
Abstract
Truffles, the symbiotic hypogeous edible fungi, have been worldwide regarded as a great delicacy because of their unique flavor and high nutritional value. By identifying their bioactive components such as phenolics, terpenoids, polysaccharides, anandamide, fatty acids, and ergosterols, researchers have paid attention to their biological activities including antitumor, antioxidant, antibacterial, anti-inflammatory, and hepatoprotective activities. In addition, numerous factors have been investigating that can affect the quality and productivity of truffles to overcome their difficulty in culturing and preserving. To provide the information for their potential applications in medicine as well as in functional food, this review summarizes the relevant literature about the biochemical composition, aromatic and nutritional benefits, and biological properties of truffles. Besides, various factors affecting their productivity and quality as well as the preservation methods are also highlighted.
Similar content being viewed by others

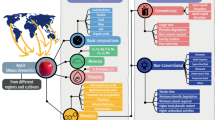

Background
Truffles are hypogeous ascomycetes fungi growing underground in depth between 5 and 10 cm. More than a hundred different kinds of truffle species were known worldwide, and new species are being discovered consistently. Taxonomically, edible truffles belong to family Tuberaceae and Pezizaceae, and to order Pezizales [1]. Tuberaceae is showing the diverse lineages of exclusively truffle forming fungi, and truffles belonging to this family have been widely consumed for centuries [2]. For example, the genus Tuber from the Tuberaceae family comprised of approximately more than 180 species globally [3]. Most culinary-grade truffles discovered in northern temperate forests of Europe, Australia, New Zealand, Asia, and North America generally belong to genus Tuber [4, 5]. On the other hand, desert truffles growing in the arid and semi-arid regions such as Syria, Iraq, Kuwait, Saudi Arabia, Morocco, Egypt, South Africa, and Tunisia, generally belong to the genus Terfezia and Tirmania [1, 6]. More details in the phylogeny of truffles are presented in Fig. 1.

A schematic overview of the three major clades identified in Pezizales. Truffles, their genus, and family that mainly introduced in this review are shown in bold
In general, truffles tend to be firm, dense, and woody in comparison with mushrooms that is soft and fragile. White truffles include Tuber magnatum, T. borchii, T. maculatum, T. oregonense, T. latisporum, T. japonicum, and Tirmania nivea [7]. Among them, T. magnatum generally found in Italy and East Europe is the high cost truffle species [8]. In 2000 and 2001, it reached €8000 per kilogram on several Italian markets [9]. Black truffles include Tuber melanosporum, T. aestivum, T. brumale, T. uncinatum, T. indicum, T. himalayense, and Terfezia claveryi [7]. Truffles exhibit a large genome size. For example, T. melanosporum showed a genome size of 125 Mb with a high transposable element and repetitive DNA content which comprised more than 58% of the genome [10].
Truffles are ectomycorrhizal having symbiotic root association. In a complex life cycle, the mycelia establish a symbiotic interaction with host organisms predominantly with the roots of various trees, both gymnosperms, and angiosperms, such as hazel, poplar, pine, eucalyptus, and oak [8, 11]. Because truffles produce their sexual fruiting bodies underground, the dispersal of spore is relying on insects and mammals [2]. Once ascospores are dispersed, the haploid spore germinates to haploid free-living mycelium establishing ectomycorrhizal association with the roots of host trees. Then, hyphae aggregate and form the sexual fruiting body which is an ascoma bearing asci (Fig. 2) [12,13,14].

Symbiotic life cycle of truffles
The cultivation and storage of truffles are hard to control due to their dependence on several biotic and abiotic factors such as climate, humidity, soil conditions, and the surrounding flora and fauna [15]. Moreover, climate change such as increased summer temperature together with increased summer hydric stress and desertification, has led to a drop in the truffle production [16]. In France harvesting predominantly T. melanosporum, truffle harvest reached 2000 tons by more than 40 departments in the nineteenth century. However, in 2003–2004, the truffle production represented only ~ 10 tons mainly by Vaucluse, Dordogne, Gard, Lot, and Alpes de Haute-Provence. Nowadays, only ~ 20 tons of truffles are harvested worldwide, and the demand for truffles exceeds their supply [11, 17]. To overcome the gap between demand and supply of truffles, some commercial farming companies such as Gazzarrini Tartufi, La Maison Plantin, and La Truffe du Ventoux, have been increasing researches and developing the cultivation techniques. It has been reported that the global truffles market size will be increased by $375.3 million during 2019–2023, and Europe will account for the highest truffles market throughout this period [18].
Although truffles are regarded as a costly delicacy, they are worldwide appreciated as valuable foodstuffs due to their distinctive flavor [19, 20]. The aroma of truffles can range from mild to intense and vary from garlicky, pungent, vanilla-like, creamy, pungent, and dusty like [21]. Among hundreds of aroma active compounds in truffles, 2-methylbutanal, 3-methylbutanal, dimethyl disulphide (DMDS), dimethyl sulphide (DMS) are the most common natural aroma compounds. However, truffle derived products have a short shelf-life and are only available during the truffle season, the food industry has been developing the synthetic flavors. The exact composition of the synthetic aroma to imitate truffle flavor is unknown, even if, for example, the truffle-flavored oils contain more than 60 volatiles, of which 2,4-dithiapentane is the most common [22]. The mixture of DMS and 2-methylbutanal has been also used for a long time by the food industry to reproduce T. melanosporum aroma [15].
Apart from the aromatic feature, their biochemical composition encompasses multiple nutritional and medicinal benefits. Truffles are rich in various types of essential nutritional products including carbohydrates, proteins, fats, minerals, lipids, and amino acids [7]. In addition, they are rich in phenolics, terpenoids, polysaccharides, and phytosterols which are related to their antitumor, antioxidant, antibacterial, hepatoprotective, anti-inflammatory, and immunomodulatory properties [11]. In recent decades, researchers have paid great attention to the bioactive compounds derived from diverse truffle species and their potential in nutritional and medicinal applications [7, 23]. This review summarizes and updates the current status of knowledge on the chemical composition of truffles, their biological activities as well as nutritional and medicinal potential, and various factors contribute to their productivity and quality.
Aroma profiles of truffles
The unique flavor of truffles is one of the main reasons to get worldwide attraction as a food product. Previous studies have focused on screening and identifying the volatile organic compounds (VOCs), namely the aroma active compounds, and characterized more than 200 VOCs in various truffle species. The major compounds responsible for the aroma in diverse truffle species were well documented in 2011 reviews [7, 15]. Our review aimed to provide an update on research conducted since 2011 about the analysis of active aroma components as well as the analytical techniques to identify them (Table 1).
Truffles possess significant variability in their aroma profiles from species to species. In general, sulfur compounds such as dimethyl sulfide (DMS) and dimethyl disulfide (DMDS), 1-octen-3-ol, and 2-methyl-1-propanol have been identified in most truffle species including Tuber melanosporum, Tuber aestivum, and Tuber borchii [15]. In Tuber magnatum (white truffle), bis (methylthio) methane (BMTM) was reported as the most critical aroma active compound [24]. To deepen the knowledge about a complex odor of various truffle species, researchers have developed multiple methods to analyze truffle’s aroma. Traditionally, VOCs have been comprehensively profiled by solid-phase microextraction (SPME), which is generally followed by gas chromatography–mass spectrometry (GC–MS) [25,26,27]. Torregiani et al. analyzed 20 raw truffles, truffle sauces, and truffle-flavored oils using headspace SPME with GC–MS (HS-SPME–GC–MS). Consequently, BMTM was predominantly detected both in raw white truffles and oils flavored with white truffle but not detected or detected only in low amounts in the T. melanosporum (black truffle) samples; instead, 1-octen-3-ol was detected in high amounts in the raw black truffle and oil samples flavored with black truffle [28]. The HS-SPME–GC–MS technique enabled that Liu and Li profiled for the first time a total of 57 volatiles in T. liyuanum; a novel truffle species that have been identified in recent years. In this new truffle species, aldehydes and aromatics were the main chemical constituents, and 3-octanone, phenylethyl alcohol, isopentane, and methylbutane contributed to the significant aromaticity [29].
The GC–MS based analysis expressed limitations to determine the correlation of quantified volatiles to the olfactory stimulus as this technique could not give information about human perception. Moreover, the perceived odor presented frequently at lower concentrations than the instrumental detection limit [30]. To overcome these limitations and to identify the key aroma contributors amongst detected volatiles, the flavor dilution (FD) factor by aroma extract dilution analysis (AEDA) or/and the odor activity value (OAV) could be determined, which are based on the gas chromatography–olfactometry (GC–O) technology [31]. Splivallo and Ebeler performed the GC–O based analysis to investigate whether thiophene derivatives contributed to the human-sensed aroma in T. borchii. Consequently, they identified a total of 13 main aroma active compounds that showed FD factor higher than 5 and determined that thiophene derivatives; particularly in 3-methyl-4,5-dihydrothiophene, were the important contributors to the human-sensed aroma in T. borchii [32]. Feng et al. performed the SPME extraction combined with GC–O technology. They determined 24 compounds as key contributors to the overall aroma in three Chinese truffle varieties which present floral, mushroom, and sweet odor [24]. In addition to these studies, numerous researchers performed GC–O based analysis together with SPME–GC–MS to find the critical aroma contributor among the comprehensively profiled truffle volatiles [19, 32,33,34].
The olfactometry based analysis considering the entire aroma mixture has been regarded as the most useful method for estimating the contribution of key aroma-active compounds. Although it represents the standardized method for determining the aroma concentration, it does not determine the single compounds and their contribution to the aroma, and the large number of dilution of AEDA is a time-consuming process [30]. Liu et al. combined the aroma evaluation method with principal component analysis (PCA) to overcome the limitations of olfactometry based methods for fermentation samples; because the fermentation condition significantly affects aroma attributes, the changing condition one-at-a-time and the repeating AEDA and OAV are laborious. The olfactometry-based method required a total of 5278 dilution samples from 26 fermentation conditions to determine the FD, while the aroma evaluation method scored 504 fermentation samples from various fermentation conditions to identify potential aroma compounds, among them the key volatiles was determined by PCA. Consequently, seven key volatiles identified by GC–O based analysis were also characterized by the new approach along with an additional two significant aroma contributors, 2,3-butanediol, and trimethylpyrazine. The results showed the potential of an aroma evaluation method combined with PCA to study key aroma compounds of fermentation samples [31]. Vita et al. employed time-of-flight (TOF–MS) based Proton Transfer Reaction-Mass Spectrometer (PTR-MS) technology which improved the GC–MS based methods and provided a fast, accurate, and direct measurement of volatiles. This innovative technique enabled the comparison of the volatiles in T. magnatum pico fruiting bodies from Tuscany region with those from the Piedmont region and to distinguish the biological phases of the fruiting bodies [36]. Meanwhile, direct solvent extraction-solvent-assisted flavor evaporation (DSE-SAFE) technique has been applied in many aspects of aroma active compounds such as in fruit and wine because DSE can extract most of the volatiles as well as non-volatiles from samples. DSE, in combination with SAFE, enables the careful and fast isolation of volatiles from solvent extracts [37,38,39]. Zhang et al. employed DSE-SAFE coupled with comprehensive two-dimensional gas chromatography (GC × GC) high-resolution-TOF/MS to distinguish the aroma profiles of Chinese white and black and truffle. They revealed for the first time that the Chinese white truffle exhibited more sulfur-containing volatiles (2.894 µg/g in total) than the Chinese black truffle (0.040 µg/g in total) in terms of both amounts and contents based on fresh weight of samples used in extraction [40].
In recent times, researchers have increasingly recognized the authenticity and traceability of flavor compounds in truffles. The authenticity and traceability can be determined by GC coupled with combustion-isotope ratio mass spectrometry (GC–C–IRMS), which exploits 13C/12C ratio abundance of the main aroma contributor in foods [41, 42]. Wernig et al. used GC-IRMS and GC–MS to discriminate the home-made and commercial truffle oils. The GC-IRMS could not differentiate between natural and synthetic flavors, while the metabolic profiling using GC–MS revealed that two sulfur-containing volatiles, dimethyl sulfone, and dimethyl sulfoxide were exclusively detected in commercial oils [22]. To discriminate the natural aroma from the synthetic one, Sciarrone et al. evaluated bis-(methylthio)methane known as one of the key aroma contributors in T. magnatum pico, using the improved multidimensional GC–C–IRMS with simultaneous quadrupole MS, by minimizing the effect of column bleeding on stable isotope ratio (δ13C) measurement. Consequently, the δ13C values of bis-(methylthio) methane from genuine truffle samples attained between − 42.6 and − 33.9‰, while those from synthetic aroma exhibited more negative values [42]. More details about the recent aroma related studies are listed in Table 1 (cited on page # 35).
The nutritional profile of truffles
Apart from the unique aroma, there are various nutritionally valuable compounds present in truffles. Although the compositional details of truffles vary from species to species and area to area, carbohydrates and proteins are generally the most abundant nutrients in truffles [1]. Besides, minerals, fibers, amino acids, fatty acids, and fats account for a significant portion of truffle composition [7, 43]. For example, a desert truffle genera Terfezia possessed 46–48 g/100 g of dry weight (DW) of carbohydrates, 32–35 g/100 g DW of protein, 14–15 g/100 g DW of ash, and 2.8–3.2 g/100 g DW of fat, while another desert truffle genera Tirmania exhibited lower protein (8–29 g/100 g DW) and ash content (5.1–5.3 g/100 g DW) but higher carbohydrate (58–83 g/100 g DW) and fat content (4–7 g/100 g DW) [44,45,46,47]. According to Yan et al., the dry matters of three Chinese truffle species, T. latisporum, T. subglobosum, and T. pseudohimalayense, were composed of 74–79 g/100 g carbohydrates, 11–15 g/100 g protein, 8.1–8.8 g/100 g ash, 2.2–2.5 g/100 g fat, 24–50 g/100 g total sugars, 96–265 mg/100 g monounsaturated fatty acid, 249–368 mg/100 g polyunsaturated fatty acid, and 70–121 mg/100 g of saturated fatty acid [48].
Various truffle species including T. aestivum, T. borchii, T. magnatum, T. melanosporum, desert truffles, and Chinese truffles exhibited a significant amount of unsaturated fatty acid (UFA) such as oleic acid and linoleic acid, accounting for more than 60% of their total fatty acid content [48, 49]. For example, the fatty acid in the fruiting bodies of T. pinoyi was mainly composed of oleic and linoleic acid, which possessed 32.29% and 29.72%, respectively [45]. Yan et al. demonstrated that linoleic and oleic acid is the most abundant fatty acid in T. latisporum, T. subglobosum, and T. pseudohimalayense. The dried powder of these three Chinese truffles contains 246.81–365.24 mg/100 g linoleic acid and 94.04–250.97 mg/100 g oleic acid, respectively [48]. Linoleic acid is an essential fatty acid and acts as a precursor of 1-octen-3-ol, which is one of the vital aroma active compounds in most truffle species [15, 50]. Oleic acid exhibited potentials in reducing cholesterol levels, preventing cardiovascular, and inhibiting cancer progression [51, 52]. In terms of their high UFA content, truffles might be considered as a potential nutritional and medicinal food.
Minerals are the essential dietary constituents that are required for numerous metabolic reactions, rigid bone formation, the transmission of nerve impulses, and regulation of water and salt balance [53]. Truffles provide a significant amount and variety of minerals. In particular, the major minerals such as potassium, magnesium, calcium, and phosphorus were abundant in European and desert truffles [44, 46, 54, 55]. Truffles are also known as a rich source of amino acids, especially the sulfur-containing amino acids (cysteine and methionine), while they generally remain limited in other plant-derived foods [56].
Extracellular enzymes play a role in nutrient acquisition and symbiotic fungus-host interaction helping truffles to penetrate and colonize into roots of the host plants [57]. T. maculatum produced seven extracellular enzymes, including amylase, catalase, cellulase, laccase, lipase, peroxidase, and xylanase; however, T. aestivum produced amylase, catalase, and peroxidase [58, 59].
Truffles can be considered as a source of protein but not appropriate for human’s daily diet because of their high price [7]. However, it is not suitable to regard them only as a costly source of protein because truffles possess great capability in both the diversity and amount of nutritionally valuable compounds such as fiber and minerals. Moreover, Bouatia et al. demonstrated that T. pinoyi either exhibited no or very low levels of antinutrient compounds including cyanide, oxalate, mercury, arsenic, and cadmium [47]. From this perspective, truffles can be consumed as a potential nutritional source with safety.
Bioactive compounds in truffles
Truffles are rich in bioactive compounds such as ascorbic acid, ergosterol, phenolics, flavonoids, terpenoids, phytosterol, and polysaccharides. In particular, truffles contain abundant flavonoids that are appreciated as an important secondary metabolite of natural products due to their diverse biological properties involving antioxidative, anti-inflammatory, anti-mutagenic, and anticancer properties, while edible mushrooms cannot synthesize, therefore, do not contain these valuable metabolites [60, 61]. From this perspective, researchers have paid attention to the biological activities of truffles and their application in medicinal domains. For example, chemical analysis of three Chinese truffle species revealed that the methanol extracts of them contained abundant antioxidant sources, including 4.59–4.63 g/100 g ascorbic acid, 144–271 mg of β-carotene/100 g carotenoid, 450–735 mg gallic acid equivalents (GAE)/100 g phenolics, and 611–1355 mg of rutin/100 g flavonoid, which could be used to prevent diseases related to oxidative damage [48]. Furthermore, the desert truffle T. nivea exhibited relatively higher total phenolic content (1.39 g GAE/100 g fresh mass (FM)) as compared to other phenolic-rich fruits such as cherries (44–88 mg GAE/100 g FM), strawberries (54–94 mg GAE/100 g FM), or onions (142–428 mg GAE/100 g FM). In particular, their methanolic extract comprised a high number of flavonoids (74.52 mg GAE/g extract) and tannins (19.78 mg catechin equivalent (CE)/g extract). These abundant bioactive compounds in truffle extract were correlated to the 2,2-diphenyl-1-picryhydrazyl (DPPH) radical-scavenging and lipid peroxidation inhibitory properties together with the antimicrobial properties against seven species of bacteria [62,63,64,65]. Ergosterol, the biological precursor of vitamin D2, is also one of the abundant bioactive compounds in diverse truffle species [66]. Villares et al. reported that T. melanosporum, T. aestivum, and T. indicum presented total ergosterol amount of 1.90 mg/g DM, 1.86 mg/g DM, and 1.37 mg/g DM, respectively [67]. This bioactive compound and its derivatives have exhibited various health-promoting activities including antioxidant, anti-inflammatory, and antihyperlipidemic activities [68,69,70].
Biological activities of truffles
Antitumor activity
Recently, many research studies have been conducted to find out the anticancer properties of truffles. According to Beara et al., methanol extracts from T. aestivum and T. magnatum possessed significant in vitro cytotoxic effects against different cancer cell lines (HeLa, MCF-7, and HT-29), besides the prominent activity of the water extract towards breast adenocarcinoma (MCF-7) [71]. In another study conducted by Khadri et al., silver nanoparticles were synthesized from aqueous extract of T. claveryi, which is particularly rich in amino acids and flavonoids. The synthesized nanoparticles exhibited significant in vitro cytotoxic effects (IC50 = 10 mg/mL) against MCF-7, which was evaluated by the colorimetric sulforhodamine B assay [72]. Using MTT (3-[4, 5-dimethylthiazol-2-yl]-2, 5 diphenyl tetrazolium bromide) assay, Dahham et al. determined the significant in vitro cytotoxic effects of hexane and ethyl acetate extract obtained from T. claveryi on different cancer cell lines, including the human brain carcinoma cell line (U-87 MG) with IC50 values of 50.3 µg/mL DW of extract. Furthermore, this study showed the anti-angiogenic efficacy of hexane extract using rat aorta ring assay, the hexane extract inducing the reduction of mitochondrial membrane potential as well as the condensation of nuclei in U-87 MG cells. Their anticancer properties would be correlated to their bioactive compounds including stigmasterol, β-sitosterol, squalene, and lupeol [73]. Phytosterols played a role in the inhibition of cancer cell growth mechanism, angiogenesis, and stimulation of cancer cell apoptosis [74]. Stigmasterol, one of the major phytosterols, downregulated lipid peroxidation and increased the levels of glutathione, superoxide dismutase, and catalase in the liver, and consequently decreased tumor volume, viable cell count, and increased mean survival time of Ehrlich ascites carcinoma tumor-bearing mice [75]. Squalene involved in natural triterpene potentially inhibited cancer cell proliferation without disturbing the normal biochemical pathways [76].
Truffles comprised significant amounts of polysaccharides possessing anticancer activity. Most polysaccharides in fungi are β-glucan polymers having the main chain which contains β-(1 → 3) linkage with some β-(1 → 6) branches [77]. Polysaccharides and their derivates play important roles in various biological processes, including pathogenesis prevention, signal recognition, and cell–cell communication [78]. It has been reported from previous studies that these macromolecules have oncogenesis- and metastasis-preventing abilities, including immunopotentiation activity against tumors. The polysaccharides in truffles could stimulate lymphocyte and macrophage division and synthesis of cytokines such as interferons, interleukins, and immunoglobulins directed against cancer antigens [79,80,81]. According to Attia et al., polysaccharides extract from T. claveryi possessed anticancer effect on Ehrlich’s ascites carcinoma cells, arresting G2 phase and reducing the percentage of G0/G1 phase in the cell cycle [82]. Zhao et al. revealed that 52 polysaccharides isolated from T. aestivum, T. indicum, T. melanosporum, and T. sinense fermentation systems as well as from their fruiting bodies showed antitumor activities against A549, HCT-116, HepG2, HL-60, and SK-BR-3 cells lines. This study found that the fraction from the fermentation system, which contained β-d-glucan having triple helix conformation (high-chain stiffness), showed significantly higher cancer cell inhibition rate than those from fruiting bodies. Moreover, the heteropolysaccharides with having lower molecular weight exhibited higher antitumor activity than those with higher molecular weight [83].
The oleic acid content in truffles also contributed to the anticancer activities. Oleic acid could suppress the overexpression of HER2, an oncogene correlated to the invasion and metastasis of cancer cells. Moreover, they could increase the intracellular ROS production and caspase 3 activity and consequently induce the cancer cell apoptosis [84, 85].
Pacioni et al. found that the fruiting bodies of T. melanosporum contained the major endocannabinoid metabolic enzymes such as N-acylphosphatidylethanolamine-specific phospholipase D (NAPE-PLD), fatty acid amide hydrolase (FAAH), diacylglycerol lipase (DAGL) and monoacylglycerol lipase (MAGL), and anandamide which is a fatty acid neurotransmitter derived from arachidonic acid and showed anticancer activity [86]. According to Picardi et al. [87], anandamide inhibited the angiogenesis of highly invasive and metastatic breast cancer cells. Patsos et al. [88] revealed that PG-EAs, the cyclooxygenase 2 (COX-2) dependent metabolites of anandamide, induced apoptosis of colorectal cancer cells, while anandamide stimulated non-apoptotic cell death in COX-2 overexpressed colorectal cancer cells. In EGFR-overexpressed prostate cancer cells, anandamide decreased EGFR levels by interacting with cannabinoid CB1 receptor, and consequently, inhibited the cancer cell proliferation [89].
Antioxidant and anti-inflammatory activities
Truffles comprise of secondary antioxidant metabolites such as phenolics, flavonoids, tocopherol, carotenoids, and phytosterols. As a free radical scavenger, phenolics can be used as an efficient antioxidant due to their reducing abilities as hydrogen- or electron-donating agents [90]. According to Beara et al. [71], water and methanolic extracts of T. aestivum and T. magnatum possessed moderate antioxidant properties related to their phenolic contents of 12–18 and 13–19 mg GAE/g DW, respectively. According to Villares et al., the methanolic extract of T. aestivum, T. indicum, and T. melanosporum comprised phenolics and ergosteryl ester of 1.88 mg/g DW and 0.36 mg/g DW, 1.52 mg/g DW and 0.09 mg/g DW, and 1.20 mg/g DW and 1.10 mg/g DW, respectively. Consequently, the highest phenolics and ergosteryl ester levels of T. aestivum were closely correlated to its greatest antioxidant power amongst these three species [67].
Desert truffle species also comprised of diverse antioxidant secondary metabolites. T. nivea showed much higher total phenolic content than that of T. pinoyi, and possessed greater antioxidant potential than T. pinoyi [19, 46]. According to Hamza and Jdir, ascorbic acid content (10.63 mg/100 g), and the combined content of carotenoids (1.17 mg/100 g), and anthocyanins (29.1 mg/100 g) in T. nivea were responsible to their antioxidant potential [46]. Doğan and Aydın revealed that the high scavenging effect of T. boudieri would correlate with catechin, the predominant phenolics present in this truffle [91].
Apart from desert truffles, Chinese truffle species also possessed high antioxidant properties. Chen et al. [92] evaluated the great DPPH radical scavenging ability of purified water-soluble polysaccharides from T. huidongense. Yan et al. demonstrated that T. latisporum, T. subglobosum, and T. pseudohimalayense revealed antioxidant activities, in particular, T. latisporum and T. pseudohimalayense, which were more abundant in phenolic compounds, possessed much stronger antioxidant potential than T. subglobosum [48].
The antioxidant compounds including phenolics, terpenoids, and polysaccharides also possess anti-inflammatory properties [77, 93]. Oxidative stress is often correlated to inflammatory disorders, particularly diabetes mellitus (DM). It is due to the activity of hyperglycemia, the typical indicator of DM, causes over-production of free radical [94]. Zhang et al. evaluated the effects of aqueous extract from T. melanosporum on the streptozotocin induced hyperglycemic rat. Consequently, the rat treated with truffle extract showed the reduced levels of glucose like the rat treated with the standard antidiabetic drug, glibenclamide. The hypoglycemic effect of truffle extract was significantly correlated with Nrf2 and NF-kB pathways, and the regulation of enzymatic and non-enzymatic antioxidant including superoxide dismutase, catalase, vitamin E, and vitamin C [95]. In addition to hypoglycemic effect, Janakat and Nassar revealed the hepatoprotective effect of aqueous extract from T. claveryi on liver damage caused by oxidative stress [96]. Moreover, Beara et al. [71] demonstrated that T. magnatum extract inhibited the COX-1 and 12-LOX pathway products, 12(S)-hydroxy-(5Z,8E,10E)-heptadecatrienoic acid (12-HHT), thromboxane B2 (TXB2), and 12(S)-hydroxy-(5Z,8Z,10E,14Z)-eicosatetraenoic acid (12-HETE), which overexpressed in various inflammatory diseases.
Antimicrobial activity
The antimicrobial activity of desert truffles has been well reported for a few decades. Janakat et al. [97] demonstrated that the aqueous extract obtained from T. claveryi inhibited the growth of Staphylococcus aureus by 66.4%. According to Owaid et al., silver nanoparticles synthesized by Tirmania sp. revealed the antibacterial activity towards gram-negative and gram-positive bacteria, predominantly the infectious bacteria of the eyes, Pseudomonas aeruginosa [98]. In another study conducted by Casarica et al., the aqueous extract from T. claveryi possessed the antimicrobial properties against Escherichia coli, Staphylococcus epidermidis, and Staphylococcus aureus, and consequently, their usage was suggested to treat eye infections caused by those three bacterial species [99]. Another desert truffle, T. nivea, from Tunisia arid zone was also exhibited by the remarkable inhibitory activity against three species of gram-positive and four species of gram-negative bacteria including Salmonella typhimurium, Escherichia coli, Pseudomonas aeruginosa, Enterococcus faecalis, Staphylococcus aureus, Staphylococcus epidermis, and Bacillus subtilis [46]. Dib-Bellahouel and Fortas found that the growth of Bacillus subtilis and Staphylococcus aureus was inhibited by ethyl acetate extract of T. pinoyi which contained pyrazines and its derivatives as the antimicrobial active compounds [100]. Stojković et al. [45] revealed that the aqueous and methanolic extract from T. pinoyi efficiently inhibited the growth of Staphylococcus aureus in chicken soup kept in the refrigerator and at room temperature. Table 2 summarizes more details about studies investigating the antimicrobial activity of truffles.
Aphrodisiac activity
Traditionally, truffles have been used as an effective sexual enhancer due to their constituent of androstenol as a steroidal pheromone [101, 102]. While truffle hunting, animals might recognize the odor of this chemical marker. Androstenol was also found in the underarm perspiration of men and urine of women and increased sexual attractiveness higher [11, 102]. The alcoholic extract of T. boudieri increased the levels of luteinizing hormone and testosterone significantly in rats and consequently possessed the aphrodisiac activity owing to its androgen enhancing properties [103]. However, Al-Damegh demonstrated that the androgenic property of truffles was more related to psychological effect because the flavonoids in truffles were present in the form of glycosides, which act as antagonists for male sex hormones [102].
Other potential applications
The bioactive compounds in truffles potentiate the use of truffles in other medicinal usages such as anti-depressants, cholesterol reducer, and immunostimulant. For example, the blood cholesterol-reducing effect of linoleic acid has been widely reported [104, 105]. In the meanwhile, truffles comprise abundant amino acids involving l-tyrosine which is a precursor of neurotransmitters catecholamines deeply involved in neural circuits [44, 106, 107]. Pattanayak et al. studied the cytotoxic effect of water-soluble heteroglycan obtained from T. rufum on human blood lymphocytes. Consequently, they discovered the potential of this polysaccharide as an immunostimulant, in terms of its ameliorative activities by maintaining the redox balance [108].
Various factors affecting productivity and quality of truffles
Like most edible fungi, the growth pattern, nutritional quality, and biological activities of truffles could be changed by various factors such as climate, soil, or the presence of contaminating microorganisms [8]. For example, dehydrative low-temperature stress induces accumulation of dehydrins in truffles proposed to play a protective role [109] and upregulation of stress-responsive genes including heat shock proteins and cell wall remodeling genes in T. melanosporum [110]. High temperatures and hydric stress in summer, together with decreased summer precipitation, could also lead to the genetic or morphological change and reduction of subsequent winter harvest [111]. Leonardi et al. evaluated the effect of heat stress on eleven T. borchii strains from different geographical and ecological provenance. Ten from eleven tested strains showed the significant growth reduction accompanying by increased hyphal septation at 34 °C, while the best performances of mycelial growth and biomass production were at 22 °C. In terms of global climate change, a suitable selection of strain which is tolerant in temperature change could be one of the successful strategies for truffle cultivation [112].
Edaphic factors such as pH, moisture, and nutrient also influence the richness and composition of truffles [113]. Hall et al. [114] demonstrated that a soil pH of 8.0–8.5 was an appropriate condition for the T. melanosporum cultivation. Ge et al. revealed that soil pH is strongly correlated to the levels of minerals in truffles such as K+, Ca2+, and Mg2+, and host plants generally produced more T. lyonii fruiting bodies at soil pH of 6.6–7.3 [115].
Meanwhile, truffle productivity and quality could be improved by regulating preharvest factors. As truffles are hypogeous fungi, they interact with soil micro-fauna and microorganisms, which can enhance the truffle formation and their antagonistic, competitive, or synergistic activities [8]. Frey-Klett et al. demonstrated that co-inoculating, adding a small amount of helper bacterium to the ectomycorrhizal fungus, could lead to much easier and faster-growing than ectomycorrhizal fungi. Thus, this method can be one of the efficient methods for optimizing the mycorrhization techniques as well as minimizing the inoculation cost [116]. Tang’s research team has developed and improved the submerged fermentation process for enhancing not only truffle biomass but also the production of polysaccharides, VOCs, and sterols in truffles [117]. For example, they demonstrated that the addition of Mg2+ and K+ could enhance the production of extracellular polysaccharides during the submerged fermentation of T. sinense. According to Vahdatzadeh and Splivallo, genetic features deeply influenced the truffle aroma formation, and the human nose could distinguish the flavor among the strains. From this regard, strain selection could be effectively used for improving the truffle flavor [118].
Truffles are highly perishable and very sensitive to pathogen contamination and water loss [119]. During storage processes such as frozen or sterilization, the aroma quality and texture of truffles had been changed. Therefore, preservation methods play a critical role in the maintenance or improving their quality. Saltarelli et al. [55] recommended a storage temperature at 4 °C to preserve the biochemical and microbiological characteristics of fresh T. aestivum, T. magnatum, and T. melanosporum. Al-Ruqaie demonstrated that freezing after blanching in 4% boiling NaCl solution is the most effective preservation method for maintaining the quality of T. claveryi and T. nivea [120]. Rivera et al. recommended lyophilization as an adequate method to better preserve the aroma of T. melanosporum, while Palacios et al. suggested that rehydration after lyophilization recovered the aromatic profile changed through lyophilization [121, 122]. According to Tirillini et al. [123], the addition of 10% linoleic acid (w/w) significantly reduced total bacterial load and enhance the nutritional value of truffle sauce.
Irradiation is one of the ways affecting microbial populations, shelf life, chemical composition, and sensory characteristics of truffles. Adamo et al. [124] demonstrated that the irradiated truffles showed a microbial load of five orders of magnitude less than the non-irradiated. Reale et al. [125] revealed that irradiation at 1.5 kilo gray (kGy) was the most effective way for extending the shelf life of truffles as well as minimizing the alteration of sensorial features of fresh truffles. According to Culleré et al. [126], the ionizing radiation at 1.5 and 2.5 kGy increased the levels of practically all the primary odorant of T. melanosporum and T. aestivum, and therefore, these type of irradiation were considered as an effective way for maintaining the truffle aroma. Tejedor-Calvo et al. recommended that electron beam or gamma irradiation (both at 1.5 kGy and 2.5 kGy) followed by storage up to 21 days were the efficient methods to enhance truffles quality and shelf life. In the study, the irradiation exhibited no influence on the total carbohydrates, β-glucans, and overall sterols levels. Although this method significantly reduced protein and chitin levels, storage after the irradiation maintained the levels of all determined compounds unchanged except for the phenolics and ergosterol levels that increased after 21 days storage [127].
Storing fresh truffles under high carbon dioxide conditions is also associated with their quality. Mencarelli et al. [128] revealed that CO2 treatment on T. aestivum at 5 °C could keep well their fragrance and robust defense system. According to Hajjar et al., storing T. uncinatum in high CO2 and low O2 atmospheres played a decisive role in maintaining their antioxidative properties by inhibiting polyphenol metabolism, anaerobic pathways, and polyamine biosynthesis. The antioxidant enzymes in truffles were active when they are kept in the soil with lower O2 conditions; therefore, removing truffles from lower to higher O2 conditions could weaken their antioxidant defense system [119].
Conclusions
The unique flavor of truffles has made them the high demanded foodstuffs; however, they are not enough to be regarded only as high-priced foodstuffs because of their nutritional and medicinal potentials as well as no known side effects. Diverse truffle species are rich in nutrients such as carbohydrates, protein, fiber, minerals, fatty acids, and amino acids. Apart from their high value in the nutritional domain, their potential in therapeutic agents is ample as well. Recent studies have identified diverse bioactive compounds such as phenolics, flavonoids, and polysaccharides in truffles and revealed their potentials as anticancer, antioxidant, antimicrobial, anti-inflammatory, antidiabetic agents. Nevertheless, most studies have focused on in vitro studies and limited researches on the application of truffles in practical therapy has been investigated. Future studies need to optimize the extraction process to obtain a high yield of the bioactive compounds. Moreover, it is needed to focus on their bioassay against different diseases and pathogens to incorporate these biological values effectively into the truffles derived natural products as well as potential medicines. In this regard, more innovative methods are also expected to overcome the difficulty in preserving truffles to enhance their shelf life whilst maintaining their bioactive compounds and causing no harmful effects. These attempts would enable the practical usage of truffles in nutritional and medicinal fields.
Availability of data and materials
Not applicable.
References
El Enshasy H, Elsayed EA, Aziz R, Wadaan MA. Mushrooms and truffles: historical biofactories for complementary medicine in Africa and in the middle east. Evid Based Complement Altern Med. 2013;2013:1–10. https://doi.org/10.1155/2014/805841.
Læssøe T, Hansen K. Truffle trouble: what happened to the Tuberales? Mycol Res. 2007;111:1075–99. https://doi.org/10.1016/j.mycres.2007.08.004.
Bonito GM, Gryganskyi AP, Trappe JM, Vilgalys R. A global meta-analysis of Tuber ITS rDNA sequences: species diversity, host associations and long-distance dispersal. Mol Ecol. 2010;19:4994–5008. https://doi.org/10.1111/j.1365-294X.2010.04855.x.
Berch SM, Bonito G. Truffle diversity (Tuber, Tuberaceae) in British Columbia. Mycorrhiza. 2016;26:587–94. https://doi.org/10.1007/s00572-016-0695-2.
Kinoshita A, Sasaki H, Nara K. Phylogeny and diversity of Japanese truffles (Tuber spp.) inferred from sequences of four nuclear loci. Mycologia. 2011;103:779–94. https://doi.org/10.3852/10-138.
Bradai L, Bissati S, Chenchouni H, Amrani K. Effects of climate on the productivity of desert truffles beneath hyper-arid conditions. Int J Biometeorol. 2015;59:907–15. https://doi.org/10.1007/s00484-014-0891-8.
Wang S, Marcone MF. The biochemistry and biological properties of the world’s most expensive underground edible mushroom: truffles. Food Res Int. 2011;44:2567–81. https://doi.org/10.1016/j.foodres.2011.06.008.
Mello A, Murat C, Bonfante P. Truffles: much more than a prized and local fungal delicacy. FEMS Microbiol Lett. 2006;260:1–8. https://doi.org/10.1111/j.1574-6968.2006.00252.x.
Jeandroz S, Murat C, Wang Y, Bonfante P, Le Tacon F. Molecular phylogeny and historical biogeography of the genus Tuber, the “true truffles”. J Biogeogr. 2008;35:815–29. https://doi.org/10.1111/j.1365-2699.2007.01851.x.
Chen PY, Montanini B, Liao WW, Morselli M, Jaroszewicz A, Lopez D, et al. A comprehensive resource of genomic, epigenomic and transcriptomic sequencing data for the black truffle Tuber melanosporum. Gigascience. 2014;3:25. https://doi.org/10.1186/2047-217X-3-25.
Patel S. Food, health and agricultural importance of truffle. Curr Trends Biotechnol Pharm. 2012;6:15–27.
Chen J, Li JM, Tang YJ, Xing YM, Qiao P, Li Y, et al. Chinese black truffle-associated bacterial communities of Tuber indicum from different geographical regions with nitrogen fixing bioactivity. Front Microbiol. 2019;10:1–14. https://doi.org/10.3389/fmicb.2019.02515.
Selosse MA, Schneider-Maunoury L, Taschen E, Rousset F, Richard F. Black truffle, a hermaphrodite with forced unisexual behaviour. Trends Microbiol. 2017;25:784–7. https://doi.org/10.1016/j.tim.2017.05.010.
Paolocci F, Rubini A, Riccioni C, Arcioni S. Reevaluation of the life cycle of Tuber magnatum. Appl Environ Microbiol. 2006;72:2390–3. https://doi.org/10.1128/AEM.72.4.2390-2393.2006.
Splivallo R, Ottonello S, Mello A, Karlovsky P. Truffle volatiles: from chemical ecology to aroma biosynthesis. New Phytol. 2011;189:688–99. https://doi.org/10.1111/j.1469-8137.2010.03523.x.
Le Tacon F, Marçais B, Courvoisier M, Murat C, Montpied P, Becker M. Climatic variations explain annual fluctuations in French Périgord black truffle wholesale markets but do not explain the decrease in black truffle production over the last 48 years. Mycorrhiza. 2014;24:115–25. https://doi.org/10.1007/s00572-014-0568-5.
Díaz P, Señoráns FJ, Reglero G, Ibañez E. Truffle aroma analysis by headspace solid phase microextraction. J Agric Food Chem. 2002;50:6468–72. https://doi.org/10.1021/jf025609t.
Technavio. Global truffles market 2019–2023. 2019.
Al-Laith AAA. Antioxidant components and antioxidant/antiradical activities of desert truffle (Tirmania nivea) from various Middle Eastern origins. J Food Compos Anal. 2010;23:15–22. https://doi.org/10.1016/j.jfca.2009.07.005.
Schmidberger PC, Schieberle P. Characterization of the key aroma compounds in white alba truffle (Tuber magnatum pico) and burgundy truffle (Tuber uncinatum) by means of the sensomics approach. J Agric Food Chem. 2017;65:9287–96. https://doi.org/10.1021/acs.jafc.7b04073.
Xiao DR, Liu RS, He L, Li HM, Tang YL, Liang XH, Chen T, Tang YJ. Aroma improvement by repeated freeze-thaw treatment during Tuber melanosporum fermentation. Sci Rep. 2015;5:17120. https://doi.org/10.1038/srep17120.
Wernig F, Buegger F, Pritsch K, Splivallo R. Composition and authentication of commercial and home-made white truffle-flavored oils. Food Control. 2018;87:9–16. https://doi.org/10.1016/j.foodcont.2017.11.045.
Patel S, Rauf A, Khan H, Khalid S, Mubarak MS. Potential health benefits of natural products derived from truffles: a review. Trends Food Sci Technol. 2017;70:1–8. https://doi.org/10.1016/j.tifs.2017.09.009.
Feng T, Shui M, Song S, Zhuang H, Sun M, Yao L. Characterization of the key aroma compounds in three truffle varieties from China by flavoromics approach. Molecules. 2019;24:3305. https://doi.org/10.3390/molecules24183305.
Kamle M, Bar E, Lewinsohn D, Shavit E, Roth-Bejerano N, Kagan-Zur V, et al. Characterization of morphology, volatile profiles, and molecular markers in edible desert truffles from the negev desert. J Agric Food Chem. 2016;65:2977–83. https://doi.org/10.1021/acs.jafc.6b04063.
Du M, Huang S, Wang J. The volatiles from fermentation product of Tuber formosanum. Open J For. 2014;4:426–9. https://doi.org/10.4236/ojf.2014.44047.
Culleré L, Ferreira V, Venturini ME, Marco P, Blanco D. Chemical and sensory effects of the freezing process on the aroma profile of black truffles (Tuber melanosporum). Food Chem. 2013;136:518–25. https://doi.org/10.1016/j.foodchem.2012.08.030.
Torregiani E, Lorier S, Sagratini G, Maggi F, Vittori S, Caprioli G. Comparative analysis of the volatile profile of 20 commercial samples of truffles, truffle sauces, and truffle-flavored oils by using HS-SPME-GC-MS. Food Anal Methods Food Anal Methods. 2017;10:1857–69. https://doi.org/10.1007/s12161-016-0749-2.
Liu C, Li Y. Evaluation of the volatile profile of Tuber liyuanum by HS-SPME with GC–MS. Nat Prod Res. 2017;31:972–5. https://doi.org/10.1080/14786419.2016.1253082.
Brattoli M, de Gennaro G, de Pinto V, Loiotile AD, Lovascio S, Penza M. Odour detection methods: olfactometry and chemical sensors. Sensors. 2011;11:5290–322. https://doi.org/10.3390/s110505290.
Liu RS, Jin GH, Xiao DR, Li HM, Bai FW, Tang YJ. Screening of the key volatile organic compounds of Tuber melanosporum fermentation by aroma sensory evaluation combination with principle component analysis. Sci Rep. 2015;5:17954. https://doi.org/10.1038/srep17954.
Splivallo R, Ebeler SE. Sulfur volatiles of microbial origin are key contributors to human-sensed truffle aroma. Appl Microbiol Biotechnol. 2015;99:2583–92. https://doi.org/10.1007/s00253-014-6360-9.
Culleré L, Ferreira V, Venturini ME, Marco P, Blanco D. Potential aromatic compounds as markers to differentiate between Tuber melanosporum and Tuber indicum truffles. Food Chem. 2013;141:105–10. https://doi.org/10.1016/j.foodchem.2013.03.027.
Liu RS, Li DC, Li HM, Tang YJ. Evaluation of aroma active compounds in Tuber fruiting bodies by gas chromatography-olfactometry in combination with aroma reconstitution and omission test. Appl Microbiol Biotechnol. 2012;94:353–63. https://doi.org/10.1007/s00253-011-3837-7.
Shimokawa T, Kinoshita A, Kusumoto N, Nakano S, Nakamura N, Yamanaka T. Component features, odor-active volatiles, and acute oral toxicity of novel white-colored truffle Tuber japonicum native to Japan. Food Sci Nutr. 2020;8:410–8. https://doi.org/10.1002/fsn3.1325.
Vita F, Taiti C, Pompeiano A, Bazihizina N, Lucarotti V, Mancuso S, et al. Volatile organic compounds in truffle (Tuber magnatum Pico): comparison of samples from different regions of Italy and from different seasons. Sci Rep. 2015;5:12629. https://doi.org/10.1038/srep12629.
Engel W, Bahr W, Schieberle P. Solvent assisted flavour evaporation—a new and versatile technique for the careful and direct isolation of aroma compounds from complex food matrices. Eur Food Res Technol. 1999;209:237–41. https://doi.org/10.1007/s002170050486.
Mo X, Yan XU, Wenlai FAN. Characterization of aroma compounds in Chinese rice wine Qu by solvent-assisted flavor evaporation and headspace solid-phase microextraction. J Agric Food Chem. 2010;58:2462–9. https://doi.org/10.1021/jf903631w.
Li N, Zheng F-P, Chen H-T, Liu S-Y, Gu C, Song Z-Y, et al. Identification of volatile components in Chinese Sinkiang fermented camel milk using SAFE, SDE, and HS-SPME-GC/MS. Food Chem. 2011;129:1242–52. https://doi.org/10.1016/j.foodchem.2011.03.115.
Zhang N, Chen H, Sun B, Mao X, Zhang Y, Zhou Y. Comparative analysis of volatile composition in chinese truffles via GC × GC/HR-TOF/MS and electronic nose. Int J Mol Sci. 2016;17:412. https://doi.org/10.3390/ijms17040412.
Camin F, Larcher R, Nicolini G, Bontempo L, Bertoldi D, Perini M, et al. Isotopic and elemental data for tracing the origin of European olive oils. J Agric Food Chem. 2010;58:570–7. https://doi.org/10.1021/jf902814s.
Sciarrone D, Schepis A, Zoccali M, Donato P, Vita F, Creti D, et al. Multidimensional gas chromatography coupled to combustion-isotope ratio mass spectrometry/quadrupole MS with a low-bleed ionic liquid secondary column for the authentication of truffles and products containing truffle. Anal Chem. 2018;90:6610–7. https://doi.org/10.1021/acs.analchem.8b00386.
Owaid MN. Bioecology and uses of desert truffles (Pezizales) in the middle east. Walailak J Sci Technol. 2018;15:179–88.
Kıvrak İ. Analytical methods applied to assess chemical composition, nutritional value and in vitro bioactivities of Terfezia olbiensis and Terfezia claveryi from Turkey. Food Anal Methods. 2015;8:1279–93. https://doi.org/10.1007/s12161-014-0009-2.
Stojković D, Reis FS, Ferreira ICFR, Barros L, Glamočlija J, Ćirić A, et al. Tirmania pinoyi: chemical composition, in vitro antioxidant and antibacterial activities and in situ control of Staphylococcus aureus in chicken soup. Food Res Int. 2013;53:56–62. https://doi.org/10.1016/j.foodres.2013.03.046.
Hamza A, Jdir H, Zouari N. Nutritional, antioxidant and antibacterial properties of Tirmania nivea, a wild edible desert truffle from Tunisia arid zone. Med Aromat Plants. 2016;5:258. https://doi.org/10.4172/2167-0412.1000258.
Bouatia M, Touré HA, Cheikh A, Eljaoudi R, Rahali Y, Oulad Bouyahya Idrissi M, et al. Analysis of nutrient and antinutrient content of the truffle (Tirmania pinoyi) from Morocco. Int Food Res J. 2018;25:174–8.
Yan X, Wang Y, Sang X, Fan L. Nutritional value, chemical composition and antioxidant activity of three Tuber species from China. AMB Express. 2017;7:136. https://doi.org/10.1186/s13568-017-0431-0.
Angelini P, Tirillini B, Properzi A, Rol C, Venanzoni R. Identification and bioactivity of the growth inhibitors in Tuber spp. methanolic extracts. Plant Biosyst. 2015;149:1000–9. https://doi.org/10.1080/11263504.2014.983575.
Ribeiro B, Pinho P, Andrade PB, Baptista P, Valentão P. Fatty acid composition of wild edible mushrooms species: a comparative study. Microchem J. 2009;93:29–35. https://doi.org/10.1016/j.microc.2009.04.005.
Pacheco YM, López S, Bermúdez B, Abia R, Villar J, Muriana FJG. A meal rich in oleic acid beneficially modulates postprandial sICAM-1 and sVCAM-1 in normotensive and hypertensive hypertriglyceridemic subjects. J Nutr Biochem. 2008;19:200–5. https://doi.org/10.1016/j.jnutbio.2007.03.002.
Puiggrós C, Chacón P, Armadans LI, Clapés J, Planas M. Effects of oleic-rich and omega-3-rich diets on serum lipid pattern and lipid oxidation in mildly hypercholesterolemic patients. Clin Nutr. 2002;21:79–87. https://doi.org/10.1054/clnu.2001.0511.
Gençcelep H, Uzun Y, Tunçtürk Y, Demirel K. Determination of mineral contents of wild-grown edible mushrooms. Food Chem. 2009;113:1033–6. https://doi.org/10.1016/j.foodchem.2008.08.058.
Vahdani M, Rastegar S, Rahimzadeh M, Ahmadi M, Karmostaji A. Physicochemical characteristics, phenolic profile, mineral and carbohydrate contents of two truffle species. J Agric Sci Technol. 2017;19:1091–101.
Saltarelli R, Ceccaroli P, Cesari P, Barbieri E, Stocchi V. Effect of storage on biochemical and microbiological parameters of edible truffle species. Food Chem. 2008;109:8–16. https://doi.org/10.1016/j.foodchem.2007.11.075.
Sawaya WN, Al-Sahlhat A, Al-Sogair A, Al-Mohammad M. Chemical composition and nutritive value of truffles of Saudi Arabia. J Food Sci. 1985;50:450–3. https://doi.org/10.1111/j.1365-2621.1985.tb13425.x.
Rejon-Palomares A, Garcia-Garrido JM, Ocampo JA, Garcia-Romera I. Presence of xyloglucan-hydrolyzing glucanases (xylogucanases) in arbuscular mycorrhizal symbiosis. Symbiosis. 1996;21:249–61.
Nadim M, Deshaware S, Saidi N, Abd-Elhakeem MA, Ojamo H, Shamekh S. Extracellular enzymatic activity of Tuber maculatum and Tuber aestivum mycelia. Adv Microbiol. 2015;5:523–30. https://doi.org/10.4236/aim.2015.57054.
Bedade D, Deska J, Bankar S, Bejar S, Singhal R, Shamekh S. Fermentative production of extracellular amylase from novel amylase producer, Tuber maculatum mycelium, and its characterization. Prep Biochem Biotechnol. 2018;48:549–55. https://doi.org/10.1080/10826068.2018.1476876.
Panche AN, Diwan AD, Chandra SR. Flavonoids: an overview. J Nutr Sci. 2016;5:1–15. https://doi.org/10.1017/jns.2016.41.
Gil-Ramírez A, Pavo-Caballero C, Baeza E, Baenas N, Garcia-Viguera C, Marín FR, Soler-Rivas C. Mushrooms do not contain flavonoids. J Funct Foods. 2016;25:1–13. https://doi.org/10.1016/j.jff.2016.05.005.
Hamza A, Zouari N, Zouari S, Jdir H, Zaidi S, Gtari M, et al. Nutraceutical potential, antioxidant and antibacterial activities of Terfezia boudieri Chatin, a wild edible desert truffle from Tunisia arid zone. Arab J Chem. 2016;9:383–9. https://doi.org/10.1016/j.arabjc.2013.06.015.
Rekika D, Khanizadeh S, Deschênes M, Levasseur A, Charles MT, Tsao R, et al. Antioxidant capacity and phenolic content of selected strawberry genotypes. HortScience. 2005;40:1777–81. https://doi.org/10.21273/hortsci.40.6.1777.
Lu X, Wang J, Al-Qadiri HM, Ross CF, Powers JR, Tang J, et al. Determination of total phenolic content and antioxidant capacity of onion (Allium cepa) and shallot (Allium oschaninii) using infrared spectroscopy. Food Chem. 2011;129:637–44. https://doi.org/10.1016/j.foodchem.2011.04.105.
Usenik V, Fabčič J, Štampar F. Sugars, organic acids, phenolic composition and antioxidant activity of sweet cherry (Prunus avium L.). Food Chem. 2008;107:185–92. https://doi.org/10.1016/j.foodchem.2007.08.004.
Tripathi NN, Singh P, Vishwakarma P. Biodiversity of macrofungi with special reference to edible forms: a review. J Indian Bot Soc. 2017;96:144–87.
Villares A, García-Lafuente A, Guillamón E, Ramos Á. Identification and quantification of ergosterol and phenolic compounds occurring in Tuber spp. truffles. J Food Compos Anal. 2012;26:177–82. https://doi.org/10.1016/j.jfca.2011.12.003.
Kuo CF, Hsieh CH, Lin WY. Proteomic response of Lab-activated RAW 264.7 macrophages to the anti-inflammatory property of fungal ergosterol. Food Chem. 2011;126:207–12. https://doi.org/10.1016/j.foodchem.2010.10.101.
Hu SH, Liang ZC, Chia YC, Lien JL, Chen KS, Lee MY, Wang JC. Antihyperlipidemic and antioxidant effects of extracts from Pleurotus citrinopileatus. J Agric Food Chem. 2006;54:2103–10. https://doi.org/10.1021/jf052890d.
Shao S, Hernandez M, Kramer JKG, Rinker DL, Tsao R. Ergosterol profiles, fatty acid composition, and antioxidant activities of button mushrooms as affected by tissue part and developmental stage. J Agric Food Chem. 2010;58:11616–25. https://doi.org/10.1021/jf102285b.
Beara IN, Lesjak MM, Četojević-Simin DD, Marjanović ŽS, Ristić JD, Mrkonjić ZO, et al. Phenolic profile, antioxidant, anti-inflammatory and cytotoxic activities of black (Tuber aestivum Vittad.) and white (Tuber magnatum Pico) truffles. Food Chem. 2014;165:460–6. https://doi.org/10.1016/j.foodchem.2014.05.116.
Khadri H, Aldebasi YH, Riazunnisa K. Truffle mediated (Terfezia claveryi) synthesis of silver nanoparticles and its potential cytotoxicity in human breast cancer cells (MCF-7). Afr J Biotechnol. 2017;16:1278–84. https://doi.org/10.5897/AJB2017.16031.
Dahham SS, Al-Rawi SS, Ibrahim AH, Majid ASA, Majid AMSA. Antioxidant, anticancer, apoptosis properties and chemical composition of black truffle Terfezia claveryi. Saudi J Biol Sci. 2016;25:1524–34. https://doi.org/10.1016/j.sjbs.2016.01.031.
Woyengo TA, Ramprasath VR, Jones PJH. Anticancer effects of phytosterols. Eur J Clin Nutr. 2009;63:813–20. https://doi.org/10.1038/ejcn.2009.29.
Ghosh T, Maity TK, Singh J. Evaluation of antitumor activity of stigmasterol, a constituent isolated from Bacopa monnieri Linn aerial parts against Ehrlich Ascites Carcinoma in mice. Orient Pharm Exp Med. 2011;11:41–9. https://doi.org/10.1007/s13596-011-0001-y.
Reddy LH, Couvreur P. Squalene: a natural triterpene for use in disease management and therapy. Adv Drug Deliv Rev. 2009;61:1412–26. https://doi.org/10.1016/j.addr.2009.09.005.
Friedman M. Mushroom polysaccharides: chemistry and antiobesity, antidiabetes, anticancer, and antibiotic properties in cells, rodents, and humans. Foods. 2016;5:80. https://doi.org/10.3390/foods5040080.
Zong A, Cao H, Wang F. Anticancer polysaccharides from natural resources: a review of recent research. Carbohydr Polym. 2012;90:1395–410. https://doi.org/10.1016/j.carbpol.2012.07.026.
Li LF, Liu H-B, Zhang QW, Li ZP, Wong TL, Fung HY, et al. Comprehensive comparison of polysaccharides from Ganoderma lucidum and G. sinense: chemical, antitumor, immunomodulating and gut-microbiota modulatory properties. Sci Rep. 2018;8:6172. https://doi.org/10.1038/s41598-018-22885-7.
Moradali MF, Mostafavi H, Ghods S, Hedjaroude GA. Immunomodulating and anticancer agents in the realm of macromycetes fungi (macrofungi). Int Immunopharmacol. 2007;7:701–24. https://doi.org/10.1016/j.intimp.2007.01.008.
Wasser S. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl Microbiol Biotechnol. 2003;60:258–74. https://doi.org/10.1007/s00253-002-1076-7.
Attia WY, El-Naggar RE, Bawadekji A, Al Ali M. Evaluation of some in vitro anti-carcinogenic activities of polysaccharides extracted from ascomata of the desert truffle Terfezia claveryi chatin. J Appl Environ Biol Sci. 2018;8:152–9.
Zhao W, Wang XH, Li HM, Wang SH, Chen T, Yuan ZP, et al. Isolation and characterization of polysaccharides with the antitumor activity from Tuber fruiting bodies and fermentation system. Appl Microbiol Biotechnol. 2014;98:1991–2002. https://doi.org/10.1007/s00253-013-5379-7.
Carrillo C, del Cavia M, Alonso-Torre SR. Antitumor effect of oleic acid; mechanisms of action. A review. Nutr Hosp. 2012;27:1860–5. https://doi.org/10.3305/nh.2012.27.6.6010.
Menendez JA, Papadimitropoulou A, Vellon L, Lupu R. A genomic explanation connecting “Mediterranean diet”, olive oil and cancer: oleic acid, the main monounsaturated Fatty acid of olive oil, induces formation of inhibitory “PEA3 transcription factor-PEA3 DNA binding site” complexes at the Her-2/neu (erbB-2). Eur J Cancer. 2006;42:2425–32. https://doi.org/10.1016/j.ejca.2005.10.016.
Pacioni G, Rapino C, Zarivi O, Falconi A, Leonardi M, Battista N, et al. Truffles contain endocannabinoid metabolic enzymes and anandamide. Phytochemistry. 2015;110:104–10. https://doi.org/10.1016/j.phytochem.2014.11.012.
Picardi P, Ciaglia E, Proto M, Pisanti S. Anandamide inhibits breast tumor-induced angiogenesis. Transl Med UniSa. 2014;10:8–12.
Patsos HA, Hicks DJ, Dobson RRH, Greenhough A, Woodman N, Lane JD, et al. The endogenous cannabinoid, anandamide, induces cell death in colorectal carcinoma cells: a possible role for cyclooxygenase 2. Gut. 2005;54:1741–50. https://doi.org/10.1136/gut.2005.073403.
Mimeault M, Pommery N, Wattez N, Bailly C, Hénichart JP. Anti-proliferative and apoptotic effects of anandamide in human prostatic cancer cell lines: implication of epidermal growth factor receptor down-regulation and ceramide production. Prostate. 2003;56:1–12. https://doi.org/10.1002/pros.10190.
Rice-Evans C. Antioxidant properties of phenolic compounas. Trends Plant Sci. 1997;2:152–9.
Doğan HH, Aydın S. Determination of antimicrobial effect, antioxidant activity and phenolic contents of desert truffle in Turkey. Afr J Tradit Complement Altern Med. 2013;10:52–8. https://doi.org/10.4314/ajtcam.v10i4.9.
Chen G, Zhang S, Ran C, Wang L, Kan J. Extraction, characterization and antioxidant activity of water-soluble polysaccharides from Tuber huidongense. Int J Biol Macromol. 2016;91:431–42. https://doi.org/10.1016/j.ijbiomac.2016.05.108.
Elsayed EA, El Enshasy H, Wadaan MAM, Aziz R. Mushrooms: a potential natural source of anti-inflammatory compounds for medical applications. Mediat Inflamm. 2014;2014:805841. https://doi.org/10.1155/2014/805841.
Muriach M, Flores-Bellver M, Romero FJ, Barcia JM. Diabetes and the brain: oxidative stress, inflammation, and autophagy. Oxid Med Cell Longev. 2014;2014:120158. https://doi.org/10.1155/2014/102158.
Zhang T, Jayachandran M, Ganesan K, Xu B. Black truffle aqueous extract attenuates oxidative stress and inflammation in STZ-induced Hyperglycemic rats via Nrf2 and NF-κB pathways. Front Pharmacol. 2018;9:1257. https://doi.org/10.3389/fphar.2018.01257.
Janakat S, Nassar M. Hepatoprotective activity of desert truffle (Terfezia claveryi) in comparison with the effect of Nigella sativa in the rat. Pak J Nutr. 2010;9:52–6. https://doi.org/10.3923/pjn.2010.52.56.
Janakat S, Al-Fakhiri S, Sallal A-K. A promising peptide antibiotic from Terfezia claveryi aqueous extract against Staphylococcus aureus in vitro. Phyther Res. 2004;18:810–3. https://doi.org/10.1002/ptr.1563.
Owaid MN, Muslim RF, Hamad HA. Mycosynthesis of silver nanoparticles using Terminia sp. Desert truffle, Pezizaceae, and their antibacterial activity. Jordan J Biol Sci. 2018;11:401–5.
Casarica A, Moscovici M, Daas M, Nicu I, Panteli M, Rasit I. A purified extract from brown truffles of the species Terfezia claveryi chatin and its antimicrobial activity. Farmacia. 2016;64:298–301.
Dib-Bellahouel S, Fortas Z. Antibacterial activity of various fractions of ethyl acetate extract from the desert truffle, Tirmania pinoyi, preliminarily analyzed by gas chromatography–mass spectrometry (GC–MS). Afr J Biotechnol. 2011;10:9694–9. https://doi.org/10.5897/AJB10.2687.
Akyüz M, Kirbaǧ S, Kurşat M. Ecological aspects of the arid and semi-arid truffle in Turkey: evaluation of soil characteristics, morphology, distribution, and mycorrhizal relationships. Turk J Bot. 2012;36:386–91. https://doi.org/10.3906/bot-1103-4.
Al-Damegh MA. Tirmania (Zubaidi) and Terfezia (Khallasi) fungi preparation method modulates body and testicular weights and blood and testicular testosterone concentration in albino rats. J Am Sci. 2014;10:60–6.
Khojasteh SM, Amiri L, Sheikhzadeh F. Effect of the alcoholic extract of terfezia boudieri on reproductive hormones in male rats. Int J Pharm Biol Sci. 2013;3:517–22.
Ramsden CE, Zamora D, Leelarthaepin B, Majchrzak-Hong SF, Faurot KR, Suchindran CM, et al. Use of dietary linoleic acid for secondary prevention of coronary heart disease and death: evaluation of recovered data from the Sydney Diet Heart Study and updated meta-analysis. BMJ. 2013;346:1–18. https://doi.org/10.1136/bmj.e8707.
Horrobin DF, Huang YS. The role of linoleic acid and its metabolites in the lowering of plasma cholesterol and the prevention of cardiovascular disease. Int J Cardiol. 1987;17:241–55.
Arnsten AFT. Catecholamine and second messenger influences on prefrontal cortical networks of “representational knowledge”: a rational bridge between genetics and the symptoms of mental illness. Cereb Cortex. 2007;17:6–15. https://doi.org/10.1093/cercor/bhm033.
Young SN. l-tyrosine to alleviate the effects of stress? J Psychiatry Neurosci. 2007;32:224.
Pattanayak M, Samanta S, Maity P, Manna DK, Sen IK, Nandi AK, et al. Polysaccharide of an edible truffle Tuber rufum: structural studies and effects on human lymphocytes. Int J Biol Macromol. 2017;95:1037–48. https://doi.org/10.1016/j.ijbiomac.2016.10.092.
Abba S, Ghignone S, Bonfante P. A dehydration-inducible gene in the truffle Tuber borchii identifies a novel group of dehydrins. BMC Genomics. 2006;7:39. https://doi.org/10.1186/1471-2164-7-39.
Zampieri E, Balestrini R, Kohler A, Abbà S, Martin F, Bonfante P. The Perigord black truffle responds to cold temperature with an extensive reprogramming of its transcriptional activity. Fungal Genet Biol. 2011;48:585–91. https://doi.org/10.1016/j.fgb.2010.09.007.
Thomas P, Büntgen U. A risk assessment of Europe’s black truffle sector under predicted climate change. Sci Total Environ. 2019;655:27–34. https://doi.org/10.1016/j.scitotenv.2018.11.252.
Leonardi P, Iotti M, Donati Zeppa S, Lancellotti E, Amicucci A, Zambonelli A. Morphological and functional changes in mycelium and mycorrhizas of Tuber borchii due to heat stress. Fungal Ecol. 2017;29:20–9. https://doi.org/10.1016/j.funeco.2017.05.003.
Pickles BJ, Gorzelak MA, Green DS, Egger KN, Massicotte HB. Host and habitat filtering in seedling root-associated fungal communities: taxonomic and functional diversity are altered in ‘novel’ soils. Mycorrhiza. 2015;25:517–31. https://doi.org/10.1007/s00572-015-0630-y.
Hall I, Brown G, Zambonelli A. Taming the truffle: the history, lore, and science of the ultimate mushroom. JSTOR. 2008;59:69.
Ge Z-W, Brenneman T, Bonito G, Smith ME. Soil pH and mineral nutrients strongly influence truffles and other ectomycorrhizal fungi associated with commercial pecans (Carya illinoinensis). Plant Soil. 2017;418:493–505. https://doi.org/10.1007/s11104-017-3312-z.
Frey-Klett P, Churin JL, Pierrat JC, Garbaye J. Dose effect in the dual inoculation of an ectomycorrhizal fungus and a mycorrhiza helper bacterium in two forest nurseries. Soil Biol Biochem. 1999;31:1555–62. https://doi.org/10.1016/S0038-0717(99)00079-6.
Tang YJ, Liu R-S, Li H-M. Current progress on truffle submerged fermentation: a promising alternative to its fruiting bodies. Appl Microbiol Biotechnol. 2015;99:2041–53. https://doi.org/10.1007/s00253-015-6379-6.
Vahdatzadeh M, Splivallo R. Improving truffle mycelium flavour through strain selection targeting volatiles of the Ehrlich pathway. Sci Rep. 2018;8:9304. https://doi.org/10.1038/s41598-018-27620-w.
Hajjar SE, Massantini R, Botondi R, Kefalas P, Mencarelli F. Influence of high carbon dioxide and low oxygen on the postharvest physiology of fresh truffles. Postharvest Biol Technol. 2010;58:36–41. https://doi.org/10.1016/j.postharvbio.2010.04.008.
Al-Ruqaie IM. Effect of different treatment processes and preservation methods on the quality of truffles: I. Conventional methods (drying/freezing). J Food Process Preserv. 2006;30:335–51. https://doi.org/10.1111/j.1745-4549.2006.00069.x.
Rivera C, Venturini ME, Marco P, Oria R, Blanco D. Effects of electron-beam and gamma irradiation treatments on the microbial populations, respiratory activity and sensory characteristics of Tuber melanosporum truffles packaged under modified atmospheres. Food Microbiol. 2011;28:1252–60. https://doi.org/10.1016/j.fm.2011.05.002.
Palacios I, Guillamón E, García-Lafuente A, Villares A. Effects of freeze-drying treatment on the aromatic profile of Tuber spp. truffles. J Food Process Preserv. 2014;38:768–73. https://doi.org/10.1111/jfpp.12028.
Tirillini B, Maggi F, Venanzoni R, Angelini P, Ortega-Toro R. Enhanced duration of truffle sauce preservation due to addition of linoleic acid. J Food Qual. 2019;2019:8421898. https://doi.org/10.1155/2019/8421898.
Adamo M, Capitani D, Mannina L, Cristinzio M, Ragni P, Tata A, et al. Truffles decontamination treatment by ionizing radiation. Radiat Phys Chem. 2004;71:165–8. https://doi.org/10.1016/j.radphyschem.2004.03.073.
Reale A, Sorrentino E, Iacumin L, Tremonte P, Manzano M, Maiuro L, et al. Irradiation treatments to improve the shelf life of fresh black truffles (Truffles Preservation by Gamma-Rays). J Food Sci. 2009;74:196–200. https://doi.org/10.1111/j.1750-3841.2009.01142.x.
Culleré L, Ferreira V, Venturini ME, Marco P, Blanco D. Evaluation of gamma and electron-beam irradiation on the aromatic profile of black truffle (Tuber melanosporum) and summer truffle (Tuber aestivum). Innov Food Sci Emerg Technol. 2012;13:151–7. https://doi.org/10.1016/j.ifset.2011.09.003.
Tejedor-Calvo E, Morales D, Maro P, Venturini ME, Blanco D, Soler-Rivas C. Effects of combining electron-beam or gamma irradiation treatments with further storage under modified atmospheres on the bioactive compounds of Tuber melanosporum truffles. Postharvest Biol Technol. 2019;155:149–55. https://doi.org/10.1016/j.postharvbio.2019.05.022.
Mencarelli F, Massantini R, Botondi R. Physiological and textural response of truffles during low-temperature storage. J Hortic Sci. 1997;72:407–14. https://doi.org/10.1080/14620316.1997.11515528.
Acknowledgements
Not applicable.
Funding
This research received no external funding.
Author information
Authors and Affiliations
Contributions
Conceptualization and design of the draft, HL; writing the manuscript, HL and KN; design of the figures, HL, KN and ZZ; collection and assembling the data, KN; data analysis and interpretation; HL, ZZ, reviewing the manuscript; ZZ, providing the critical revision and supervision; MQF. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Lee, H., Nam, K., Zahra, Z. et al. Potentials of truffles in nutritional and medicinal applications: a review. Fungal Biol Biotechnol 7, 9 (2020). https://doi.org/10.1186/s40694-020-00097-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40694-020-00097-x