress oral cancer cell proliferation, invasion, and migration. Mol Cell Biochem. 2019;458(1–2):11–26.
Zhang L, et al. Exosomes from bone marrow mesenchymal stem cells enhance fracture healing through the promotion of osteogenesis and angiogenesis in a rat model of nonunion. Stem Cell Res Ther. 2020;11(1):38.
Zhao C, et al. Exosomes Derived From Bone Marrow Mesenchymal Stem Cells Inhibit Complement Activation In Rats With Spinal Cord Injury. Drug Des Devel Ther. 2019;13:3693–704.
Rong X, et al. Human bone marrow mesenchymal stem cells-derived exosomes alleviate liver fibrosis through the Wnt/beta-catenin pathway. Stem Cell Res Ther. 2019;10(1):98.
He JG, et al. Exosomes Derived from IDO1-Overexpressing Rat Bone Marrow Mesenchymal Stem Cells Promote Immunotolerance of Cardiac Allografts. Cell Transplant. 2018;27(11):1657–83.
Xiong L, et al. Exosomes from Bone Marrow Mesenchymal Stem Cells Can Alleviate Early Brain Injury After Subarachnoid Hemorrhage Through miRNA129-5p-HMGB1 Pathway. Stem Cells Dev. 2020;29(4):212–21.
Mianehsaz E, et al. Mesenchymal stem cell-derived exosomes: a new therapeutic approach to osteoarthritis? Stem Cell Res Ther. 2019;10(1):340.
Asghar S, et al. Exosomes in intercellular communication and implications for osteoarthritis. Rheumatology (Oxford). 2020;59(1):57–68.
Ding J, et al. Exosomes Derived from Human Bone Marrow Mesenchymal Stem Cells Stimulated by Deferoxamine Accelerate Cutaneous Wound Healing by Promoting Angiogenesis. Biomed Res Int. 2019;2019:9742765.
Zhu G, et al. Exosomes from human-bone-marrow-derived mesenchymal stem cells protect against renal ischemia/reperfusion injury via transferring miR-199a-3p. J Cell Physiol. 2019;234(12):23736–49.
Zhao L, et al. Exosomes derived from bone marrow mesenchymal stem cells overexpressing microRNA-25 protect spinal cords against transient ischemia. J Thorac Cardiovasc Surg. 2019;157(2):508–17.
Li J, et al. BMSCs-Derived Exosomes Ameliorate Pain Via Abrogation of Aberrant Nerve Invasion in Subchondral Bone in Lumbar Facet Joint Osteoarthritis. J Orthop Res. 2020;38(3):670–9.
Liu C, et al. Kartogenin enhances the therapeutic effect of bone marrow mesenchymal stem cells derived exosomes in cartilage repair. Nanomedicine (Lond). 2020;15(3):273–88.
Zhang YN, Lie PC, Wei X. Differentiation of mesenchymal stromal cells derived from umbilical cord Wharton’s jelly into hepatocyte-like cells. Cytotherapy. 2009;11(5):548–58.
Zhao Q, et al. Differentiation of human umbilical cord mesenchymal stromal cells into low immunogenic hepatocyte-like cells. Cytotherapy. 2009;11(4):414–26.
Ji GZ, Wei X, Chen GQ. Growth of human umbilical cord Wharton’s Jelly-derived mesenchymal stem cells on the terpolyester poly(3-hydroxybutyrate-co-3-hydroxyvalerate-co-3-hydroxyhexanoate). J Biomater Sci Polym Ed. 2009;20(3):325–39.
Zhang S, et al. MSC exosomes mediate cartilage repair by enhancing proliferation, attenuating apoptosis and modulating immune reactivity. Biomaterials. 2018;156:16–27.
Liu X, et al. Integration of stem cell-derived exosomes with in situ hydrogel glue as a promising tissue patch for articular cartilage regeneration. Nanoscale. 2017;9(13):4430–8.
Sang X, et al. Thermosensitive Hydrogel Loaded with Primary Chondrocyte-Derived Exosomes Promotes Cartilage Repair by Regulating Macrophage Polarization in Osteoarthritis. Tissue Eng Regen Med. 2022;19(3):629–42.
Zheng L, et al. Primary chondrocyte exosomes mediate osteoarthritis progression by regulating mitochondrion and immune reactivity. Nanomedicine (Lond). 2019;14(24):3193–212.
Da-Wa ZX, et al. Exosomes Derived from M2 Macrophages Exert a Therapeutic Effect via Inhibition of the PI3K/AKT/mTOR Pathway in Rats with Knee Osteoarthritic. Biomed Res Int. 2021;2021:7218067.
Tan F, Wang D, Yuan Z. The Fibroblast-Like Synoviocyte Derived Exosomal Long Non-coding RNA H19 Alleviates Osteoarthritis Progression Through the miR-106b-5p/TIMP2 Axis. Inflammation. 2020;43(4):1498–509.
Liang Y, et al. Chondrocyte-Targeted MicroRNA Delivery by Engineered Exosomes toward a Cell-Free Osteoarthritis Therapy. ACS Appl Mater Interfaces. 2020;12(33):36938–47.
Xu X, et al. Exosome-mediated delivery of kartogenin for chondrogenesis of synovial fluid-derived mesenchymal stem cells and cartilage regeneration. Biomaterials. 2021;269:120539.
Heard BJ, et al. Single intra-articular dexamethasone injection immediately post-surgery in a rabbit model mitigates early inflammatory responses and post-traumatic osteoarthritis-like alterations. J Orthop Res. 2015;33(12):1826–34.
Xia B, et al. Osteoarthritis pathogenesis: a review of molecular mechanisms. Calcif Tissue Int. 2014;95(6):495–505.
Zhao C, et al. Exosomes from adiposederived stem cells promote chondrogenesis and suppress inflammation by upregulating miR145 and miR221. Mol Med Rep. 2020;21(4):1881–9.
Haslauer CM, et al. Loss of extracellular matrix from articular cartilage is mediated by the synovium and ligament after anterior cruciate ligament injury. Osteoarthritis Cartilage. 2013;21(12):1950–7.
Fahy N, et al. Human osteoarthritic synovium impacts chondrogenic differentiation of mesenchymal stem cells via macrophage polarisation state. Osteoarthritis Cartilage. 2014;22(8):1167–75.
Ding J, et al. Bone Marrow Mesenchymal Stem Cell-Based Engineered Cartilage Ameliorates Polyglycolic Acid/Polylactic Acid Scaffold-Induced Inflammation Through M2 Polarization of Macrophages in a Pig Model. Stem Cells Transl Med. 2016;5(8):1079–89.
Oishi Y, Manabe I. Macrophages in inflammation, repair and regeneration. Int Immunol. 2018;30(11):511–28.
Wynn TA, Vannella KM. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity. 2016;44(3):450–62.
Fernandes TL, et al. Macrophage: A Potential Target on Cartilage Regeneration. Front Immunol. 2020;11:111.
Zhang H, et al. Synovial macrophage M1 polarisation exacerbates experimental osteoarthritis partially through R-spondin-2. Ann Rheum Dis. 2018;77(10):1524–34.
Cosenza S, et al. Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis. Theranostics. 2018;8(5):1399–410.
Chen W, et al. Immunomodulatory effects of mesenchymal stromal cells-derived exosome. Immunol Res. 2016;64(4):831–40.
Tkach M, et al. Qualitative differences in T-cell activation by dendritic cell-derived extracellular vesicle subtypes. EMBO J. 2017;36(20):3012–28.
Pang XL, et al. Immature dendritic cells derived exosomes promotes immune tolerance by regulating T cell differentiation in renal transplantation. Aging (Albany NY). 2019;11(20):8911–24.
Hosseini R, et al. Cancer exosomes and natural killer cells dysfunction: biological roles, clinical significance and implications for immunotherapy. Mol Cancer. 2022;21(1):15.
Aung T, et al. Exosomal evasion of humoral immunotherapy in aggressive B-cell lymphoma modulated by ATP-binding cassette transporter A3. Proc Natl Acad Sci U S A. 2011;108(37):15336–41.
Riazifar M, et al. Stem Cell Extracellular Vesicles: Extended Messages of Regeneration. Annu Rev Pharmacol Toxicol. 2017;57:125–54.
Vonk LA, et al. Mesenchymal Stromal/stem Cell-derived Extracellular Vesicles Promote Human Cartilage Regeneration In Vitro. Theranostics. 2018;8(4):906–20.
Ragni E, et al. Secreted Factors and EV-miRNAs Orchestrate the Healing Capacity of Adipose Mesenchymal Stem Cells for the Treatment of Knee Osteoarthritis. Int J Mol Sci. 2020;21(5):1582.
Chen P, et al. Desktop-stereolithography 3D printing of a radially oriented extracellular matrix/mesenchymal stem cell exosome bioink for osteochondral defect regeneration. Theranostics. 2019;9(9):2439–59.
Li Q, et al. The tissue origin effect of extracellular vesicles on cartilage and bone regeneration. Acta Biomater. 2021;125:253–66.
Chen X, et al. Mesenchymal stem cell-derived exosomal microRNA-136-5p inhibits chondrocyte degeneration in traumatic osteoarthritis by targeting ELF3. Arthritis Res Ther. 2020;22(1):256.
Mao G, et al. Exosomes derived from miR-92a-3p-overexpressing human mesenchymal stem cells enhance chondrogenesis and suppress cartilage degradation via targeting WNT5A. Stem Cell Res Ther. 2018;9(1):247.
Zhang R, et al. Mesenchymal stem cell related therapies for cartilage lesions and osteoarthritis. Am J Transl Res. 2019;11(10):6275–89.
Vizoso FJ, et al. Mesenchymal Stem Cells in Homeostasis and Systemic Diseases: Hypothesis, Evidences, and Therapeutic Opportunities. Int J Mol Sci. 2019;20(15):3738.
Fang W, Vangsness CT Jr. Implications of Anti-Inflammatory Nature of Exosomes in Knee Arthritis. Cartilage. 2021;13(2_suppl):204S-212S.
Zhang S, et al. Exosomes derived from human embryonic mesenchymal stem cells promote osteochondral regeneration. Osteoarthritis Cartilage. 2016;24(12):2135–40.
Liu Y, et al. Exosomal KLF3-AS1 from hMSCs promoted cartilage repair and chondrocyte proliferation in osteoarthritis. Biochem J. 2018;475(22):3629–38.
Li LL, et al. Repair of osteoarthritis in animal model with exosomes derived from BMSCs transfected by the siRNA -Piezo1 through CT navigation. Zhongguo Gu Shang. 2021;34(12):1171–8.
Katsuda T, et al. The therapeutic potential of mesenchymal stem cell-derived extracellular vesicles. Proteomics. 2013;13(10–11):1637–53.
Acknowledgements
This work was supported by the National Key R&D Program of China (2019YFA0110600), National Natural Science Foundation of China (82102552, 81900958, 81772319), the National Science Foundation of Guangdong Province (2020A1515010059), and the China Postdoctoral Science Foundation Grant (2019TQ0379, 2019M663262).
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
The manuscript was mainly designed by QG, WG, JZ and SL. HY, ML and GT wrote the manuscript. YM, CN, and ZY contributed to the collection of data. JW, QG and XS revised the content of the manuscript. All authors reviewed the manuscript, contributed to its content and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors have no competing interests to declare.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yin, H., Li, M., Tian, G. et al. The role of extracellular vesicles in osteoarthritis treatment via microenvironment regulation. Biomater Res 26, 52 (2022). https://doi.org/10.1186/s40824-022-00300-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40824-022-00300-7